

ABSTRACT
Flagella of the unicellular green alga Chlamydomonas reinhardtii are nearly identical to cilia of mammalian cells and provide an excellent model to study ciliogenesis. These biflagellated cells have two actin genes: one encoding a conventional actin (IDA5) and the other encoding a divergent novel actin-like protein (NAP1). Previously, we described a role for actin in regulation of flagella-building intraflagellar transport machinery and now probe how actin redundancy contributes to this process using a nap1 mutant Chlamydomonas strain. Treatment with Latrunculin B, a potent actin polymerization inhibitor on the nap1 mutant background acutely disrupts all filamentous actins in the cell. We find that actins are an absolute requirement for flagellar growth when the preexisting pool of flagellar precursors is depleted. Nap1 mutants treated with Latrunculin B also showed reduced protein synthesis during regeneration. Finally, loss of functional actins reduced the incorporation of existing flagellar proteins as well as caused mislocalization of a key transition zone gating protein, NPHP-4. These experiments demonstrate that each stage of flagellar biogenesis requires redundant actin function to varying degrees, with an absolute requirement for actin in incorporation of newly synthesized flagellar proteins.
INTRODUCTION
Assembly and composition of the eukaryotic flagellum (also known as the cilium) are critical for signaling and development in nearly all cells of the human body. The flagella of the green alga Chalmydomonas reinhardtii are nearly identical to cilia of mammalian cells and provide an excellent model to study cell signaling, cell motility, and regulation of ciliary assembly. To date, the known mechanisms dictating the behavior of these organelles are largely dependent upon the microtubule cytoskeleton. The flagellum is composed of microtubules, extends from a microtubule organizing center known as the basal body, and its assembly requires control of microtubule dynamics. Trafficking from sites of cellular protein synthesis to their ultimate destination in flagella is also thought to occur on microtubule tracks (Tai et al., 1999)
However, evidence for the role of another major cytoskeletal component, actin, in ciliary regulation is increasing. In mammalian cells, disruption of actin leads to increases in both ciliary length and percentage of ciliated cells (Kohli et al., 2017; Sharma et al., 2011), which may be due to roles for actin in basal body migration, docking, and stabilization (Kim et al., 2010; Pan et al., 2007; Park et al., 2008; Tu et al., 2017; Yeyati et al., 2017). Actin is also required for vesicle budding in the endocytic pathway (Girao et al., 2008), which may influence ciliary assembly as there is a trafficking pathway connecting the endocytic compartments to a vesicular compartment involved in ciliary assembly (Kim et al., 2010). Membrane remodeling for both ciliary exocytosis (Nager et al., 2017) and ciliary resorption are actin dependent processes (Saito et al., 2017).
We previously identified functions of actin in ciliary assembly using the flagella of Chlamydomonas reinhardtii as a model. In this system, we are able to induce flagellar severing on demand to allow synchronous and successive rounds of flagellar regeneration. Chlamydomonas has two actin genes, one that encodes a conventional actin (IDA5) and another that encodes a novel actin-like protein (NAP1) with ~65% homology to mammalian actin (Hirono et al., 2003). Ida5 mutants and myosin inhibited cells had impaired flagellar motor recruitment to basal bodies, impaired entry of motors into the flagellar compartment, and ultimately a reduced initial flagellar growth rate (Avasthi et al., 2014).
Despite defects in flagellar protein recruitment and flagellar assembly in ida5 conventional actin mutants, these mutants ultimately grow flagella to wild-type lengths. If normal lengths can be maintained without actin, is it an important regulator of flagellar assembly? In this study, we asked if more severe flagellar biogenesis phenotypes were masked by redundant roles of the second Chlamydomonas actin NAP1, as NAP1 expression increases upon actin loss or disruption (Onishi et al., 2016). Through genetic and chemical perturbation of both Chlamydomonas actins, we found that the two actin genes have overlapping functions that include flagellar protein synthesis and composition of the flagellar gate. We also found an absolute requirement for these actins in flagellar incorporation of newly synthesized flagellar proteins, suggesting that a necessary trafficking step for flagellar proteins requires actin in addition to microtubules.
