

Abstract
Originality-significance statement Increased incidence of food-borne disease outbreaks caused by fresh produce contaminated with Escherichia coli O157:H7 and Salmonella spp. has prompted researchers to examine the interaction between these bacteria and various plant species under different environmental conditions. Although studies show that human enteropathogenic bacteria survive both on the surface of and inside plants, little is known about the molecular mechanism underlying plant invasion and colonization. Here, we examined the interaction between the human pathogenic bacterium Shigella and the model plant Arabidopsis. We found that four Shigella spp. strains proliferated successfully in Arabidopsis, causing symptom-like lesions in tissues. Using mutants lacking T3S effectors (i.e., noninvasive human strains), we demonstrated that effectors regulating pathogenesis of shigellosis in humans also play a central role in bacteria-plant interactions. To the best of our knowledge, this is the first study to examine Shigella-mediated virulence and host immune suppression in a plant host at a molecular level.
Summary Although there is debate about whether human intestinal pathogenic bacteria are also plant pathogens, it is clear that these bacteria use plants as an alternative host. Shigella, which infects primates, is reportedly transmitted by fresh vegetables; however, its molecular interactions with plants have not been extensively studied. Here, we show that four Shigella strains, S. boydii (S. b), S. sonnei (S. s), S. flexneri 2a (S. f 2a), and S. flexneri 5a (S. f 5a), proliferated at different levels in Arabidopsis thaliana. Microscopic studies revealed that these bacteria were present inside leaves and damaged plant cells. GFP-labeled S. b, S. f 2a, and S. f 5a entered plants via guard cells, and S. f 2a infiltrated root tissues and colonized roots. Using mutants lacking type III secretion systems (T3SS), we found that T3SS of Shigella that regulate the pathogenesis of shigellosis in humans also play a central role in attachment and multiplication in Arabidopsis. Furthermore, the immunosuppressive activity of two T3S effectors, OspF and OspG, were needed for the proliferation of Shigella in Arabidopsis. These findings demonstrate that Shigella-mediated virulence determinants are expressed, and pathogenic symptoms are observed, in model plants.
Introduction
Shigella is a Gram-negative bacterium from the Enterobacteriaceae family. It is a non-motile, rod-shaped, facultative intracellular and invasive pathogen, very closely related to Escherichia coli. Based on the carbohydrate composition of the O-antigen, i.e., the polysaccharide component of the lipopolysaccharide molecule that is the major bacterial surface antigen, Shigella is classified into four serogroups. These have been given species designations, namely, S. dysenteriae 1 (serogroup A), S. flexneri (serogroup B), S. boydii (serogroup C), and S. sonnei (serogroup D) (Lindberg et al., 1991; Schroeder and Hilbi, 2008). Shigella spp. are important epidemic pathogens and a serious public health concern in developed and developing countries. Although about 164,300 deaths in all age groups, and 54,900 deaths in children younger than 5 years, were reported globally in 2015, the actual number of infections might be higher because mild symptoms are not reported (Mortality and Causes of Death, 2016). This means that the microorganism may employ a variety of survival strategies not only for infection but also for survival in a restricted host. In recent years, the morbidity or mortality of the infection caused by the emergence of Shigella that produces β-lactamase, an enzyme that underpins the resistance to the third-generation antibiotics cephalosporins, has been increasing (Zhang et al., 2011).
Shigella is a human-adapted pathogen that infects the host via multiple transmission routes. Human hosts are primarily infected by the ingestion of water or food contaminated with bacteria from feces (the fecal-oral route), including ingestion of undercooked contaminated meats and improperly washed contaminated vegetables (Mead et al., 1999; Weir, 2002; Gupta et al., 2004). Fresh fruits and vegetables, such as lettuce, tomatoes and green peppers, are responsible for the widespread transmission of food-borne infections by Salmonella or Shigella (Guchi and Ashenafi, 2010; Semenov et al., 2010; Gu et al., 2013). These observations suggest that human pathogenic bacteria use plants as alternative hosts as a stable environmental niche. Several human pathogenic bacteria, including Salmonella enterica serovar Typhimurium, Escherichia coli O157:H7, and Pseudomonas aeruginosa, are known to use plants as alternative hosts (Plotnikova et al., 2000; Semenov et al., 2010). These bacteria can attach to the plant surface, and actively invade and proliferate in plant tissues (Cevallos-Cevallos et al., 2012; Martínez-Vaz et al., 2014). In particular, several enteropathogenic bacteria, including E. coli O157:H7 and Salmonella, spread within plants through vascular tissues after infection via contaminated water (Solomon et al., 2002). Although Shigella contamination has also been reported in plants, it is not yet known whether the bacterium actively invades and/or proliferates inside the plant.
