

Abstract
Alzheimer’s disease (AD) clinically presents with significant cognitive deficits in memory and executive function while pathologically displaying neuronal atrophy, the buildup of amyloid beta plaques and the presence of neurofibrillary tangles. Current therapies have modest effects on cognition and no effect on neurodegeneration. In AD, the GABAergic system shows reduced neuron populations, less GABA release and altered GABAA receptor (GABAAR) functioning. GABAergic somatostatin (SST) interneurons are particularly vulnerable. These SST cells target GABAAR’s containing the α5 subunit which are linked to cognition, but remain untargeted for therapeutic interventions. When lost, SST interneurons no longer coordinate signals and this presents as cognitive impairment. Using α5-GABAAR positive allosteric modulation (α5-PAM), there is the potential to restore their signaling and improve cognitive performance despite the presence of amyloid. Here we tested a selective α5-PAM (GL-II-73) at early (2 months) and late (5 months) stages of amyloid progression using the 5XFAD model for cognitive benefits, neurotrophic effects and amyloid clearance ability (N=48/study; 50% female). We found age-dependent deficits in spatial working memory and GL-II-73 dose dependently improves this deficit at 5 months of age. Chronic treatment with GL-II-73 showed spine density, spine count and dendritic length recovery at both time points. However, amyloid deposition progressed with age, and GL-II-73 did not have an effect on amyloid build up, suggesting that the effects observed are directly due to the impact of GL-II-73 on α5-GABAARs, and not related to amyloid plaques. These results show that despite the presence of amyloid GL-II-73’s α5-PAM activity overcomes cognitive deficits even at later stages of amyloid progression and demonstrates neurotrophic effects. Overall, results support GL-II-73 and its selective α5-PAM activity as a promising therapeutic option for cognitive deficits in AD.
Introduction
Cognitive deficits, such as altered executive functions, reduced working memory, impaired spatial memory or simply daily planning have been long reported in patients suffering from dementia and more specifically Alzheimer’s disease (AD)1. While being a very complex pathology with multiple underlying biology changes, one of the most reported hallmarks for AD is the buildup of amyloid beta plaques2. Clinical evidence support that elevated amyloid load in patients increases the risk for cognitive decline and for the development of Alzheimer’s disease3, even if therapies targeting the amyloid load still fail to alleviate the symptoms4. Four currently approved pharmacological therapies to help with symptoms include Donepezil, Galantamine and Rivastigmine which target the acetylcholine system as cholinesterase inhibitors, or Memantine which is a NMDA receptor antagonist aiming to inhibit aberrant neuronal activity5. These therapies however only offer modest cognitive symptom relief and do not prevent brain atrophy or neuron loss. Most recently FDA-approved and first of its kind for AD, Aducanumab is a monoclonal antibody therapy designed to clear amyloid from the brain. Results from this drug are controversial and studies are ongoing to confirm its efficacy6. Studies have found evidence for reduced amyloid load in rodent7 and AD patient brains8 however, correlating amyloid reductions to clinically relevant cognitive outcomes have been debated9. Therapies that are clinically efficacious for cognition are therefore still in high demand.
Designing therapies that target the underlying pathophysiology of AD can help efficacious therapy use. We know that AD and prodromal state risk factors include age, stress and genetics. A commonality between age, stress and familial AD is the reduction and dysfunction of somatostatin (SST) interneurons10-12. Somatostatin interneurons show a particular vulnerability in several disease states including AD and has been reproduced in transgenic rodent models13. This vulnerability has been linked to ER stress and the processing of immature preproSST14. In AD, it has been well documented that there are reduced SST expression levels, reduced SST+ neuron numbers and co-localization with β-amyloid plaques15-17. Functional SST cells in cortical layers mediate a “blanket of inhibition” that balances signal to noise ratios by inhibiting the distal dendrites of pyramidal neurons 18, releasing GABA that signals through both GABA-B 19 and GABA-A receptors.