RESULTS
Flagellar length maintenance requires at least one functional actin
To investigate if NAP1 participates in the actin-dependent flagellar assembly functions previously identified for the conventional actin IDA5 (Avasthi et al., 2014), we obtained a nap1 null mutant strain isolated on the basis of its sensitivity to Latrunculin B (LatB), a conventional actin depolymerizing agent (Onishi et al., 2016). Cells with genetic disruption of NAP1 and chemical perturbation of IDA5 filaments lack all filamentous actins and ultimately cannot survive for long periods of time. However, acute perturbation using LatB on the nap1 mutant background allow us to probe the functions of both actins on the shorter time scales needed for assessing flagellar dynamics (Figure S1). When wild-type and nap1 mutants are treated with 10μM LatB, wild-type flagella shorten but eventually recover while nap1 mutant flagella continue to shorten (Figure 1A). NAP1 is upregulated upon LatB treatment (Onishi et al., 2016) in wild-type cells but cannot be upregulated in the nap1 mutant. In the event that flagellar defects are due to indirect effects of actin disruption on microtubule organization, we confirmed that there were no gross defects in microtubule organization or microtubule number in LatB treated nap1 mutants (Figure S2). These data suggest that some form of filamentous actin, either IDA5 or NAP1, is required for flagellar maintenance.

A. Cells of each type were exposed to 10μM Latrunculin B. WT flagella shorten but recover while nap1 mutants + Latrunculin B cannot recover. Error bars are 95% confidence interval (n=50). B. When all actin is disrupted in nap1 mutant cells we see the flagella only grow to half length. This suggests functional actin is required for full flagellar assembly. Flagellar lengths were assessed at various time points. Error bars are 95% confidence interval.
Functional actins are required for full flagellar assembly
Ida5 mutants initially assemble their flagella more slowly but ultimately reach wild-type length (Avasthi et al., 2014). However, given that loss of all actins prevented flagellar maintenance (Figure 1), we investigated if flagella could fully assemble when both actins are perturbed by deflagellating wild-type and nap1 mutants via pH shock and regenerating flagella in the presence of LatB. When IDA5 is disrupted on the nap1 mutant background, flagella cannot grow beyond half-length of a typical flagellum (Figure 1B). Nap1 mutants (Figure 1B), ida5 mutants (Avasthi et al., 2014), and wild-type cells that can upregulate NAP1 in response to LatB treatment, can ultimately assemble flagella to full length. This suggests that some form of actin, either filamentous IDA5 or NAP1, is required for full flagellar assembly.
Complete actin disruption blocks all flagellar growth when flagellar precursors are depleted
In Chlamydomonas, cells can assemble half-length flagella following severing (deflagellation) without additional new protein synthesis (Rosenbaum et al., 1969). The existing unincorporated flagellar proteins are often referred to as the flagellar precursor pool. Given that flagella can assemble to half-length when both actins are disrupted, we asked whether the remaining flagellar growth represented incorporation of the precursor pool. To test this, nap1 mutants were deflagellated and treated with cycloheximide to block new protein synthesis and allow the existing precursor pool to be fully incorporated into flagella. This depletes the cell of proteins that are limiting for flagellar growth. Cells are then deflagelledated a second time, cycloheximide is washed out, and LatB is added in to disrupt all actin in the cell. A schematic of this experiment is shown in Figure 2A. With no pool of limiting flagellar precursors, the cell must synthesize, traffic and incorporate newly synthesized flagellar proteins. Under these conditions, complete disruption of actin blocks all flagellar growth (Figure 2B). This suggests that 1) the regeneration of half-length flagella in LatB treated nap1 mutants is due to incorporation of the existing flagellar precursor pool, and 2) that some form of actin is an absolute requirement for newly synthesized flagellar proteins to contribute to flagellar assembly.

A. Schematic of new flagellar protein incorporation assay. B. New flagellar protein incorporation assay, measuring the effects of Latrunculin B on nap1 mutant flagellar regeneration under precursor pool depleting conditions. There is no flagellar growth using newly synthesized protein in nap1 mutants when IDA5 actin is disrupted. Error bars are 95% confidence interval (n=50).