Unlike animals, plants have no adaptive immune system; instead, each cell possesses an innate immune system. The innate immune system in plants and animals recognizes and suppresses pathogens and has common features that are preserved throughout evolution. Plant pattern recognition receptors recognize conserved microbial or pathogen-associated molecular patterns; the pattern-triggered immunity (PTI) is activated via the mitogen-activated protein kinase (MAPK) cascades (Jones and Dangl, 2006). To suppress PTI, bacteria inject effector proteins into plant cells using type III secretion systems (T3SS). To counteract this PTI evasion response, the plant nucleotide binding-leucine rich repeat proteins recognize the pathogen effectors; effector-triggered immunity is then activated to accompany the hypersensitive response (Jones and Dangl, 2006). For human or animal intestinal bacteria to infect plants, the PTI must first be disabled. S. enterica serovar Typhimurium, similar to its activity in the mammalian host, uses T3SS to suppress plant immune responses (Schikora et al., 2011; Schikora et al., 2012). In particular, SpvC, one of the T3S effector proteins of S. enterica, targets the MAPK signaling system in both plant and animal hosts to suppress the host PTI (Neumann et al., 2014).
Here, we examined the ability of four Shigella strains (S. s (Holt et al., 2012), S. b and S. f 2a (Wei et al., 2003), and S. f 5a (Onodera et al., 2012)) to proliferate in Arabidopsis plants. We found that the four strains invaded and proliferated differently in plant tissues. Proliferation of mutants lacking the T3S effectors, i.e., noninvasive human strains, was reduced in planta. Reverse genetics and molecular biology experiments demonstrated that the immunosuppressive function of Shigella T3S effectors OspF and OspG was important for Shigella proliferation in plants. These observations indicate that Arabidopsis may be useful as a model host for studying the pathogenesis of Shigella.
Results
Four Shigella spp. strains interact differently with Arabidopsis
To observe the behavior of the human pathogen Shigella in plants, we investigated the interaction of four Shigella spp. strains representing three serogroups (S. b, S. s, S. f 2a, and S. f 5a) with the model plant A. thaliana. Shigella is a water-borne pathogen, with infection spreading via contaminated water (Pandey et al., 2014). Hence, we chose to use a flood-inoculation approach (Ishiga et al., 2011), which is thought to mimic natural inoculation closely. Similarly to the phytopathogen Pseudomonas syringae pv. tomato DC3000 (Pst), when 2-week-old Arabidopsis seedlings were inoculated with S. s or S. f 2a, clear symptoms were observed, such as yellowing and necrosis of the leaves (Fig. 1A). On the other hand, no obvious symptoms were apparent after inoculation with S. b and S. f 5a (Fig. 1A) (Liu et al., 2015). Pst ΔhrcC, a mutant lacking the T3SS of Pst, and the non-pathogenic bacterium E. coli DH5a were used as negative controls for infection. In addition to observing the symptoms, the bacterial growth in planta was also evaluated, to detect initial plant adherence and proliferation. Early attachment of all Shigella strains and DH5a was more than 10 times lower than that of Pst and Pst ΔhrcC (Fig. 1B). However, at 3 days post-inoculation (dpi), the extent of cell amplification differed depending on the Shigella strain; nevertheless, the cell number of all strains increased more than 1 000 times. Notably, S. s cell numbers increased over 105 times. This level of proliferation was comparable to that of plant pathogens. By contrast, the interaction between S. f 5a and Arabidopsis (in terms of symptoms and/or proliferation) was similar to that between the plant and non-pathogenic DH5α.
