

Abstract
Male crickets attract females by producing calls with their forewings. Louder calls travel further and are more effective at attracting mates. However, crickets are small, and therefore inefficient, dipole sound-sources. Only a small group called tree crickets make acoustic tools called baffles which reduce acoustic short-circuiting, a source of dipole inefficiency. Here, we ask why baffling is uncommon among crickets. We hypothesize that baffling may be rare, because like other tools they offer insufficient advantage for most species. To test this, we modelled the calling efficiencies of crickets within the full space of possible natural wing sizes and call frequencies, in multiple acoustic environments. We then generated efficiency landscapes, within which we plotted the positions of 111 cricket species across 7 phylogenetic clades. We found that all sampled crickets, in all conditions, could gain efficiency from tool use. Surprisingly, however, we also found that calling from the ground significantly increased efficiency, with or without a baffle, by as much as an order of magnitude. We found that the ground provides some reduction of acoustic short-circuiting but also halves the air volume within which sound is radiated. It simultaneously reflects sound upwards, allowing recapture of a significant amount of acoustic energy through constructive interference. Thus, using the ground as a reflective baffle is a very effective strategy for increasing calling efficiency. Indeed, theory suggests that this increase in efficiency is accessible not just to crickets, but to all acoustically communicating animals whether they are dipole or monopole sound sources.
Significance Statement Loudness is a crucial functional feature of calls in acoustically communicating animals. Animals attempting to reach prospective mates or ward off predators are expected to make themselves as loud as possible. Given the importance of loudness, there are two long-standing, seemingly unrelated paradoxes in acoustic communication. The first is the rarity of acoustic tool use. The second is the high number of animals that call from reflective surfaces, like the ground, known be an impediment to sound propagation. We resolve both paradoxes and show them to be related. By refocusing analysis from sound propagation to sound radiation, we show that the ground is the opposite of an impediment and can, in fact, boost sound loudness more than tool use. Thus, we show that calling from a reflective surface is an alternative strategy for maximizing call loudness, and one that is available to all animals.
Introduction
Male crickets make loud advertisement calls to attract females who use these calls to locate mates (1). Louder calls travel further, cover more area, and attract more females (2–4). When faced with a choice, females prefer louder calls (2, 5). Being louder therefore has implications for mating success and evolutionary fitness in these singing insects. However, despite the apparent loudness of a nighttime chorus, cricket calls are acoustically constrained by a phenomenon known as ‘acoustic short-circuiting’ specific to dipole sound sources (6, 7). Cricket wings are sound radiators that vibrate back and forth in the air like pistons. As a wing moves in one direction, the air in front of the wing is compressed, and the air behind it is rarified. These two changes in pressure travel away as waves as the motion is periodically repeated. However, the waves on either side of the wing are of opposite phase and interfere destructively where they meet, at the edges of the wing. Thus, less sound is radiated, reducing sound radiation efficiency (6, 8). The smaller the wings of a cricket with respect to the wavelength of the sound it makes, the higher the short-circuiting and associated loss of efficiency. Indeed, the few crickets that have been studied are small and experience significant short-circuiting (9).
To reduce the efficiency lost to acoustic short-circuiting, a few tree cricket species build and use an acoustic tool known as a baffle (5–7, 10). A baffle consists of a leaf with a hole chewed by the cricket near the middle of the leaf. When the size of the leaf and hole are optimal, such structures reduce acoustic short-circuiting and increase efficiency by as much as 10 dB compared to unbaffled calling, reflecting a tripling of sound pressure levels (7). However, despite their benefits, only a handful of species among thousands make baffles, all within the sub-family Oecanthinae (5, 7, 10, 11).
Given the obvious benefits, why is acoustic baffle use rare in crickets? Tree cricket baffles are tools, and tool use is generally rare (12–14). Indeed, few species use tools, whether crickets, other invertebrates or even vertebrates. Invertebrate tool use, however, seems especially rare. For example, 56 independent occurrences of tool use were found in mammals, whereas only 13 occurrences were found in the significantly more speciose insects (14). Two hypotheses from the tool use literature, the “cognitive capacity” and the “lack of utility” hypotheses offer two different reasons for this rarity. The “cognitive capacity” hypothesis suggests that complex tool use behaviors are less likely to evolve in animals with smaller brains and lower cognitive capacity. This is an unlikely explanation since many animals with relatively low cognitive capacities do use, and even make, tools which themselves are not necessarily complex objects. Small-brained animals are even known to make very complex and highly functionally optimized habitation structures which do not require high cognitive capacity (7, 15).
A competing hypothesis is the “lack of utility” hypothesis which posits that tool behavior can evolve regardless of cognitive capacity, but that its evolution requires an ecological context in which it confers sufficient selective advantage (15). Only species that can achieve higher gains from tool use than from other strategies (e.g., morphological features, site selection) are likely to evolve tool using behavior.
To test the lack of utility hypothesis, we must quantify tool utility and use of the tool must have implications for evolutionary fitness. It is often difficult to meet these two conditions. Work in chimpanzees has directly quantified tool utility by evaluating how much caloric value can be gained by using a tool to exploit an otherwise unexploitable resource (16). Other studies have made more indirect arguments; work in sea otters has shown that tools are employed more frequently in populations in which tough prey require tools to access them (17). In capuchin monkeys, larger individuals who can more effectively use tools to crack nuts are more likely to use tools (18). However, few studies quantitatively assess the lack of utility hypothesis, particularly outside the context of food.
Baffle use in crickets is an ideal system in which to test the lack of utility hypothesis. First, baffle use is rare and second, we can directly measure its acoustic utility in terms of increase in sound radiation efficiency (12). Finally, baffle use has been shown to have real fitness implications, by increasing the number of mates attracted to a given male, and also by increase mating duration, both processes likely to increase reproductive success (2).
Therefore, in this study, we tested the lack of utility hypothesis across a large group of singing insects, the true crickets or Grylloidea. We used finite element models to quantify baffle utility in two ways. First, we ascertained the range of sizes of the sound radiator (cricket wings) and frequency ranges of the calls used by 111 crickets, spread over the cricket phylogeny and used it to define an acoustic-morphospace (Fig 1). Then we quantified a volumetric sound radiation efficiency (SREv), averaged across all positions in this space, specifically the relation between the radiator vibration amplitude, and the sound levels that are generated, similar to the previous study on tree cricket baffle efficiency (7). Since the amplitude of the radiator vibration would reflect the effort applied by the animal, this measure of efficiency captures a significant component of the relationship between singing effort and output. By plotting SREv over the complete acoustic-morphospace, we were able to generate efficiency landscapes, which encompassed baffled and unbaffled crickets and enabled us to fully investigate all possible crickets, even those that did not appear in our sample.
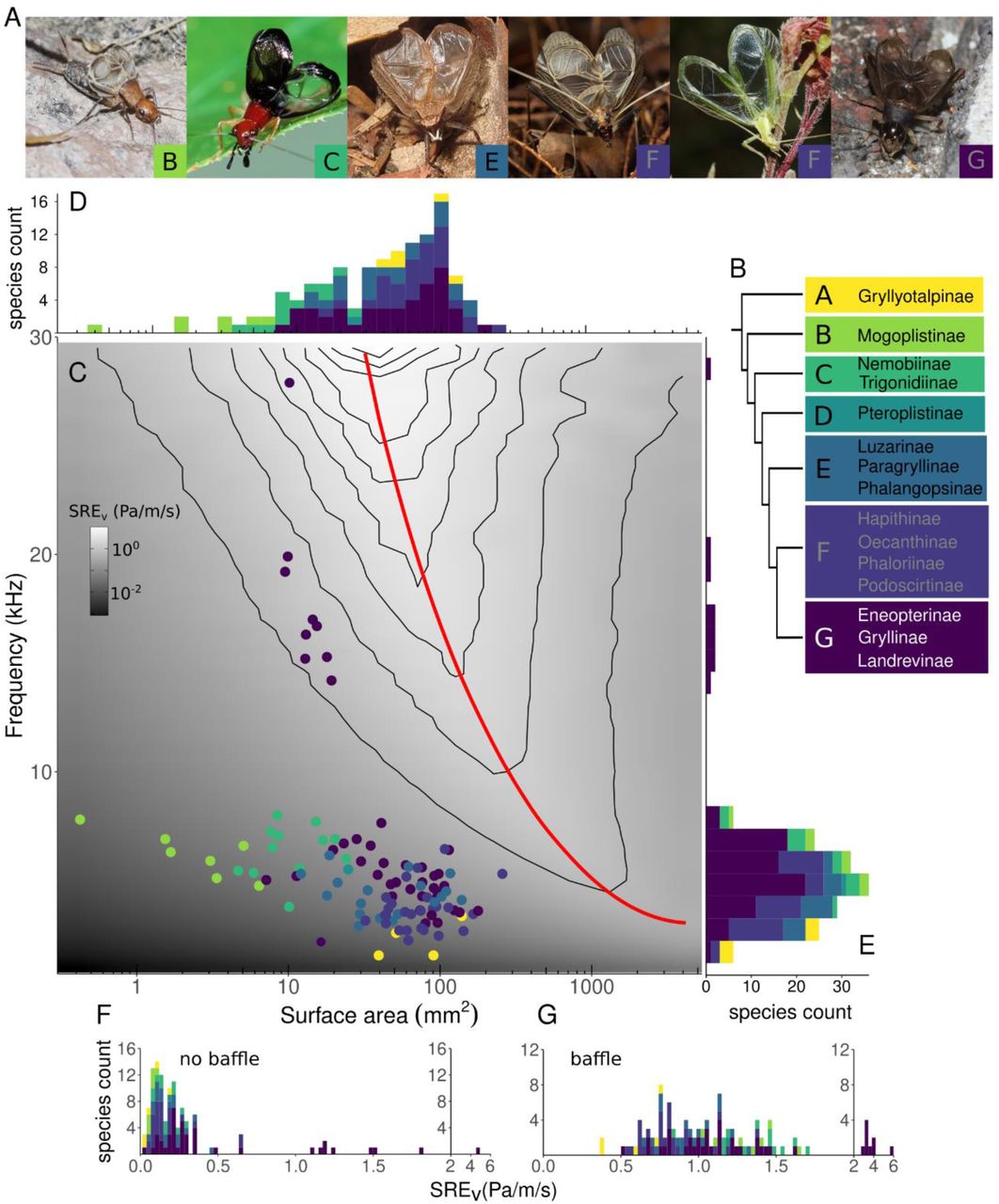
All crickets could increase efficiency by baffled calling. Sound radiation efficiency (SRE) landscape across the acoustic-morphospace of crickets. A. Representative images of cricket males with wings raised in calling posture. From upper left to lower right, species pictured are: Hoplosphyrum boreale (photo: James P. Bailey), Phyllopalpus pulchellus (photo: Wilbur Hershberger), Lerneca inalata (photo: Richard C. Hoyer), Meloimorpha japonica (photo: Ryosuke Kuwahara), Oecanthus quadripunctatus (photo: James P. Bailey), and Turanogryllus eous (photo: Taewoo Kim). Boxes with letter indicate the clade to which each species belongs. B. Phylogeny illustrating the seven clades defined by Chintauan-Marquier et al (2016) along with subfamilies within each clade (branch lengths not to scale). C. SREv achieved by different combinations of wings sizes vibrating at different frequencies. Within this space of possibilities the wing areas and call frequencies of all sampled animals are shown overlaid as points on the SRE landscape. This SREv is calculated from finite element models. Red line indicates optimal efficiency ridge, or the size at each frequency that would produce an ideally baffled calling scenario. D, E. Distributions of the calling song frequency and wing size of different animals, respectively. Histograms include additional species for which only wing or call measurements were available. F. SREv of each species without use of a baffle. G. SREv of each species with use of an ideal baffle.