GABA is an inhibitory neurotransmitter that signals through ionotropic GABAA-Rs and GABAB receptors (GABAB-Rs). GABAA-Rs are pentameric in nature and often consist of a combination of two α, two β and one other subunit type (γ, δ, ε, π or θ)20. Of the α-subunits, α1-α6 subtypes exist and each generally contributes to specialized functions. There is significant evidence showing α1 containing GABAA-Rs mediate sedative effects and are bound by benzodiazepines21. α5 containing GABAA-Rs seem to be more involved in learning and memory functionally and by their localization within the hippocampus and prefrontal cortex21-24. The α5-GABAA-Rs are located synaptically and extrasynaptically in distal dendrites of pyramidal neurons 25,26 where they regulate tonic inhibition via chloride ion influx into the postsynaptic pyramidal cell 27,28. The loss of SST / α5-GABAergic signaling alters information processing by cortical microcircuits and this contribute to cognitive dysfunction17. By targeting the α5-GABAA receptors pharmacologically, we can theoretically restore the “blanket of inhibition” and regulate temporal and spatial signal to noise ratios to improve cognition.
A ligand binding profile is important for therapeutic efficacy in order to produce the desired effect with binding specificity while limiting off target binding that can produce side effects. Previously, our lab has described an α5 selective positive allosteric modulator (α5-PAM) with importantly negligible potentiation at α1 subunits, making the potential therapeutic cognitive efficacy with lack of side effects highly desirable. Selectivity for α5-GABAA-Rs is therefore a potential therapeutic target for cognitive symptoms of AD.
We showed that the use of an α5-PAM has pro-cognitive properties in stress induced and aging models 24,29, which are models exhibiting SST deficits. In these models, we also showed that chronic treatment with an α5-PAM can increase dendritic length and spine density, on postsynaptic cells, which altogether suggests a combined symptomatic and disease-modifying effect of the intervention. However, there has been no work using transgenic models that express amyloid to consider potential benefits despite this pathological hallmark, and such demonstration could represent a breakthrough in the development of therapies that provide cognitive benefits as well as better brain functions in patients building amyloid load. Here, we use the 5XFAD amyloid progression transgenic model to investigate if acute or chronic α5-PAM interventions have pro-cognitive effects at early (2months of age) or late (5 months of age), if neurotrophic effects occur at these two time points with chronic treatment and if chronic α5-PAM has an effect on amyloid clearance.
Methods
Ethical Statement
All animal work was completed in accordance with the Ontario Animals for Research Act (RSO 1990, Chapter A.22), Canadian Council on Animal Care (CCAC), and was approved by the Institutes Animal Care Committees. For study timelines please refer to Figure 1.

Animals
5XFAD breeders (Jackson Laboratories; Cat #034840-JAX) were paired with WT breeders (Jackson Laboratories; Cat #10012). Experimental mice were generated in house using 5XFAD and WT littermates. “5XFAD” experimental animals were heterozygous and their WT litter mates were all aged in house to 2 months of age for “early stage” testing (Fig 1A,B) and 5 months of age for “late stage” testing (Fig1A,C). Based on breeding outcomes the following cohorts were used. 1) Acute studies N=46 ∼50% female, 2) Early stage chronic studies (2/3 months old): N=48, 50% female 3) Late stage chronic (5/6 months old): N=48, 50% female.
Animals were group housed until the beginning of the study at which time they were single housed. Animals had ad libitum access to food and water with the exception of food restriction outlined in the Noldus Phenotyper Cognition Wall paradigm. All animals were maintained on a 12 hour light/dark cycle. Animals were handled following the technique described in Marcotte et al 30 to limit their reactivity to the experimenter, critical for proper assessment of behavioral outcomes.
Drug Preparation
The PAM with preferential activity at the α5-GABAA-R (α5-PAM, code name GL-II-73) was synthesized in collaboration with Dr. Cook’s group (University of Wisconsin-Milwaukee) as described31. GL-II-73 was dissolved in vehicle solution (85% distilled H2O, 14%propylene glycol (Sigma Aldrich, 1% Tween 80 (sigma Aldrich) for acute administration, intraperitoneally (IP). The 2-3 month acute study used GL-II-73 at 10mg/kg for all behavoural tasks administered via IP. The 5-6 month acute study used GL-II-73 at 5mg/kg and 10mg/kg in the Y maze via IP injection. In both chronic studies, 30mg/kg of GL-II-73 was dissolved in tap water and given to mice in their water for per oral (PO) administration.
Behavioural Testing
All equipment was wiped down with 70% ethanol between trials and animals to remove olfactory cues.