Flagellar protein synthesis is reduced upon actin disruption
During flagellar regeneration in Chlamydomonas, new flagellar proteins are synthesized to allow full assembly (Lefebvre et al., 1978). Since LatB treated nap1 mutant flagella only assemble to half-length (similar to growth in cycloheximide when no proteins can be synthesized) and depletion of the precursor pool completely blocks flagellar assembly, we next tested if actin loss blocked flagellar protein synthesis. Actin has known roles in transcription (Miralles and Visa, 2006) and may be involved in the deflagellation induced expression of flagellar proteins. Synthesis of new flagellar proteins can be quantified using cumulative flagellar growth beyond half-length (the amount of existing flagellar precursor) (Lefebvre et al., 1978). To test actin’s effect on the amount of new protein synthesis at different time points following flagellar severing, nap1 mutants are deflagellated and treated with 10μM LatB at 30 minute intervals. Cells are deflagellated a second time, LatB is washed out, and cycloheximide is added to incorporate the pool of existing limiting flagellar proteins (Figure 3A). By measuring the length of flagella under actin perturbed conditions and subtracting the length of flagella in cycloheximide (when zero protein synthesis takes place), we can infer whether new proteins were synthesized beyond what existed in the precursor pool. When LatB treated nap1 mutants are compared with DMSO treated controls, we see that loss of all actins does indeed reduce the amount of limiting flagellar proteins synthesized (Figure 3B). This experiment uses flagellar length/growth as a proxy for limiting flagellar protein synthesis. Flagellar proteins are so strongly upregulated following deflagellation (Lefebvre, 1986) that global transcriptome analysis of strongly induced genes can be used to identify flagellar components (Stolc V, 2005). This allowed us to use a more direct method to assay protein synthesis by visualizing nascent proteins through labeled puromycin (Goodman and Hornberger, 2013). Puromycin is an antibiotic that blocks translation by incorporating into nascent proteins. Nap1 mutants were deflagellated via pH shock, and treated with 10μM LatB for 30 minutes, when flagellar protein synthesis is expected to be strongly upregulated (Lefebvre et al., 1978). Cells, were then incubated in puromycin, which was subsequently labeled to visualize newly synthesized proteins. Compared to DMSO treated cells, LatB treated cells showed a large decrease in fluorescence (Figure 3C). This result recapitulates the flagellar length data which indicates that less flagellar proteins are synthesized in cells with no functional actin.

A. Schematic representation of new protein synthesis assay. B. Cells were deflagellated in the presence of 10μM Latruculin B and deflagellated a second time washing out the Latruclin B and adding in 10μg/mL Cycloheximide. This process of double deflagellation allows for quantification of newly synthesized limiting flagellar protein when actins are disrupted. Fewer new flagellar proteins are synthesized upon disruption of both actins C. Saturated images to display cells are present in all treatments. We visualized nascent proteins by puromycin labeling. Nap1 mutant cells treated with 10μM Latruculin B and 10μg/mL of cycloheximide are much dimmer than the DMSO control. This recapitulates the flagellar measurement data indicating less proteins synthesized when the cell has no functional actins. For all measurements (n=30).
Actin loss produces flagellar gating defects
During regeneration in LatB treated nap1 mutants (Figure 2B), we noticed that the flagellar growth to half-length appeared slower than cells containing both actins. To test if this represents an inability to incorporate proteins from the existing pool of flagellar precursors, we simultaneously treated nap1 mutants with cycloheximide and LatB following deflagellation. LatB treated cells (both wild-type cells that take time to upregulate NAP1 and nap1 mutants) incorporate the existing pool of flagellar precursors more slowly than cells with intact actins (Figure 4A). We also previously found that during flagellar assembly, there was a small range of flagellar lengths (7-9μm) in which flagellar motor recruitment to basal bodies in ida5 mutants was comparable to controls but motor entry into flagella was reduced (Avasthi et al., 2014). This along with slow incorporation of the precursor pool suggests a role for actin in gating of material already accumulated at the flagellar base. The region thought to be critical for gating functions is the transition zone at the base of flagella. It has connectors between the microtubule core and flagellar membrane. It also houses proteins found to regulate the composition of flagella. Actin itself is found in the transition zone of flagella (Diener DR, 2015) and may function to transport or anchor the transition zone gating proteins. NPHP-4, a gene mutated in the cilium-related kidney disease nephronophthisis, is a crucial component of the ciliary gate that controls entry of both membrane-associated and soluble proteins (Awata et al., 2014). We used a strain expressing HA-tagged NPHP-4 to test the effects of functional actin loss on NPHP-4 localization. In control cells stained with anti-HA antibody, NPHP-4 is seen as two apical spots at the base of flagella (Figure 4C). In cells treated with LatB for 30 minutes, we saw a dramatic loss of apical NPHP-4 localization (Figure 4D). Occasionally we could see some remaining apical staining and the results are quantified in Figure 4B. These results demonstrate that acute disruption of actin with LatB treatment (at a time point prior to NAP1 upregulation) causes a reorganization of the transition zone for loss of a known transition zone gating protein, NPHP-4.