We also considered natural ecological conditions in which crickets call, such as, from close to the ground, and from within vegetation. In both these cases, the environment interacts with the sound radiator and the sound emanating from it across spatial scales, and may effectively remove any gains from baffling (19, 20). To capture spatial effects, we generated a second metric sound radiation efficiency, this time measured along a transect, (SREt). Here we used the boundary element method and quantified the effect of interacting acoustic surfaces on acoustic efficiency as sound propagates away from the singer, under a range of environmental conditions.
Here we considered calling from the ground more carefully, in terms of its effect on efficiency. In the animal communication literature, typically the ground is considered during propagation and not in relation of sound radiator efficiency. In this context, it is typically seen as a severe impediment to sound propagation by causing significant amount of excess attenuation (21–24). Researchers also focus on the ground effect as degrading temporal structure (25) and directional information (26, 27). While we cannot address temporal structure in our examination of sound radiation efficiency, excess attenuation is accounted for. Additionally, using the same models, we also quantified directionality to test how efficiency might trade off with this biologically crucial feature.
Using these data, we asked whether the rarity of baffle use in crickets is explained by the lack of utility hypothesis. We examined the differences between baffled and unbaffled calling in different realistic scenarios. We expect that known baffle-users will be animals who benefit most from baffle use, and non baffle-users might not accrue as many benefits due to acoustic, morphological or environmental constraints.
Results
All crickets would benefit from baffle use in idealized conditions
To capture the natural range of wing sizes and calling frequencies among true crickets, we collected wing surface area and call frequency data for 111 cricket species from a large range of sources (Fig 1, Tables S11, S12, Fig S1). Species were distributed across 7 clades as described by the most recent phylogeny of the Grylloidae or “true cricket” super family (28) (Fig 1, SI 1).
We then constructed finite element models which predicted the sound fields produced by wings of different sizes at different call frequencies for 1086 different combinations which encompassed all the observed frequencies and wing sizes, i.e. the full acoustic-morphospace (Fig 2). In all conditions, wings were modelled as suspended in free space, vibrating with a uniform velocity perpendicular to the wing plane (SI 2). The model predicts the resulting sound field (see Supplementary methods for details). We then calculate sound radiation efficiency (SREv (Pa/m/s)), by taking a volumetric average of the sound pressure level generated (Pa), over a sphere of radius 20 cm around the wing, divided by the time-space average of the wing vibration velocity (m/s). This normalized measure of efficiency enables comparison between species, no matter their actual wing velocity or sound pressure level.

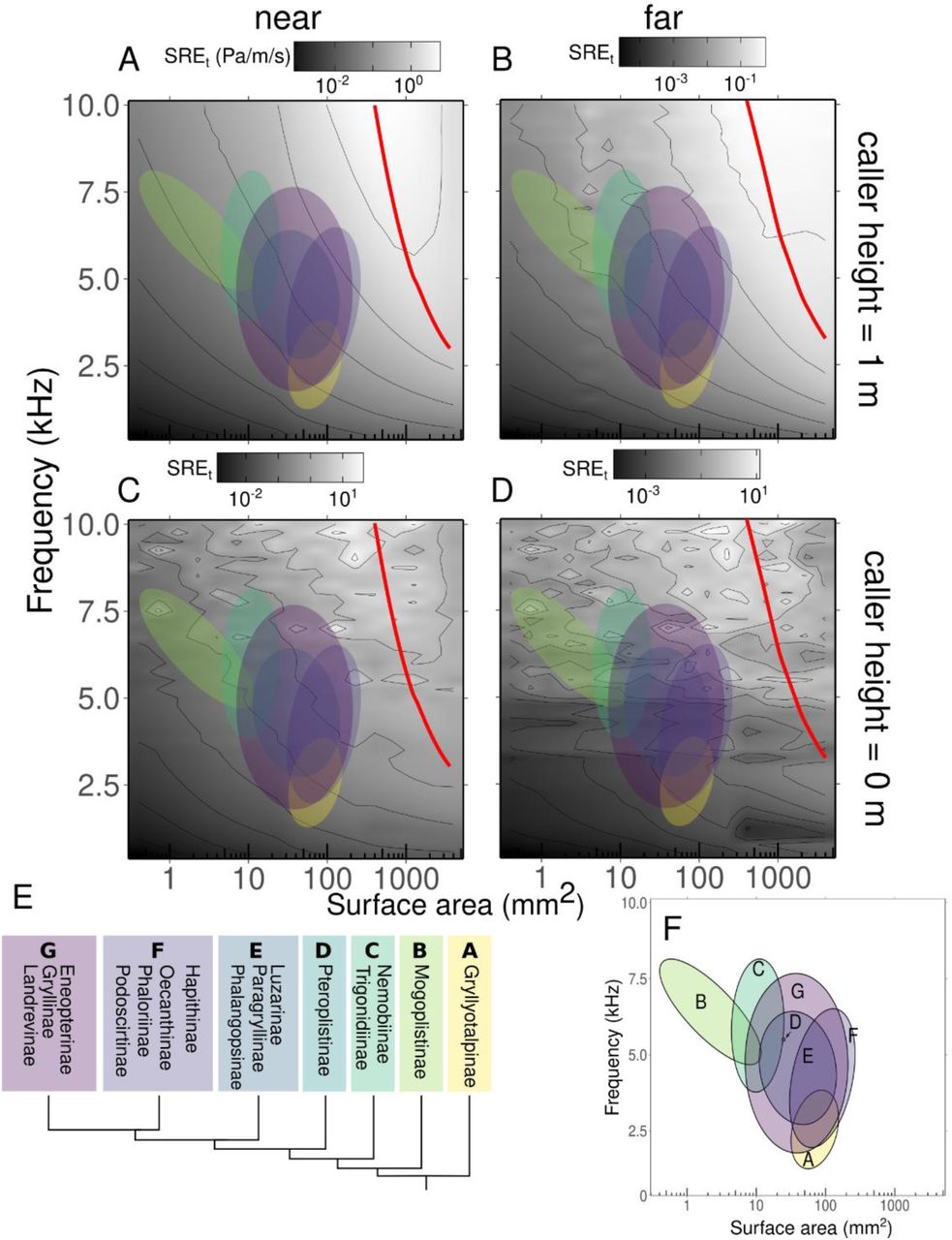
Sound propagation efficiency when the cricket sings near the bare hard ground is orders of magnitude higher than in free space. Each panel represents a combination of caller height above ground (0 m or 1 m) and receiver distance from caller (0.05 – 02 m “near” and 0.8 – 0.9 m “far”). A. Distance = near, height = 1m, B. Distance = far, height = 1 m; C. Distance = near, height = 1 m, D. Distance = far, height = 0 m. Red lines indicate optimal efficiency ridge, or the size at each frequency that would produce an ideally baffled calling scenario. Note that the frequency range is reduced compared to figure 2, in order to exclude high frequency callers which likely use alternative vibrational modes. Each clade of animals is represented by a colored ellipse. E. Phylogeny representing each clade F. Key to clade represented by each ellipse.
Next we plotted an SREv landscape (Fig 1) for the full acoustic-morphospace. On this landscape, we plotted the locations of the 111 species of crickets with known wing size and frequency allowing us to infer their SREv (Fig 1). These data therefore demonstrate precisely how suboptimal crickets are in terms of their efficiency (Fig 1F), and how much they could gain through use of an optimal baffle (Fig 1G).
Previous work examining four species of crickets and a small number of other insects determined that they each perform less efficiently than the theoretical optimal level for dipole sound sources (9). In our larger dataset, there is a clear ridge of high efficiency running through the SREv landscape (red line in Fig 1c, hereafter referred to as “optimal efficiency ridge”), which shows the optimal radiator size for every frequency. Despite sampling species with a wide range of wing areas (0.4 – 258 mm2) and call frequencies (1.6 – 27.9 kHz), all species lie below this optimal efficiency ridge. The efficiency distribution among crickets is somewhat bimodal (Fig 1F, G). The nine species with an SREv above 1 Pa/m/s all had calling frequencies above 14 kHz and belonged to the subfamily Eneopterinae, in clade G. Given the difference in their song radiation mechanics (29) we excluded these individuals from subsequent analyses (see supplementary methods). After excluding these high frequency callers, we found that SREv ranges from 0.02 to 0.67 Pa/m/s, mean: 0.18 ± 0.01 SE, n = 101.
