

Abstract
Hunger evokes stereotypic behaviors that favor the discovery of nutrients. The neural pathways that coordinate internal and external cues to motivate food seeking behaviors are only partly known. Drosophila that are food deprived increase locomotor activity, are more efficient in locating a discrete source of nutrition, and are willing to overcome adversity to obtain food. Here we developed a semi-naturalistic assay and show that two distinct dopaminergic neural circuits regulate food-seeking behaviors. One group, the PAM neurons, functions in food deprived flies while the other functions in well fed flies, and both promote food seeking. These satiation state-dependent circuits converge on dopamine D1 receptor-expressing Kenyon cells of the mushroom body, where neural activity promotes food seeking behavior independent of satiation state. These findings provide evidence for active food seeking in well-fed flies that is separable from hunger-driven seeking.
Introduction
The neural mechanisms that regulate feeding motivation are ancient, fundamental for survival, and under complex regulation, and yet they remain partially defined and understood. Feeding motivation is classically divided into pre-ingestive and consummatory phases1,2. In the pre-ingestive phase, nutritional deficits cause release of hormonal signals that act on the brain to bias behavioral states towards seeking food, including heightened attention to food-related environmental cues, increased locomotion, and suppression of incompatible behaviors such as sleep. Once a nutritional source is encountered, homeostatic mechanisms in concert with sensory and nutrient detectors cause a cessation of locomotion and engagement of motor programs for food intake. Both pre-ingestive and consummatory phase behaviors are motivated and goal-directed. However, the goals and the conditions for their completion are different, suggesting that the neural circuits controlling each phase are also different. Defining the neural mechanisms of feeding motivation is important in part because the dysregulation of feeding behavior is intimately tied to obesity and eating disorders, as well as to other pathological alterations of motivation, including drug addiction3,4.
Simpler organisms such as Drosophila hold promise for uncovering the neural circuit mechanisms for motivated feeding behavior. In Drosophila, feeding behavior studies have focused mostly on the consummatory phase, and have revealed satiation state-dependent effects on sensory5–7, motor8–10, and central processing of feeding11–14. Appetitive associative conditioning with feeding has defined detailed neural circuits implicated in reward learning15–18. Drosophila behavioral studies of the pre-ingestive phase have focused mostly on sensory perception of appetitive stimuli, including odor tracking, satiation state-dependent olfactory acuity, and search strategies19–23. Here we report the development of a semi-naturalistic assay for innate pre-ingestive behaviors in Drosophila, in which flies search in an open arena for a discrete source of food. Semi-naturalistic assays may offer advantages over task-specific assays in defining how complex information is processed to drive behavior. We demonstrate specific roles for distinct dopaminergic neural circuits in the well-fed and food-deprived states for regulating food seeking behavior.
Results
Parametric Analysis of Drosophila Food Seeking Behavior
We developed a semi-naturalistic paradigm to measure various aspects of food seeking in freely behaving flies. Flies placed into a translucent arena (Fig. 1A) are tracked with a video camera (Fig. 1B). After a set acclimation period, a small volume of food is introduced at the center of the arena. Increasing lengths of food deprivation (wet starvation with water only) increased the number of flies in contact with the food, the food occupancy rate (Fig. 1C). Locomotor speed in the absence of food increased with increasing lengths of food deprivation time (Fig. 1D). Introduction of food into the arena rapidly decreased the locomotor speed of food deprived flies that were not in contact with the food source. Food intake scaled with deprivation time, as measured in a separate assay that minimizes seeking time (Fig. 1E). For subsequent experiments, ‘food-deprived’ indicates 16–20 hr of a water only diet, unless otherwise noted.

Food deprivation effect on food seeking behavior. A. Two-sided chamber for food seeking assays. Flies and 200 ul of cornmeal molasses food on Parafilm placed in each chamber via sliding side doors. The chamber is lit from below. Fly locomotion is recorded from above. B. 10 sec locomotor traces of 20 flies each filmed soon after addition of food (yellow dot). C. Left: The percent of flies on food over time for a food deprivation time course. Right, food occupancy averaged at 25–30 min. P<0.0001, ANOVA/Bonferroni comparison to 0 hr. n=17–18 groups. D. Locomotor speed. Left, speed at 20 min of acclimation, without food. Right, speed averaged over 0–10 min after food introduction. P=0.0091 no food, P=0.0066 food, ANOVA/Bonferroni compared to 0 hr. n=9–15 groups. E. Intake with increasing food deprivation time. P<0.0001, ANOVA/Bonferroni comparison to 0 hr. n=9 groups. *P<0.05, **P<0.01.
