

Abstract
Learning is a general mechanism of adaptive behavioural plasticity whose benefits and costs depend on the environment. Thus, seasonal oscillations in temperate environments between winter and summer might produce cyclical selection pressures that would drive rapid evolution of learning performance in multivoltine populations. To test this hypothesis, we investigated the evolutionary dynamics of learning ability over this rapid seasonal timescale in a natural population of Drosophila melanogaster. Associative learning was tested in common garden-raised flies collected from nature in the spring and fall over three consecutive years. The spring flies consistently learned better than fall flies, revealing seasonal evolution of improved learning performance in nature. Fecundity showed the opposite seasonal pattern, suggesting a trade-off between learning and reproduction. This trade-off also held within population: more fecund individual females learned less well. This trade-off is mediated at least in part by natural polymorphism in the RNA binding protein couch potato (cpo), with a genotype favoured during summer showing poorer learning performance and higher fecundity than a genotype favoured over winter. Thus, seasonal environments can drive rapid cyclical evolution of learning performance, but the evolutionary dynamics may be driven by trade-offs generated by pleiotropic effects of causative alleles selected for other reasons.
1. Introduction
Learning is a powerful mechanism of plasticity that allows animals to behaviourally respond to changing environments without genetically-based evolutionary change [1,2]. Learning plays an important role in many aspects of animal biology including foraging, spatial orientation, predator avoidance, aggression, social interactions, and sexual behaviour [1,3–7], and thus is likely to have manifold effects on fitness.
The ability to learn is itself a product of evolution, but we know very little about how rapidly characteristics of the learning process evolve in nature. Examples of genetic differences in learning performance have been reported between closely related species [8,9] and between allopatric populations within a species [10,11], but the timescale over which these differences have evolved is unknown. Laboratory selection experiments in a range of taxa indicate that natural populations have copious standing genetic variation for learning ability and that learning performance can improve rapidly under direct selection [12–16]. However, selection on learning in nature may be indirect and mediated by the fitness consequences of the many behaviours it modifies. An environmental change that results in an increased advantage of learning in the context of one behaviour (e.g., foraging) might have no effect on, or even decrease, its benefits in a different context (e.g., predator avoidance), thus dampening net selection on learning.
Here we test if learning performance evolves rapidly over an annual timescale in a natural population of D. melanogaster. This multivoltine species is a promising system to study rapid evolution because each generation experiences different conditions as seasons progress. The alternating conditions between different selection regimes of winter and summer drive rapid cyclical evolution of Drosophila melanogaster life history, stress resistance and immunity traits [17–19]. One might predict that increased ability to learn should be favoured in the summer because learning modulates behaviours that are key to fitness in the summer, such as foraging, pathogen avoidance, and sexual and social interactions [20–25]. However, learning might also contribute to behaviours that are important for overwintering survival (e.g., shelter identification, foraging, pathogen avoidance). Finally, evolutionary dynamics of learning ability may be driven by selection on genetically correlated traits. Such genetic correlations may reflect the high energetic costs of the central nervous system [26,27], leading to trade-offs with other fitness traits. Such trade-offs have been reported between learning and reproduction in Pieris rapae [28] and Drosophila [29], as well as between learning and stress resistance in Drosophila [30,31]. Thus, seasonal evolution in learning ability could be driven by indirect effects of selection on correlated traits – particularly on fecundity in the summer or stress tolerance in the winter.
We assessed aversive and appetitive learning in common-garden-raised flies derived from spring and fall collections made at the same locality in the wild over three consecutive years. We found that learning performance was consistently higher in the post-winter (i.e., spring) collections compared to the post-summer collections in the fall. A reproductive cost of learning ability was indicated by an inverse relationship with fecundity between seasonal populations and at an individual level.
Next, we tested if natural variants of the pleiotropic RNA-binding protein couch potato (cpo) contribute to the relationship between learning and fecundity. Fecundity and reproductive dormancy differ among natural variants in cpo that change in frequency with latitude and season [32–35]. We hypothesized that cpo may be involved in learning because it is highly expressed in the nervous system including the centre of associative learning, the mushroom body [36–38]. We found that learning performance and fecundity in flies carrying natural cpo genotypes that are more common in spring versus fall paralleled the seasonal differences in learning within the natural population. These results suggest that rapid fluctuating evolution of learning ability in wild Drosophila is at least in part driven by pleiotropic effects of cpo polymorphism.