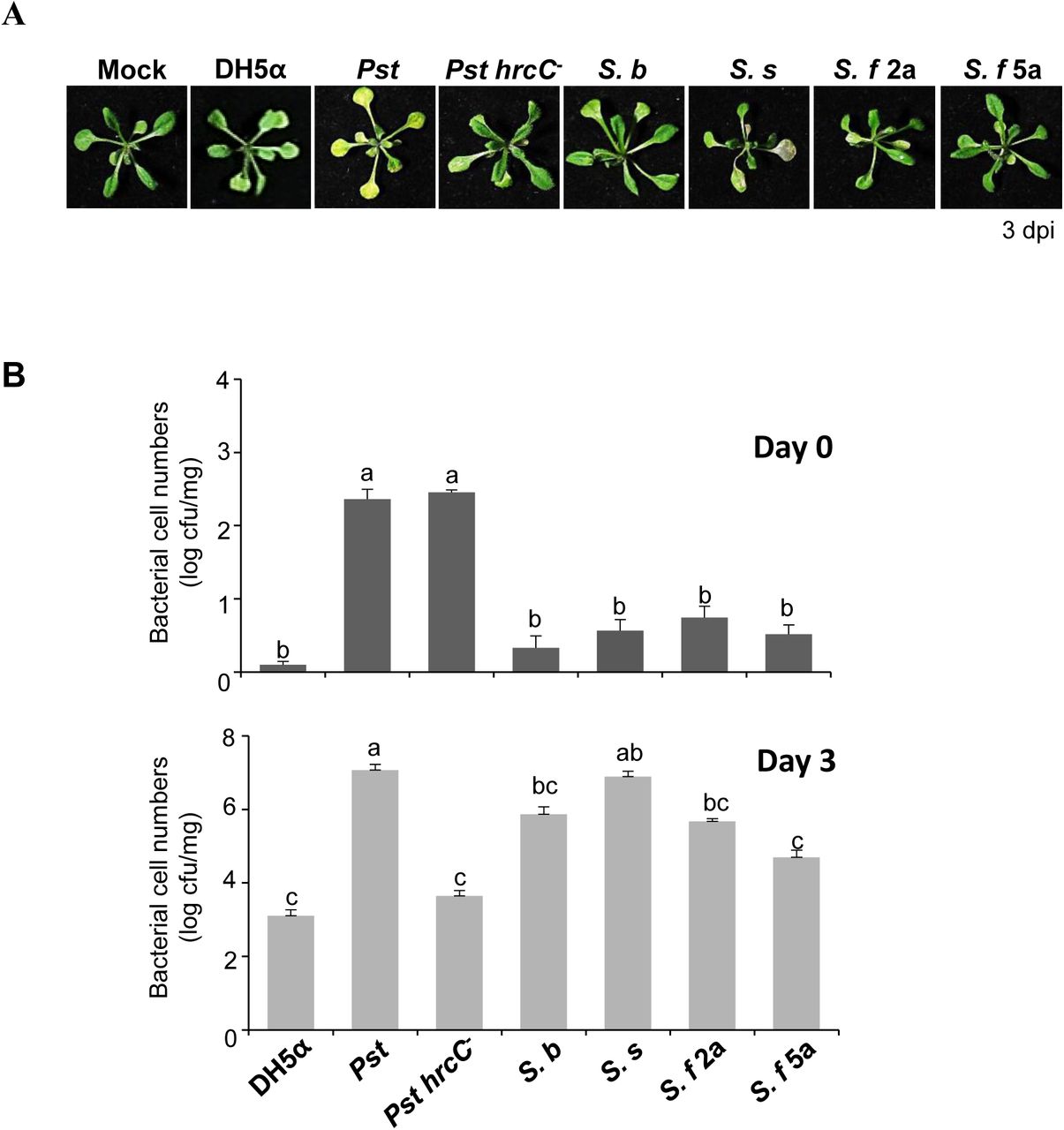
Shigella proliferates and induces disease symptoms in Arabidopsis plants. (A, B) Arabidopsis seedlings in 1/2 MS medium were flood-inoculated with sterile water containing 0.025% Silwet L-77 (Mock) or bacterial suspensions (5 × 105 cfu/ml). (A) The symptoms of Arabidopsis inoculated with Shigella spp. by flood-inoculation. Photographs of representative symptoms were taken at 3 dpi. (B) Bacterial cell numbers were evaluated on Days 0 and 3 after flood-inoculation. The bars represent the mean ± SD of three replicates and the different letters indicate significant differences between samples (P < 0.05, one-way ANOVA). All experiments were repeated three times independently and representative results are shown.