Next, we calculated how much each species could gain simply by using a baffle. To calculate this gain, we used the optimal efficiency ridge, since optimal radiator efficiency is the same as optimal baffled efficiency (8). If animals were to continue using the same call frequency, but used an ideal baffle, each species stood to gain between 1.7 – 35 times (5 – 30 dB) above their baseline SREv (mean: 7.6 ± 0.41 times, 16 ± 0.37 dB, n = 99). Among those who stood to gain the most included animals in clade B, specifically in the subfamily Mogoplistinae (scaly crickets). These animals have very small wings (mostly under 5 mm2), suggesting a poor match between sound wavelength and wing size. On the other hand, the animal closest to the optimal efficiency ridge, was Madasumma affinis, belonging to the subfamily Podoscirtinae in clade F. This animal has the largest wing at 258 mm2, however, even this animal stood to gain 0.5 Pa/m/s (5 dB or 1.6x increase) with the use of an ideal baffle. Taken together, these data suggest that all crickets could increase SREv, and therefore, stand to benefit from use of a baffle.
Ground calling emerges as an alternative strategy to baffle use in complex environments
While analysis of SREv suggests that all crickets should use baffles, this prediction is based on sound fields travelling in free space and over short distances. It is possible that efficiency advantages from baffle use become negligible as sounds interact with objects in the cricket’s local environment such as the ground or the vegetation. Many baffle users are low frequency callers, and it is also possible that higher frequency crickets lose all advantage from baffling in complex acoustic conditions. Either of these scenarios would lend support to the lack of utility hypothesis.
To address whether and how the efficiency landscape is changed by realistic calling conditions, we used boundary element modeling. Specifically, we used this method to add a “ground” component to our existing models, where the ground could have different characteristics including vegetation cover. In these models, sound can be reflected and dissipated by the ground and the effect of the vegetation is captured by an excess attenuation term based on empirical data (see supplementary methods). We used empirical measurements of ground impedance and although our modeled ground is flat and smooth, these measures should take realistic ground variability into account. We also varied the height of the caller above the ground (ground calling: 0 m, elevated calling: 1 m). We measured efficiency again by normalizing sound levels against radiator vibration levels (see methods). Sound levels were measured at two distances from the caller: near (averaged from 0.05 – 0.2 m away), and far (averaged from 0.8 – 0.9 m away). To simplify analysis, we always measured efficiency at the same height as the caller. To differentiate this measure of efficiency from SREv, we call it sound radiation efficiency along a transect, or SREt (Pa/m/s).
The most striking and unexpected result from our analysis was that calling from the ground (Fig 2C, D) yielded much higher SREt than calling from one meter above it (Fig 2A, B). This is reflected in the efficiency landscapes by an average increase of about 5x (14.5 dB) across the entire acoustic-morphospace that we measured. Indeed, the highest SREt observed with a ground caller was 4 Pa/m/s (Fig 2C), two orders of magnitude higher than peak SREt with an elevated caller (0.06 Pa/m/s, Fig 2A). This increase in efficiency is likely due to two phenomena. The first is partial baffling offered by the ground, which will prevent some acoustic short-circuiting. A second phenomenon is likely the ground effect, in which the pressure field that would normally propagate below the radiator is instead reflected upward from the ground and mixes with the directly propagated field. Here, we see that the ground effect leads to constructive interference between the direct, and the reflected and the ground wave and an increase in sound pressure levels (30). We find that even optimally baffled animals could gain an average of 2.8x efficiency (9.2 dB) along the measurement transect, by calling from the ground compared to elevated calling.
On the other hand, calling from far above the ground yields SREt values that are similar in level to SREv values calculated in the ideal free-field scenario modeled previously. At further distances, the values decrease as predicted by spreading in open air. Taken together, our models posit that ground calling and elevated baffled calling are two potential alternative strategies to maximize efficiency.
Calling from the ground is still efficient when ground properties and vegetation are varied
All grounds are not equivalent and the increase in the net increase SREt may depend on their properties. For instance, soft grounds or those covered with vegetation would be much more dissipative and may eliminate the advantage accrued from ground calling. To test this possibility, we investigated whether this alternative strategy framework holds up when these properties of the environment are varied. We found few differences in SREt with different types of grounds (SI 7). SREt tends to be slightly higher with a “soft” ground, which is better at dissipating sound, similar to a freshly tilled agricultural field (see methods) and this effect is magnified further away from the caller. This suggests dissipation has a higher effect further from the source, and primarily on propagation, whereas here the phase shift is more appropriate for constructive interference near the source. With a harder, more reflective ground, similar to a tightly packed forest floor, SREt is slightly lower. However, significant differences between these two ground types occur at wing sizes well outside the natural range for crickets. At close distances, and particularly above the ground, the differences between ground types are very small (SI 7). Therefore, all future analyses assume a “hard” ground.
Finally, we tested whether vegetation would reduce the predicted SREt landscapes for ground calling. Vegetation does slightly decrease the magnitude of SREt overall and unsurprisingly, there is a slight frequency dependence where SREt is lowered slightly more at high frequencies (Fig SI 8). This suggests that high frequency callers may be at an increased disadvantage when calling in vegetation as suggested before (23), and will see diminishing returns when using a baffle. However, we found that excess attenuation due to vegetation does not significantly change the overall patterns of efficiency. By and large, it shifts the efficiency landscape to a lower point at most points within the parameter space (Fig 2, SI 8) (31). However, the efficiency near the ground remains higher than the efficiency 1m above the ground (SI 9). Finally, the effects of vegetation on SREt are undoubtedly more complicated than an excess attenuation factor. Modeling plants explicitly, at a variety of shapes and sizes, would be a useful extension to this study. However, since the efficiency of ground calling remains higher than calling from 1m above ground, we conclude that calling from the ground remains an effective alternative strategy, even if the ground is soft, or covered with some vegetation.
Ground calling does not significantly degrade call directionality
So far, our analysis has used the loudness of calls to define efficiency. However, a call must be both loud and directional to be effective. That is, the call must present a spatial gradient that a potential mate can follow to the source. Previous data has suggested that such gradients are severely degraded in ground calling crickets (23, 27, 32, 33), but not in elevated calling (34). This suggests that SREt gains from ground calling may trade off against call directionality. To test this, we analyzed call directionality by designing a directionality metric that captured how difficult it would be for a female cricket to follow an acoustic gradient back to the call’s source (see methods). A value of one indicates that the gradient along a transect perpendicular to the wing planes is always in the “correct” direction, that is, sound pressure level increases as the female moves toward the caller in steps of ∼ 2 body lengths (2 cm). A lower value means that over some stretches of this transect, SPL increases and at other steps it decreases. A value of 0.5, for instance, means that the SPL decreases over 50% of the steps as the female moves closer.
We find that directionality varies with respect to frequency, radiator size, and height from ground (Fig SI 10). Although ground calling does experience a loss of directionality compared to elevated calling, these losses are small. Near a ground caller, calls are all strongly directional (> 0.9) below about 5 kHz, except for very small wings. Further from the caller, calls are strongly directional below about 3.5 kHz. Therefore, high frequency callers would be most susceptible to the gradient effects. However, even below these cutoffs, directionality rarely drops below 0.5 in any condition, and ground calling remains a viable strategy. It should be noted that other studies have found more substantial degradations in call directionality in sounds traveling along the ground, but over greater distances than our current models (27). However, data for both field crickets and tree crickets suggest that the SPL of typical cricket calls drop below threshold at about 1 m from the caller (32, 34), and therefore we considered this a biologically relevant distance over which to measure directionality.
Alternative calling strategies are likely in use by some cricket species
Based on the overall efficiency landscape, ground calling and baffled calling are potential alternative strategies to maximize efficiency. However, we have so far considered the full acoustic-morphospace, i.e. all possible combinations of radiator (wing) size and call frequency, but most of these combinations are not used by real crickets.
To shift our focus to the sampled crickets, we tested where alternate calling strategies may offer the largest advantage to actual crickets. We divided all sampled animals into their respective clades and calculated clade level SREt for each of three alternative strategies as measured far from the caller: calling from the open hard ground (ground calling), and from within vegetation 1 m off the ground with no baffle (unaided calling), and from within vegetation with a baffle (baffled calling) (Fig. 3). We compared both baffled calling and ground calling to unaided calling as a baseline. It would have been ideal to determine whether animals, in fact, use the strategy that we predict should maximize efficiency based on known calling preferences. Unfortunately, we do not have data on calling preferences of many sampled animals. However, some clade-level similarities have been observed in calling behaviors, and different clades show some clustering in the wing size-frequency space (Figs 3, SI 9). We give three examples below of clades with some known information about calling behavior.

The most effective calling strategy (ground vs baffled) varies depending on clade, however directionality does not vary significantly. A, B. Comparison of SREt on bare ground with no baffle, and 1 m in vegetation with and without an ideal baffle. These measurements were taken far from the caller, i.e. an average of the SPL at a distance of 0.8-0.9 m from wings, directly in front of the wings was used. A. indicates extreme outliers, which only occur when we consider animals calling from the bare ground. Baffled and ground calling do not substantially differ with respect to directionality, although it does vary somewhat by clade. C. Directionality in near condition, D. Directionality in far condition.