A. Cyclohexamide is used to stop new protein synthesis. The addition of Latrunculin B (or 1% DMSO control) is to test the affect actin disruption has on incorporating flagellar protein. Actin disruption results in slower incorporation. Flagellar lengths were assessed every 30 minutes for 2 hours. Error bars represent 95 % confidence. (**:p<0.05). B. Percentage of cells with two apical spots upon NPHP-4 labeling are quantified .C. Cells expressing HA tagged NPHP-4 were stained with NPHP4 primary antibody and localization is seen at the base of the flagella (white arrows). D. Cells treated with 10 μM Latrunculin B for 30 min were stained with the NPHP4 antibody. We see mislocalizaiton of the NPHP4 protein. White arrows indicate a cell with some remaining apical NPHP4 localization. Red arrows show loss of NPHP4 at the base of the flagella.
DISCUSSION
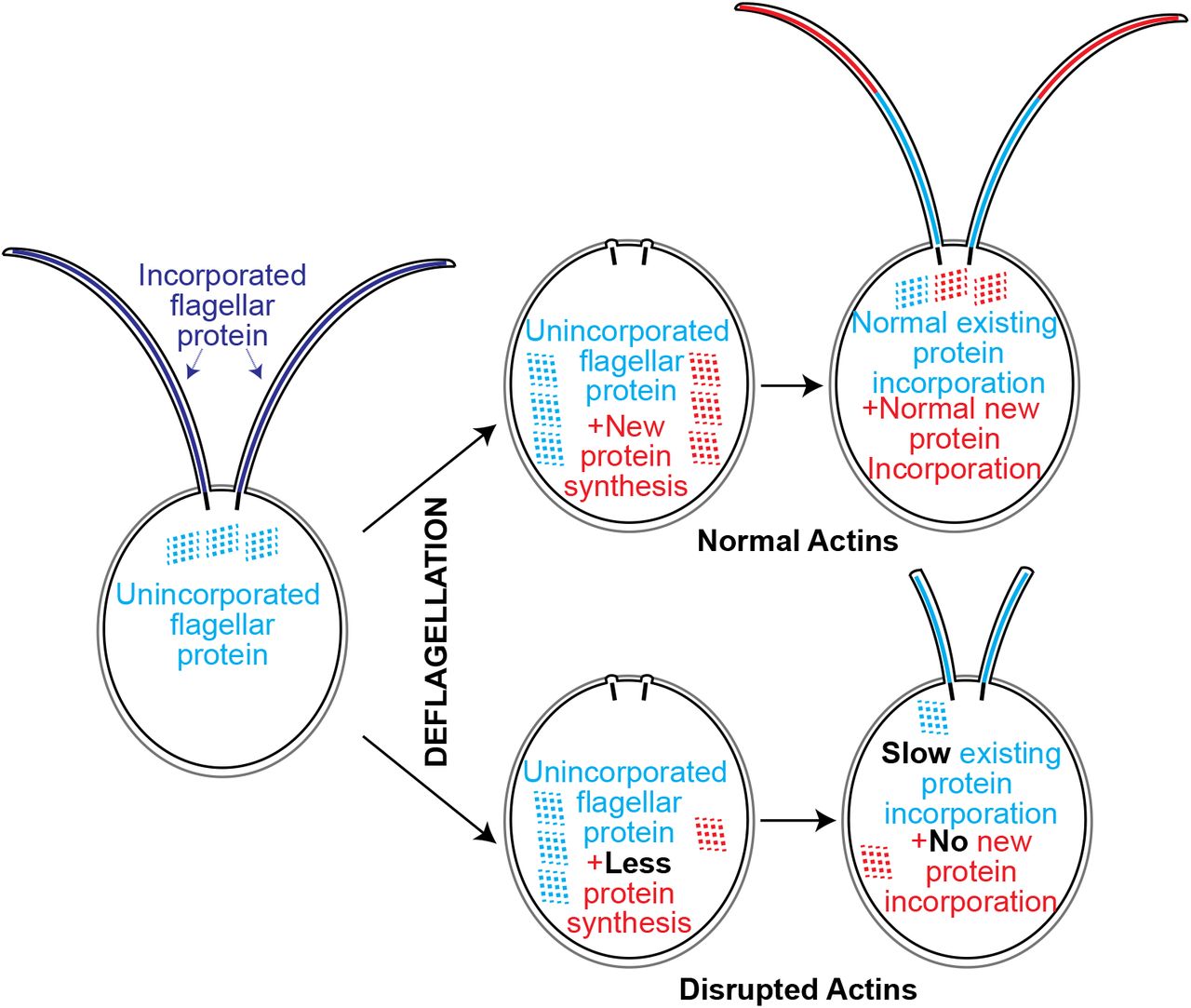
Here we report a requirement of at least one form of Chlamydomonas actin for flagellar length maintenance and full flagellar assembly. Anterograde flagellar motor complexes, which are required for assembly of the flagellum, utilize newly recruited proteins from the cell body pool (Wingfield et al., 2017). With constant turnover and a demand for continuous recruitment to the base of flagella it appears that actin plays a role in this recruitment. Additionally, functional actin is required for normal flagellar protein synthesis and normal incorporation of existing proteins (Figure 5). Importantly, we found that actin-deficient cells cannot grow flagella at all when new flagellar proteins must be 1) synthesized, 2) trafficked, and 3) incorporated (Figure 2). However, while actins are involved, some level of flagellar protein synthesis and incorporation occurs (given the reduced but non-zero values for LatB treated nap1 mutants in Figures 3B and 4A). Together, these data support a model in which actin is an absolute requirement for the trafficking step of growth using newly synthesized flagellar proteins. This may include defects at any step after protein synthesis and prior to entering a cytoplasmic protein pool utilized for flagellar growth. For example, actin may play a role in proper recruitment, compartmentalization, or budding of essential flagellar membrane proteins from the Golgi. Actin may also be required for long range transport of flagellum-bound vesicles from sites of protein synthesis. Further studies looking into these possibilities and the role of myosins that have been implicated by us (Avasthi et al., 2014) and others (Assis et al., 2017) are currently underway.