

ABSTRACT
Aeromonas dhakensis is the most virulent Aeromonas species in tropical and subtropical areas and causes a variety of human diseases. Owing to its resistance against some antibiotics, there is an urgent need for new strategies against this pathogen. This is the first study to isolate and characterize lytic phages against A. dhakensis. Of Aeromonas isolates, only isolate AM could be used to isolate phages using the enrichment technique. This strain was identified via biochemical tests, 16S rDNA sequencing, and whole-genome analyses. All results confirmed that strain AM was A. dhakensis. In silico detection of antimicrobial resistance genes and virulence factors corresponding to the main bacterial virulence determinants were observed in A. dhakensis AM, which was then used as the host for phage isolation. Four lytic phages, designated vB_AdhS_TS3, vB_AdhM_TS9, vB_AdhM_DL, and vB_AdhS_M4, were isolated. Transmission electron micrographs showed that vB_AdhS_TS3 and vB_AdhS_M4 belonged to Siphoviridae family, whereas vB_AdhM_TS9 and vB_AdhM_DL belonged to Myoviridae family. Host-range determination demonstrated that all phages were capable of infecting only A. dhakensis. Three phages, vB_AdhS_TS3, vB_AdhM_TS9, and vB_AdhM_DL, were selected since they had a shorter latency period and larger burst sizes. All phages were resistant to a wide range of pH values and remained relatively stable after a 60-minute incubation at 4 °C, 25 °C, 30 °C, and 37 °C but were sensitive to higher temperatures. The pre-treatment (co-inoculation of A. dhakensis and phage) with individual phages and phage cocktails reduced bacterial numbers in the range of 2.82-6.67 and 5.19-5.43 log CFU/mL, respectively, after 6 h of incubation. In post-treatment, the maximum inactivation was achieved with the log reduction of bacterial number in the range of 3.06-5.25 and 4.01-6.49 log CFU/mL after 6 and 12 h of incubation, respectively. A combination of phage cocktail with amoxicillin at sub-MIC showed complete inactivation in pre-treatment and post-treatment in a volume of 200 µL; however, an incomplete inhibition was observed in post-treatment in the volume of 20 mL but still decreased by about 1.2-1.7 log CFU/mL when compared to the control and other individuals used after incubation for 48 h. The complete genome and G+C content of phages vB_AdhS_TS3, vB_AdhM_DL, and vB_AdhM_TS9 were 115,560, 61,429, and 115,503 bp, respectively, with G+C contents of 41.10%, 61.7%, and 35.34%, respectively. This study demonstrated using phages as an adjuvant with a sublethal concentration of antibiotics as an effective therapeutic strategy.
IMPORTANCE Aeromonas dhakensis is widely distributed in the environment and can cause a variety of infections both in human and animals and is often misidentified with other members of the Aeromonas family, A. hydrophila, A. veronii, or A. caviae using the commercial test kits which has led to challenges in treating this pathogen. Hence, our study isolated phages against this bacterial strain and extensively characterized their efficacy with common antibiotics used to treat the pathogen.
INTRODUCTION
Aeromonas species are gram-negative, facultatively anaerobic, rod-shaped chemoorganotrophs with both oxidative and fermentative metabolism and oxidase- and catalase-positive bacteria. Aeromonas spp. are abundant in aquatic environments and are found in food, and animals and lead to various infections in humans (1–3). Among the species, A. hydrophila, A. caviae, and A. veronii were the most prevalent species prior to the proposal of a new species, A. dhakensis. A. dhakensis (synonymized with A. hydrophila subsp. dhakensis (4) and A. aquariorum (5) is often misidentified as A. hydrophila (6, 7) by phenotypic methods (8), and 16S rRNA sequencing is considered unreliable for identifying Aeromonas at the species level (3). This strain has gained increasing interest because it is widely distributed in the environment and can cause infections such as gastroenteritis, wound infection, bacteremia, skin and soft-tissue infections, and respiratory infections (9, 10). Previous studies have shown that A. dhakensis was the most virulent Aeromonas species in tropical and subtropical regions (11). It causes soft tissue infection and bacteremia, especially in individuals with malignancy or liver cirrhosis (12, 13). This species was more prevalent in countries with hot climates, such as Bangladesh (4), Taiwan (12, 14), Australia (15), Philippines (formerly A. hydrophila) (16), Egypt (17), Malaysia (18) and Thailand (19). In Thailand, A. dhakensis (formerly A. aquariorum) was first reported in marine shrimps cultured in low-salinity inland areas and is the second most prevalent species next to A. veronii. A. dhakensis contains many virulence factors, including hemolysins and various extracellular hydrolytic enzymes, which play a significant role in the invasiveness and establishment of infections (20, 21). Clinical strains have been isolated from stool, blood, wounds, and other extra-intestinal samples of humans worldwide (12). Antibiotic treatment is critical for preventing and treating infections; however, resistance to antimicrobial agents, including amoxicillin, cephalothin, and cefoxitin, is rising (8). Moreover, some A. dhakensis can produce biofilms to adhere to different surfaces; thus, treating A. dhakensis infections with common medications becomes further challenging. Therefore, there is a need to develop alternative treatments for A. dhakensis infections other than antimicrobial agents.
Bacteriophages are viruses that kill specific bacteria without disturbing other flora. Bacteriophages are used to control pathogenic bacteria and potential treatments have been reported for humans, animals, crops, and aquaculture (22–24). Many studies have isolated phages against A. hydrophila and have determined their efficacy for protective and therapeutic effects against disease (25–28). However, there have been no reports on the isolation and characterization of lytic phages specific to A. dhakensis. The objective of this study was to isolate and characterize a new lytic phage from water that infects A. dhakensis. This study also investigated the lytic activity of the isolated phage and its combination with antibiotics against A. dhakensis in vitro.