Sensory and Nutritional Inputs to Food Seeking
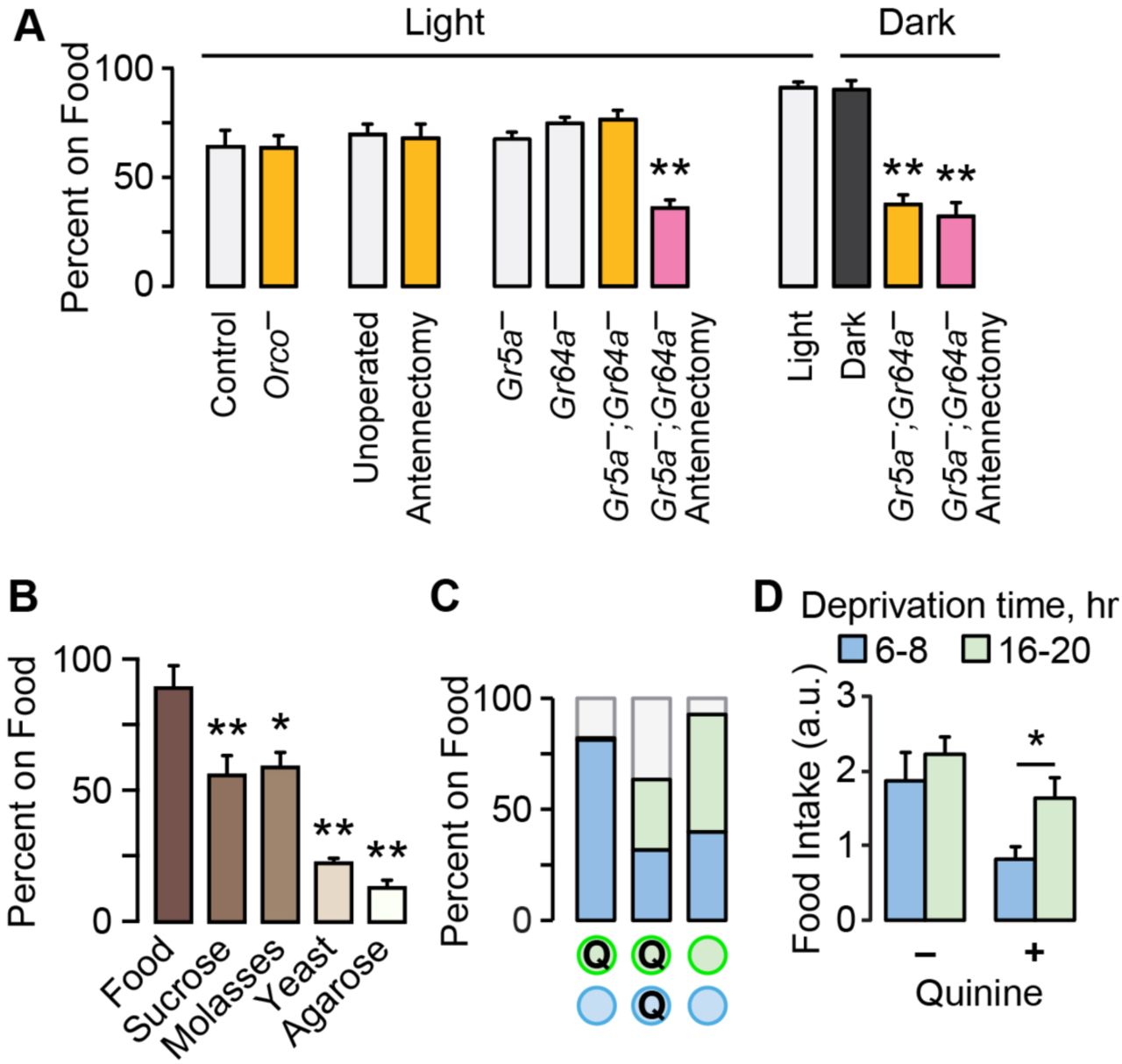
We tested for the role of olfaction, taste, and vision in food seeking in food-deprived flies (Fig. 2A). Neither genetic nor surgical ablation of food odor-detecting neurons - olfactory coreceptor mutant Orco1 or removal of the third antennal segment - affected food seeking24,25.

