

Abstract
Eusocial insects live in teeming societies with thousands of their kin. In this crowded environment, workers combat disease by removing or burying their dead or diseased nestmates. For honey bees, we found that hygienic brood-removal behavior is triggered by two odors–β-ocimene and oleic acid–which are released from brood upon death. β-ocimene is a co-opted pheromone that normally signals larval food-begging, whereas oleic acid is a conserved necromone across arthropod taxa. Interestingly, the odor blend induces hygienic behavior more consistently than either odor alone. We suggest that the volatile β-ocimene flags hygienic workers’ attention, while oleic acid is the death cue, triggering removal. Hygienic bees detect and remove brood with these odors faster than non-hygienic bees, and both molecules are ligands for hygienic behavior-associated odorant binding proteins. These are key mechanistic insights into some of the molecular interactions that govern disease resistance in a domesticated eusocial insect.
Introduction
Disease and parasite transmission is a constant threat in dense insect societies (1-3). Ants (4-8), termites (9-11), and honey bees (12-16) have evolved social mechanisms of disease resistance which mitigate this risk and improve the collective health of their colonies. Ants transport dead nestmates to their midden heaps, termites bury or entomb their dead in graves, and honey bees remove dead and diseased brood from the hive. E. O. Wilson described these processes as ‘necrophoresis,’ (4) or the movement of dead individuals away from the living colony. Necrophoresis reduces pathogen reservoirs, inhibiting the spread of diseases and parasites between fallen nestmates and those who endure (1).
In honey bees (Apis mellifera), one dominant form of necrophoresis is hygienic behavior (13, 14). Hygienic honey bee workers will identify and remove diseased, dead, and sometimes parasitized brood from the colony. This is an effective defense against major brood diseases, including chalkbrood (Ascosphaera apis) (17, 18), American foulbrood (Paenibacillus larvae) (13, 19), and the devastating varroa mite (Varroa destructor) (19-21). When highly hygienic colonies are inoculated with these pests or pathogens, they are less likely to develop clinical symptoms than non-hygienic hives (19, 22, 23). Furthermore, if hygienic hives do develop symptoms, they are more likely to recover and survive.
The underlying mechanism of the behavior has only been partially deciphered. Like other social insects, honey bees identify their diseased nestmates via chemical cues; however, since their brood develop in the confines of a sealed wax cell, they have an added challenge. The physical barrier between the workers who execute the behavior and the brood interferes with their ability to detect their targets. Detecting the dead brood is thought to rely on volatile odor signals that permeate the brood cell cap (24), but very few hygienic behavior-inducing odors have been identified and confirmed behaviorally (25, 26). Swanson et al. (25) found that a volatile chalkbrood odor (phenethyl acetate) was a strong hygienic behavior-inducer, and Nazzi et al. showed that a volatile varroa-associated odor ((Z)-6-pentadecene) does the same (26). Non-volatile cues have not yet been investigated behaviorally, despite including some of the most taxonomically conserved necrophoretic compounds (e.g. oleic acid and linoleic acid).
Hygienic honey bees have superior olfactory sensitivity compared to non-hygienic honey bees (24, 27), which likely depends in part on differences in antennal gene expression (22, 28-32). In a search for antennal biomarkers for hygienic behavior, we previously identified two odorant binding proteins – OBP16 and OBP18 – that significantly correlated with colony hygienic score (31). Antennae are honey bees’ main olfactory appendages, and OBPs aid odor signal detection by binding and transporting hydrophobic odor molecules from the antennal pores to the olfactory nerves (33). Despite some tantalizing inferences, OBP16 and 18 have not been mechanistically linked to hygienic behavior.
Previously, we compared odor profiles of freeze-killed (dead) brood and healthy brood to find candidate hygienic behavior-inducing compounds (29). We identified two new candidate compounds that were significantly more abundant in freeze-killed brood: oleic acid and β-ocimene. Oleic acid is a minimally volatile, oily substance which acts as a death cue in eusocial and non-eusocial arthropods (4, 7, 34-37). β-ocimene, on the other hand, is a volatile honey bee brood pheromone that is normally a larval food-begging signal (38, 39). Prior to our work in 2017 (29), β-ocimene and oleic acid had not been linked to honey bee hygienic behavior.