RESULTS AND DISCUSSION
Aeromonas isolation and identification
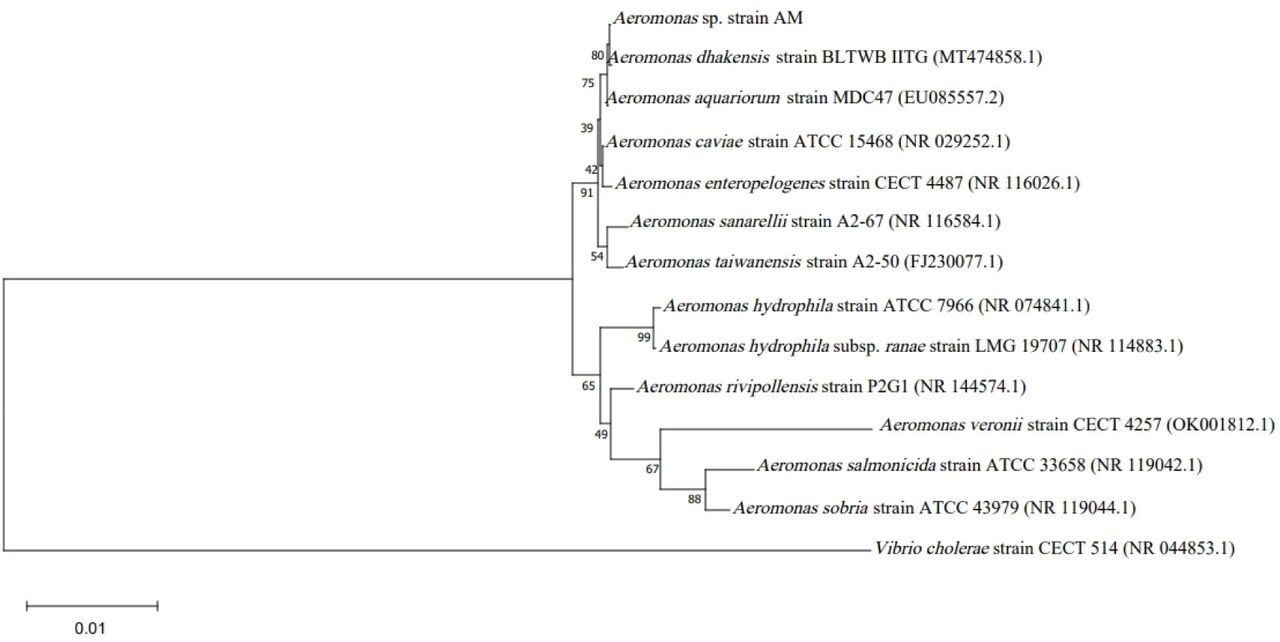
Three of the 40 isolates from 30 collection sites were preliminarily identified as Aeromonas by biochemical tests. These three Aeromonas strains were then used as hosts for phage isolation. However, only the Aeromonas isolate AM was able to isolate phages using the enrichment technique. The isolated AM was further characterized using 16S rRNA gene sequence analysis which revealed gene sequences with a length of 1,546 nucleotides, and overlapped A. dhakensis with 99% identity. The neighbor-joining tree revealed that strain AM was most closely related to A. dhakensis (Fig. 1). The biochemical tests of A. dhakensis AM are shown in Table 1. However, A. dhakensis is often misidentified as A. hydrophila since both species can be isolated from Aeromonas isolation agar supplemented with ampicillin. However, we added two biochemical tests (L-arabinose and salicin fermentation) to differentiate A. dhakensis from A. hydrophila subsp. hydrophila and A. hydrophila subsp. ranae (10). The results showed that strain AM was negative for L-arabinose and positive for salicin fermentation, confirming that this strain is A. dhakensis. However, to identify more characteristics, a whole genome sequence analysis of strain AM was included in this study.

Phylogenetic tree of Aeromonas spp. based on the 16S rRNA gene using neighbor-joining method. Bootstrap values (%) of 1,000 replicates are represented on the branches.
Biochemical tests of A. dhakensis AM
Antimicrobial susceptibility of A. dhakensis AM
The MICs of six antimicrobial agents against A. dhakensis AM were evaluated (Table 2). Amoxicillin had the highest MIC of 24 µg/mL among the antibiotics, much higher than the CLSI MIC breakpoints (>8 µg/mL). As the information about the susceptibility profiles of A. dhakensis is rare, our findings show that A. dhakensis AM is susceptible to chloramphenicol, doxycycline, and gentamicin. From in silico genome analyses, tetracycline resistance genes were identified in the genome of A. dhakensis AM (data not shown); however, the MIC value of tetracycline was intermediate (12 µg/mL) (Table 5). Aeromonads are usually susceptible to 4th-generation cephalosporins, aminoglycosides, fluoroquinolones, tetracycline, and trimethoprim-sulfamethoxazole (15, 50, 51). However, increasing resistance rates to fluoroquinolones, tetracyclines, and trimethoprim-sulfamethoxazole have been reported (52–54). According to Chen et al. (2014) (12), all strains of A. dhakensis (n=37) were susceptible to gentamicin, and 90% of isolates gave the MIC of 0.5 µg/mL, which was similar to our study. A few antimicrobial agents (oxytetracycline, amoxicillin, sulfadimethoxine/ormetoprim, and enrofloxacin) have been approved for use in aquaculture in Thailand (55). The results of this study indicated that the resistance rate to amoxicillin was per Aravena-Roman et al. (2011) (56), who reported that only 1.6% of 193 Aeromonas isolates were susceptible to amoxicillin. Therefore, we used amoxicillin at sub-MIC to evaluate the synergism between the antibiotics and phage cocktail.
Antibiotic susceptibility test
Genomic features of A. dhakensis AM
The in silico genome of A. dhakensis AM comprises one circular chromosome of 4,884,279 bp with a G+C content of 61.9% (Fig. 2, Table 3). The genome contained 4,256 coding DNA sequences (CDSs). We emphasized the antimicrobial resistance genes and virulence factors corresponding to the main bacterial virulence determinants. Antimicrobial resistance genes were identified in the genome of A. dhakensis AM (data not shown). Genes for antibiotic inactivation were identified, including members of the resistance-nodulation-cell division (RND) antibiotic efflux pump, major facilitator superfamily (MFS) antibiotic efflux pump, small multidrug resistance (SMR), AQU beta-lactamase, OXA beta-lactamase, elfamycin resistant EF-Tu, and gene antibiotic target alterations. Virulence factor genes were identified in the genome of A. dhakensis AM (data not shown). Several type II secretion system (T2SS) component genes and type VI secretion system (T6SS) with T6SS effectors, including two hemolysin-coregulated (hcp) and two valine-glycine repeats G (vgrG), have been identified, suggesting that T6SS can play a role in the pathogenicity of this strain. Several typical toxin-encoding genes have been identified, including aerolysin (aerA), extracellular hemolysin (ahh1), hemolysin (hlyA), hemolysin III, repeat in toxin (rtxA, rtxB, rtxC, rtxD, rtxE, and rtxH), thermostable hemolysin, and exotoxin A (toxA). Five prophages were identified in the genome (data not shown), and no plasmids were found during genome analysis.