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Loss of all functional actins via depolymerization of IDA5 actin in nap1 mutants results in no flagellar growth using newlhy synthesized protein. Disruption of both actins causes slower protein incorporation and less newly synthesized flagellar protein but cannot account for the inability to grow flagella. We hypothesize functional actin is required for trafficking of newly synthesized protein to the flagella base.
Also novel is the finding that actin is involved in flagellar protein synthesis (Figure 3). While actins are known to localize to the nucleus (Pederson and Aebi, 2002) and interact with transcription factors, chromatin remodeling complexes and RNA polymerases (Miralles and Visa, 2006), it is not well defined whether all or a subset of transcripts require actin function. Given the interaction of actin with core transcriptional machinery, perhaps it is more unexpected that some flagellar protein synthesis can indeed occur in the absence of functional actins.
Lastly, we found a dramatic loss of transition zone localization of NPHP-4 (Figure 4B-D). In Chlamydomonas, actin was also identified through biochemical purification as a transition zone protein and may serve as a scaffold in the region. While NPHP-4 turnover appears more static in Chlamydomonas cells (Awata et al., 2014), actin and myosins found in the peri-basal body region (Assis et al., 2017; Avasthi et al., 2014) may be involved in turnover of transition zone components in mammalian cells where NPHP-4 the turnover rate is high (Takao et al., 2017). Chlamydomonas Nphp-4 mutants in exhibit slow flagellar assembly that ultimately reach wild-type flagellar lengths, similar to ida5 mutants (Avasthi et al., 2014). This may partially explain the slow incorporation of flagellar proteins we see in actin disrupted cells (Figure 4A). Nphp-4 mutants also show decreased membrane protein entry into flagella, suggesting the protein’s importance in regulating flagellar composition. Membrane associated proteins in C. elegans (Williams et al., 2011) and soluble housekeeping proteins in Chlamydomonas (Awata et al., 2014) also inappropriately accumulate within cilia in nphp-4 mutants, suggesting that there is a more general dysregulation of ciliary gating upon NPHP-4 loss. Finally, nphp-4 mutants in C. elegans have ultrastructural abnormalities (Jauregui et al., 2008; Lambacher et al., 2016). Therefore, at minimum, actin disruption, which significantly affects NPHP-4 localization, is expected to encompass the full range of ciliary phenotypes associated with NPHP-4 loss.