In the present work, we investigate oleic acid and β-ocimene’s roles in hygienic behavior using behavioral assays, electrophysiology, and OBP ligand binding assays. Our behavioral assay overcomes a major hurdle in testing the hygienic behavior-inducing capacity of odors: by adding odors through the resealable cells of Jenter™ queen cages, we can add individual odors to brood cells while maintaining perfect integrity of the wax cell walls and cap. By this method, even the non-volatile oleic acid induces hygienic behavior, but the blend of β-ocimene and oleic acid induced hygienic behavior most strongly and consistently. Electroantennogram (EAG) recordings of bees from a hygienic colony show that at a hive-realistic temperature and humidity, oleic acid induces only slightly above-background antennal nerve responses, suggesting that it is only detectable upon contact or extremely close proximity. In vitro ligand binding assays show that both β-ocimene and oleic acid are strong ligands for at least one of OBP16 or OBP18 (which are upregulated in hygienic bees’ antennae). Taken together, we propose a mechanistic model where the co-opted, volatile brood pheromone (β-ocimene) works together with an evolutionarily conserved death cue (oleic acid) via interactions with hygienic behavior-associated odorant binding proteins (OBP16 and OBP18) to induce hygienic behavior.
Results
Previously, we found that β-ocimene and oleic acid were both emitted more strongly from freeze-killed brood compared to live brood, making them promising candidates as putative hygienic behavior-inducers. To test if β-ocimene and oleic acid are sufficient to induce brood removal, we uncapped patches of brood and dispensed 1 μl of neat (100%) or diluted (1%) synthetic odors on the brood (Figure 1A). We call this method the front-way odor assay (FOA).

A) Schematic of the FOA. Patches of capped brood (~30 cells in technical duplicate) developing naturally in a standard frame were uncapped (white patches) and 1 μl of odors (β-ocimene, oleic acid, a 1:1 v/v mix of the two, phenethyl acetate or hexane) at either 1% or 100% concentrations (v/v in hexane) were dispensed onto the brood. Frames were incubated in the colony's brood box for 3 hours before recording removal rates. B) Post-FOA recapping frequencies. Data from N = 9 colonies was analyzed with a one-way ANOVA (F = 13.3, p = 2.4e-8). C) Post-FOA removal frequencies. Hexane is the negative control and phenethyl acetate (a chalkbrood odorant) was meant to be the positive control. Data from 5 hygienic hives and 5 non-hygienic hives was analyzed with a three-factor ANOVA (levels: dose, odor, hygienicity; dose: F = 61.2, p = 4.3e-11; odor: F = 19.8; p = 7.1e-11; hygienicity: F = 20.2, p = 2.7e-5), followed by a Tukey HSD post-hoc test. Significance code (Tukey HSD): * p < 0.05, ** p < 0.01, **** p < 0.0001. For both B) and C), boxes depict the interquartile range (IQR) and whiskers span 1.5*IQR. Letters indicate groups that are significantly different from one another (Tukey HSD P < 0.05).
First, we confirmed that hexane was an appropriate negative control by recording the recapping frequencies following the treatments. We found that after just three hours, an average of 44% of the hexane-treated cells were recapped, which was significantly higher than for all other odors (Figure 1B; one-way ANOVA followed by Tukey HSD; β-ocimene: p = 2e-7; oleic acid: p = 1e-8; mix: p = 1e-8; phenethyl acetate: p = 1e-8). The next highest was β-ocimene, with 5.4% recapped. The others all had recapping frequencies of 1% or less, indicating that the brood were no longer accepted by the workers. In a preliminary test, we confirmed there was no effect of patch proximity on brood removal (N = 5; two-way ANOVA; F = 0.025, p = 0.876). In a second test, we found that workers removed treated pupae and prepupae at similar rates (three-factor ANOVA; levels: dose, odor, age; F = 0.84; p = 0.36). Therefore, we combined data for the two ages and used the FOA to test 3 more colonies, yielding groups of N = 5 hygienic and 5 non-hygienic colonies. We found significant effects of dose, odor, and hygienicity (Figure 1C; three-factor ANOVA; dose: F = 61.2, p = 4.3e-11; odor: F = 19.8; p = 7.1e-11; hygienicity: F = 20.2, p = 2.7e-5).
As expected, brood treated with neat odors were removed significantly more frequently compared to those treated with diluted odors. Furthermore, brood treated with β-ocimene, oleic acid, or their blend tended to be removed more often than those treated with hexane or phenethyl acetate. Specifically, in the neat odor treatments, β-ocimene, oleic acid and their blend all induced significantly higher brood removal relative to hexane (Tukey HSD; p = 0.0034, p = 0.0075, and p = 0.0049 respectively), but in the diluted odor treatments, none of the odors induced significantly different brood removal. However, their relative patterns still reflect what's observed in the neat odor treatments.