Circular representation of the A. dhakensis AM genome. The following characteristics are shown from the outside to the center of the diagram. Circle 1: coding sequence (CDS) on the reverse strand, circle 2: coding sequence (CDS) on the forward strand, circle 3: GC contents, circle 5: GC skew values (GC skew+ shown in green, GC skew-shown in pink).
Genome features of A. dhakensis AM
Phage isolation, purification and phage morphology
Four phages, designated as vB_AdhS_TS3, vB_AdhM_TS9, vB_AdhM_DL, and vB_AdhS_M4 were isolated using A. dhakensis AM as the host. The phages were purified using three successive single-plaque isolations. These phages exhibited clear plaques with diameters ranging from 1.7 to 2.0 mm (Fig. 3). Electron micrographs revealed that two isolated phages vB_AdhS_TS3 and vB_AdhS_M4 belonged to the family Siphoviridae, displaying an icosahedral head of approximately 75.2 nm and 64.8 nm, respectively, a contractile tail with the length of 225.3 nm and 185.4 nm, respectively. Phages vB_AdhM_DL and vB_AdhM_TS9 belonged to the family Myoviridae, possessing an icosahedral head with dimensions of 50.4 and 85.1 nm, respectively and a tail length of 210.4 nm and 101.4 nm, respectively (Fig. 3). Bai et al. (2019) (57) reported that from 51 complete genome sequences of Aeromonas phages in GenBank, the majority of Aeromonas phages were double-strand DNA phages belonging to the Myoviridae (33/51), Podoviridae (7/51) and Siphoviridae (5/51) families, along with one ssDNA virus and five unclassified bacteriophages (58–63).

Plaques and TEM images of phage vB_AdhS_TS3 (A), vB_AdhS_M4 (B), vB_AdhM_DL (C), and vB_AdhM_TS9 (D). Scale bar = 100 nm.
Host range determination
All phages were infected only with A. dhakensis and did not infect other Aeromonas spp., such as A. hydrophila, A. caviae, A. sobria, A. trota, or A. veronii (Table 4). Bacteriophage vB_AdhM_DL is the broadest host range phage, able to infect A. dhakensis in five out of the six strains tested. However, for future applications, there is a need to search for additional A. dhakensis hosts. Our results indicate that all phages are species-specific, as reported elsewhere for most Aerononas phages (64).
Host range of four A. dhakensis phages. Clear lysis zone (+++), turbid lysis (++), weak lysis (+) and no lysis zone (−).
Optimal multiplicity of infection determination (MOI) and one-step growth curve
Phage vB_AdhS_TS3, vB_AdhM_DL, vB_AdhS_M4 and vB_AdhM_TS9 generated a maximum titre of 9.68±0.05, 9.94±0.05, 10.41±0.06 and 8.85±0.25 PFU/mL when infected at an optimal MOI of 10 (Table 5). A one-step growth curve of the phages vB_AdhS_TS3, vB_AdhM_DL, vB_AdhS_M4, and vB_AdhM_TS9 was obtained by propagation on A. dhakensis AM as the host at 37 °C (Fig. 4). The latent period of phages vB_AdhS_TS3, vB_AdhM_DL, vB_AdhS_M4, and vB_AdhM_TS9 was approximately 40, 30, 50, and 30 min, respectively, and the burst sizes of phage vB_AdhS_TS3, vB_AdhM_DL, vB_AdhS_M4, and vB_AdhM_TS9 were estimated as 1380, 1280, 253, and 6300 PFUs/infected cells, respectively. Several parameters affect phage therapy, including burst size, latent period, and initial phage dose (65–67). Apart from a wide host range, phages with short latent periods and/or high burst sizes are preferred as biocontrol agents. Among the four phages, phage vB_AdhS_M4 had the longest latent period, smallest burst size, and narrowest host range. Therefore, we selected the other three phages, vB_AdhS_TS3, vB_AdhM_DL, and vB_AdhM_TS9, for further studies.
The optimal MOI of phage vB_AdhS_TS3, vB_AdhM_TS9, vB_AdhM_DL, and vB_AdhS_M4

One-step growth curve of vB_AdhS_TS3 (A), vB_AdhM_DL (B), vB_AdhM_TS9 (C), and vB_AdhS_M4 (D). Data are the mean of triplicate independent experiments with standard deviation.
pH and thermal stability
The pH and thermal stability of phages were determined by the change in the number of plaque-forming units (PFU). As shown in Fig.5, all phages were resistant to a wide range of pH values after 2 h of incubation, and the optimum range was pH–6-8 (Fig. 5). However, phages vB_AdhS_TS3 and vB_AdhM_DL showed a significant decrease at pH 3 and 12, whereas phage vB_AdhM_TS9 was inactivated at pH 12. No plaques were seen at pH 2.
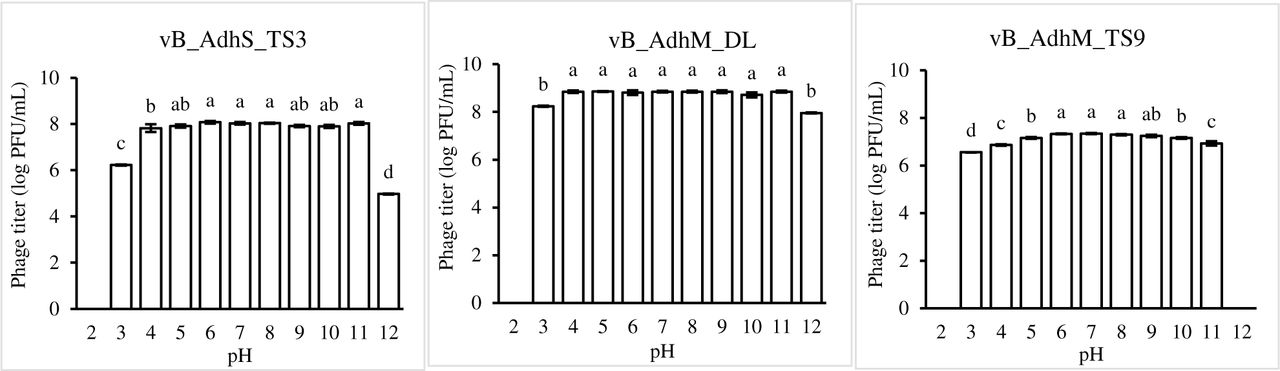
pH stability of phages vB_AdhS_TS3, vB_AdhM_DL, and vB_AdhM_TS9. Phages were incubated for 1 h under different pH values. The values are the mean of three determinations. Different letters indicate significant differences between pH treatments (p <0.05).