Environmental and sensory information in food seeking. A. Food occupancy following sensory ablations in 16–20 hr food deprived flies. Antennectomy is surgical removal of the third antennal segment. Orco− flies lack the Orco olfactory coreceptor; Gr5a− and Gr64a− are taste receptor mutants. P<0.0001 for both Light and Dark, ANOVA/Bonferroni compared to control, n=8–12 groups. B. Occupancy of 16–20 hr food deprived flies to agarose with the indicated food component. P<0.0001, ANOVA/Bonferroni comparison to Food. n=4–5 groups. C. Two-choice tests with unadulterated (open circles) and 10 mM quinine food (Q). n=5 groups. D. Flies consumed greater quantities of quinine food (3 mM) when food-deprived for 16–20 hr (long) versus 6–8 hr (short). P=0.0251, Mann Whitney test, n=12. *P<0.05, **P<0.01. See also Figure S1.
Similarly, flies lacking a subset of sugar sensing taste receptors had no effect on food seeking for sucrose. These experiments suggested that flies may use more than one sensory modality when seeking nearby food. Flies with both ablated antennae and taste receptor mutations showed decreased food occupancy, suggesting coordination between olfaction and taste. Food seeking remained robust in complete darkness. However, taste receptor mutant flies showed reduced food occupancy in total darkness, and additionally removing olfactory input did not further reduce occupancy. These results indicate that flies use a combination of taste, olfactory, and visual cues to find and occupy a discrete food source.
Flies may seek one or more food constituents. Food deprived flies were most attracted to complete food, then sugars, and then protein (Fig. 2B). When given a binary choice, flies preferred complete food over any other option, and preferred sugars over yeast (Supplementary Fig. S1). Similarly, flies occupied sweet and nutritious sucrose more than either sweet-only sucralose or nutritious-only sorbitol (Supplementary Fig. S1). Finally, nutrition appears to be important for switching the locomotor state of food deprived flies: flies slowed in the presence of sucrose or D-glucose, whereas they did not in the presence of sucralose or L-glucose (Fig. S1D). These findings suggest that sweetness is a mechanism that captures flies on a food source, and that nutritional content of the food source is important for fully switching flies from the pre-ingestive to consummatory phase of food seeking.