2. Methods
(a) Seasonal populations derived from nature
The evolution of learning differences over seasonal time was assessed by comparing seasonal reconstructed outbred populations (ROP) from isofemale lines, hereafter referred to as spring and fall ROPs. Gravid females aspirated off decaying fruit in the spring (June) and fall (November) at Linvilla Orchards in Media, PA (39.9°N, 75.4°W), across three consecutive years (2012-2014) were used to establish isofemale lines that were maintained in standard laboratory conditions (25°C, 12L:12D on cornmeal-molasses medium) on a four-week transfer cycle. Populations representing each collection were constructed after all collections were complete using 40 isofemale lines per collection. The populations were subsequently maintained in common garden culture for more than 10 non-overlapping generations. Systematic differences in traits between spring and fall ROPs assessed in the common garden can be attributed to genetic differences of the spring and fall allele frequencies.
(b) cpo recombinant outbred populations
Three intronic SNPs in cpo are associated with climatic adaptation: 3R: 17964408, 3R: 17965558, 3R:17967866 (D. melanogaster reference genome v.6). The temperate genotype (cpoTTA) is more common at high latitudes and in the spring while the tropical genotype (cpoCGT) is more common at low latitudes and in the fall [35]. The polymorphism at 3R:17967866 was identified as a non-synonymous coding change in exon 5 with functional effects on reproductive dormancy and other life history traits [33,37], however, its specific molecular function has been re-evaluated as it not identified in the modENCODE classification of functional elements [39]. Learning performance of the genotypes was assessed using ROPs that were each fixed for one genotype with a randomized genetic background [40]. ROPs for cpoTTA and cpoCGT were constructed using nine independent, homozygous lines from the Drosophila Genetics Reference Panel [35,41]. Ten gravid females from each line were permitted to lay eggs for 48h; after at least 10 non-overlapping generations of recombination with the other lines containing the same genotype each ROP was fixed for either cpoTTA or cpoCGT, and any linked loci, in a heterogeneous, outbred background.
(c) Aversive shock learning in the laboratory
We quantified the performance of flies in an aversive olfactory learning assay that assessed their ability to remember and avoid an odour previously associated with mechanical shock [30]. Flies were reared in common laboratory conditions (25°C, 12L:12D) at standardized density of 100 eggs per vial, sorted into groups by sex under light CO2 anaesthesia 24 hour prior to the learning assessment, and then assayed at 3-5 days of age. Single-sex groups of 30 flies were conditioned to associate an odorant (the conditioned stimulus, CS+) with a mechanical shock. Three cycles of conditioning were conducted, each consisting of the sequence: 30 seconds exposure to CS+ paired with mechanical shock (pulsating 1 second every 5 seconds), 60 seconds break of humid air, 30 seconds exposure to another odorant (CS−) with no shock, and 60 seconds humid air. Methylcyclohexanol (MCH, 800 uL/L) and 3-octanol (OCT, 600 uL/L) were used as odorants; half of the fly groups were conditioned to avoid each odorant to account for innate preference for either odorant (defining two conditioning directions). After a one-hour memory retention interval we tested odorant preference of the conditioned flies by giving them 60 seconds to choose between the odorants in an elevator T-maze. The cpo genotypes were also assessed for immediate response (5-minute retention) and long-term memory (24-hour retention).
To exclude impairment in odour perception or differences in innate preference we performed two additional controls. Controls for absolute preference subjected flies to mechanical shock cycle without odorant before presenting a choice between a single odorant and air in the T-maze. Controls for relative preference tested innate odour preference in naïve flies by giving a choice between the two odorants in the T-maze.