The thermal stability of the isolated phages was determined at pH 7.0. Figure 6 shows that all the phages remained relatively stable after a 60-min incubation at 4 °C, 25 °C, 30 °C, and 37 °C but were sensitive to higher temperatures (Fig. 6). At 100 °C, phage vB_AdhS_TS3 was completely inactivated within 5 min, whereas phages vB_AdhM_DL and vB_AdhM_TS9 were undetectable after 10 min (Fig. 7).

Thermal stability of phages vB_AdhS_TS3, vB_AdhM_DL, and vB_AdhM_TS9. Phages were incubated for 1 h at different temperatures. The values are the mean of three determinations. Different letters indicate significant differences between pH treatments (p < 0.05).

Thermal stability of phages vB_AdhS_TS3, vB_AdhM_DL, and vB_AdhM_TS9 at 100°C.
Whole-genome sequencing of phages and in silico analysis
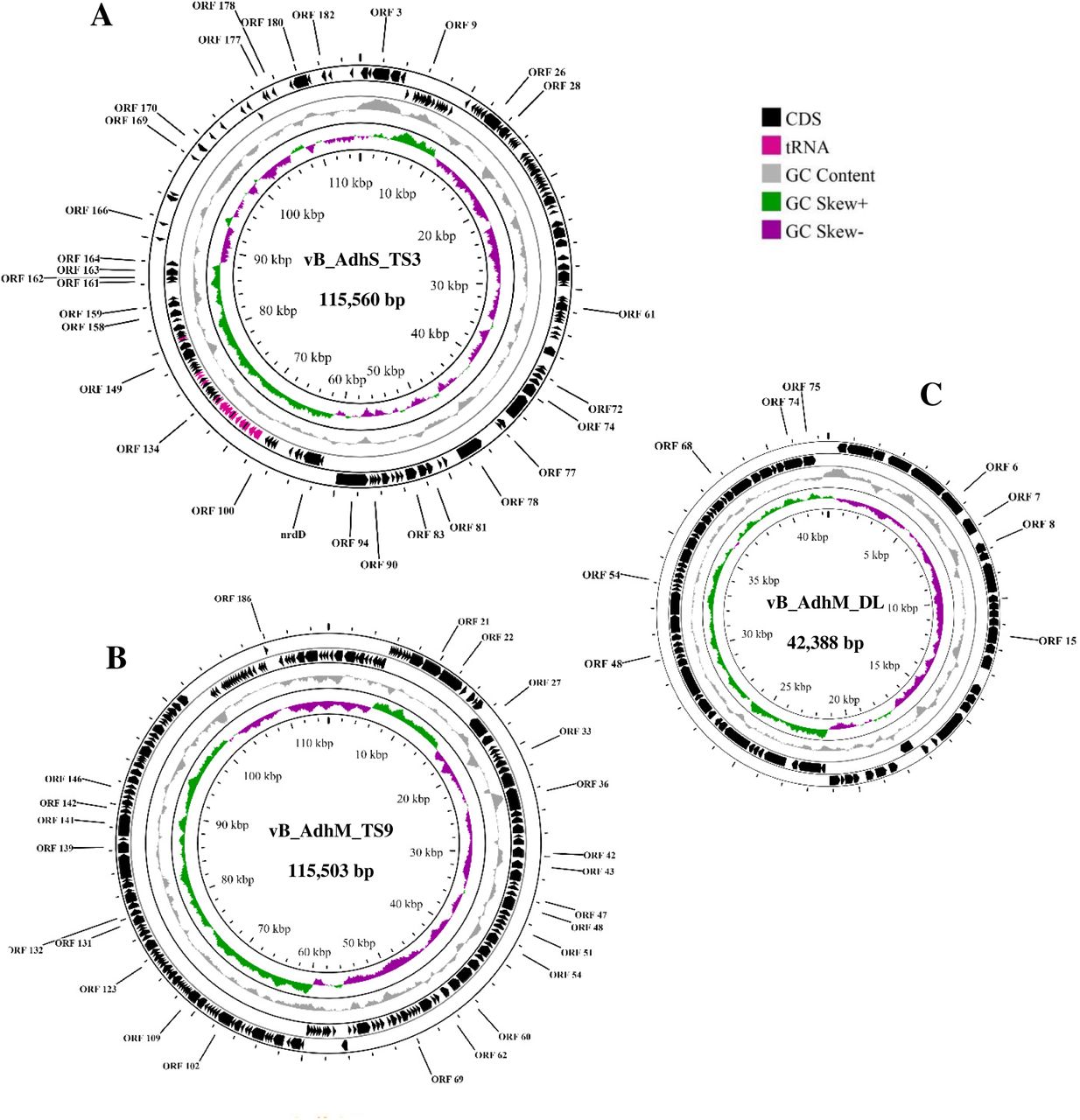
The genome size of vB_AdhS_TS3 was 115,560 bp with a G + C content of 41.10%. BLASTn analysis showed that the genome sequence of vB_AdhS_TS3 had the highest similarity to that of Aeromonas phage Akh-2 (81.90%). Furthermore, vB_AdhS_TS3 encodes 30 transfer-RNAs (tRNA), as shown in Table 6. With regards to the gene function, the open reading frames (ORFs) of vB_AdhS_TS3 were identified by BLASTp, and a total of 151 ORFs were predicted. Among these genes, 30 were predicted to have known functions (Table 7, Fig 8A), and 121 ORFs were predicted to encode hypothetical proteins. Similarly, the genome size of vB_AdhM_TS9 was 115,503 bp, with a G + C content of 35.34%, and encoded 195 proteins. BLASTn analysis showed that the genome sequence of vB_AdhM_TS9 had the highest similarity to the Aeromonas phage phiA047 (94.43%). Phage vB_AdhM_TS9 does not appear to encode tRNA genes. Out of 195 ORFs, 175 ORFs were hypothetical, whereas only 25 ORFs predicted functions (Table 8, Fig 8B). The genome size of vB_AdhM_DL was 42,388 bp, with a G+C content of 34.43% and 75 proteins, respectively. BLASTn analysis showed that the genome sequence of vB_AdhM_DL had the highest similarity to the Aeromonas phage phiA019 (94.43%). Of the 75 encoded proteins, Only 9 out of 75 encoded predicted functions, whereas 66 ORFs were hypothetical (Table 9, Fig 8C). We did not find an ORF encoding a protein with known toxins, antibiotic-resistant genes (ARGs), virulent factors (VFs) of bacterial origin, or lysogenic markers such as integrase, recombinase, repressor/anti-repressor protein, and excisionase in all three phage genomes. Furthermore, PhageAI was used to classify vB_AdhS_TS3, vB_AdhM_TS9, and vB_AdhM_DL as virulent (lytic phage), with high confidence (99.10%, 90.70% and 91.21% respectively).