A characteristic of motivated behavior is the willingness to overcome negative consequences. Flies will eat substantially less food when it is adulterated with bitter compounds, and this scales with satiation state13. In a binary choice competition, food deprived flies occupied quinine-containing food, but only if there was no better choice (Fig. 2C). Furthermore, food intake was less suppressed by quinine with longer deprivation (Fig. 2D). We used a sucrose food source for all subsequent experiments.
Role of Dopaminergic Neurons in Food Seeking
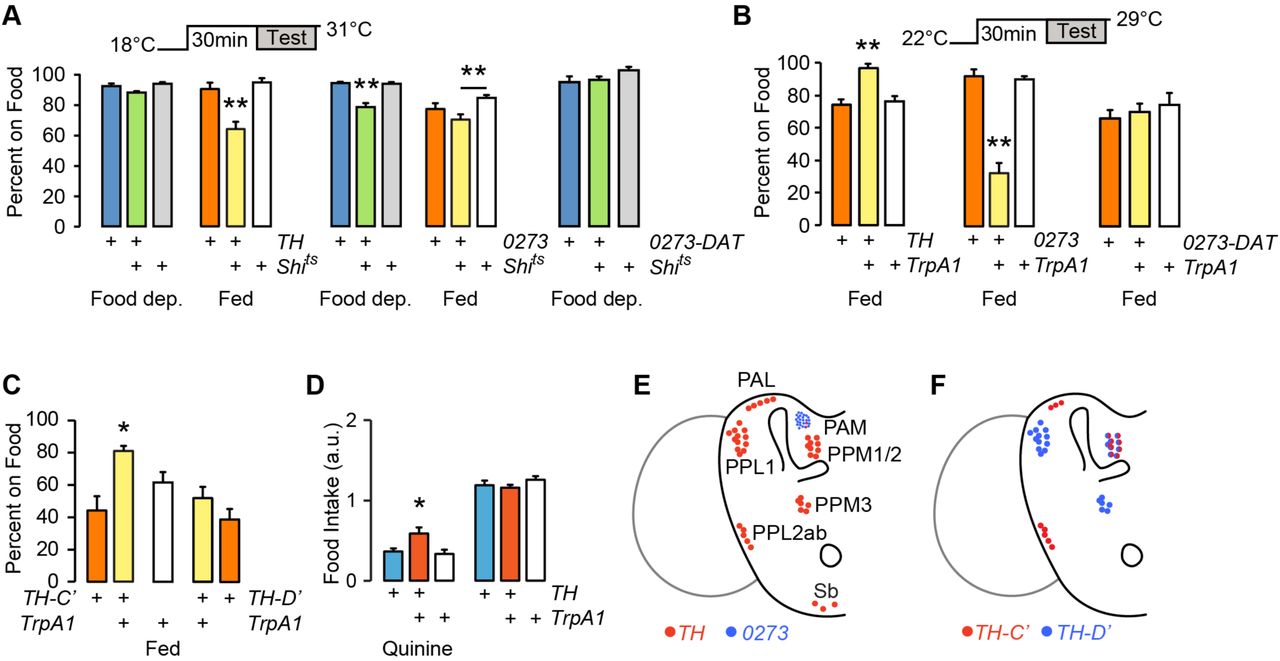
Dopaminergic neural circuits are critical for motivation, reward, and food seeking in mammals, and for many similar functions in flies26. To test the role of dopamine in food seeking in flies, we acutely inactivated and activated subsets of dopamine neurons in fed and food-de-prived flies and assessed occupancy of sucrose. Dopamine neurons group into several discrete anatomical and functional clusters in the adult fly brain (Fig. 3E). TH-Gal4 labels most dopamine neuron clusters, but is largely absent from the PAM (protocerebral anterior medial) cluster of approximately 100 dopamine neurons. 0273-Gal4 labels most or all dopamine neurons in the PAM cluster but not in other dopamine neurons. Acutely blocking transmitter release in TH-Gal4 neurons with the temperature-sensitive dynamin Shibire (Shits) had no effect on food occupancy in food deprived animals (Fig. 3A). Food occupancy was decreased when TH-Gal4 neurons were transiently inactivated in fed animals. There was no effect of inactivation on locomotor activity (Supplementary Fig. S2). Conversely, inactivation of 0273-Gal4 neurons specifically decreased food occupancy in food deprived animals. DAT-Gal80 (also named R58E02-Gal80) expresses the GAL4 inhibitor GAL80 exclusively in PAM neurons: DAT-Gal80 blocked the 0273>Shits food occupancy phenotype (Fig. 3A). Finally, chemical depletion of dopamine with 3-iodotyrosine also decreased food occupancy, indicating that dopamine is a neurotransmitter for food seeking (Supplementary Fig. S2). Thus, dopamine neurons in the TH-Gal4 pattern promote food occupancy in fed animals, and PAM dopamine neurons in the 0273-Gal4 pattern promote food occupancy in food deprived animals.