Y maze
The Y maze uses spontaneous alternation to investigate spatial working memory. This test was adapted from the T maze paradigm described in Faucher et al.32 The maze consists of 3 arms (2 goal arms and 1 starting arm) each separated by 120°. The two “goal” arms have a sliding door near the center of the maze and closest to the experimenter is the starting arm with a sliding door at the distal end of the arm that is used as a “start box”. The first phase of the test involves a 2-day habituation phase where mice can freely explore the maze for 10 minutes each day. On the third day, a training phase is conducted. Training consists of placing the animal inside the start box and after a 30 seconds intertrial interval, the sliding door is opened. The mouse can then enter either of the goal arms. Once chosen, the sliding door of the goal arm is closed and the mouse remains in the chosen arm for 30 seconds before being returned to the “start box”. The chosen arm and latency to decide are recorded. This is repeated for 7 trials. The maze is wiped down with 70% ethanol between each trial to remove olfactory cues. On the 4th day a similar procedure is used however the intertrial interval was increased to 60 seconds and an 8th trial is conducted with a 10 second intertrial interval. This 8th trials evaluates an animals motivation to alternate therefore if no alternation is made the animal is excluded from the analysis to avoid biases from lack of motivation instead of cognitive performance. In acute studies, animals were injected with either vehicle or GL-II-73 30 min prior to testing. Mean % alternation was calculated using Mean alternation %=[(Number of alternations/number of trials)*100]. In the acute study 5 month study, 5mg/kg and 10mg/kg were tested separately in the same animals. Control and PS19 Vehicle animals ran the experiment twice and the average of their scores with completion of the 8th trial are reported.
Golgi Staining and Analysis
Golgi staining was used to investigate neuronal morphology. Brains were extracted and immediately placed in Golgi Solution for 2 weeks. Brains were then transferred to a shipping solution provided by and sent to NeuroDigitech (San Fransisco, CA, USA) for analysis. 100µm thick serial, coronal slices were made using a crystat from anterior to posterior. Slices were mounted on glass slides and the basal and apical dendrites of pyramidal neurons of layers II/III were identified as regions of interest (ROIs) in the PFC and CA1 of the hippocampus. NeuroLucida v10 (Microbrightfield, VT) software was used for quantification on a Dell PC computer and controlled a Zeiss Axioplan 2 image microscope with Optronics MicroFire CCD cameria (1600×1200). Motorized X, Y and Z focus enabled high resolution images and later quantification. Low magnification (10x and 20x) was used to identify ROIs with the least truncations of distal dendrites at high magnification (40x-60x). Using immersion oil and Zeiss 100x, 3D images were constructed to allow for spine counting along the complete dendritic tree. Inclusion and exclusion criteria were adapted from Wu et al.33 with inclusion requiring chosen neurons (n=6 per animal) to visually have the dendrites and soma completely filled, no overlapping with other soma and have complete 3D visualization of the dendritic tree. Exclusion criteria was neurons with incomplete staining and/or incomplete visualization. Spine sampling only included those that were well characterized and orthogonal to the dendritic shaft because those above or below are unable to be distinguished well. Raw data was extrapolated and quantified using NeuroExplorer (Microbrightfield, VT, USA).
Thioflavin S Staining and Analysis
Upon brain extraction one hemisphere was drop fixed in 4% PFA for 48 hours then immersed in 30% sucrose for 48 hours. Using a cryostat 30µm serial, coronal slices were taken and stored in cryoprotectant. The free floating Thioflavin S (ThioS) protocol involved 3 washes in PBS for 5 min followed by a 10 min wash in ThioS (1% weight/volume dissolved in ddH2O). 2 washes for were then completed in 75% Ethanol for 5 minutes each wash followed by rehydration via 3 washes in PBS, each for 5 min. Slices were then mounted on glass slides. Slides were imaged using an Olympus IX73 inverted LED fluorescence microscope and analyzed using Image J software. Image processing involved selecting the ROI (PFC, overall hippocampus), thresholding the image, converting to binary, converting to masks and analyzing particles.
Statistics
Y maze data was analyzed using One-Way ANOVA followed by Fisher’s LSD. Morphology Golgi stain results were analyzed using a Repeated Measure ANOVA with Tukey’s post hoc testing. Finally, the ThioS amyloid analysis used a Two-Way ANOVA with a post hoc Bonferroni Multiple Comparisons Test.
Results
For all statistical values please refer to Supplementary Tables referenced.