All statistical analyses were performed using the R software (R Core Team, version R 3.2.2). To analyse learning performance, we modelled proportion of flies selecting the odour not previously associated with shock (CS−) using generalized mixed models with logit link and binomial error distribution, fitted by maximum likelihood (package lme4, Bates et al. 2014). To test for the significance of main effects and interactions, we used likelihood ratio test from the package afex [43] and functional analysis of variance (ANOVA, Fox and Weisberg 2011). Flies that chose no odour (i.e., remained in the centre of the T-maze) were excluded from analysis. For illustration purposes, the proportion of flies learning was rescaled as 2 × proportion – 1 to make it range from – 1 to 1 with 0 indicating no learning. We used the following models where ROP (season or cpo genotype), year, sex, and conditioning treatment are fixed effects and block is a random effect grouping samples tested at the same time:
Natural populations:
Proportion = Year + ROP + Sex + Conditioning treatment + Year:Conditioning treatment + Sex:Conditioning treatment + Block
cpo genotypes:
Proportion = ROP + Sex + Conditioning treatment + Retention Interval + ROP:Conditioning treatment + ROP:Retention Interval + Block
(d) Appetitive learning in a natural environment
The appetitive conditioning learning assays tested the flies’ ability to remember and prefer the flavour previously associated with food, with preference assessed as number of flies caught in traps of two alternative flavours. It was adapted from [15,24], but implemented in a natural setting in outdoor cages over two replicate days (August 30 and 31, 2016).
Flies were reared in standard laboratory (25°C, 12L:12D) conditions at controlled density (approximately 200 eggs per bottle), and 3-day cohorts were sorted into groups of 50 by sex. Flies were marked with fluorescent powder using four colours (Blue 1162B, Red 1162R, Yellow 1162Y and White 1162W; Bioquip, Rancho Dominguez, CA) according to learning treatment and source population. A pinch of powder was added to an empty vial before adding the flies and gently rolling the vial until the flies were coated. Flies were then transferred to a non-nutritional agarose for 12 hours to recover from the marking process and to become starved, so they were more motivated to feed. The flies were subsequently exposed to either strawberry or apple flavoured food (two directions of conditioning) for an 8-hour training period, followed by a 4-hour retention period on a fresh agar substrate, before being released into the outdoor cages for testing. The use of four marking colours permitted the simultaneous release in the same cage of flies from the two genotypes being tested (either spring and fall or cpoTTA and cpoCGT) conditioned with either flavour (400 flies per conditioning direction, gentoype season and sex). The cages (0.6 × 0.6 × 1.8 m), made of mesh and with plant bedding covering the ground, were placed underneath a peach tree. Four pairs of strawberry and apple unidirectional traps were dispersed around the cage. Flies trapped in each trap within 14 hours after release were sexed and attributed to season and conditioning direction based on colouring. All of the trapped flies showed traces of the powder; thus, marking was effective, and no flies infiltrated the experiment from outside.
Learning performance was quantified as proportion of flies selecting traps flavoured with the same flavour that they had been previously exposed to during the conditioning. The same statistical analyses were performed as on the aversive conditioning, with year, ROP (season or cpo genotype), sex, and conditioning treatment are fixed effects and trap and day are random effects.
Natural populations:
Proportion = Year + ROP + Sex + Conditioning treatment + Year:Conditioning treatment + Sex:Conditioning treatment + Trap + Day
cpo genotypes:
Proportion = ROP + Sex + Conditioning treatment + ROP:Sex + Trap + Day
(e) Fecundity
Fecundity of the reconstructed seasonal populations was measured by placing 25 virgin females from each reconstructed seasonal population into culture bottles [40] with 25 males from a standardized stock (BDSC-3605). Food plates were changed daily for ten days to count the number of eggs and record any mortalities. Daily mean fecundity per female was calculated across 10 replicate bottles per population, taking into account any mortality that occurred. Therefore, population measurement of fecundity was assessed using mixed model ANOVAs, with the fixed effects of season and year (seasonal ROP) and genotype (cpo ROP).
Within-population relationship between learning and reproduction was assessed in the spring reconstructed seasonal populations from 2013 and 2014. Virgins were collected, aged in vials of standard food for 3 days, and subject to the aversive olfactory conditioning and test described above. The flies were thus divided in “learners” (those that chose CS-) and “non-learners” (those that chose CS+). From each replicate test, five “learner” and five “non-learner” females were placed onto food with 5 males from a tester stock; all available flies were used in the few replicates with fewer than five flies. The flies were housed on food supplemented with topical yeast to promote egg production before the assay and on grape-agar substrate during the egg collection. Daily fecundity was counted for 3 days post learning assessment and the mean fecundity per day per female was calculated based on the number of females in the vial. It was then compared between “learners” and “non-learners” from the same replicate test with a paired T-test.