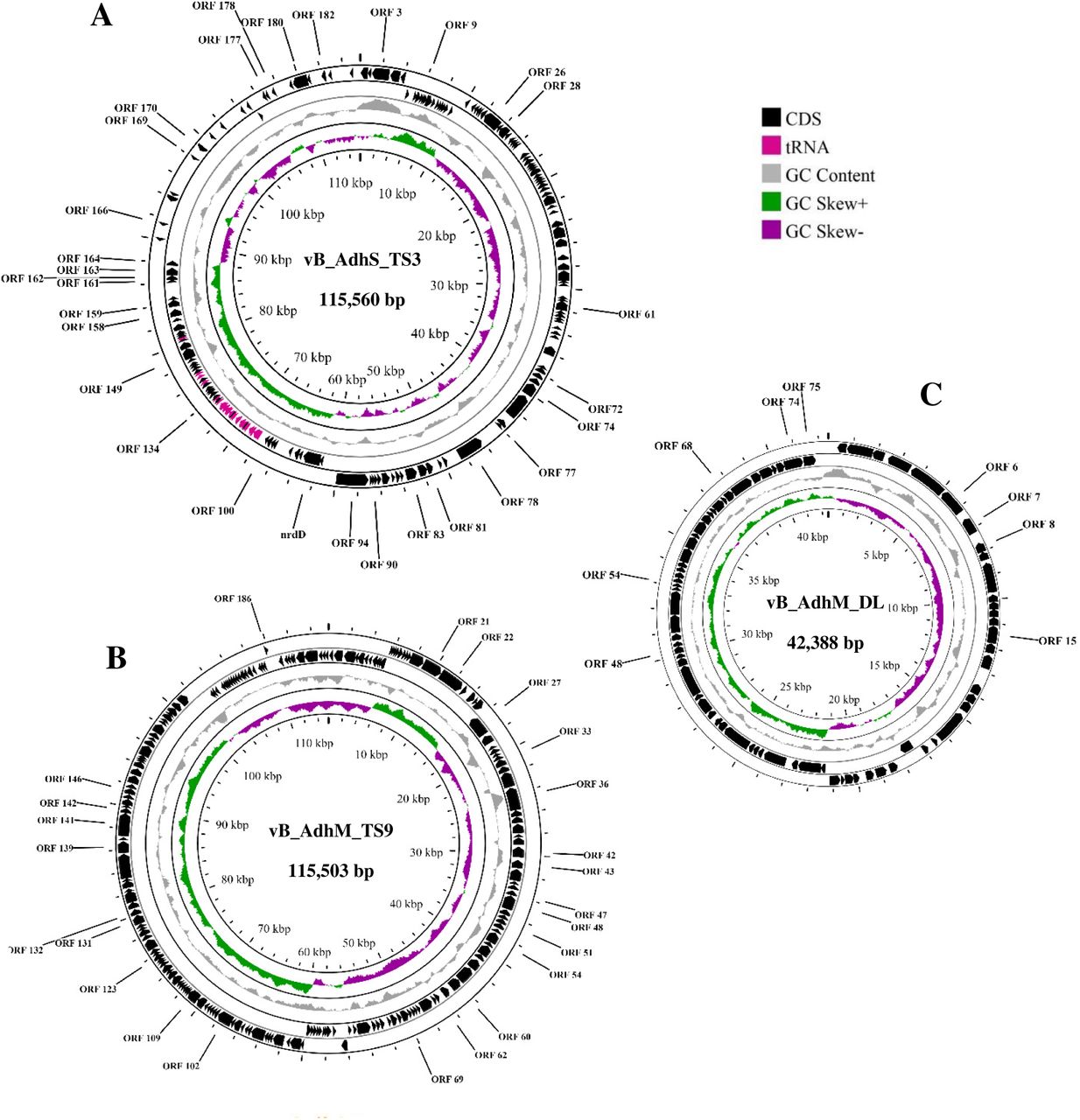
The genome map of three phages A) vB_AdhS_TS3 B) vB_AdhM_TS9 C) vB_AdhM_DL. Circles from outermost to innermost correspond to predicted genes (BLASTp, nr database, E value of <10-5) on the forward strand, reverse strand, and GC content.
Genomic characterization of three bacteriophages targeting A. dhakensis AM
Features of the ORFs of phage vB_AdhS_TS3, Predicted functions of proteins, and best matches with databases
Features of the ORFs of phage vB_AdhM_TS9, predicted functions of proteins, and best matches with databases
Features of the ORFs of phage vB_AdhM_DL, Predicted functions of proteins, and best matches with databases
Effect of single in pre- and post-treatment to control A. dhakensis AM growth
The lytic effect of individual phages on the growth of A. dhakensis AM was evaluated at different MOIs. Both pre-and post-treatment, the maximum cell decrease for all phages was observed during 6-12 h of incubation at all MOIs compared with the uninfected bacterial control. The pre-treatment with phages vB_AdhS_TS3, vB_AdhM_DL, and vB_AdhM_TS9 reduced the maximum bacterial count by 5.40, 6.67 and 3.91 log CFU/mL, respectively, after 6 h of incubation. In post-treatment, the maximum inactivation was achieved at 12 h with the log reduction number of 4.68, 5.25 and 4.43 log CFU/mL, respectively. The growth of bacteria cultured with phages decreased remarkably depending on the regrowth of bacteria at 48 h in all treatments. The maximum growth of the bacterial control was approximately 6-7 log CFU/mL after 48 h of incubation (Fig. 9), whereas the CFU levels of the experimental group treated with only phage remained constant throughout. When the phages were incubated in the presence of the host, the phages gradually increased and then became stable over 48 h of incubation. Based on the maximum inhibition, the combination of two phages as a phage cocktail in pre- and post-treatment with optimal MOIs was chosen, as shown in Fig. 10.