We start with clade F, consisting primarily of the baffle making Oecanthines, or tree crickets. Members of this group stand to gain efficiency on the order of about 4.5x, (13 dB) from ground calling compared to unaided calling according to our data (Fig 3). However, they could gain 9x, (19 dB) it they baffled. Indeed, what is known of Oecanthine natural history bears out our predictions; tree crickets are known to mostly call from vegetation, including vegetation that is suitable for baffle building and use (6, 34). In fact, all known cricket baffle users are in this clade, as predicted based on the lack of utility hypothesis. For clade G, on the other hand, consisting mostly of the Gryllinae, or field crickets, we predict the opposite. On average, ground calling gives an advantage of 9x (19 dB) above unaided for species in this group, whereas baffling gives an advantage of about 8x (18 dB) above unaided. Again, behavioral data suggests that many field crickets indeed prefer to call from the open ground habitats that we predict would maximize their efficiency (35).
Clade A, the Gryllotalpidae or the mole crickets, represents an interesting exception to this alterative strategy framework. This group stands to gain the most from baffled calling of all clades (13x, 22 dB) compared to ground calling (2x, 6 dB). Yet, species in this group are all known to exclusively call from the ground and do not use baffles. However, they do use an acoustic aide. Mole crickets build and call from burrows which function as resonators and convert them into monopole sound sources, eliminating acoustic short-circuiting through a different mechanism than baffling (22, 36). Indeed, it is possible that other acoustic means of maximizing call efficiency exist and could in the future add further complexity to our hypotheses.
Finally, if an animal baffled, but its call propagated though vegetation compared with no vegetation, the gains would be relatively small in most cases (< 6 dB SPL) (SI 9), We also performed a similar analysis for call directionality (Fig 3C, D). However, since directionality was quite high for all calling conditions, we therefore suggest that directionality does not preclude one alternative strategy over another.
Discussion
Why would baffle use evolve among crickets in the first place?
From our data, exploiting the ground effect by calling from the ground emerges as a viable alternative to tool use in crickets. This simple site selection strategy can even exceed the efficiency gains of tool use in some scenarios. Given that making and using tools like baffles requires a specialized behavioral repertoire, and precise execution of these behaviors (7, 12), the real question becomes why a species would ever use this strategy if simpler site selection preference for the ground could give similar increases in efficiency.
There is evidence that crickets have been singing as early as the Cretaceous period (37). These early calling crickets were likely ground dwellers, with some species subsequently moving up into vegetation as the group diversified (38). We therefore suggest that baffle-using crickets may have originally moved up into vegetation for non-acoustic reasons, whether it was to exploit additional food resources or avoid predators. Baffle use would have then evolved secondarily to recover some of the efficiency lost when abandoning ground calling. The biophysical modeling methods presented here open the door to testing such a hypothesis about baffle use.
It is also known that crickets call from other “ground” substrates such as tree trunks, cave walls, or artificial structures (19, 22), which our data suggest could also increase calling efficiency (35, Fig 4C, D). The efficiency effects of these substrates could be further investigated using biophysical models contributing to our knowledge of acoustic ecology. In principle, we could even model the wings of extinct crickets, and estimate calling frequency based on the stridulatory apparatus on the wing (39, 40). By bringing extinct crickets “back to life” in this way we could ask questions about the evolution of acoustic tool use and calling ecology more broadly. We suggest that biophysical modeling, grounded with data from real animals, can be a valuable tool for any biologist wishing to better characterize and understand tool use in the context of animal communication.

Hard reflective surfaces like the ground or water offer all acoustically active animals an increase in sound radiation efficiency. A. The presence of a perfectly reflective surface near an acoustic source can lead to an increase of the power radiated in the hemisphere above the source, relative to the power that same source would radiate over a full spherical volume in free space. Both monopole and horizontal dipole sources (whose vibration axis is parallel to the reflective surface) can get a large boost in power when they are close to the reflective surface, relative to the wavelength of sound being radiated. (Reproduced from equations in ref. 41). In contrast, vertically oriented dipoles need to be further from the reflective surface to get an equivalent gain in efficiency. B. In fact, near the ground, the sound field from a dipole source is expected to become less directional and we observe this in our models. Data shown here are from a radiator of surface area of 126 mm2 and at frequency 5 kHz, which is seen to be directional 1 m above the ground, but much less so when right next to the ground. C-E. Different animals, especially small animals who have low sound radiation efficiency whether they are monopole or dipole sources may call from near acoustically hard and reflective surfaces, like C. water (photo: Doug Mills), D. tree trunks (photo: Natasha Mhatre), or even the E. ground (photo: Brandon Woo) may be exploiting this effect. Animals that are monopole-like sources are likely to orient themselves parallel to the surface since this is advantageous for camouflage as well. However, crickets which are dipole-like acoustic sources, are constrained to expose themselves by calling with their wings perpendicular to the ground surface to maintain the advantageous horizontal dipole orientation.
Sound radiation efficiency in animal acoustic communication
In the field of animal acoustic communication, calling from the near the ground has been thought of as severely disadvantageous, whether in insects, birds or even primates (21, 22, 41–46). One reason for this is that reflections from the ground degrade temporal cues in calls and songs (25). However, most animal calls are repetitive and redundant, since they are used for functions such as sexual or territorial signaling, or as contact or alarm calls. Here, fine temporal structure is not crucial and the primary functional factor is how loud the sound is (22). How then is loudness affected by the presence of a reflective surface such as the ground? In animal acoustics, it is widely believed that the ground generates high amounts of excess attenuation compared to simple spherical spreading of sound (21, 22, 41–46). Yet, paradoxically, many animals call from the ground, tree trunks, or other vertical surfaces such as cave walls in many ecological contexts (22).
Previous acoustic analyses have typically measured excess attenuation as observed at two distances from the source. Thus, these analyses only account for propagation losses and fail to consider the first step in the process, sound radiation efficiency, i.e., the relationship between the vibration amplitude of the radiator and the level of the sound field that is generated. Here we show that for dipole singers like crickets, calling from the ground can actually provide a substantial boost in sound radiation efficiency, outweighing propagation losses at distances and receiver positions relevant to cricket behavior. It is only when we take this first step in the process of sound generation into account, that it becomes clear that calling from the ground may be an advantage rather than an impediment.
In fact, this phenomenon may be much more general than currently appreciated; analytical findings from as far back as 1957 show that the acoustic power radiated by monopoles and horizontal dipoles (such as cricket wings) increases as they get closer to a perfectly reflecting surface, by as much as a factor of two (Fig 4) (47). When acoustic sources are close to the ground, the reflected sound field effectively sums with the direct sound field, and at very close distances, this summation is near perfect. In particular, the radiated field from dipoles becomes almost monopole like when the source is close to a reflective ground (Fig 4) (48).
Of course, real grounds are not perfectly reflective and do have a finite impedance; they absorb some sound energy (30, 49). However, this does not significantly alter the theoretical expectation of increased sound radiation, except that an additional ground wave is formed in addition to the reflected wave (30). Nonetheless, dipole and monopole sources near the ground are still theoretically expected to have higher sound radiation efficiencies compared to the free or direct field condition (30, 50). In our analyses, we considered dipole sound radiators of finite size, above realistically parametrized grounds. We found sound radiation efficiency increases considerably near the ground. In fact, in some cases efficiency increases by even more than a factor of 2, likely through the baffling effect offered by the ground against acoustic short circuiting.
These findings extend beyond crickets, as they are true for both horizontal dipoles and monopoles (Fig. 4). All acoustically communicating animals are considered to be either dipole or monopole-like sound sources. Among non-cricket invertebrates, both fruit flies and mosquitoes produce sounds with their wings, functioning as dipoles (9). Other invertebrates that use tymbal-based sound production like cicadas and wax moths behave like monopoles (9). Among vertebrates, the sound fields produced by bats are well captured by a baffled-dipole piston model (52, 53) and whales have a similarly directional sound field (54). Most acoustically active vertebrates such as fish, frogs, reptiles, birds and mammals are considered to be monopole sound sources (21, 51). Thus all these animals can exploit this mechanism for gaining efficiency (9, 21, 51). In effect, by shifting the focus from sound propagation to sound radiation efficiency, we are able to posit an explanation for both the paradoxical rarity of tool use and paradoxically high number of animals that call from the ground. Indeed, our findings, and a reconsideration of established acoustics theory, leads us to the exciting discovery of a hitherto unknown mechanism for increasing calling efficiency available to all acoustically communicating animals.
Materials and Methods
Specimen Data
We collected data on wing surface area and call frequency for each of 111 cricket species distributed across the 7 clades described by Chintauan-Marquier et al. (28) (Fig 1, SI 1). We restrict our analysis to this group, since these species are known to raise their wings when singing (55). This behavior means that they are dipole sources of sound, and acutely affected by acoustic short-circuiting (5, 9). Each species was assigned to a clade based on the following two criteria: (1) the species itself was included in the Chintauan-Marquier et al (2016) dataset or (2) the subfamily of the species was included in the dataset and it was found to be monophyletic within one of the seven Chintauan-Marquier clades. Data were obtained from a variety of databases including Orthoptera Species File (56), Crickets North of Mexico, and numerous publications (all references are available in SI 11-12). For a few species of Oecanthines, wings were provided by Nancy Collins and photographed in the lab under a dissecting microscope. All specimens measured were adult males, identified by wing morphology and lack of ovipositor. We measured surface area of the entire left forewing including the lateral field. Fitting an ellipse to the wing, we calculated aspect ratio (length of ellipse/width of ellipse). All image data were gathered using ImageJ version 1.53 (57). We next calculated the fundamental frequency of cricket advertisement calls using Raven Lite version 2.0 (Cornell Lab of Ornithology, 2020). When multiple specimens of a single species were analyzed, averages were used for wing size and call variables. To better represent the full range of wing size and call frequency in our dataset, we included some specimens in the histograms showing wing size and frequency (Fig 1D, E) for which we had only one type of data. Twelve animals had only wing size, but not call data, and 57 animals had call, but not wing size data (see SI 11-12 for details).
