

Abstract
Several sublineages of omicron have emerged with additional mutations that may afford further antibody evasion. Here, we characterise the sensitivity of emerging omicron sublineages BA.2.75.2, BA.4.6, and BA.2.10.4 to antibody-mediated neutralisation, and identify extensive escape by BA.2.75.2. BA.2.75.2 was resistant to neutralisation by Evusheld (tixagevimab + cilgavimab), but remained sensitive to bebtelovimab. In recent serum samples from blood donors in Stockholm, Sweden, BA.2.75.2 was neutralised, on average, five-fold less potently than BA.5, representing the most neutralisation resistant variant evaluated to date. These data raise concerns that BA.2.75.2 may effectively evade humoral immunity in the population.
Main text
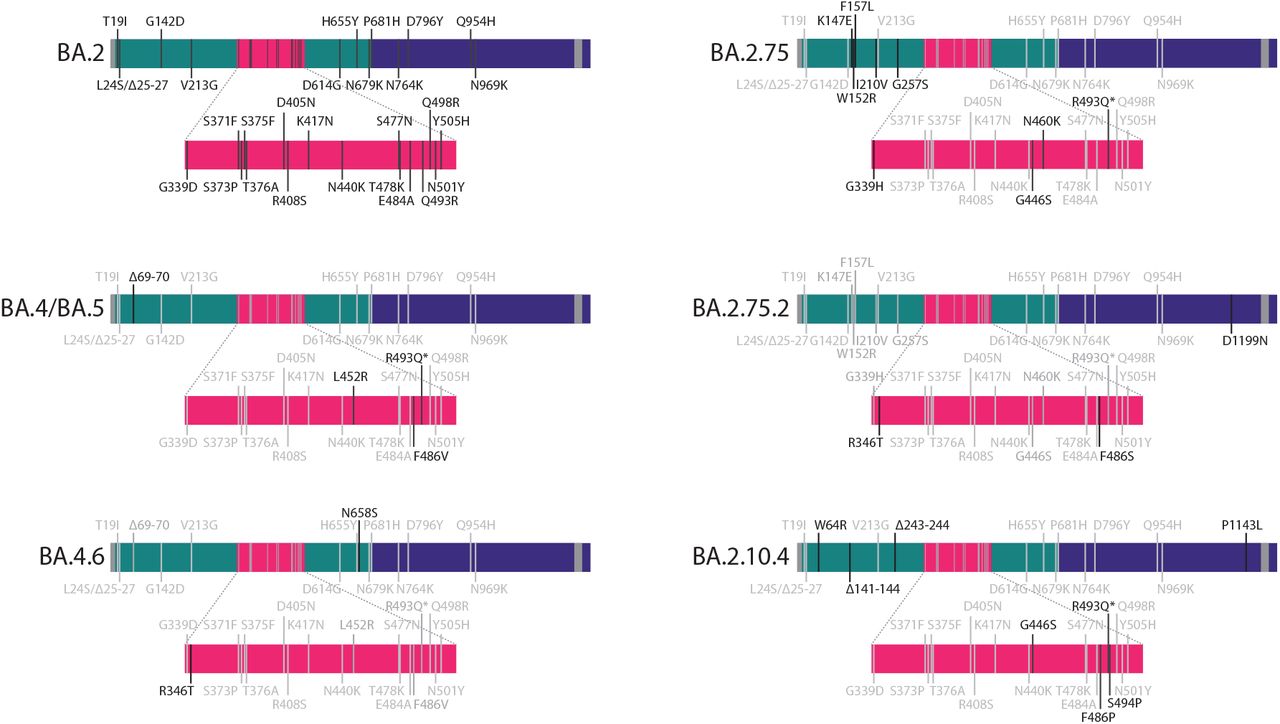
SARS-CoV-2 omicron sublineage BA.2.75 expanded rapidly in some parts of the world, but has so-far not outcompeted BA.5 globally. Despite similar geometric mean neutralising titers (GMT) to BA.5, BA.2.75 remained sensitive to classes of antibodies that BA.5 had escaped1,2, suggesting significant scope for further antibody evasion3. These included potent cross-neutralising epitope group B and D1 antibodies1. The emergence and rapid growth of a sublineage of BA.2.75 carrying additional mutations R346T, F486S, and D1199N (BA.2.75.2) is therefore suggestive of more extensive escape from neutralising antibodies.