3. Results
(a) Rapid evolution of learning and fecundity in a natural population
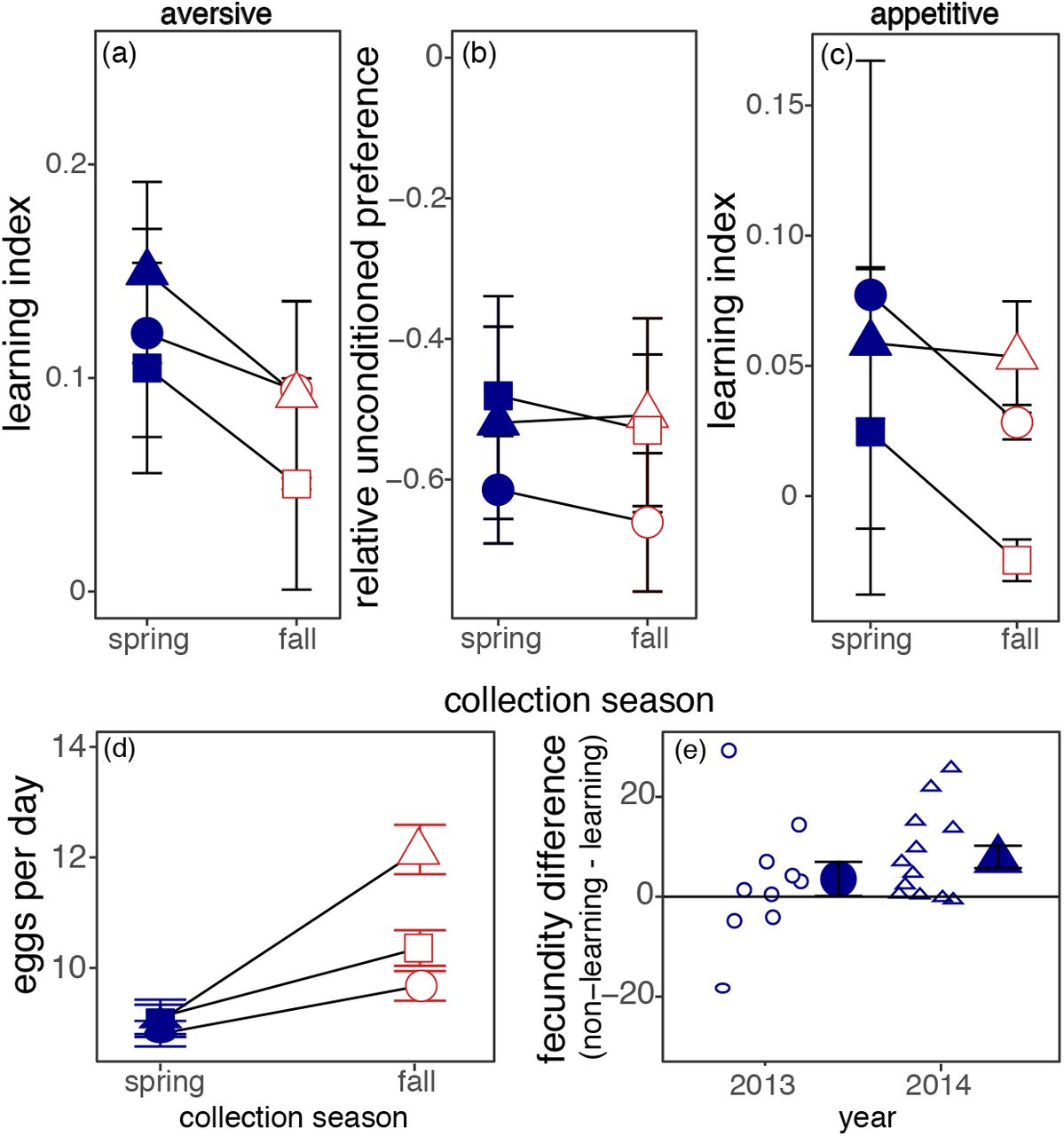
Learning ability in the wild D. melanogaster population evolved rapidly and repeatedly across seasonal time. The spring collections learned better than the fall from the same year when assessed using laboratory aversive conditioning ( , p = 0.03; Figure 1a, Table 1). These behavioural differences were not due to differences in ability to perceive the odorants, as there was no effect of season in absolute preference between odorant and solvent in unconditioned flies (
, p = 0.03; Figure 1a, Table 1). These behavioural differences were not due to differences in ability to perceive the odorants, as there was no effect of season in absolute preference between odorant and solvent in unconditioned flies ( , p = 0.85 Figure 1b).