Finite Element Models for Sound Radiation Efficiency
We first estimated the sound radiation efficiency of crickets calling in open air using finite element (FE) analysis (SI 2). We built this model and all subsequent models in COMSOL Multiphysics version 5.5. All models used the pressure acoustics module and were solved in the frequency domain assuming a steady state. The Helmholtz equation was the governing equation.
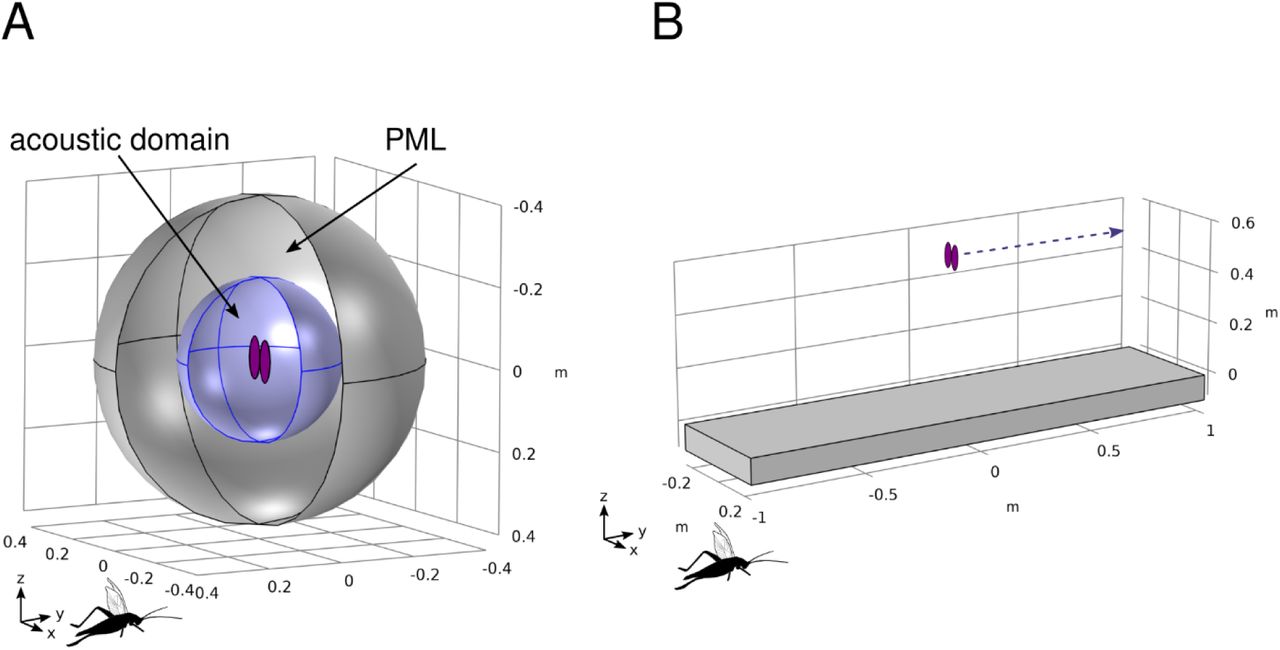
Model geometry, boundary conditions, symmetry, and vibration
Animals were represented by two ellipses which modelled the forewings sitting next to each other along the long axis, in the same plane (SI 2). These ellipses should accurately approximate total radiated sound power and the true shape of the sound field, and it is the size and shape of the radiator and frequency of the call that determines radiation efficiency, and not the radiator’s material properties (8). Surrounding the wings was a spherical acoustic domain consisting of air with a 20 cm radius. The acoustic domain was suspended inside a second spherical domain of 40 cm radius (SI 2. A perfectly-matched layer (PML) was applied to the area between the acoustic domain and larger outer sphere. PMLs in finite element modelling are used to mimic an open and non-reflecting infinite acoustic domain, which absorbs all sound energy from the acoustic domain (59). Thus PMLs mitigate modelling artefacts such as the effects of sound reflecting from the edges of the acoustic domain (7). To reduce computational time, our model was constructed using ¼ symmetry. That is, we modeled ½ of one wing and ¼ of the two spherical domains. We then mirrored this model twice, once about the short axis of the wing to create a whole wing and ½ of the spherical domains, and a second time about a line between and parallel to the long axis of the wings to create a second wing and the whole spherical domains. We applied a time- and space-averaged velocity normal to the entire surface of the wings at 0.13m/s. This was the value measured from the wings of singing Oecanthus henryi and is the only known estimate for crickets (7). However, given that we are calculating efficiency rather than reporting actual sound pressure levels, normalizing using this known value allows comparison between species. We vibrated the wings at a frequency range of 0.5 - 32 kHz, in increments of 0.25 kHz. SREv was subsequently calculated from each of these model outputs as a volumetric average of the absolute pressure in the acoustic domain, divided by the time- and space-averaged velocity of 0.13 m/s, resulting in units of Pa/m/s.
Finite Elements
3D tetrahedral elements were used in both the acoustic domain and PML. After undertaking a mesh size sensitivity study (SI 5), we chose the “extra fine” mesh setting in COMSOL, with about 60000 elements in the acoustic domain. This number did vary somewhat with wing size, as fewer elements are used with very small wings.
Model Parameters
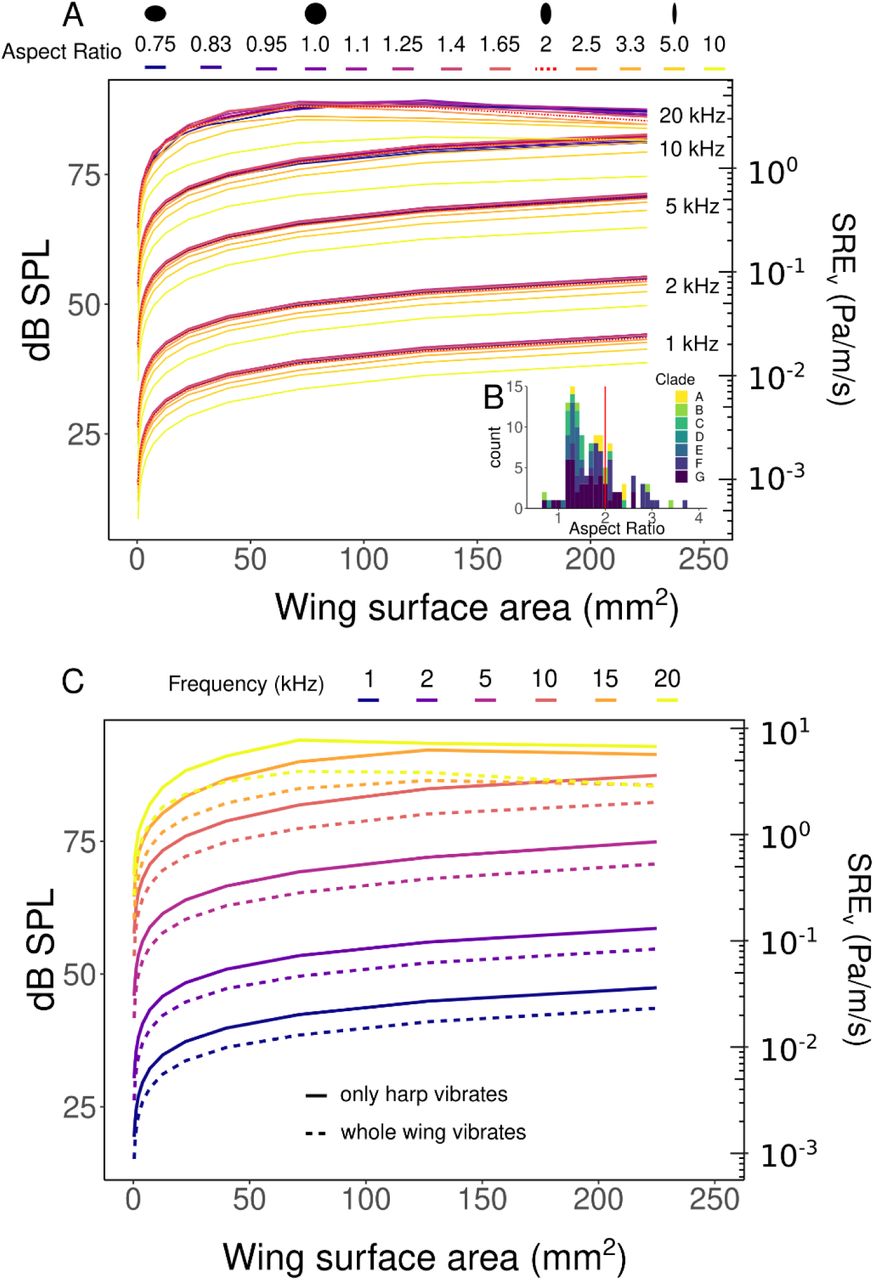
We ran the finite element model at a range of wing surface areas from 0.4 – 4000 mm2, scaled logarithmically by the equation 4 × 10x, where x ranges from -1 to 3). We used an aspect ratio of 2 (wings are twice as long as they were wide). Our chosen aspect ratio of 2 was well within the range of most cricket species (median: 1.6, range: 0.7 - 3.7). Aspect ratio did not play a large role in sound production, except at aspect ratios > 5 (length of wing 5x the width), which were not observed in real wings (SI 4). For aspect ratios within the range of crickets, differences in SREv between aspect ratios at a given wing area and frequency never exceeded 3 dB.
Other Modeling Considerations
The cricket body was not included in our models as it was found to have negligible effects on SREv at all wing sizes and frequencies (mean difference: 0.05 ± 0.01 dB). We also evaluated whether applying vibration to only a part of the wing (a “harp”) influenced sound production. Some cricket species (though not all) use this sound production method (60). We found only minor increases in SREv between vibrating only a harp or vibrating the entire wing (mean: 4 ± 0.08 dB), except at wing sizes well outside the range of the real wings that we measured (SI 4).