If β-ocimene and oleic acid are closely linked to hygienic behavior, we would expect hygienic colonies to respond more strongly to the odor stimuli than non-hygienic colonies. We found that indeed, hygienic colonies removed significantly more treated brood in both the neat odor treatments (Tukey HSD; p = 0.0084) as well as the diluted treatments (p =0.011). This agrees with previous electroantennography studies showing that hygienic bees’ antennae are more sensitive to disease odors than non-hygienic bees (40).
The front-way odor assay is a quick method of gauging if odors can induce brood removal, but it cannot test for odor transmission through the physical barrier of the wax cap. To investigate the odors in a more realistic scenario, we devised a new assay using the Jenter™ system that allows us to treat brood with odors while maintaining the integrity of the brood cells. We call this the back-way odor assay (BOA; Figure 2A), since we add the odors through the back of the brood cell. Briefly, we place a queen in a Jenter™ cage until she lays eggs in the comb of the cage, then release her and allow the workers to rear the brood until it is capped. The back of the Jenter™ cage is equipped with removable plugs that enable odors to be added inside the cell without disturbing the delicate wax cell cap, and plugged again to close the brood cell. We used this method to add neat hexane, β-ocimene, oleic acid and the odor blend to brood cells before and after pupation (N = 5 for each age). We found that after incubating in the hive for 20 h, β-ocimene did not induce significantly more brood removal relative to hexane (Figure 2B; two-factor ANOVA followed by Tukey HSD; p = 0.82 for pre-pupal brood and p = 0.10 for post-pupal brood). However, oleic acid strongly induced pre-pupal removal (p = 0.0004) but only marginally significant post-pupal removal (p = 0.057). The odor blend induced the most consistently high brood removal of them all, which was significant for both brood ages (p = 0.0004 for pre-pupal and p = 0.0003 for post-pupal).

A) Schematic of the BOA. Queens were caged in a Jenter™ queen rearing cage (a hanging square of artificial comb) until she populated the cells with eggs. The queens were released and brood were allowed to develop until capping (front view). We treated brood cells with neat odors in a semi-random design through the cell plugs (back view, brown circles), then the odor-impregnated brood was incubated in the colony for 20 h to allow time for odor diffusion, uncapping, and removal. Diagrams are not to scale. The actual Jenter™ cage has ~ 100 removable plugs (one every 3rd cell). B.) We treated pre-pupal and post-pupal brood