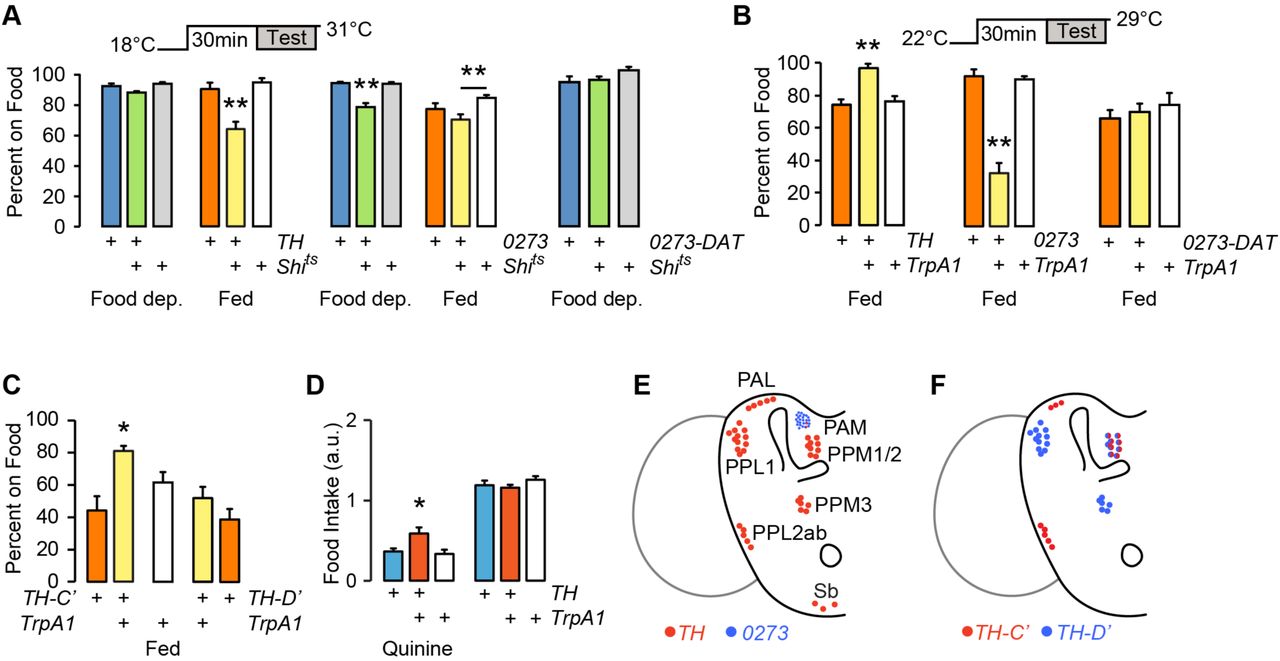
Satiation state-dependent effects of dopamine neuron activity on food seeking. A. Acute inactivation of dopamine neurons with Shibirets (Shits), food occupancy in fed and 16–20 hr food-deprived flies. P=0.0012 ANOVA/Tukey’s, n=8–11 groups with TH-Gal4. P=0.0001 Kruskal-Wallis/Dunn’s, n=8–10 groups food deprived; P=0.0139 ANOVA/Tukey’s, n=8–9 groups fed, with 0273-Gal4. 0273-DAT: 0273-Gal4 with R58E02-Gal80 to specifically block GAL4 activity in the PAM cluster dopamine neurons. n=6 groups. B. Acute activation of dopamine neurons in fed flies, food occupancy. P=0.0002, ANOVA/Tukey’s, n=8–11 groups with TH-Gal4. P=0.0002, Kruskal-Wallis/Dunn’s, n=8 groups with 0273-Gal4. 0273-DAT: n=8 groups. C. Acute activation of subsets of TH-Gal4 neurons, food occupancy in fed flies. P=0.0002, ANOVA/Tukey’s, n=8–11 groups. D. Food intake in 4–6 hr food-deprived flies. P=0.0053, ANOVA/Tukey’s, n=15–19 groups. E. Dopamine neuron clusters in the adult brain that express TH-Gal4 and 0273-Gal4. F. Dopamine neurons that express TH-C’-Gal4 and TH-D’-Gal4. *P<0.05, **P<0.01. See also Figure S2.
To test if dopamine neurons are permissive or instructive, we acutely activated them using the temperature-sensitive cation channel TrpA1. Consistent with an instructive role, activating TH-Gal4 neurons in fed flies increased food occupancy (Fig. 3C). Fed 0273>TrpA1 flies showed a marked decrease in food occupancy, and this was due to PAM dopaminergic activation in the 0273-Gal4 pattern. To identify the relevant neurons in the TH-Gal4 pattern, we used transgenes that differentially label specific clusters of dopamine neurons (Fig. 3F)17. Activation of patterns that included the PPL2ab, PPM2, and PAL, but not the PPL1, PPM1, or PPM3 dopamine neuron clusters increased food occupancy in fed flies (Fig. 3C). To test if the identified dopaminergic neurons may regulate feeding motivation, we activated TH-Gal4 neurons in mildly (4 hr) food-deprived flies. Under these conditions, activation of TH-Gal4 neurons specifically increased consumption of quinine adulterated food (Fig. 3D). Taken together, these experiments are consistent with dual roles for dopamine in food-seeking behavior: a PAM dopamine neuron-mediated promotion of food seeking in the food-deprived state, and a TH-Gal4 dopamine neuron-mediated promotion of food seeking in the fed state. In the fed state, PAM dopamine neurons can block food seeking.