Boundary Element Models for sound propagation efficiency
To test hypotheses about how cricket calls interact with objects in the environment, we needed to include an additional domain in the model: a “ground” with realistic parameterized acoustic impedance. To make this model as realistic as possible and to minimize boundary effects, we needed to make the ground element as large as possible relative to the size of the wings. The combination of the large size of ground and the high sound frequencies of interest resulted in finite element models that were too computationally intensive to run. We therefore turned to boundary element modeling as an alternative means of assessing sound propagation efficiency.
Both acoustic boundary element models and finite element models are numerical methods for solving the Helmholtz equation to capture a developing sound field within a medium. However, they differ in how they discretize space within the model. Finite element models discretize volumes by partitioning into a 3-dimensional mesh of finite elements. This allows detailed descriptions of the medium in which the field develops. Boundary element models on the other hand reduce computational cost by discretizing only the boundaries of the acoustic domain and assume a linear homogenous medium in all other spaces. The boundary element formulation therefore trades off some specificity in exchange for computational efficiency, allowing us to make relatively large, more biologically relevant models to assess sound propagation in a spatially explicit manner.
We ran our boundary element models using the pressure acoustics, boundary elements module in COMSOL. All models were run in the frequency domain and assumed steady-state behavior. The Helmholtz equation does not take attenuation due to damping into account, which can become an issue at distances far from the source. However, at frequencies >500 Hz, attenuation due to damping is only about 2 dB per kilometer (26), so we considered this effect to be negligible over the distances of interest for this study (0.2 – 1m).
Model geometry, forcing, and boundary conditions
Wings in the boundary element model were modeled the same way as in the finite element model (SI 2), with no material properties and one-way coupling between the wings and sound fields. Wings were positioned perpendicular to the top surface of the ground, with the flat surfaces of the wings aligned with the short axis of the ground. The wings were centered with respect to ground. The wings were placed above the ground at either 0 m, or 1 m. The same time- and space-averaged velocity was applied as above, and the same set of wing surface areas were used. The ground was modeled as a rectangular slab, 0.5 m wide, 2 m long, and 0.10 m thick. A sound-hard boundary was applied to the bottom surface of the ground slab. Because we were interested in spatially-explicit measures of efficiency as sound propagates across ground, we did not use symmetry conditions to create this model. However, because the sound fields should be symmetric on either side of the wings, we only measured the sound field on one side.
Model Parameters
We used a restricted frequency range for the boundary element models, ranging from of 0.5 – 10 kHz, in increments of 0.25 kHz. We chose 10 kHz as the cutoff because very few of our measured animals call above this frequency, and those that do were Eneopterine species who were likely using a vibrational mode inconsistent with the piston mode that we have implemented (29). High frequency (> 10 kHz) callers were included in the finite element models to give a general sense of where they might fit in with the other animals, but in reality no animals occupy this space in the landscape and all analyses explicitly comparing species exclude them.
To model how a real ground interacts with sound, we applied an acoustic impedance to our modeled ground. Acoustic impedance quantitatively describes how much sound energy is dissipated by the ground, compared to the energy reflected. We used the Attenborough slit-pore model to implement ground impedance. This model uses three parameters to capture both dissipative and reflective properties: flow resistivity, pore density, and porous layer depth. We modeled two different types of ground, a “soft” ground (flow resistivity: 2000 kPa×s/m2, porosity: 0.6) which is less reflective and a “hard” ground (flow resistivity: 9 kPa×s/m2, porosity: 0.4), which is more dissipative. Porous layer depth was held constant for both treatments, at 0.04 m. Both ground parameters were taken from empirical measurements of a “soft” freshly-tilled field and a “hard” forest floor (49).
Sound propagation efficiency definition
In the finite element models, we calculated a volumetric average of absolute pressure within the acoustic domain. However, such a measure would not be appropriate to assess propagation efficiency, as the sound waves’ interactions with the ground would accumulate as distance from the source increases. Therefore, we calculated SREt in a spatially-explicit manner. We measured absolute pressure at 50 points along a 1m long line parallel to the long axis of the ground, at the same height as the wings. The line originated at the center between the two elliptical ‘wings’. We divided this line into “near” and “far” from the caller: near = 0.05 – 0.2 m from wings, far = 0.8 – 0.9 m from wings. Efficiency was calculated as before, by dividing sound pressure level (Pa) by 0.13 m/s, the space and time averaged velocity applied to the wings. We also created an additional boundary element model with no ground, to allow for direct comparisons between ground and no ground and to sanity check the boundary element method compared with the previous finite element models.
Finite Elements
Tetrahedral elements were used on the surface of the ground and 2D triangular elements on the wings. After performing a similar sensitivity study as with the finite element models, we decided on a maximum element size for the wing surfaces of 0.5 cm and 1 cm for the surface of the ground. Since the sound wave is not explicitly modelled, this element size is not related to sound frequency, and instead captures boundary conditions and hence can be larger than in the finite element models.
Excess attenuation due to vegetation
To calculate the effect of vegetation, we used existing models to calculate how standing vegetation is expected to impact call efficiency. We then subtracted this excess attenuation from the COMSOL result. We calculated excess attenuation using the following empirically derived equation from Bashir et al 2015:
 Where EA(Pa) represents excess attenuation due to foliage, F = foliage area per unit volume, L = path length, k = wavenumber, and a = average leaf size. We used values empirically derived for dense foliage with relatively large leaves (31), such as those used to construct baffles in known baffle-using species. To approximate the foliage area and leaf size that a typical baffle user would prefer, we used F = 6.3m-1 and a = 0.0784m in our measures of excess attenuation.
Where EA(Pa) represents excess attenuation due to foliage, F = foliage area per unit volume, L = path length, k = wavenumber, and a = average leaf size. We used values empirically derived for dense foliage with relatively large leaves (31), such as those used to construct baffles in known baffle-using species. To approximate the foliage area and leaf size that a typical baffle user would prefer, we used F = 6.3m-1 and a = 0.0784m in our measures of excess attenuation.
Efficiency of individual species and how much they gain with baffle
To test the lack of utility hypothesis, we used the output of our models to estimate the gains in efficiency that each species could attain if it used a baffle. We did this for both the idealized measure of efficiency (SREv) and the more realistic scenario involving a ground and vegetation (SREt). For each modeling scenario, we first estimated the efficiency of each cricket species in our dataset, given their wing area and call frequency. Next, we calculated the efficiency that each species would have if it used an ideal baffle. To do this, we first determined the size at which the wings and baffle working together as a single radiator would reach maximal efficiency. The optimal size is a function of the wavelength of that sound frequency in that medium. We calculated the quantity ka for each surface area-frequency combination in the model, where k is the wavenumber and a is the effective radius of the sound radiating plates (8). ka is a dimensionless quantity often used in acoustics, as it helps define when a radiator of a particular size transitions from being inefficient sound radiator at low frequencies to an efficient high frequency radiator. For instance, an optimally sized circular piston has ka = 1 (8).
However, the radiators being considered here are two aligned ellipses which not perfectly circular in shape. Hence the particular value of maximal efficiency ka will be different in this configuration. So, to estimate optimal ka for cricket wings, we plotted ka versus efficiency as measured from our models, with a separate trace for each frequency (SI 6). We then identified the ka at which maximal efficiency was reached for all frequencies. This value represents the size at which highest attainable efficiency is reached, consistent with an optimally baffled condition (8) (SI 6). In our finite element models, we found optimal ka to be about 1.3. For the boundary element models at the far distance, optimal ka was approximately 1.55 (SI 6).
Next, we performed a simple linear regression between frequency and maximal efficiency at optimal ka, then calculated the slope and y-intercept of this regression (SI 6). We used this equation to calculate optimal baffled efficiency for each species. The relationship between frequency and efficiency differed depending on condition (open ground vs ground + vegetation) (SI 6), so this regression was performed separately for each environmental condition when calculating optimal baffled efficiency for a given condition. It should be noted that due to modelling constraints we were only able to calculate baffled efficiency on the ground for animals with a carrier frequency >3.5 kHz.
Directionality index
To address how difficult it would be for a female to localize a male call, we assessed the directionality of the call in each modeling scenario. In an open field with no ground, the sound level is expected to decrease smoothly following the inverse square law (61, 62). A cricket moving toward the source of a call should therefore always experience either an increase in loudness, or, if the increase is below the animal’s difference threshold, no change in loudness. A cricket should always move in the direction of increasing SPL to locate the singing male and therefore a mating opportunity. However, in reality, sound fields become more complicated when they interact with the ground, resulting in a noisy relationship between SPL and distance (63). In such sound fields, female phonotaxis may fail as there is no clear acoustic gradient to follow to the source. To quantify this degree of potential “confusion”, we calculated a directionality index for each modeling scenario. First, we calculated Δ SPL between each two adjacent points 2 cm apart (∼ 2 body lengths for most animals in this analysis). Δ SPL was calculated starting at 1 m away and moving toward the source. Next, we classified each of these values as either consistent with expected change in SPL or inconsistent. Consistent values represented either an increase, no change, or a decrease smaller than Δ 3dB SPL (a factor of about 1.4), which is thought to be close to the detectable threshold for crickets (64). See Römer, 2021 for a more complex treatment of such thresholds. For our purposes, inconsistent values represented a decrease in SPL greater than 3 dB. For each modeling scenario, we calculated the proportion of Δ SPLs classified as consistent. This resulting value we call “Directionality” ranging from 0 to 1 (SI 10). We calculated directionality for two different distance treatments, “near” was calculated from 0.05 – 0.2 m from the wings, and “far” was calculated from 0.5 – 1m from the wings.