with each odor (9-10 brood cells for each age and odor, N = 5 colonies). Data was analyzed using a two-factor ANOVA (levels: age and odor) followed by a Tukey HSD post hoc test. There was a significant effect of odor (F = 20.3, p = 1.51e-7), no significant effect of age (F = 0.16, p = 0.694), and an insignificant but noteworthy interactive effect (F = 1.9, p = 0.157). Letters indicate groups that are significantly different from one another (Tukey HSD p < 0.05). Boxes depict the interquartile range (IQR) and whiskers span 1.5*IQR.
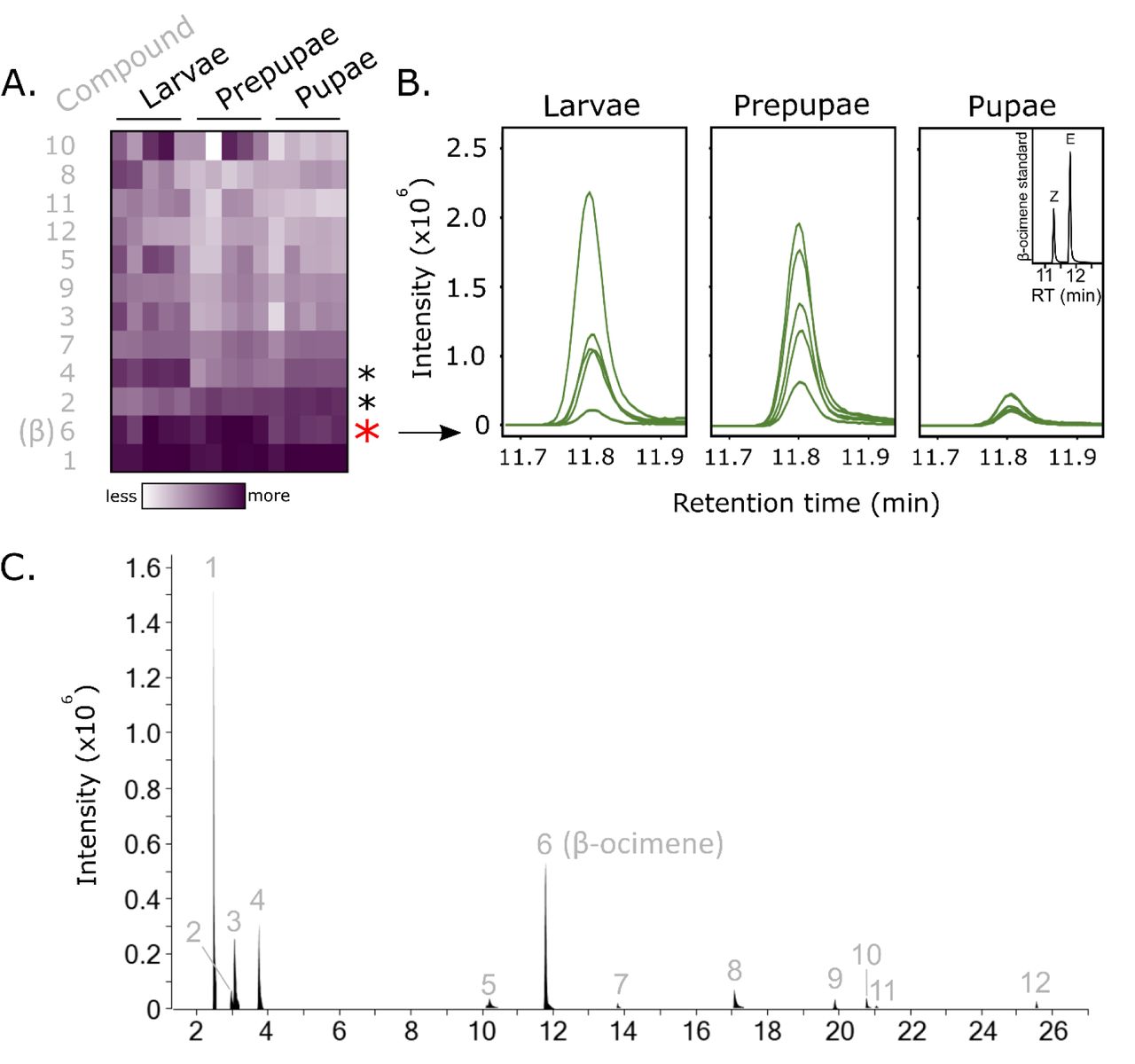
To try to explain the patterns of pre-pupation and post-pupation brood removal, we investigated changes in the background volatile and non-volatile odor profiles that could confound with our odor treatments. To do this, we performed solid-phase micro-extraction gas chromatography-mass spectrometry (SPME GC-MS) on 5th instar larvae, prepupae, and pupae. We also used a hexane wash to extract cuticle compounds from these life stages and analyzed them by GC-MS as well, capturing the less volatile signals. We found that of all the identified volatiles (Figure 3), β-ocimene abundance changed most significantly according to age (one-way ANOVA, Benjamini-Hochberg corrected 1% FDR; p=0.0010, q=0.01), with relatively high amounts emitted in 5th instar larvae and prepupae, and low amounts in pupae (Figure 3B). Other minor chromatogram components were also differentially emitted (compounds 2 and 4, corresponding to isopropanol and 2-pentanone, respectively). The hexane wash identified many branched chain hydrocarbons which were differentially emitted with age but importantly, oleic acid was not among the identified molecules (Figure S1).

We performed solid phase micro-extraction gas chromatography mass spectrometry (SPME-GC-MS) on 5th instar larvae, prepupae and pupae (N = 5 colonies each). A) Heatmap showing intensities of all integrated peaks. Chromatogram peaks were manually integrated and areas under the curve were compared between ages using a one-way ANOVA and Benjamini-Hochberg correction (5% FDR). Each row corresponds to peak intensities belonging to a different compound. β-ocimene, the most significantly different compound, is indicated with a red asterisk, while two other significantly different compounds (matching to isopropanol (2) and 2-pentanone (4) are indicated with black asterisks. B) Chromatogram traces of the β-ocimene peak. Its identity was confirmed with a synthetic standard (inset chromatogram). Based on its retention time, only the E isomer was identified in the brood. C) Example SPME-GC-MS total ion chromatogram. Numbers correspond to compounds labelled in 3A.