, p = 0.85 Figure 1b).

Rapid evolution of learning and reproduction in natural populations of Drosophila melanogaster. (a) Higher learning (mean +/− S.E) in the spring populations compared to the fall replicated across three years: 2012 (square), 2013 (circle), 2014 (triangle). (b) No difference in absolute preference in either population when unconditioned flies are given the choice between one odour and solvent. (c) Spring populations return to positive conditioning food at a higher rate than fall populations. (d) Fewer eggs laid per day in the spring populations compared to the fall averaged over ten days. (e) Individuals that do not learn in the aversive conditioning assay have higher daily reproductive output over three days than those that learn. Difference between individuals is shown in small outlined shapes and the mean difference +/− SE in the large, filled shapes.
Analysis of Deviance (Likelihood ratio test)
The spring collections also were better at learning an association between food and flavour tested in outdoor mesocosms. The spring flies were more likely than the fall flies to be trapped in traps flavoured with the same flavour as the food they previously encountered ( , p = 0.017, Figure 1c).
, p = 0.017, Figure 1c).
The seasonal pattern of fecundity was opposite to that for learning ability. At the population level, the fall populations laid on average 30% more eggs per female per day than the spring populations (Figure 1d; F1,57 = 44.2, p < 0.0001). A phenotypic trade-off between learning and reproduction was also apparent at the individual level within the spring populations. Females within each population that chose the “wrong” odour (i.e., the odour associated with shock) in the aversive learning assay subsequently laid an average of six more eggs (25% more) per day compared to the females that made the “correct” choice, i.e., avoided the shock-associated odor (t14= −2.84, p = 0.013, Figure 1e).
(b) Natural variation in cpo sequence affects learning and fecundity
Recombinant outbred populations (ROPs) homozygous for the spring cpoTTA genotype had higher aversive learning than the fall ROP; this pattern persisted across all retention intervals between conditioning and testing ( , p = 1.91×10-7; Figure 2a, Table 1). In the absence of conditioning, the spring ROPs showed a weaker avoidance of the odorants compared to the fall with 1-hour between shock exposure and the test (
, p = 1.91×10-7; Figure 2a, Table 1). In the absence of conditioning, the spring ROPs showed a weaker avoidance of the odorants compared to the fall with 1-hour between shock exposure and the test ( , p=6.52 × 10-3, Figure 2b), but not at 5-minutes or 24-hour retention intervals. The cpo ROPs did not differ in the relative preference of naïve flies choosing between the odorants (
, p=6.52 × 10-3, Figure 2b), but not at 5-minutes or 24-hour retention intervals. The cpo ROPs did not differ in the relative preference of naïve flies choosing between the odorants ( , p = 0.29, Figure 2c), although there was an interaction between genotype and odorant (
, p = 0.29, Figure 2c), although there was an interaction between genotype and odorant ( , p = 0.012) with the spring ROP having a slight preference for MCH and the fall for OCT. However, replicate flies were conditioned in both directions to avoid either odour so this slight preference should not affect the quantification of learning ability. Thus, the difference in learning performance is not explainable by differences in odour perception. The spring ROPs also had a higher appetitive associative learning when assessed in the natural mesocosms (
, p = 0.012) with the spring ROP having a slight preference for MCH and the fall for OCT. However, replicate flies were conditioned in both directions to avoid either odour so this slight preference should not affect the quantification of learning ability. Thus, the difference in learning performance is not explainable by differences in odour perception. The spring ROPs also had a higher appetitive associative learning when assessed in the natural mesocosms ( , p=0.039, Figure 2a). The spring cpoTTA ROP was characterized by lower fecundity, with females laying 36% fewer eggs than females from the fall cpoCGT ROP (Figure 2d, F2=8.23, p=0.01).
, p=0.039, Figure 2a). The spring cpoTTA ROP was characterized by lower fecundity, with females laying 36% fewer eggs than females from the fall cpoCGT ROP (Figure 2d, F2=8.23, p=0.01).