The log reduction in A. dhakensis number in pre- and post-treatment using single phage vB_AdhS_TS3, vB_AdhM_DL and vB_AdhM_TS9. The data were expressed as mean ± SD. All assays were carried out in triplicates.

The log reduction in A. dhakensis number in pre- and post-treatment groups using phage cocktail vB_AdhS_TS3, vB_AdhM_DL and vB_AdhM_TS9. The data are expressed as mean ± SD. All assays were carried out in triplicates.
Effect of phage cocktail in pre- and post-treatment to control A. dhakensis AM growth
The effectiveness of the phage cocktail in the reduction of A. dhakensis AM is shown in Fig. 10. Cocktail 3, composed of phages vB_AdhM_TS9 and vB_AdhM_DL, was more effective against A. dhakensis AM than the other cocktails. Upon pre-treatment, the maximum inactivation with cocktail 3 (vB_AdhM_TS9 (MOI 1)+ vB_AdhM_DL (MOI 0.1)) was 5.08 ± 0.51 log CFU/mL after 6 h of incubation compared with uninfected control. In post-treatment, the maximum reduction with cocktail 3 (vB_AdhM_TS9 (MOI 1) + vB_AdhM_DL (MOI 1)) was 4.71 ± 0.49 log CFU/mL after 12 h of incubation when compared with those of the bacterial control. Bacterial regrowth was observed at 24 h in all treatments. The phage alone was constant throughout the experiment. Although phage cocktails are promising for preventing the emergence of phage-resistant mutants, the incubation of phages and bacteria for longer periods may result in the appearance of phage-resistant strains (68). Therefore, the combination of phages and antibiotics has also been studied to minimize the negative side effects of antibiotics.
A. dhakensis growth inhibition by phage cocktail and antibiotics combination
To establish the phage-antibiotic synergy (PAS) effect, we determined the bacterial inactivation by three combinations of phage cocktails with amoxicillin at sub-MIC (32 μg/mL) in different volumes (200 µL and 20 mL). In the presence of amoxicillin and phage alone, the antibiotic- and phage-resistant variants rapidly grew after 6 h of incubation. In the pre-treatment, the combination of phage cocktail 1 or 2 with amoxicillin at sub-MIC resulted in complete inhibition during 48 h and 12 h in a volume of 200 µL and 20 mL, respectively (Fig. 11). At a volume of 20 mL, a significant reduction in bacterial numbers was observed when treated with a combination of phage cocktail 1 or 2 and sub-MIC amoxicillin at 48 h of incubation (p < 0.05). After post-treatment, the combination of phage cocktail 1 or 2 with amoxicillin at sub-MIC resulted in complete inhibition for 48 h in 200 µL (Fig. 12). However, only partial inhibition was observed after 12 h at a volume of 20 mL. Bacterial regrowth gradually increased after 12 h, and no significant reduction in viable bacteria was observed after 48 h of incubation compared to the phage cocktail of antibiotics alone. In this study, the bacterial concentration in this treatment (1 × 105 CFU/mL) was much higher than in natural bacterial contamination. Moreover, this study was performed in a higher volume of medium (20 mL), which may reduce the interaction between phages and/or antibiotics before reaching the bacteria. However, phage cocktails 1 and 2 decreased the CFU 1.2-1.7 log CFU/mL compared to the control and other groups treated individually after incubation for 48 h. Our study strongly suggests that the synergistic antibacterial effects of antibiotics and phages should be performed in the early stages when the bacterial number is low. This indicates the very efficient inactivation of bacteria by the antibiotic at sub-MIC in combination with the phage cocktail. The first use of the phage‒antibiotic synergy (PAS) strategy was described by Comeau (2007) (69). Sublethal concentrations of antibiotics may help lytic bacteriophages reproduce rapidly and promote their antibacterial effects. Moreover, when bacteriophages are used in combination with antibiotics, there may be a profound order effect; bacteriophage treatment implemented before drug treatment results in maximum bacterial killing.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of phage cocktail and amoxicillin combination at 1/2 MIC against A. dhakensis AM. The bar graph represents the bacterial concentration (log CFU/ mL), and the line graph represents the phage concentration (log PFU/ mL). The data are expressed as mean ± SD. All assays were carried out in triplicates. Each lowercase label corresponds to a significantly diffrerent (p < 0.05) bacterial concentration within each time point. Capital letters denote significantly distinct (p < 0.05) bacterial concentrations and time points compared to each other time point within the same conditions.
Our study demonstrates that phage-based approaches are an attractive way to inactivate A. dhakensis in vitro. The cocktail of three different bacteriophages (phage vB_AdhS_TS3, vB_AdhM_DL and vB_AdhM_TS9) revealed promising in vitro lytic activity on A. dhakensis. Furthermore, the combination therapy using phage cocktails and antibiotics showed greater promise compared with either therapy alone. Moreover, combination therapy can also prevent the development of resistant mutants that would otherwise develop rapidly when exposed to antibiotics or phages. This demonstrates that using phages as an adjuvant with a sublethal concentration of antibiotics is an effective therapeutic strategy.
MATERIALS AND METHODS
Bacterial strains and culture conditions
All Aeromonas strains and other Aeromonas reference strains were grown on nutrient agar (NA) plates or in nutrient broth (NB) (Himedia, India). All Aeromonas stock cultures were stored at -20 °C in NB containing 20% (v/v) glycerol. Before use, frozen cultures were cultivated at 30 °C with shaking at 200 rpm in NB. The culture broth was then streaked on NA, and a single colony was transferred to the NA slant for further study.