Previously, we reported that stimulating honey bee antennae with oleic acid yielded no measurable nerve depolarization signal above the background stimulus of air alone (29). Since we clearly observe that oleic acid can induce hygienic behavior in brood removal assays (including when the brood cell cap remains in-tact), we questioned if the workers were detecting oleic acid-treated cells by olfaction or some other sense (e.g. gustation). To investigate this further, we replicated the electroantennography experiment comparing oleic acid to background stimulation, but at a temperature that better-matches in-hive conditions. When we administered warmed oleic acid (at approximately 33°C) we found that it stimulates worker antennae only slightly more than blank stimuli (Figure 4). There was also a significant effect of odor (two-way ANOVA; levels: odor, side; F=12.4;p =2.3e-5), with β-ocimene and the odor blend inducing significantly higher antennal nerve depolarizations than oleic acid in left antennae (p = 0. 011 and p = 0.016, respectively). The same comparisons yielded a marginally significant response in the right antennae (p = 0.085 and p = 0.086, respectively).

We excised left (N = 13) and right (N = 14) antennae from honey bees in a single highly hygienic colony (score = 95%) and measured the EAG response to neat odors (Syntech™ CS-55) at hive-realistic temperatures (around 33°C). The EAG response represents blank-subtracted odor stimuli. We found a significant effect of odor (two-way ANOVA; levels: side, odor; F = 12.4, p = 2.3e-5), and letters indicate groups that are significantly different from one another (Tukey HSD p < 0.05). Boxes depict the interquartile range (IQR) and whiskers span 1.5*IQR.
Recently, several antennal protein biomarkers for hygienic behavior have emerged, including two odorant binding proteins (OBP16 and OBP18) which are up-regulated in hygienic bees. To test if β-ocimene and oleic acid are strong ligands for these proteins, we performed in vitro binding assays with OBP16 and OBP18. Like our behavioral assays, we used hexane as the odor negative control. We found that of the three tested odors, hexane and phenethyl acetate consistently had the lowest binding affinity (Figure 5, Table S1), which mirrors the behavioral response to these compounds (Figure 1C). β-ocimene bound OBP16 strongly, but not OBP18. Oleic acid, however, bound both OBPs strongly, with OBP18 being the strongest.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
We used an NPN (N-Phenyl-1-naphthylamine) competitive binding assay to measure affinities of β-ocimene, oleic acid, phenethyl acetate, and hexane (negative control). Assays were performed in technical duplicate with 2 μM protein and 2 μM NPN in all cases. Lower NPN fluorescence intensity indicates stronger ligand binding. The high NPN fluorescence intensity for the high oleic acid concentrations is due to the formation of micelles at higher concentrations of the ligands (41, 42). Error bars are standard error of the mean.
Discussion
Hygienic behavior has been studied in honey bees since at least the 1960s (15), but our knowledge of the molecular mechanism behind it is incomplete. In the present work, we investigate two candidate honey bee necromones – β-ocimene (a co-opted brood pheromone (38, 39, 43, 44)) and oleic acid (a well-known necromone in other arthropods (4-6, 9, 35, 36)) – using in vivo, ex vivo, and in vitro techniques. We demonstrated 1) that the odors are sufficient to induce hygienic behavior in realistic behavioral assays (Figure 1C), 2) despite being a viscous compound, oleic acid can stimulate nerve depolarizations worker antennae at hive temperatures (Figure 4), and 3) oleic acid and β-ocimene have high affinities to odorant binding proteins that are upregulated in hygienic honey bees (Figure 5). These data offer important new insights into the mechanisms underlying this complex behavioral process.
β-ocimene and oleic acid have very different chemical properties: β-ocimene is a volatile alkene (boiling point: 65-66°C) and oleic acid is a viscous, mono-unsaturated carboxylic acid (boiling point: 360°C). Both are emitted more strongly in freeze-killed honey bee brood compared to live brood (29), but based on their differences in volatility, we expect them to permeate the brood cell cap at different rates. In a biologically relevant scenario, this spatial diffusion should be necessary for adult workers to detect odor signals evolving under the cap. Since the odor blend induces brood removal most consistently in the back-way odor assays (Figure 2), but not the front-way odor assays (Figure 1), we suggest that β-ocimene and oleic acid may be acting in a cooperative manner when they have to diffuse through the cap. Since our electroantennogram recordings show that there is no synergistic effect at the level of antennal odor detection, we suggest they could instead be cooperating via volatility mechanics. For example, a potential mechanism is that β-ocimene diffuses rapidly and attracts worker visits (as it is already known to do for larval feeding (39)) and after subsequent cell inspection, oleic acid acts as the determinant death cue that stimulates brood removal. In the front-way odor assay, however, the workers are in constant contact with the odorants (since there is no cap acting as a barrier); therefore, oleic acid is readily detectable even in the absence of an attractant.