At 2 months of age, 5XFAD animals show some working memory deficits
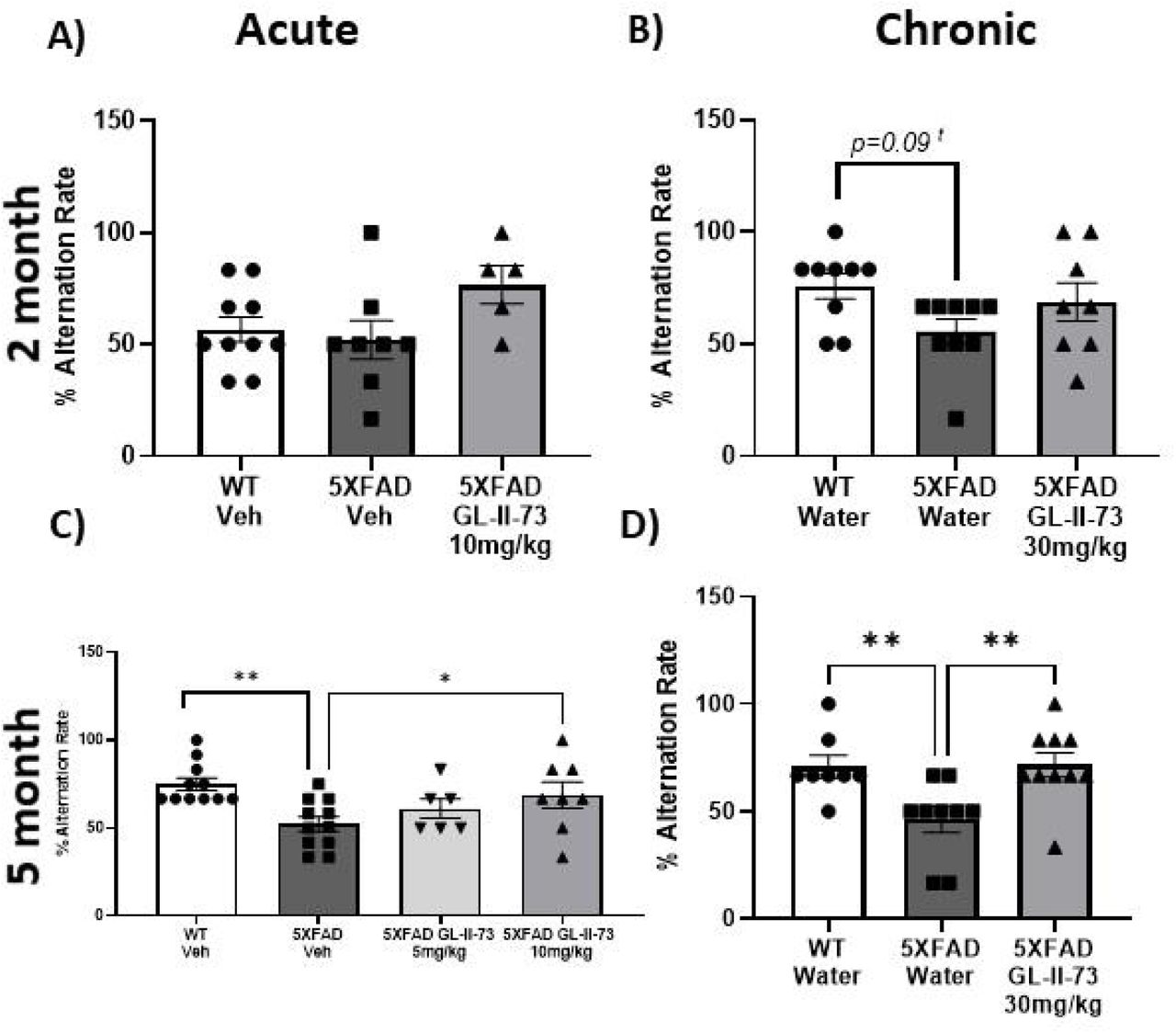
The Y maze was used to test spatial working memory in 2 month old 5XFAD mice, at an early stage in the amyloid 5XFAD model. In the acute study, WT animals performed poorly compared to what is expected from WT at this age, making the results inconclusive and one way ANOVA found no significant group differences (Fig.2A). However, in the chronic study ANOVA found a trend level difference between groups (Fig. 2B) (Supplemental Material Table 1). Results showed 5XFAD animals are beginning to show some spatial working memory deficits at this age.