Dopamine Receptor Regulation of Food Seeking
Dop1R1 encodes a D1-like dopamine receptor that functions in motivation-related behaviors, including arousal state, drug reward, and learning and memory27–29. We tested flies with strongly reduced expression of Dop1R1 for food seeking behaviors. Food-deprived Dop1R1 mutant flies were hyperactive and appeared to ignore food (Fig. 4A). Moreover, Dop1R1 mutant food occupancy was reduced when fed or food deprived (Fig. 4B). Loss of the dopamine D2-like receptor D2R did not affect food occupancy, but restored normal food occupancy to Dop1R1 mutants. These data suggest that Dop1R1 promotes food seeking, and that an opposite role for D2R is uncovered in the absence of Dop1R1. Food intake was unaffected in food-deprived flies of these genotypes (Supplementary Fig. S3).

Dopamine receptor-expressing neurons in the mushroom body control food seeking. A. Locomotor traces of food-deprived flies 5 min after addition of food. Dop1R1 mutant f02676 vs. Berlin genetic background control. B. Food occupancy for the indicated genotypes that were fed or food deprived. t-test P=0.0492 fed (n=16–20 groups), P=0.001 food deprived (n=16–20 groups). D2R: the loss-of-function mutation f06521. C. Location of Dop1R1 enhancer fragments. D. Genetic rescue of Dop1R1 mutant food occupancy in 16–20 hr food deprived animals. Dop1R1-Gal4 strains (blue) were made heterozygous in f02676 homozygotes (rescuing configuration, green). P<0.0001 ANOVA/Bonferroni’s comparison to f02676, n=8–16 groups. E. Inclusion of MB-Gal80, preventing GAL4 activity in the mushroom bodies blocks B12 rescue. P<0.0001 ANOVA/Tukey’s, n=10–19 groups. F-H. Expression pattern of Dop1R1-Gal4 strains (CD8-GFP, green), and bruchpilot (magenta) to show the synaptic neuropil. I. Acute silencing of B12 Dop1R1-Gal4 neurons with Shits, food occupancy, food deprived and fed. Food deprived: P<0.0001 Kruskal-Wallis/Dunn’s, n=4 groups. Fed: P=0.0002 Kruskal-Wallis/Dunn’s, n=7–8 groups. J. Addition of MB-Gal80 in B12 Dop1R1-Gal4>Shits fed flies, food occupancy. P<0.0001 Kruskal-Wallis/Dunn’s, n=6–10 groups. K. Activation of B12 Dop1R1-Gal4 neurons in fed flies increased food occupancy. P=0.0054, ANOVA/Tukey’s, n=7–9 groups. *P<0.05, **P<0.01. See also Figure S3.
The Mushroom Bodies Promote Food Seeking Independent of Satiation State
We performed genetic rescue experiments to ask where Dop1R1 functions for food seeking in food deprived flies. To bias the rescue towards functionally relevant brain regions, we utilized Dop1R1-Gal4 strains that expressed GAL4 under the control of short non-coding genomic DNA fragments cloned from the Dop1R1 locus (Fig. 4C). Food occupancy was partially rescued when Dop1R1 was expressed with three different Dop1R1-Gal4 strains in food-deprived Dop1R1 mutants: B07, B12, and C02 (Fig. 4E). Anatomical analysis of the expression patterns for the rescuing Dop1R1-Gal4 drivers revealed expression overlap. In the B12 and C02 strains, the mushroom bodies were prominently labeled, as were regions of the central complex, including the fan-shaped body and protocerebral bridge (Fig. 4F,G). The B07 strain prominently labeled the ellipsoid body of the central complex (Fig. 4H). We failed to rescue Dop1R1 mutant food occupancy using GAL4 drivers that label the ellipsoid body, fan-shaped body, or the protocerebral bridge (not shown). By contrast, decreasing GAL4 activity with mushroom body-specific expression of GAL80 eliminated B12 rescue of the Dop1R1 mutant food occupancy phenotypes (Fig. 4E). Moreover, restoring Dop1R1 with the mushroom body-specific driver MB247-Gal4 rescued Dop1R1 food seeking (Supplementary Fig. S3). Thus, Dop1R1 expression in the mushroom bodies is sufficient to promote food seeking in food deprived animals.