The back-way odor assay we describe here is the most biologically relevant assay for testing different odors’ abilities to induce hygienic behavior. Unlike other behavioral assays where cells are either uncapped (as in our front-way odor assay) or filled with odor-impregnated brood dummies (25, 26), this assay fully maintains comb integrity, uses real brood, and allows the workers to perform the complete behavior (uncapping and removal). While the odor blend was the most consistently high inducer of brood removal, oleic acid alone also induced significant brood removal for young (pre-pupal) brood, but not post-pupal brood (Figure 2B). Based on our analysis of the background brood odor profile, this could be because of naturally released β-ocimene (Figure 3) interfering with the synthetic odor treatments. Since the younger brood emitted significantly more natural β-ocimene compared to the older brood, the young brood treated with oleic acid was, in a way, also a blend, which could explain why this treatment induced similar removal to the synthetic blend. Very few pre-pupal β-ocimene-treated brood were removed (28%), which is consistent with young brood emitting their own β-ocimene already. Post-pupal β-ocimene-treated brood, which emit very little natural β-ocimene, were removed at higher rates (54%), although this was not statistically significant (p = 0.10).
With a large body of research showing that olfaction is important for hygienic behavior, combined with two odorant binding proteins (OBP16 and OBP18) emerging as protein biomarkers for hygienic behavior, a tempting hypothesis is that the OBPs are aiding the detection of odors associated with disease or death. After showing that hygienic bees remove more β-ocimene- and oleic acid-treated brood compared to non-hygienic bees, we performed ligand binding assays with OBP16 and OBP18 to test if the OBPs linked to hygienic behavior have a high affinity to these necromones (Figure 5). Interestingly, both hexane and phenethyl acetate had a low affinity to both OBPs, which is consistent with both odors being poor inducers of hygienic behavior (as demonstrated in our behavioral assays). β-ocimene, however, displayed strong affinity for OBP16. Oleic acid was a strong ligand for both OBPs, and bound OBP18 the strongest of all those we tested. Since β-ocimene and the odor blend induced significantly higher antennal nerve depolarizations (measured by EAG) than oleic acid, this suggests the worker bees must be very close to the emanating cell (or possibly even contacting the source) to sense it.
Swanson et al. (25) originally identified phenethyl acetate as a strong hygienic behavior-inducing compound emitted from chalkbrood-infected larvae; however, in our experiments, we found that it induces similar levels of hygienic behavior relative to the negative control in both the diluted (P = 0.99) and neat (P = 0.97) odor treatments, which is less than both oleic acid and β-ocimene. In fact, Swanson et al. found that phenethyl acetate induced 40-100% brood removal using just 5% of the odor amount we used. One reason why we did not observe high phenethyl acetate removal rates could simply be because the colonies used by Swanson et al. were from a genetic lineage that was more sensitive to chalkbrood odors than ours.
Based on our data, we cannot yet rule out the possibility that some of the behavioral response toward odor-treated brood was a result of toxicity of the odor itself. In an acute toxicity assay we found that 1 μl of oleic acid was sufficient to cause contact toxicity when dispensed on the abdomen of pupae, halting the development of 40% of treated individuals and inducing a prophenoloxidase immune response after 2 d (Table S2). However, 100% of hexane- and β-ocimene-treated brood developed normally, and the odor blend caused a response midway between these extremes. This toxicity does not mirror the differential hygienic response we observed in the front-way nor the back-way odor addition assays; therefore, it is unlikely to be a main contributor to the hygienic response we see here.
In summary, this data demonstrates for the first time that oleic acid and β-ocimene induce brood removal in honey bees. Hygienic bees respond to the odors more strongly than non-hygienic bees, and the blend induces brood removal most consistently in a biologically realistic brood removal assay. Despite being poorly volatile, oleic acid is still detectable even beneath a brood cell cap, and electrophysiology tests show it only marginally stimulates antennal nerve responses in environmental conditions similar to those inside a hive, suggesting that extremely close proximity is necessary for bees to detect this odor. Both odors are strong ligands for at least one of the OBPs linked to hygienic behavior. Furthermore, oleic acid elicits necrophoretic behavior across phylum Arthropoda (1, 4, 6, 7, 34-36), and these data piece its activity in honey bees into the phylogenetic puzzle. To the best of our knowledge, our data shows for the first time that this ‘death cue’ function is evolutionarily conserved in honey bees, and that oleic acid may be working in concert with β-ocimene as an attractant.