At 5 months of age 5XFAD animals show a spatial learning deficit that is reversed by GL-II-73
The Y maze was used again to test spatial working memory when animals reached 5 months of age. At 5 months old one way ANOVAs with Fisher’s LSD post hoc testing found significantly lower alternation rates between WT and 5XFAD animals without GL-II-73 in both acute and chronic studies. In the acute study, the dose response of GL-II-73 to improve deficits was tested using 5mg/kg and 10 mg/kg (Fig 2C). Percent alternation of 5XFAD animals treated with 5mg/kg GL-II-73 did not significantly differ from 5XFAD vehicle or WT animals. 5XFAD animals receiving 10mg/kg GL-II-73 had significantly higher percent alternation than 5XFAD animals receiving vehicle (Supplementary Material Table 1). In the chronic study, 5XFAD animals receiving GL-II-73 showed improved alternation rate compared to 5XFAD animals with water (Fig. 2D) (Supplementary Material Table 1).
Neurotrophic effects are found in 2 and 5 month old GL-II-73 treated animals
Pyramidal neuron morphological changes in the PFC and CA1 of the hippocampus were quantified using Golgi staining in chronic studies at 2 and 5 months of age. Full statistics are found in Supplementary Material Table 2). In the PFC at 2 months of age, one way ANOVA found group differences and Tukey’s post hoc test showed spine density was significantly lower in 5XFAD water animals compared to WT and 5XFAD GL-II-73 animals. This was true in both basal and apical dendrites. No difference between WT and 5XFAD GL-II-73 mice were identified (Fig. 3A-F,J-L, P). 5XFAD water animals had lower spine count and lower dendritic length compared to WT and 5XFAD GL-II-73. This was again present at both basal and apical dendrites. In the CA1 of 2 month old animals, one way ANOVA detected group differences and Tukey’s post hoc found lower spine densities in 5XFAD water animals compared to WT and 5XFAD GL-II-73 mice in both basal and apical dendrites. The 5XFAD water group had significantly lower spine count and dendritic length compared to WT and 5XFAD GL-II-73 in basal and apical segments. In 5 month old animals, one way ANOVA detected group differences and Tukey’s post hoc testing found 5XFAD water animals had lower spine density in basal and apical dendrites compared to WT and 5XFAD GL-II-73 animals. Spine density of 5XFAD GL-II-73 was significantly lower than WT controls (Fig. 3 G-I,M-O,Q). ANOVA again found group differences in spine count followed by Tukey’s post hoc testing in basal and apical dendrites showing lower counts in 5XFAD Water animals compared to both WT and 5XFAD GL-II-Post hoc testing found no difference between WT and 5XFAD GL-II-73 in apical or basal dendrite spine count. Dendritic length group differences were detected with one-way ANOVA in basal and apical dendrites. Tukey’s post hoc testing found the 5XFAD water group had lower dendritic length than WT and 5XFAD GL-II-73 groups. In basal dendrites, 5XFAD GL-II-73 dendritic length was lower than WT controls but in apical dendrites there was no significant difference between these two groups.

GL-II-73 does not affect amyloid clearance
Amyloid load was investigated using ThioS staining and statistics are reported in Supplementary Material Table 4. In 5XFAD animals the amyloid load increased over time as expected. 5 month old mice had higher amyloid plaque density in both the PFC and the hippocampus compared to 2 month old mice receiving the same treatment (Fig.4 A-N). The 5XFAD model uses human transgenes regulated by the constitutive, brain specific Thy1 promoter, therefore we used amyloid plaque density to determine if GL-II-73 had an effect on amyloid clearance. One way ANOVA with Tukey’s post hoc testing at both 2 and 5 months of age found the Het water group had significantly high plaque density than WT controls in the PFC and hippocampus of that age. In both brain regions, Het GL-II-73 animals had significantly more amyloid compared to WT and were not significantly different from Het water animals.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Discussion
This study was designed to investigate the impact of modulation of α5-GABAA receptors at early and late amyloid deposition to determine functional, morphological and histological effects related to cognition and pathology. This work was driven by observations that SST interneurons and the GABAergic system is affected in AD, and that selective α5 GABA-A-R modulation has pro-cognitive effects in models of stress and aging which are also well documented risk factors for AD24,29. Inthe 5XFAD model of amyloid load, we tested acute and chronic GL-II-73 at 2 months of age and 5 months of age on spatial working memory. We found that amyloid load happening in the 5xFAD mouse brain induced cognitive decline, which was reversed even at later stages of amyloid deposition with GL-II-73 in a dose dependent manner and with chronic administration. We also demonstrated that chronic treatment reverses dendritic shrinkage and spine loss, at both ages and in both regions investigated being the PFC and CA1 of the hippocampus, where α5-GABAARs are expressed. However, and as expected, chronic treatment does not change the amyloid load, showing that the effects observed are mediated by activity on the pyramidal cells directly, but do not alleviate amyloid build-ups.