Isolation of Aeromonas
To isolate Aeromonas species, 30 samples were collected from different sources, including fishponds, canal water, and rivers in Bangkok, Thailand. All samples were collected in sterile bottles, stored in a cooler, and transported to the laboratory for analysis. The samples were streaked onto an Aeromonas isolation medium (HiMedia, India) supplemented with ampicillin. The plates were incubated for 24 h at 30 °C. The dark green, opaque, dark center colonies resembling Aeromonas sp. were re-streaked on fresh Aeromonas isolation medium until a pure culture was obtained. The pure isolates were subjected to gram staining, nitrate reduction test, glucose OF (oxidation-fermentation) test, and oxidase and catalase test. Gram-negative bacteria capable of degrading nitrates to nitrites, glucose fermenters, oxidase, and catalase-positive isolates resembling the genus Aeromonas were selected for 16s rRNA gene sequencing analysis. Other biochemical tests were used to differentiate between Aeromonas genera. L-arabinose fermentation was also differentiated between A. hydrophila and A. dhakensis. Likewise, salicin fermentation allowed differentiation between A. hydrophila and A. dhakensis from A. hydrophila subsp. ranae (10). Hemolysis test was performed on Columbia agar supplemented with sheep blood. All Aeromonas spp. isolates were stored at 20 °C in nutrient broth (NB) (Himedia, India) supplemented with 20% glycerol until further use.
Identification of Aeromonas spp
The genomic DNA of the Aeromonas isolate was extracted using an AccuPrep® Genomic DNA Extraction Kit (Bioneer, Korea) following the manufacturer’s instructions and used as templates for PCR amplification. A pair of universal primers, 27F:5′-AGAGTTTGATCCTGGC TCAG-3′ and 1492R:5′-GGCTACCTTGTTACGACTT-3′ was used to amplify the 16S rRNA gene. PCR amplification was performed in a thermal cycler ((Mastercycler, Eppendorf, USA) using the conditions described by Pringsulaka, Patarasinpaiboon, Suwannasai, Atthakor, & Rangsiruji (2011) (29). The amplification products were separated on a 0.6% agarose gel and visualized under UV light after staining with ethidium bromide (1 mg/mL). A MinElute Gel Extraction Kit (Qiagen, USA) was used to purify the 16S rDNA fragments, which were sequenced and compared to the GenBank database using the Basic Local Alignment Search Tool (BLAST). Phylogenetic trees were established using the neighbor-joining method in the MEGA 5.1 software package following Han et al. (2017) (30). The amplification products obtained based on the 16S rDNA specific for the A. hydrophila gene primers were compared with the corresponding PCR fragments of the reference strain of A. hydrophila TISTR 1321.
Antimicrobial susceptibilities of Aeromonas isolates
Aeromonas isolates were cultured on nutrient agar (Himedia, India) for 24 h at 30 °C, after which a suspension was prepared in sterile saline, adjusted to 0.5 McFarland standard, and diluted to reach a final bacterial inoculum concentration of 5 × 105 CFU/mL. Next, 100 μL was spread onto Mueller Hinton (MH) agar plates (Himedia, India). Then, the MIC test strips containing amoxicillin, chloramphenicol, doxycycline, gentamicin, and tetracycline (Liofilchem® MTS™, Italy) were placed at the center of the plate and incubated for approximately 24 h at 30 °C. The results were then read by evaluating the ellipsoid zones of bacterial growth inhibition and examining the intersection of this zone and the concentration mark on the test strip, which indicated the MIC. The interpretative criteria were derived from those described by the Clinical and Laboratory Standards Institute (CLSI) VET04 (31).
Phage isolation and detection
The isolated Aeromonas strains were used as hosts for bacteriophage isolation. Fresh overnight cultures of Aeromonas were propagated in 25 mL of double-strength NB in a 250-mL Erlenmeyer flask for 4-6 h. Then, 25 mL of canal and fishpond samples were added and further incubated at 30 °C for 24 h. The culture broth was centrifuged at 8000xg for 10 min, and the supernatant was filtered using a 0.45-μm pore-size syringe filter. The double-layer agar plate method was used on NA medium to determine the presence of phages (32, 33). Bacteriophage plaques were enumerated after overnight incubation at 30 ° °C and calculated as plaque-forming units (PFU/mL). Single plaques were isolated and purified by picking plaques into 300 μL of the NB containing early exponential phase Aeromonas spp. and incubating at 30 °C overnight. This step was repeated three times to ensure the isolation of a pure phage. Phage stocks were prepared in NB supplemented with 30 mM CaCl2 (NB-Ca) and then stored in a dark bottle at 4 ° C.
Electron microscopy
Phage morphology was visualized by transmission electron microscopy (TEM). Carbon-formvar-coated grids were gently placed on fresh overnight plaques and negatively stained with 1% (w/v) uranyl acetate (pH 4.5). Phage morphology was examined using a TECNAI 20 TWIN transmission electron microscope operated at 120 kV and a magnification of x120,000.
Host-range determination
The host range of the isolated phages was determined using the spot test method (34). Other reference strains of aeromonads were tested for susceptibility to phages. Briefly, 100 μL of each reference strain (OD600 =1) was added to 4 mL of liquefied NB soft agar (NB broth with 0.5% agar), mixed gently, and poured over the NA plate. After the agar solidified, 10 μL of phage filtrate was spotted onto NB soft agar and incubated overnight at 30 ° °C and bacterial sensitivity to the phage was indicated by a plaque at the spot. The clarity of the spots was recorded into three categories: clear (transparent) spots (++), turbid spots (+), and no lysis zones (-).
Determination of optimal multiplicity of infection (MOI)
A host strain suspension (108 CFU/mL) in NB was mixed with the phage stock at four different ratios (0.01, 0.1, 1, and 10 PFU/CFU) to determine the optimal MOI. After 3.5 h incubation at 30 °C, the samples were harvested and assayed to determine the phage titer using the double-layer agar plate method (29). The ratio with the highest phage titer was the optimal MOI.