Supplementary Material

Measuring acoustic-morphospace of crickets. Wing area and call frequency were quantified. A. Wing surface area was calculated as the area of the entire forewing. B. The carrier frequency (sometimes called fundamental frequency) of the call was identified from spectrograms and recorded. If the fundamental frequency occupied a sweep over a range of frequencies (as shown), an average was taken. C. Specimen sampling scheme. Distribution of all species described across each of the seven clades in Grylloidea (according to Orthoptera Species File, as of February 16, 2022). D. Distribution of species that were sampled for this study.

Geometry of biophysical models. A. Geometry of finite element (FE) models. Inner blue sphere is the acoustic domain. The wings are represented by purple ellipses in the center. Outer sphere is a perfectly-matched layer, to mitigate boundary effects caused by the finite acoustic domain. SPL is averaged over the inner sphere for the calculation of SREv. Wings vibrate along the y axis, as indicated by the cricket silhouette. B. Geometry of boundary element (BE) models. Grey shape represents a ground of defined acoustic impedance. Purple ellipses represent wings. Dotted line illustrates the line (transect) along which measurements were taken to assess SREt.

Sound fields produced by modeled wings vary with respect to call frequency and wing size. Fields are oriented such that wings are perpendicular to page and vibrate left to right, as indicated by the silhouette cricket. Sound fields are given for the following combinations of wing size and frequency: A. wing size = 0.04 mm2, frequency = 10 kHz; B. wing size = 225 mm2, frequency = 10 kHz; C. wing size = 0.4 mm2, frequency = 1.5 kHz; D. wing size = 225 mm2, frequency = 1.5 kHz. Spatial scale given in C applies to all sound fields. Efficiency (here, size and color of sound field) increases with improved match between wavelength of sound and size of radiator (wing). Cricket wings in general are small so this match is poor except at the extreme high end of radiator size and call frequency (B).

Wing aspect ratio and use of a “harp” resonator do not significantly impact SREv within biologically-relevant ranges of wing size and call frequency. A. The effect of wing aspect ratio on SREv at six different call frequencies. The aspect ratio that was used on all models in this study (2) is shown by red dotted line. B. Actual distribution of aspect ratios among species. Red line indicates aspect ratio that was used in our models (2). We see that while aspect ratio influences SREv, this effect is minor within the realistic range of aspect ratios (typically <3 dB for ARs from 1 to 3.3). B. The effect on SREv of vibration spread over a small area (harp) compared with the whole wing. Some species of crickets restrict the vibrating portion of the wing to a “harp” region. However, we find that this does not strongly affect SRE at any frequency within our range of interest.

Mesh sensitivity analysis for models based on the finite element method. Each line represents the SPL of a single wing size with a different number of mesh elements. Each panel shows this analysis at a different frequency: A. 30 kHz, B. 20 kHz, C. 10 kHz, D. 5 kHz. As the difference between the second-largest and largest number of elements was small, we proceeded with the largest number of elements shown here for the analysis.

Calculating efficiency with optimal baffle. A. ka at which optimal efficiency occurs for each frequency. Each frequency is represented by a different line. Line at which efficiency is maximized for each frequency (maximum efficiency ridge) is shown by red dashed line. B. Efficiency at this optimal ka (1.55) in open and vegetation conditions.

Acoustic hardness of ground does not significantly influence SREt at biologically relevant ranges of wing size and call frequency. Comparison of SREt over hard vs soft ground. Each panel represents a combination of caller height above ground (0 m or 1 m) and receiver distance from caller (0.05 – 02 m “near” and 0.8 – 0.9 m “far”). A. Distance = near, height = 1m, B. Distance = far, height = 1 m; C. Distance = near, height = 1 m, D. Distance = far, height = 0 m. Color indicates whether higher SRE is found with hard ground (red shades), soft ground (blue shades) or no difference (white). Data are presented as a log2 ratio instead of a straight proportion. Log2 ratios are scaled such that the ranges above and below 1 are proportional, rather than values below 1 being compressed between 0 and 1. Each clade of animals is represented by a colored ellipse. E. Phylogeny representing each clade F. Key to clade represented by each ellipse.

Vegetation decreases efficiency overall but does not substantially change the landscape pattern of efficiency. Each panel represents a combination of caller height above ground (0 m or 1 m) and receiver distance from caller (0.05 – 02 m “near” and 0.8 – 0.9 m “far”). In each height and distance scenario, an excess attenuation factor due to vegetation was also applied. A. Distance = near, height = 1m, B. Distance = far, height = 1 m; C. Distance = near, height = 1 m, D. Distance = far, height = 0 m. Red lines indicate optimal efficiency ridge, or the size at each frequency that would produce an ideally baffled calling scenario. Each clade of animals is represented by a colored ellipse. E. Phylogeny representing each clade F. Key to clade represented by each ellipse.

Vegetation somewhat decreases efficiency in baffled and grounded calling conditions. Differences in SREt by clade depending on vegetation and baffle use. For each clade, bars with black outline represent SREt without baffle and bars with red outline represent SREt with ideal baffle. Background color of the bars indicate vegetation or no vegetation.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Call directionality decreases at higher frequencies, particularly with grounded calling. Each panel represents a combination of caller height above ground (0 m or 1 m) and receiver distance from caller (0.05 – 02 m “near” and 0.8 – 0.9 m “far”). A. Distance = near, height = 1m, B. Distance = far, height = 1 m; C. Distance = near, height = 1 m, D. Distance = far, height = 0 m. Each clade of animals is represented by a colored ellipse. E. Phylogeny representing each clade F. Key to clade represented by each ellipse.
Sources for morphological data. Asterisk after species name indicates species is represented in both frequency and wing size datasets
Sources for call frequency data. Asterisk after species name indicates species is represented in both frequency and wing size datasets
Full citations for Specimen data:
Anso J, Barrabé L, Desutter-Grandcolas L, Jourdan H, Grandcolas P, Dong J, Robillard T. 2016. Old Lineage on an Old Island: Pixibinthus, a New Cricket Genus Endemic to New Caledonia Shed Light on Gryllid Diversification in a Hotspot of Biodiversity. PLOS ONE 11:e0150920. doi:10.1371/journal.pone.0150920
Barranco P, Gilgado JD, Ortuño VM. 2013. A new mute species of the genus Nemobius Serville (Orthoptera, Gryllidae, Nemobiinae) discovered in colluvial, stony debris in the Iberian Peninsula: A biological, phenological and biometric study. Zootaxa 3691:201–219. doi:10.11646/zootaxa.3691.2.1
Castro-Souza RA, Zefa E, Ferreira RL. 2020. New troglobitic and troglophilic syntopic species of Endecous (Orthoptera, Grylloidea, Phalangopsidae) from a Brazilian cave: a case of sympatric speciation? Zootaxa 4810:271–304. doi:10.11646/zootaxa.4810.2.3
Cole JA, Funk DH. 2019. Anaxipha hyalicetra sp. n. (Gryllidae: Trigonidiinae), a new sword-tailed cricket species from Arizona. Journal of Orthoptera Research 28:3–9. Dambach M, Gras A. 1995. Bioacoustics of a miniature cricket, Cycloptiloides canariensis (Orthoptera: Gryllidae: Mogoplistinae). Journal of Experimental Biology 198:721–728. doi:10.1242/jeb.198.3.721
Desutter-Grandcolas L. 2009. New and little known crickets from Espiritu Santo Island, Vanuatu (Insecta, Orthoptera, Grylloidea, Pseudotrigonidium Chopard, 1915, Phaloriinae and Nemobiinae p.p.). zoos 31:619–659. doi:10.5252/z2009n3a12
Desutter-Grandcolas L, Robillard T. 2006. Phylogenetic systematics and evolution of Agnotecous in New Caledonia (Orthoptera: Grylloidea, Eneopteridae). Systematic Entomology 31:65–92. doi:10.1111/j.1365-3113.2005.00299.x
Gorochov AV, Tan MK. 2012. New crickets of the subfamilies Phaloriinae and Pteroplistinae (Orthoptera: Gryllidae) from Singapore. Zootaxa 3525:18–34. doi:10.11646/zootaxa.3525.1.2
He Z. 2012. A new species of Truljalia Gorochov 1985 from Taiwan (Orthoptera: Gryllidae; Podoscirtinae; Podoscirtini). Zootaxa 3591:79–83. doi:10.11646/zootaxa.3591.1.5
He Z, Lu H, Liu Y, Li K. 2017. A new species of Ornebius Guérin-Méneville, 1844 from East China (Orthoptera: Mogoplistidae: Mogoplistinae). Zootaxa 4303:445–450. doi:10.11646/zootaxa.4303.3.10
Hugel S. 2009. New Landrevinae from Mascarene islands and little known Landrevinae from Africa and Comoros (Grylloidea: Landrevinae). Annales de la Société entomologique de France (NS) 45:193–215. doi:10.1080/00379271.2009.10697602
Jaiswara R, Dong J, Ma L, Yin H, Robillard T. 2019. Taxonomic revision of the genus Xenogryllus Bolívar, 1890 (Orthoptera, Gryllidae, Eneopterinae, Xenogryllini). Zootaxa 4545:301–338. doi:10.11646/zootaxa.4545.3.1
Kavanagh MW, Young D. 1989. Bilateral symmetry of sound production in the mole cricket, Gryllotalpa australis. J Comp Physiol A 166:43–49. doi:10.1007/BF00190208
Kim T. 2011. A Taxonomic Study of the Scaly Cricket Family Mogoplistidae (Orthoptera: Ensifera: Grylloidea) in Korea. Zootaxa 2928:41–48. doi:10.11646/zootaxa.2928.1.4
Lima RM, Schuchmann K-L, Tissiani AS, Nunes LA, Jahn O, Ganchev TD, Lhano MG, Marques MI. 2018. Tegmina-size variation in a Neotropical cricket with implications on spectral song properties. Journal of Natural History 52:1225–1241. doi:10.1080/00222933.2018.1457728
Love RE, Walker TJ. 1979. Systematics and Acoustic Behavior of Scaly Crickets (Orthoptera: Gryllidae: Mogoplistinae) of Eastern United States. Transactions of the American Entomological Society (1890-) 105:1–66.