We next tested the role of neurotransmission in Dop1R1-expressing mushroom body neurons in food seeking. Similar to loss of Dop1R1, acute blockade of synaptic output in B12 neurons decreased food occupancy in both fed and food-deprived flies (Fig. 4I). Importantly, this effect also localized to the mushroom bodies (Fig. 4J). B12>Shits flies also showed reduced locomotion, however this phenotype persisted when the mushroom body neurons were subtracted from B12 (Supplementary Fig. S3), suggesting that distinct Dop1R1 neurons control food occupancy and locomotion. Finally, acute activation of B12 neurons in fed flies increased food occupancy (Fig. 4K). Taken together, these results indicate that the activity of Dop1R1-expressing mushroom body neurons promote motivated food seeking in both the fed and food-deprived state.
Discussion
Distinct dopaminergic circuitry promotes food seeking under well fed and food deprived conditions. Dopamine neurons in the TH-C’ pattern promote seeking in well fed flies, and dopamine neurons in the PAM cluster promote seeking in food deprived flies. The PAM neurons likely function directly upstream of Dop1R1-expressing neurons of the mushroom body that promote food seeking in both the fed and food-deprived states. These circuits function in food seeking under semi-naturalistic conditions, where flies can freely perform many steps of food seeking behavior. Understanding how these dopaminergic circuits contribute to discrete steps of feeding behavior, from local search through to repletion and disengagement from a food source, will help define how motivational states transition from task to task.
Roles of Dopamine in Appetitive Behaviors
Dopaminergic neurons are critical for many appetitive and aversive behavioral responses across animal species. Dopamine may act as a salience, arousal, or attention signal that gives importance to specific valence information arriving from other circuit elements26,30,31. In rodents, genetic, pharmacological, and lesioning studies indicate that striatal dopaminergic pathways can selectively function in the pre-ingestive phase to promote food seeking30,32,33. We found that acute activation of dopamine neurons in fed flies increased food occupancy, yet it did not cause increased food intake. Likewise, genetic elimination of the Dop1R1 receptor decreased food occupancy without affecting food intake. In contrast, inactivation of Dop1R1 receptor neurons decreased food intake in the food-deprived state, possibly reflecting their key role in integrating sensory and internal state information. These findings suggest that dopaminergic pathways promote pre-ingestive food seeking. However, the role of dopamine is more complex. For example, the PAM dopamine neurons are activated by ingestion of sugar, and their activation is greater in food-deprived flies, indicating that dopaminergic neurons are engaged during the consummatory phase of feeding, and they may be sensitized to responding to input during the pre-ingestive phase17.
Prior studies assigned dopamine to particular aspects of feeding behavior and also to motor functions that are critical to feeding14,23. In particular, dopamine neurons in the TH-Gal4 pattern are implicated in controlling motor output: TH-Gal4 neuron hyperpolarization, blocking synaptic input, interferes with motor performance and aspects of food seeking behavior in food deprived flies23,34. We did not detect differences in unstimulated motor activity or in the magnitude of an olfactory-stimulated startle response when we blocked synaptic output from TH-Gal4 neurons, indicating that flies exhibited grossly normal motor behavior in our as-say35. The differences in observed phenotypes may reflect the multifunctional roles of TH-Gal4 dopamine neurons that are revealed by specific types of manipulation.
Which dopamine neurons are responsible for food seeking? In well-fed flies, neurons in the TH-C’ pattern promote seeking. This pattern includes dopamine neurons in the PAL, PPM2, and PPL2 clusters, and this group of neurons was previously shown to promote female egg-laying preference on sucrose17,36. The neurons in these clusters project to many specific regions of the brain, and their individual functions remain largely unknown. One exception is that individual neurons in the PPM2 cluster, the DA-WED neurons, support protein consumption preference in protein deprived flies14. The DA-WED neurons synapse to Dop1R1 neurons in the B03 pattern, which did not support rescue of food seeking in our experiments. These findings argue that there are distinct dopaminergic circuits in the TH-C’ pattern that control different forms of nutrient seeking. The PAM neurons are also heterogeneous, sending projections that tile to well-defined regions of the mushroom body and to regions of the protocerebrum. Specific subsets of PAM neurons that are included in the 0273-Gal4 pattern have been implicated in various forms of appetitive learning and memory, however their inactivation did not impact food seeking in food deprived flies (not shown)15,17,37–39. This suggests that there may be further segregation of PAM dopamine neuron function, possibly according to innate and learned appetitive responses.