One-step growth curve experiments
A one-step growth curve for each phage isolate was performed as Sunthornthummas et al (2017) (35) with some modifications. The A. dhakensis culture (108 CFU/mL) was resuspended in 1 ml of NB and incubated with an equal volume of phage (107 PFU/mL) to generate an MOI of 0.1. After 10 min of adsorption at 37°C, the suspension was filtered through a 0.45 µm pore size syringe filter. Unadsorbed phages were removed from the filter membrane by washing the filter several times with MRS broth. Infected cells were harvested and resuspended in 20 ml of MRS broth. An aliquot of each dilution was withdrawn at intervals for phage counts using the double-layer agar plate method (36). The latent period, rise period, and burst size were calculated using the one-step growth curve (32).
pH and thermal stability
For the pH stability tests, NB was pre-adjusted to a wide range of pH values (pH 2.0, 3.0, 4.0, 5.0, 8.0, 9.0, 10.0, and 11.0) with 1N HCl or 1N NaOH. A phage suspension (1010 PFU/mL) was inoculated and incubated. After 90 min of incubation at 30 °C, the phage titer was determined using the double-layer agar plate method. For thermal inactivation experiments, phage lysates (1010 PFU/mL) were subjected to heat treatment at 4, 30, 37, 45, 63, 72, and 100 ° °C in NB. The tubes were harvested at regular intervals and placed in an ice-water bath. The phage titer was determined using the double-layer agar plate method, and the phage survival rates were calculated by dividing the PFU at each time point by that at the primary PFU. All tests were performed in triplicates.
Whole Genome sequencing and in silico analyses
Genomic DNA of Aeromonas sp. AM was extracted using an AccuPrep Genomic DNA Extraction Kit (Bioneer, Daejeon, Korea). Phage DNA was isolated as previously described (35). The phage DNA pellet was resuspended in 100 mL TE buffer (pH 8.0). The concentration and DNA quality were evaluated using a NanoDrop™ 2000 spectrophotometer (Thermo Fisher Scientific, USA). High-purity genomic DNA with an OD260/OD280 ratio of 1.8–2.0 and an OD260/OD230 ratio of 2.0–2.2 was used for whole-genome sequencing. The purified genomic DNA was sent to the Beijing Genomics Institute (BGI) in China for short-read sequencing.
De novo assembly of Aeromonas sp. AM and three phage genome sequences were constructed using SPAdes 3.12 (37). The quality of the reads was investigated using FASTQC (38) and trimmed with Trimmomatic 0.39 (39). Functional annotation was performed using Prokka v1.14 (40) and tRNAs and rRNAs were identified using tRNAscan-SE 2.0 (41).
Bioinformatics analyses
Nucleotide and amino acid sequences were compared using Blastn software. Translated open reading frames (ORFs) were compared to the non-redundant GenBank protein database using the Blastp software. Additionally, the genomic DNA of Aeromonas dhakensis AM and three phages were screened for the presence of virulence genes using the Virulence Factors of Pathogenic Bacteria (VFDB) (42), Comprehensive Antibiotic Resistance Database (CARD) databases (43), and PlasmidFinder 2.1 (44, 45). PHASTER was used to identify prophages in bacterial genomes (46, 47), and the lifestyles of the three phages were predicted computationally using PhageAI (https://phage.ai/) (48). The genome of Aeromonas sp. AM and the three phages were visualized using the (CGView) (https://beta.proksee.ca/) webserver (49).
Accession Numbers
The genome sequences of Aeromons dhakensis AM were deposited in the NCBI database under accession number JAPHNH000000000, and the genome sequences of phage vB_AdhS_TS3, vB_AdhM_TS9, and vB_AdhM_DL were deposited in the NCBI database under accession number OP820700, OP820701 and OP820702, respectively.
A. dhakensis growth inhibition by single phage and phage cocktail in vitro
Phage therapy was divided into two treatments: pre- and post-treatment. In the pre-treatment experiment, phage vB_AdhS_TS3, vB_AdhM_TS9, vB_AdhM_DL, and vB_AdhS_M4 or phage cocktail were added before inoculation with A. dhakensis AM (1 × 108 CFU/mL), resulting in MOIs of 0.1, 1, and 10, respectively. In the post-treatment experiment, A. dhakensis AM suspensions (1 × 108 CFU/mL) were inoculated into NB and incubated for 3 h (logarithmic growth phase). Equal volumes of phage vB_AdhS_TS3, vB_AdhM_TS9, vB_AdhM_DL, and vB_AdhS_M4 or phage cocktail were added at MOIs of 0.1, 1, and 10, respectively. Both treatments were performed in a 250 mL Erlenmeyer flask containing 50 mL of NB at 200 rpm and incubated at 30 °C for 48 h. For each assay, two control samples were set: the bacterial control and the phage control. The bacterial control was inoculated with A. dhakensis but not phages, and the phage controls were inoculated with phages but not bacteria. The control and test samples were incubated under the same conditions. Aliquots of the test samples and their controls were sampled at 0, 6, 12, and 24 h of incubation. In all assays, phage titer was determined in triplicate using the double-layer agar plate method. The bacterial concentration was determined in triplicate in the NA medium. Three independent experiments were performed for each condition.
A. dhakensis growth inhibition by phage cocktail and antibiotics combination
The inhibitory effects of the selected two-phage cocktail with effective MOIs in combination with antibiotics at sub-MIC (1/2 MIC) were determined as previously described. In the pre-treatment experiment, a combination of the selected two-phage cocktail with effective MOIs and amoxicillin at sub-MIC was added before inoculation with A. dhakensis AM (1 × 105 CFU/mL). In the post-treatment experiment, A. dhakensis AM suspensions (1 × 105 CFU/mL) were inoculated into NB and incubated for 3 h. Equal volumes of the selected three-phage cocktail with effective MOIs and amoxicillin at sub-MICs were added. Only the phage cocktail and antibiotics at MIC were also administered in both pre-and post-treatment. For each assay, two control samples were set up: one containing only bacteria and the other containing only phage. Phage and bacterial counts were determined in NB in two different volumes: 200 μL in 96-well microtiter plates and 20 mL in 250 mL Erlenmeyer flasks. The latter was incubated on an orbital shaker with a shaking speed of 200 rpm. After incubation at 30°C, the aliquots of each sample and their controls were collected every 6 h for 48 h and were serially diluted to determine viable bacteria (CFU/mL) in NA plates incubated for 24 h at 30°C. The experiment was performed in triplicate.
Statistical Analysis
Statistically significant differences in all experiments were determined by one-way analysis of variance (ANOVA), and post-hoc Tukey’s test was applied to illustrate significant differences between bacterial concentrations between treatment groups over time. A p-value <0.05 was considered to indicate statistical significance. SPSS statistical software package (version 13.0) was used for all analyses.
ACKNOWLEDGEMENT
This work was supported by an annual government statement of expenditure, Srinakharinwirot University (grant number 031/2564), and graduate school fund, Faculty of Science, Srinakarinwirot University.
REFERENCES