Methods
Honey bee colonies and hygienic testing
We kept honey bee colonies at four separate apiaries in Greater Vancouver, Canada, and performed hygienic testing as previously described (22). All hygienic testing, sampling and odor assays were completed during the summer of 2017.
Front-way odor assays
To perform the front-way odor assays, we retrieved two brood frames from each colony, uncapped patches of brood with tweezers and dispensed 1 μl of odor treatments onto the exposed brood (Figure 1A). We tested the odors β-ocimene, oleic acid, a 1:1 v/v blend of the two, phenethyl acetate (positive control), and hexane (negative control) at concentrations of 100% and 1% (v/v in hexane). Phenethyl acetate was not included in the blend because it is not known to co-occur with the other odors (phenethyl acetate is from chalkbrood, while β-ocimene and oleic acid are associated with freeze-killed brood). For each odor and concentration, we performed two technical replicates (2 patches of 30 brood cells each, one on each frame). We tested the different concentrations on different days. After treating the brood patches with odors, we photographed, traced, and labelled each patch on a transparency and replaced the brood frame in the hive. After 3 h, we returned to the hive and recorded the number of brood removed.
Brood patches were composed of variable developmental stages (5th instar larvae, prepupae and pupae), so in a preliminary test we used the photographs pre- and post-incubation to assess the fraction of each developmental stage that were removed and/or recapped by the workers. Due to variable patch composition, we did not obtain the same number of biological replicates for every developmental stage and odor (see Table 1 for the replicate break-down). Data for 5th instar larvae are not shown because replication was too low, so these data were excluded from subsequent analyses. Brood removal data was analyzed with a three-factor ANOVA (levels: dose, odor, age) followed by a Tukey HSD post-hoc test. All statistical analyses were performed in R unless otherwise specified.
Replicate information for age-related brood removal measurements
In a second preliminary experiment, we confirmed that there was no effect of patch proximity in the front-way odor assay. We varied proximity by testing two patches of brood per colony that were either separated by a single capped cell-width on the same side of a frame, or on different frames with two brood frames located between them. One microliter of oleic acid or β-ocimene was added to the cells of each patch.
To assess the relationship between hygienicity and odor-treated brood removal, we performed the front-way odor assay on 5 hygienic (scoring > 80% on the freeze-killed brood assay) and 5 non-hygienic (scoring< 80%) hives. Hygienic scores ranged from 39% to 100%. These data were analyzed using a three-factor ANOVA (levels: dose, odor, hygienic sore) followed by a Tukey HSD post hoc test. Brood recapping data was derived from the same assays (N = 9 for each odor (data was unavailable for one colony) using a one-way ANOVA (level: odor).
Back-way odor assays (BOAs)
To test the effects of β-ocimene, oleic acid and their 1:1 v/v blend in a more biologically realistic scenario, we developed the BOA assay (Figure 2A). This assay adapts artificial comb cages of the Jenter™ queen rearing system to instead rear worker brood in situ. The Jenter™ set features removable plastic plugs from the rear of the comb – usually used to harvest eggs/larvae for queen rearing-which provide convenient access points for odor addition without damaging the wax brood cell caps or the brood itself.
We conditioned the Jenter™ comb cages by placing them in a colony for several days, allowing the bees to draw out full-height comb cells. We then caged the queens and allowed them sufficient time to populate the combs with eggs (typically overnight). We released the queens and allowed the workers to rear the brood in situ. Once capped, we inspected the brood via the removable plugs to confirm the developmental stage. Through this small posterior window, 5th instar larvae and prepupae are indistinguishable, but pupae are easily recognized by their clearly developed abdomen and hind feet.
We removed the plugs for patches of 9-10 brood cells and dispensed odors (1 μl of neat solutions) onto the brood through the back of the comb and re-plugged each cell. We traced a map of the odor-treated patches and placed the combs in colonies for 20 h to allow workers to detect the odor signals through the cap and respond. We performed five biological replicates for each odor and developmental stage (pre-pupation and post-pupation). Since the 5th instar larvae and pre-pupae are indistinguishable, the ‘pre-pupation’ group contains both stages. After incubation, we removed the comb and counted the number of brood cells from each odor treatment that were removed and/or partially cannibalized. Removal data was analyzed as described above except we used a two-factor ANOVA (levels: odor, age). Due to spontaneous re-queening events, the hygienic scores are not known for all of the colonies in this experiment.