{kind=link}
{kind=link}
Natural variants in couch potato (cpo) involved in trade-off between learning and reproduction. (a) Flies containing the spring (cpoTTA) genotype show better learning than flies that contain the fall (cpoCGT) genotype, across all retention intervals between conditioning and test in the aversive shock assay, as well as in the appetitive conditioning of the natural mesocosm. (b) Unconditioned flies containing the spring genotype have a slightly higher absolute preference for odour instead of solvent compared to the flies containing the fall genotype. (c) However, there is no difference in relative preference for either of the experimental odours in unconditioned flies. (d) Flies containing the fall cpo genotype had higher daily reproductive output than flies containing the spring cpo genotype. In all panels the symbols are mean +/- SE.
4. Discussion
(a) Rapid evolution in natural populations
Our data show that learning and fecundity both evolve rapidly in a natural population of D. melanogaster over the scale of approximately 10-15 generations from spring to fall and approximately 1-2 generations between fall and spring. The differences can be attributed to annual cyclical genetic changes because environmental effects are removed by rearing and testing these populations in common laboratory conditions. The repeatability across years indicates that this is not a result of genetic drift but instead of deterministic evolutionary processes. Genomic data strongly suggest that gene flow through migration is not generating the observed seasonal cycles in allele frequency [45,46]. Therefore, the rapid and repeatable seasonal changes in learning and reproduction are more likely to reflect seasonally fluctuating selection. Learning is thus not only a mechanism of plasticity that allows organisms to respond rapidly to environmental change; we demonstrate that learning ability itself can evolve rapidly in nature.
The learning differences between spring and fall populations were consistently observed in a laboratory aversive conditioning assay and in an appetitive conditioning assay performed in outdoor mesocosms. In both assays, flies originating from the spring collections performed better than those from the fall. Learning associations between odours and both aversive and appetitive stimuli appear to be mediated by the same molecular mechanisms of synaptic plasticity in the mushroom body of the brain [47]. The memory of appetitive and aversive associations is formed at least partly in different subsets of mushroom body neurons, and acting on the appetitive and aversive memories involves different neurotransmitters [47]. Here, the similar pattern in learning performance in both assays implies that the evolutionary change affected a shared component of the learning process. This is consistent with an experimental evolution study in which Drosophila populations selected for appetitive learning also performed better in the aversive shock-odour assay [48].
The higher learning in the spring population is counter to the prediction that learning should improve during the summer when flies are active and perform many behaviours known to depend on learning. However, cognitive abilities may evolve to match the demands posed by an organism’s biological and physical environment [49,50]. Energetically challenging environments (e.g., cold winters or severe droughts) are hypothesized to favour cognitive performance at the cost of other physiological systems receiving less resources [51]. For example, cache seed recovery success in several bird species suggests that learning may be important for overwintering survival in harsh environments [52–54]. Although flies do not cache food, it is possible that learning is also important for other aspects of D. melanogaster overwintering survival in hash climates, such as the ability to find a suitable overwintering site or temperature dependent patterns of foraging [55]. However, it is also possible that seasonal evolution of learning may be driven by selection on genetically correlated traits, such as fecundity.
(b) Links between resilience, fecundity and learning
We found that flies from fall collections show higher fecundity than those from the spring. The opposite trend has previously been reported in wild-caught females: higher fecundity during the first 24-hours after capture in flies collected in the spring compared to those collected in the fall [18]. However, many factors that change with season (e.g., age and nutrition) also affect fecundity; thus, the previously identified differences among wild-caught flies could have reflected these environmental effects rather than genetic differences. In the present study these environmental effects were excluded by rearing flies in a common garden, exposing for the first time a genetic component of seasonal changes in reproductive output. Fecundity is likely to be under strong selection during the summer months of rapid population expansion. Furthermore, previous studies have demonstrated that fly lines collected in the spring are more resilient than those from the fall, with higher propensity for reproductive dormancy [17], greater resistance to thermal stress [18], starvation [18] and bacterial pathogens [56]. The winter conditions are expected to favour such resilience traits, and at least some of these traits are known to trade-off with fecundity [57,58]. Altogether, these results support the hypothesis of alternating selection for resilience required for survival in the harsh winter conditions and for reproduction during the summer expansion [59,60].