Sensory Tuning of Food Seeking Motivation
Appetitive olfactory cues such as those emitted from palatable food elicit approach and can activate neurons important for feeding. Olfactory receptor neurons that respond to appetitive odors increase sensitivity through the actions of the neuropeptides sNPF and SIFamide 21,40. Further, neurons that release the neuropeptide NPF are activated to a greater extent in response to food odors in food-deprived flies; their activation promotes and inactivation inhibits odor attraction41. In well-fed larvae, the attractive odor pentyl acetate increases food intake through the actions of NPF and dopamine11. Therefore, food-related odors not only elicit approach behavior in a satiation state dependent manner, but also increase the activity of neurons expressing neuropeptides that regulate feeding behavior. Our results indicate that, under semi-naturalistic conditions, olfaction is important but apparently not crucial for food seeking in food-deprived flies: neither surgical nor genetic ablation of olfaction decreased food occupancy, and its role was only revealed by simultaneous partial ablation of taste responses. Further, flies were efficient in seeking odorless sucrose. Taken together, olfaction, hygrosensation, visual cues, and taste responses likely act in concert with internal cues to set the intensity of food seeking when freely behaving flies are in close proximity to a food source.
Methods
Strains and Culturing
All strains were outcrossed for five generations to the Berlin genetic background prior to behavioral testing. Flies were raised on standard food containing agar (1.2% w/v), cornmeal (6.75% w/v), molasses (9% v/v), and yeast (1.7% w/v) at 25°C and 70% humidity unless otherwise indicated. Dop1R1-Gal4 (R72B03, R72B05, R72B06, R72B07, R72B09, R72B12, R72C01, R72C02) strains were generated by the FlyLight project39, TH-C’-Gal4 and TH-D’-Gal4 were from Mark Wu, Gr5aEP-5 and Gr64a1 were from Anupama Dahanukar, 0273-Gal4 from Daryl Gohl and Thomas Clandinin, MB-Gal80 from Hiro-mu Tanimoto, Orco1 from Leslie Vosshall, and others from the Bloomington Stock Center.
Behavioral Measurements
Groups of 21 males were collected 1–2 days prior to the experiment. For food deprivation flies were placed into empty culture vials containing water saturated Whatman filter paper. For 3-iodotyrosine treatment flies were cultured for 30 hr with 5% sucrose/2% yeast/10 mg/mL 3-iodotyrosine (3IY), and treated an additional 16 hr with 3IY in water for food deprivation. Standard fly food was used for all experiments except where indicated. Thin-walled Plexiglas behavioral chambers were designed with two side-by-side arenas, each arena measuring 45x75x10 mm, or 85×135×10 mm for experiments with Shibirets. Chambers were designed and built by IO Rodeo (Pasadena, CA). Flies were filmed from above at 10 fps with the arena placed on white light LED panel (Edmund Optics). Filmed flies were tracked with customized software40. For food occupancy, the number of flies off food was subtracted from the total number of flies and divided by total number of flies. Percent on food was calculated as the average of the last two measured time points. Locomotor activity was the average speed of all flies in 20 sec bins.
To measure food intake, 5 ml standard fly food with 2% erioglaucine (Sigma) with or without 3mM quinine was striped onto 1/4 of the inner surface of a wide fly vial, and condensation removed. 30–50 flies were introduced and the vial laid on its side so that the food edge was at the apex. After 30 min, the flies were homogenized in a volume adjusted to the number of flies and consumption was determined spectrophotometrically.
Statistical measurements were made with Prism 6.0 (GraphPad). Error bars are the SEM. Data is available upon request.
Author Contributions
DL, DSF, and FWW conceived of and carried out the experiments, and analyzed the results. FWW wrote the paper.
Acknowledgements
We thank the members of the laboratories of Fred Wolf and Michael Cleary for advice, and Daryl Gohl and Thomas Clandinin for unpublished strains. This work was supported by grants from the NIH (AA018799), The Hellman Fellowship Fund, and the University of California, Merced. The authors declare no competing financial interests.
Footnotes
↵* co-first authors





{kind=link}
{kind=link}
{kind=link}
{kind=link}