Our previous work (Avasthi et al., 2014) and the data presented here allow us to isolate specific steps of flagellar assembly that require normal actin dynamics. Actin has many cellular functions, including in organelle morphology, transcription, membrane dynamics, and polarized trafficking. We are now beginning to see how these conserved actin functions can dramatically influence flagellar biogenesis, a process previously thought to be controlled primarily through microtubule regulation.
MATERIALS AND METHODS
Compounds
Latrunculin B, DMSO and Cycloheximide were purchased from Sigma and used at indicated concentrations and incubated for specified times.
Strains
Wild-type, ida5 mutants NPHP4-HAC were obtained from the Chlamydomonas stock center as CC-125 mt +, CC-3420, and CC-5115. The nap1 mutant was a generous gift from Fred Cross, Masayuki Onishi, and John Pringle.
Inhibitor Treatment and Flagellar Length Measurements
All strains were grown in liquid TAP medium for 24 hours prior to incubation with 10 μM Latrunculin B or 10 μg/mL for indicated times. Cells were fixed in 1% glutaraldehyde and imaged by DIC microscopy at 40 X magnification. Flagellar lengths were measured using ImageJ software by line segment tracing and spline fitting.
Flagellar Regeneration
Flagellar regeneration was induced by deflagellating cells via pH shock by adding 60μL of 0.5N acetic acid followed by 70 μL of 0.5N KOH to 1 mL of cells in liquid TAP. Cells were fixed with 1% glutaraldehyde at 0,30,60,90,120, and 240 minutes. Flagella were measured as described in the method above. Double deflagellation experiments for new protein incorporation were performed as described above (Figure 2A). The cells were deflagellated via pH shock and treated with 10μg/mL of Cycloheximide for 2 hours. The cells were deflagellated a second time and the cycloheximide was washed out and 10μM Lat B was added. The cells were treated for 4 hours and samples were taken every 30 minutes. Flagella lengths were measured using ImageJ software by line segment tracing and spline fitting.
Immunofluorescence Microscopy of NPHP4-HAC
Cells were grown in liquid TAP media 24 hours prior to experiment. Cells were deflagellated prior to incubation with either 10 μM Lat B or 1% DMSO for 30 min. Cells were fixed to coverslips with 4% paraformaldehyde fixation and stained with primary HA anti-rat purchased from Sigma at (1:1000) dilution. The secondary antibody used was also purchased from Sigma and used at (1:1000) dilution as well. Slides were visualized on Nikon TiS microscope on the FITC channel.
New protein synthesis quantification
To determine if the cell can synthesize protein under actin depleted conditions, we performed a double deflagellation experiment to quantify the amount of newly synthesized protein. The method is described in Figure 3A but briefly, the initial deflagellation in this experiment follows treatment with 10 μM Lat B to allow protein synthesis under actin depleted conditions. The first deflagellation runs for a total of 2 hours but cells are deflagellated a second time every 30 minutes where the Lat B is washed out and 10μg/mL cycloheximide is added in to prevent any new protein synthesis. The addition of cycloheximide allows only protein that was synthesized during Lat B treatment to be incorporated into the assembling flagellum. Growth beyond half-length is quantified as newly synthesized protein.
Puromycin labeling for new protein synthesis
The click-it opp kit was purchased from Thermo-Fisher which utilizes a puromycin analog to be incorporated into nascent proteins and then labeled with an antibody to be visualized in the FITC channel on a fluorescent microscope. The protocol was followed according to the instructions provided.
ACKNOWLEDGEMENTS
We would like to thank Masayuki Onishi, John Pringle, and Fred Cross for providing the nap1 mutant. Thank you to memebers of the Avasthi Lab for trouble shooting and manuscript feedback. This work was funded by P20 GM104936-09 (PA) and NSF GRFP 1518767 (BJ).
REFERENCES