Mutations to other residues at these positions have been previously characterised: F486V mediates significant escape in BA.54; R346K contributed to antibody escape in the pre-omicron Mu variant5, and occurred in some early omicron BA.1 isolates6, and the widespread BA.1.1 lineage; and R346S occurred in a spike evolved in vitro to escape polyclonal antibody responses (PMS20)7. Spike residue 346 has been specifically highlighted for its capacity for additional escape in omicron8, and numerous sublineages of omicron have been detected carrying convergent mutations there. BA.4.6, carrying R346T and N658S, is currently the dominant 346T-carrying lineage, and has been detected across a wide geographic distribution. Similarly, several lineages are emerging carrying mutations at 486, including BA.2.10.4 harbouring F486P.
Here we report the sensitivity of emerging omicron sublineages BA.2.75.2, BA.4.6, and BA.2.10.4 to neutralisation by a panel of clinically relevant and pre-clinical monoclonal antibodies, as well as by serum (N=18) from blood recently donated in Stockholm, Sweden during week 35, 2022 (29 Aug - 4 Sept 2022).
BA.2.75.2 and BA.4.6 both show complete escape from cilgavimab and the Evusheld combination, while BA.2.10.4 retains some sensitivity to cilgavimab (Fig 1A). S309 (sotrovimab) exhibits similarly low potency against BA.5, BA.2.75.2, and BA.2.10.4, with some further reduction against BA.4.6. Bebtelovimab still potently neutralises all variants tested.

Sensitivity of SARS-CoV-2 omicron sublineages relative to B.1 (D614G) to neutralisation by (A) monoclonal antibodies, and (B) randomly sampled sera from blood recently donated in Stockholm, Sweden between 29 Aug 2022 - 4 Sept 2022 (N=18). Sera with neutralisation <50% at the lowest dilution tested (20) are plotted as 20 (dotted line). ID50, 50% inhibitory dilution; IC50, 50% inhibitory concentration.
Against recently sampled serum, BA.4.6 and BA.2.10.4 (GMTs of 311 and 356 respectively) were moderately more resistant to neutralisation than BA.5 (GMT = 453). However, BA.2.75.2 exhibited substantial escape from neutralisation. Neutralising GMT against BA.2.75.2 was 85, more than 5-times lower than against BA.5 (Fig 1B), representing the most resistant variant characterised to date.
Taken together, these data identify profound antibody escape by the emerging omicron sublineage BA.2.75.2, suggesting that it effectively evades current humoral immunity in the population.
Methods
Cell culture
HEK293T cells (ATCC CRL-3216) and HEK293T-ACE2 cells (stably expressing human ACE2) were cultured in Dulbecco’s Modified Eagle Medium (high glucose, with sodium pyruvate) supplemented with 10% fetal bovine serum, 100 units/ml Penicillin, and 100 μg/ml Streptomycin. Cultures were maintained in a humidified 37°C incubator (5% CO2).
Pseudovirus Neutralisation Assay
Pseudovirus neutralisation assay was performed as previously4. Briefly, spike-pseudotyped lentivirus particles were generated by co-transfection of HEK293T cells with a relevant spike plasmid, an HIV gag-pol packaging plasmid (Addgene #8455), and a lentiviral transfer plasmid encoding firefly luciferase (Addgene #170674) using polyethylenimine. Spike variants were generated by multi-site directed mutagenesis of BA.2, BA.4, or BA.2.75 expression plasmids5, and all mutated plasmids were subsequently confirmed by Sanger sequencing.