1. Age dependent deficits are found in spatial working memory and GL-II-73 dose dependently improves the deficit at 5 months of age
First, we characterized spatial working memory deficits at early (2 months of age) and late (5 months of age) time points in the 5XFAD amyloid load model and determined if GL-II-73 was able to reconcile deficits. At our earlier time point, we found trend level spatial working memory deficits that are consistent with the timeline of other 5XFAD cognitive decline onset34,35. Also, consistent with previous characterization of the 5XFAD model, the spatial working memory deficits progressed with age becoming significantly impaired by 5 months of age. At 5 months of age, we found GL-II-73 had a dose dependent effect in remedying the cognitive deficit. This is consistent with our previous work showing dose dependent benefits of 10mg/kg over 5mg/kg in the unpredictable chronic mild stress model and aging model24,29. Here, the 5 month chronic study also showed spatial working memory deficits reversed by GL-II-73. This supports that a5 GABAA-R modulation is able to recover deficits at later stages of amyloid deposition, providing symptomatic relief.
2. GL-II-73 has neurotropic effects on pyramidal neurons in the PFC and CA1
The progression of neuronal degeneration in AD is detrimental to neuronal function, behaviour and cognitive performance36. The 5XFAD model clearly demonstrated neuronal degeneration through the loss of spine density, spine count and lower dendritic length. Interestingly, GL-II-73 showed neurotrophic effects across the PFC and the hippocampus at early and late time points assessed. This could have clinical relevance because there is potential for this intervention to induce neurotrophic effects even at later stages of the disease to restore functionality to remaining neurons. The mechanism for neurotrophic effects is unknown and would be of interest in future studies. Ketamine studies have proposed GABAergic disinhibition activates AMPARs that stimulates TrkB and BDNF neurotrophic pathways37,38.
3. Amyloid clearance is not affected in PFC or hippocampus by GL-II-73
The 5XFAD model uses the constitutively active Thy1 promoter to induce the transgenes that increase amyloid load. Here, amyloid was visualized using ThioS staining. We showed very clear evidence that the model increases amyloid density with age in the PFC and the hippocampus and investigated if GL-II-73 was able to affect amyloid clearance. We did not identify an effect on clearance by α5-GABRA modulation, however despite no change in amyloid load there was still behavioral cognitive improvement which could be clinically relevant. Cognitive improvements seen here can likely be attributed to GL-II-73 modulating a5-GABRA activity in combination with the neurotrophic effects on pyramidal neuron spines.
Limitations
Limitations to consider here are the poor performance of the WT animals in the 2 month acute study. This will be repeated and retested with 5mg/kg dose to see any dose dependent changes. Another limitation of this work is that the 5XFAD model cannot determine if GL-II-73 can impact plaque deposition and this should be studied in a different model. Also important to note, is that Alzheimer’s disease is not restricted to amyloid load, so while interesting in nature, this data does not predict efficacy in patients with more complex pathophysiology, but this intervention demonstrats efficacy at reversing cognitive decline and neuronal atrophy despite amyloid buildup.
Conclusion
In the 5XFAD amyloid load model, acute and chronic a5-GABRA modulation using GL-II-73 improved spatial working memory deficits that occur at stages of early and later amyloid deposition. Chronic administration with GL-II-73 also produced promising morphological changes. Together this data supports the role of a5-GABRA modulation in cognition and demonstrates therapeutic relevance for a5-GABRA modulation for diseases such as AD.
Funding
This work has been funded by the Weston Brain Institute (TR192043).
Disclosures
JMC, MM, ES and TP are listed inventors on patents covering syntheses and use of the compound. ES is Founder of Damona Pharmaceuticals, a biopharma dedicated to bring novel GABAergic compounds to the clinic. AB, KW, KP and DS declare no conflicts of interest.
Acknowledgements
Authors thank Mehrab Ali for support with administrative tasks throughout the study. They thank CAMH animal facility staff for the caring for our animals over the study duration. Authors further thank the members NeuroDigitech for their contribution to data generation.
References