Gas chromatography mass spectrometry (GC-MS)
We performed GC-MS on larvae, prepupae and pupae to detect differences in their natural odor profiles. We collected capped 5th instar larvae, prepupae and pupae from five different colonies and performed solid-phase micro-extraction GC-MS as well as cuticle hexane wash GC-MS exactly as previously described (29). Capped larvae were distinguished from prepupae based on their orientation in the cell (prepupae are elongated while larvae are curled) and the presence or absence of visible feces in the gut system (5th instar larvae defecate before elongating). Spectral data was searched using Mass Hunter Qualitative Analysis software (vB.06.00) and the Wiley Chemical Compound Library (W9N08.L). Peak areas were manually integrated and compound profiles were compared between developmental stages using a one-way ANOVA followed by a Benjamini-Hochberg correction (5% FDR) performed in Perseus (v1.5.5.3). We confirmed the identity of β-ocimene against a synthetic standard (Sigma).
Electroantennography (EAG) recordings
We obtained EAG recordings on bees collected from a single highly hygienic colony (score = 95%) maintained at the University of British Columbia. For ex vivo EAG analysis, we sampled adult nurses from an open brood frame and kept them in a humid incubator (36°C) with access to sucrose water (1:1) until antennal excision. Immediately prior to EAG testing, either the left or right antenna was removed from individual bees (according to a priori randomization) by cutting at the base of the scape. We trimmed the last flagellum segment with dissection scissors, then connected the ends to recording electrodes via glass capillary tubes filled with insect saline solution (210 mM NaCl, 3.1 mM KCl, 10 mM CaCl2, 2.1 mM NaCO3, and 0.1 mM NaH2PO4, pH 7.2) as previously described (45). After data was acquired for the first antenna, the second was collected and the bee was euthanized.
During EAG acquisition, we used a Syntech™ CS-55 stimulus controller to continuously pass humidified air over the antenna and to deliver 1 s pulses of odorized air. To produce odorized air, we cut 1 cm2 slips of No. 1 Whatman filter paper and inserted them into glass Pasteur pipette cartridges. We heated the cartridges to 37°C using a flexible chromatography column heater, at which time we dispensed onto the filter paper 5 μl of distilled water (blank), β-ocimene, oleic acid, or a 1:1 v/v blend of β-ocimene and oleic acid (mix). After allowing 30 s of initial evaporation and slight cooling for the cartridge to reach approximately 33°C, we aimed away from the antenna and passed a 1 s burst of room-temperature air through the pipette before stimulating the antennae with the odors. We then exposed the antennae to a set of 3 consecutive 1 s bursts for each odorant in a randomly-determined order. Between 0.5 and 1 min was allowed between each presentation to allow antennal electrical activity returned to baseline. Blank stimuli (also 3 consecutive 1 s bursts each) were performed at two randomly determined times during acquisition. For each antenna, we subtracted the average blank intensity from the odor EAG intensities, then compared odor groups with a two-way ANOVA (levels: odor, side).
Ligand affinity assays for odorant binding proteins (OBPs)
Recombinant OBP16 and OBP18 were cloned, expressed, and purified exactly as previously described (31). We then used an NPN (N-Phenyl-1-naphthylamine) competitive binding assay to measure affinities of β-ocimene, oleic acid, phenethyl acetate, and hexane (negative control). Binding assays were also conducted as previously described (31), except they were performed in technical duplicate with 2 μM protein, 2 μM NPN, and between 0 and 8 μM of hexane and phenethyl acetate or between 0 and 6 μM of β-ocimene and oleic acid. Dissociation constants of the ligands were calculated from the corresponding IC50 values (concentrations of ligands halving the initial fluorescence value of 1-NPN), using the equation: KD=[IC50]/(1+[1 - NPN]/K(1-NPN))where[1-NPN] is the free concentration of 1-NPN and K(1-NPN) is the dissociation constant of the complex protein/[1-NPN].
Odor toxicity assays
To test the toxicity of the odors, we retrieved 60 purple-eyed, white body pupae and applied 1 μl of neat odor (phenethyl acetate was not included) to the dorsal abdominal area (n = 15 each). We placed the pupae in tissue-lined petri dishes and incubated them at 33°C for 2.5 d. We then scored the pupae for whether their development was halted (i.e. their cuticle did not begin to brown) and whether a prophenoloxidase response had initiated (i.e. the dorsal abdominal region became black). All pupae with halted development also had a prophenoloxidase response.
References