Martins L de P, Redü DR, Oliveira GL de, Zefa E. 2012. Recognition characters and new records of two species of Phylloscyrtini (Orthoptera, Gryllidae, Trigonidiinae) from southern Brazil. Iheringia, Sér Zool 102:95–98. doi:10.1590/S0073-47212012000100013
Martins LDP, Silva LGD, Henriques AL, Zefa E. 2013. First record of the genera Luzarida Hebard, 1928 and Luzaridella Desutter-Grandcolas, 1992 (Orthoptera, Gryllidae, Phalangopsinae) from Brazil, including a new species and description of the female of Luzarida lata Gorochov, 2011. Zootaxa 3609:421–430. doi:10.11646/zootaxa.3609.4.4
McIntyre ME. 1977. Acoustical communication in the field crickets Pteronemobius nigrovus and P. bigelowi (Orthoptera: Gryllidae). New Zealand Journal of Zoology 4:63–72. doi:10.1080/03014223.1977.9517937
Mello F de AG de, Pellegatti-Franco F. 1998. A New Cave Cricket of the Genus Endecous from Southeastern Brazil and Characterization of Male and Female Genitalia of E. itatibensis Rehn, 1918 (Orthoptera: Grylloidea: Phalangopsidae: Luzarinae). Journal of Orthoptera Research 185–188. doi:10.2307/3503517
Mello F de AG de, Reis JC dos. 1994. Substrate Drumming and Wing Stridulation Performed during Courtship by a New Brazilian Cricket (Orthoptera: Grylloidea: Phalangopsidae). Journal of Orthoptera Research 21–24. doi:10.2307/3503603
Milach EM, Costa MKMD, Martins LDP, Nunes LA, Silva DSM, Garcia FRM, Oliveira ECD, Zefa E. 2016. New species of tree cricket Oecanthus Serville, 1831 (Orthoptera: Gryllidae: Oecanthinae) from Reserva Natural Vale, Espírito Santo, Brazil, with chromosome complement. Zootaxa 4173:137–146. doi:10.11646/zootaxa.4173.2.4
Milach EM, Martins LDP, Costa MKMD, Gottschalk MS, Oliveira GLD, Redü DR, Neutzling AS, Dornelles JEF, Vasconcellos LA, Zefa E. 2015. A new species of tree crickets Oecanthus (Orthoptera, Gryllidae, Oecanthinae) in tobacco plantation from Southern Brazil, with body color variation. Zootaxa 4018:266–278. doi:10.11646/zootaxa.4018.2.6
Nischk F, Otte D. 2000. Bioacoustics, Ecology and Systematics of Ecuadorian Rainforest Crickets (Orthoptera: Gryllidae: Phalangopsinae), with a Description of Four New Genera and Ten New Species. Journal of Orthoptera Research 229–254. doi:10.2307/3503651
Orsini MP, Costa MKMD, Szinwelski N, Martins LDP, Corrêa RC, Timm VF, Zefa E. 2017. A new species of Miogryllus Saussure, 1877 and new record of Miogryllus piracicabensis Piza, 1960 (Orthoptera: Gryllidae) from State of Rio Grande do Sul, Brazil, with calling song and chromosome complement. Zootaxa 4291:361– 372. doi:10.11646/zootaxa.4291.2.8
Otte D. 2007. Australian Crickets (Orthoptera: Gryllidae). Academy of Natural Sciences.
Pereira MR, Miyoshi AR, Martins LDP, Fernandes ML, Sperber CF, Mesa A. 2013. New Neotropical species of Hygronemobius Hebard, 1913 (Orthoptera: Grylloidea: Nemobiinae), including a brief discussion of male genitalia morphology and preliminary biogeographic considerations of the genus. Zootaxa 3641:1–20. doi:10.11646/zootaxa.3641.1.1
Robillard T, Desutter-Grandcolas L. 2004. High-frequency calling in Eneopterinae crickets (Orthoptera, Grylloidea, Eneopteridae): adaptive radiation revealed by phylogenetic analysis. Biological Journal of the Linnean Society 83:577–584. doi:10.1111/j.1095-8312.2004.00417.x
Robillard T, Ichikawa A. 2009. Redescription of Two Cardiodactylus Species (Orthoptera, Grylloidea, Eneopterinae): The Supposedly Well-Known C. novaeguineae (Haan, 1842), and the Semi-Forgotten C. guttulus (Matsumura, 1913) from Japan. jzoo 26:878–891. doi:10.2108/zsj.26.878
Robillard T, Nattier R, Desutter-Grandcolas L. 2010. New species of the New Caledonian endemic genus Agnotecous (Orthoptera, Grylloidea, Eneopterinae, Lebinthini). Zootaxa 2559:17–35. doi:10.11646/zootaxa.2559.1.2
Robillard T, Su YN. 2018. New lineages of Lebinthini from Australia (Orthoptera: Gryllidae: Eneopterinae). Zootaxa 4392:241–266. doi:10.11646/zootaxa.4392.2.2 Robillard T, Tan MK. 2013. A taxonomic review of common but little knwon crickets from Singapore and the Philippines (Insecta: Orthoptera: Eneoptera: Eneopterinae) 21.
Robillard T, Yap S, Yngente MV. 2013. Systematics of cryptic species of Lebinthus crickets in Mount Makiling (Grylloidea, Eneopterinae). Zootaxa 3693:49–63. doi:10.11646/zootaxa.3693.1.3
Souza-Dias PGB, Szinwelski N, Fianco M, Oliveira ECD, Mello FD a. GD, Zefa E. 2017. New species of Endecous (Grylloidea, Phalangopsidae, Luzarinae) from the Iguaçu National Park (Brazil), including bioacoustics, cytogenetic and distribution data. Zootaxa 4237:454–470. doi:10.11646/zootaxa.4237.3.2
Su YN, Rentz DCF. 2000. Australian Nemobiine Crickets: Behavioral Observations and New Species of Bobilla Otte & Alexander (Orthoptera: Gryllidae: Nemobiinae). Journal of Orthoptera Research 5–20. doi:10.2307/3503626
Symes LB, Collins N. 2013. Oecanthus Texensis: A New Species of Tree Cricket from the Western United States. orth 22:87–91. doi:10.1665/034.022.0203
Tan MK, Japir R, Chung AYC, Wahab RBHA. 2020. New taxa of crickets (Orthoptera: Grylloidea: Phaloriinae, Phalangopsinae, Itarinae and Podoscirtinae) from Borneo (Brunei Darussalam and Sandakan). Zootaxa 4810:244–270. doi:10.11646/zootaxa.4810.2.2
Tan MK, Kamaruddin KN. 2016. A new species of Gryllotalpa mole cricket (Orthoptera: Gryllotalpidae: (Gryllotalpinae) from Peninsular Malaysia. Zootaxa 4066:552– 560. doi:10.11646/zootaxa.4066.5.3
Vicente NM, Olivero P, Lafond A, Dong J, Robillard T. 2015. Gnominthus gen. nov., a new genus of crickets endemic to Papua New Guinea with novel acoustic and behavioral diversity (Insecta, Orthoptera, Gryllidae, Eneopterinae). Zoologischer Anzeiger - A Journal of Comparative Zoology 258:82–91. doi:10.1016/j.jcz.2015.06.005
Weissman DB, Gray DA. 2019. Crickets of the genus Gryllus in the United States (Orthoptera: Gryllidae: Gryllinae). Zootaxa 4705:1–277. doi:10.11646/zootaxa.4705.1.1
Zefa E, Acosta RC, Timm VF, Szinwelski N, Marinho MAT, Costa MKMD. 2018. The Tree Cricket Neoxabea brevipes Rehn, 1913 (Orthoptera: Gryllidae: Oecanthinae) from the Brazilian southern Atlantic Forest: morphology, bioacoustics and cytogenetics. Zootaxa 4531:554–566. doi:10.11646/zootaxa.4531.4.6
Zefa E, Neutzling AS, Redü DR, Oliveira GLD, Martins LDP. 2012. A new species of Oecanthus and Oecanthus lineolatus Saussure, 1897 from Southern Brazil: species description, including phallic sclerites, metanotal glands and calling song (Orthoptera: Gryllidae: Oecanthinae). Zootaxa 3360:53–67. doi:10.11646/zootaxa.3360.1.2
Acknowledgments
We wish to acknowledge a number of undergraduate students who assisted with the collection and databasing of cricket acoustic and morphometric data: Morteza Al Rabya, Nancy Kim, Shanker (Matthew) Nadarajah, and Daniel Xie. Nancy Collins provided specimens for several of the wing measurements. Graduate students Hossein Asgari and Reese Gartly assisted with multiplexing dozens of model runs on multiple machines. Emine Celiker and Damian Elias provided feedback on an earlier version of this manuscript. This work would have not been possible without an extensive worldwide network of specimen databases. We especially wish to thank those who contribute to, curate, or maintain Orthoptera Species File, Singing Insects of North America, Museum D’Historie Naturelle, and numerous university insect collections. We also wish to thank the photographers who graciously provided their images free of charge for inclusion in this manuscript.
We would also like to thank the following funding sources for their support of this research: NSERC Discovery (Grant no. 687216), and early career supplement (675248), and an NSERC Canada research chair (Grant no. 693206) to NM; the Western University Postdoctoral Fellowship to EEB; Western Undergraduate Work Study to SD and HW; Western Undergrad Summer Research Internship to HW.
Footnotes
Competing Interest Statement: The authors declare no competing interests.
We have added a new analysis making our results more general across acoustically active animals.
References