Neutralisation was assessed in HEK293T-ACE2 cells. Pseudoviruses titrated to produce approximately 100,000 RLU were incubated with serial 3-fold dilutions of serum or monoclonal antibody for 60 minutes at 37°C in a black-walled 96-well plate. 10,000 HEK293T-ACE2 cells were then added to each well, and plates were incubated at 37°C for 44-48 hours. Luminescence was measured using Bright-Glo (Promega) on a GloMax Navigator Luminometer (Promega). Neutralisation was calculated relative to the average of 8 control wells infected in the absence of antibody. Samples were run against all variants ‘head-to-head’ using the same dilutions.
Monoclonal antibodies
Cilgavimab and tixagevimab were evaluated as their clinical formulations. For the rest of the monoclonal antibodies evaluated, antibody sequences were extracted from deposited RCSB entries, synthesized as gene fragments, cloned into pTWIST transient expression vectors by Gibson assembly or restriction cloning, expressed and purified, all as previously described6.
Serum samples
Serum samples from anonymized blood donors from Stockholm, Sweden, were obtained from from week 35, 2022 (29 August 2022 - 4 September 2022). Sera were heat inactivated at 56°C for 60 minutes prior to use in neutralisation assays.
Ethical Statement
The blood donor samples were anonymized, and not subject to ethical approvals, as per advisory statement 2020–01807 from the Swedish Ethical Review Authority.
Statistical analysis
Individual ID50 and IC50 values for each sample against each variant were calculated in Prism v9 (GraphPad Software) by fitting a four-parameter logistic curve to neutralisation by serial 3-fold dilutions of serum/antibody.
Author contributions
Conceptualization, D.J.S., T.P.P., B.M.;
Formal analysis, D.J.S.;
Conducted the assays, D.J.S., C.K., J.F., T.P.P.;
Designed the methodology, D.J.S., C.K., R.E., T.P.P, B.M.;
Responsible for figures and tables, D.J.S., T.P.P., B.M.;
Resources, S.M., R.E., S.R, N.K.B., G.B.K.H., J.A., B.M.;
Oversaw the study, D.J.S., G.B.K.H., S.T.R., J.A., B.M;
Funding Acquisition, S.T.R., G.B.K.H., J.A., T.P.P., B.M.
Writing – original draft, D.J.S., B.M.;
Writing – review & editing, D.J.S., G.B.K.H., J.A., T.P.P., B.M.
D.J.S and B.M. were responsible for the decision to submit the manuscript for publication.
Competing Interests
STR is a cofounder of and held shares in deepCDR Biologics, which has been acquired by Alloy Therapeutics. DJS, GBKH, and BM have intellectual property rights associated with antibodies that neutralise omicron variants. All other authors declare no competing interests.
Funding
This project was supported by funding from SciLifeLab’s Pandemic Laboratory Preparedness program to B.M. (Reg no. VC-2022-0028) and to J.A. (Reg no. VC-2021-0033); from the Erling Persson Foundation to B.M and G.B.K.H; by the European Union’s Horizon 2020 research and innovation programme under grant agreement no. 101003653 (CoroNAb) to G.B.K.H., S.T.R., and B.M; and by the G2P-UK National Virology consortium funded by MRC/UKRI (grant ref: MR/W005611/1) (T.P.P).
Acknowledgements
pCMV-dR8.2 dvpr was a gift from Bob Weinberg (Addgene plasmid # 8455; http://n2t.net/addgene:8455; RRID:Addgene_8455). pBOBI-FLuc was a gift from David Nemazee (Addgene plasmid # 170674; http://n2t.net/addgene:170674; RRID:Addgene_170674). We also wish to thank the clinicians and researchers, globally, who contributed to the sampling, sequencing, and identification of emerging SARS-CoV-2 variants.
Appendix
Supplementary Figures

{kind=link}
{kind=link}
Novel substitutions compared to the corresponding parental lineage are shown in black, with shared substitutions shown in grey. Depicted are BA.2, BA.4 and BA.5 (which have an identical spike amino acid sequence), BA.4.61, BA.2.75, BA.2.75.22, and BA.2.10.43.