The seasonal changes learning performance could in principle have evolved for reasons unrelated to those driving the seasonal fluctuations in resilience and fecundity. However, in addition to the inverse relationship between learning performance and fecundity in spring versus fall populations, we found a negative phenotypic correlation between these traits at the individual level within populations. Females that made the “correct” odour choice in the aversive assay had lower fecundity that females from the same populations that made the “incorrect” choice (i.e., chose the odour previously associated with shock). The inverse relationship between learning and fecundity is consistent with previously reported trade-offs between these traits in a range of taxa [28,29,61]. The trade-off supports the hypothesis that the decline in learning performance from spring to fall in the natural Drosophila population is connected with selection for fecundity during the summer months. If so, the concerted annual evolutionary changes in learning performance, fecundity and resilience traits would be mediated by seasonal fluctuations in allele frequencies at loci with pleiotropic effects on all those traits.
(c) Candidate polymorphism at cpo
Our results suggest cpo as one such pleiotropic candidate gene that mediates the annual seasonal evolutionary cycle. Variants of cpo show latitudinal clines and seasonal fluctuations in frequency [33,34,45,46,62,63] and affect many traits including activity, dormancy, fecundity and lifespan [33,35,37]. We show that genetic variants of cpo also affect learning performance: the ROP homozygous for the cpo genotype that is more frequent in the spring than in the fall and increases in frequency with latitude [35] performed better in the aversive learning assay than the ROP homozygous for the alternative fall genotype. Furthermore the spring ROP had a lower fecundity than the fall ROP [35]. The cpo ROPs that represent annual frequency oscillations and latitudinal patterns of three loci in cpo are able to reconstruct the seasonal antagonistic effects on fecundity and learning performance that we observed in the wild-derived collections. This makes cpo a strong candidate to contribute to the cyclic annual evolution of these traits in the natural population of D. melanogaster.
Learning is a new phenotype associated with cpo even though the RNA-binding protein is known to be expressed in the mushroom bodies and cpo mutants have behaviour phenotypes [36,64]. cpo knockdown mutants have reduced odour avoidance [65], which we saw in flies carrying the spring cpo genotype that had weaker avoidance of odours when paired with air during the control. This leaves the possibility that the spring cpo genotype impairs perception of odours; however, this should have led to impaired rather than improved learning performance. Thus, the difference in learning between the genotypes is unlikely to be due to differences in olfaction, also confirmed by the odour response controls we report above. It is more plausible that the natural cpo genotypes we studied, or genetic variants linked to them, affect the neural processes of memory formation, storage and/or recall, although the mechanism by this RNA-binding protein acts on the function of neurons remain known.
(d) Pleiotropy as a driving force in evolution of learning
The discussion of learning evolution often assumes that inter- and intra-specific differences in learning performance are mainly driven by ecological factors that affect the benefits of learning, such as environmental predictability, reliability of cues, opportunities for learning and costs of making mistakes [1,50,66,67]. However, our study points out that natural genetic variants that affect learning genes are likely to have broad pleiotropic effects. Similarly, a natural polymorphism in D. melanogaster foraging (for) gene is also pleiotropic: in addition to learning [68,69] it affects many traits, including foraging behaviour [70], aggregation [71] and competitive ability [69]. Highly pleiotropic effects of genes involved in learning also emerges from the study of mutant phenotypes [72,73]. The overall force of selection acting on such pleiotropic polymorphisms in natural populations reflects their aggregate impact on survival and reproduction mediated by the diverse ecologically relevant traits they influence. This makes it difficult to disentangle which traits are directly under selection for their fitness benefits and which mainly evolve as a by-product of natural selection on correlated traits. It is unclear in our results if changes in learning performance are driven by ecological factors that affect the usefulness of learning or if they evolve as a pleiotropic effect of selection acting on the focal polymorphisms for reasons unrelated to learning performance. Such pleiotropy may reflect alternative coadapted strategies or it may be a non-adaptive mechanistic consequence of gene action. Nonetheless, even if evolving mainly as correlated response to selection on other traits, the learning differences that we identify between our spring and fall populations and between the cpo genotypes may have ecologically relevant consequences for behaviour.
Author contributions
ELB, TJK and PS designed the experiment. ELB performed the experiment and did the analyses. ELB, TJK and PS wrote the manuscript.
Acknowledgements
This work was supported by NSF GRF DGE-0822 (ELB), the Rosemary Grant Award from the Society for the Study of Evolution (ELB), the Teece Dissertation Research Fellowship from the University of Pennsylvania (ELB), the Peachey Environmental Fund from the Department of Biology, University of Pennsylvania (ELB), and NIH R01GM100366 (PS), as well as research funding from the University of Lausanne to TJK.
References