Apparently different symptoms and bacterial proliferation rates were observed for different Shigella strains. This suggested that a variability of the plant interaction mechanisms among the strains, e.g., the adherence and multiplication, might contribute to the differences. Differences in the nutritional requirements of bacterial strains may constitute another reason for the differences in the growth rates within the Arabidopsis host.
Penetration of the plant leaf surface and subsequent internalization of Shigella spp
Since we observed that Shigella proliferate and induce disease-like symptoms in Arabidopsis, we used scanning electron microscopy (SEM) and transmission electron microscopy (TEM) to examine whether the bacterium multiplies on the leaf surface or in the intercellular space (apoplast). The cuticle of a plant leaf is the first physical barrier encountered by the pathogen; known plant pathogens enter the host plant cell through a wound, an open stomata, or a vector (Yeats and Rose, 2013). As anticipated, Pst, which infects plants via open stomata (Panchal et al., 2016), colonized guard cells at 24 h post-infection (Fig. 2). All tested Shigella strains clustered around guard cells and the surface of epidermal cells (Fig. S2). S. s and S. b formed relatively wide clusters in the surrounding areas, including guard cells (Fig. 2 and S2). In particular, S. b and S. f 2a intensely colonized guard cells (Fig. 2), leading us to speculate that they enter plants via open stomata, similar to Pst.

Shigella adheres to the Arabidopsis epidermis and clusters around guard cells. Arabidopsis seedlings were flood-inoculated with bacterial suspensions (5 × 106 cfu/ml). After 24 h, the leaves were fixed and analyzed under a SEM. The Pst cells were observed over the stomata. Shigella strains were associated with the stomata. Bar, 10 μ,m. Representative image showing bacteria around guard cells (indicated by white arrows). The results are representative of two independent experiments.
Most plant pathogenic bacteria in planta infect and colonize the apoplast (Abramovitch et al., 2006; Gao et al., 2016). We used TEM to investigate whether Shigella is capable of intercellular colonization and causing damage to plant cells by analyzing Arabidopsis leaves flood-inoculated with Shigella (Fig. 3). Indeed, the TEM images revealed Shigella colonization of the intercellular spaces and bacterial attachment to the host cell walls. The presence of the microbes in the intercellular space resulted in the alteration of the host organelle structure, such as the separation of the plasma membrane from the cell wall, the liberation of cell organelles, and disruption of chloroplasts (Fig. 3). This effect was most pronounced in plant cells inoculated with S. b and S. s; further, S. s and S. b were more commonly found in the intercellular spaces than S. f 2a and S. f 5a.

Shigella localizes in the apoplast and affects plant cell structures. Arabidopsis seedlings were flood-inoculated with bacterial suspensions (5 × 106 cfu/ml). After 24 h, the leaves were evaluated under a TEM. The Shigella strains colonized the intercellular spaces. Bar, 5 μ,m (left panels). Bar, 1 μ,m (right panels). Each column of micrographs represents a different magnification. TEM images revealed the Shigella (white arrows) attached to the cell wall in the intercellular spaces and altered mesophyll cells. Black arrows indicate separation of the plasma membrane from the cell wall, the abnormal organelles and disruption of chloroplasts. The results are representative of two independent experiments.
Taken together, these results indicated that, although the proliferative capacity of the Shigella strains differs, the cells can invade and colonize the apoplast of plants, thereby causing structural damage to the host.
Penetration of Arabidopsis root tissue by Shigella strains
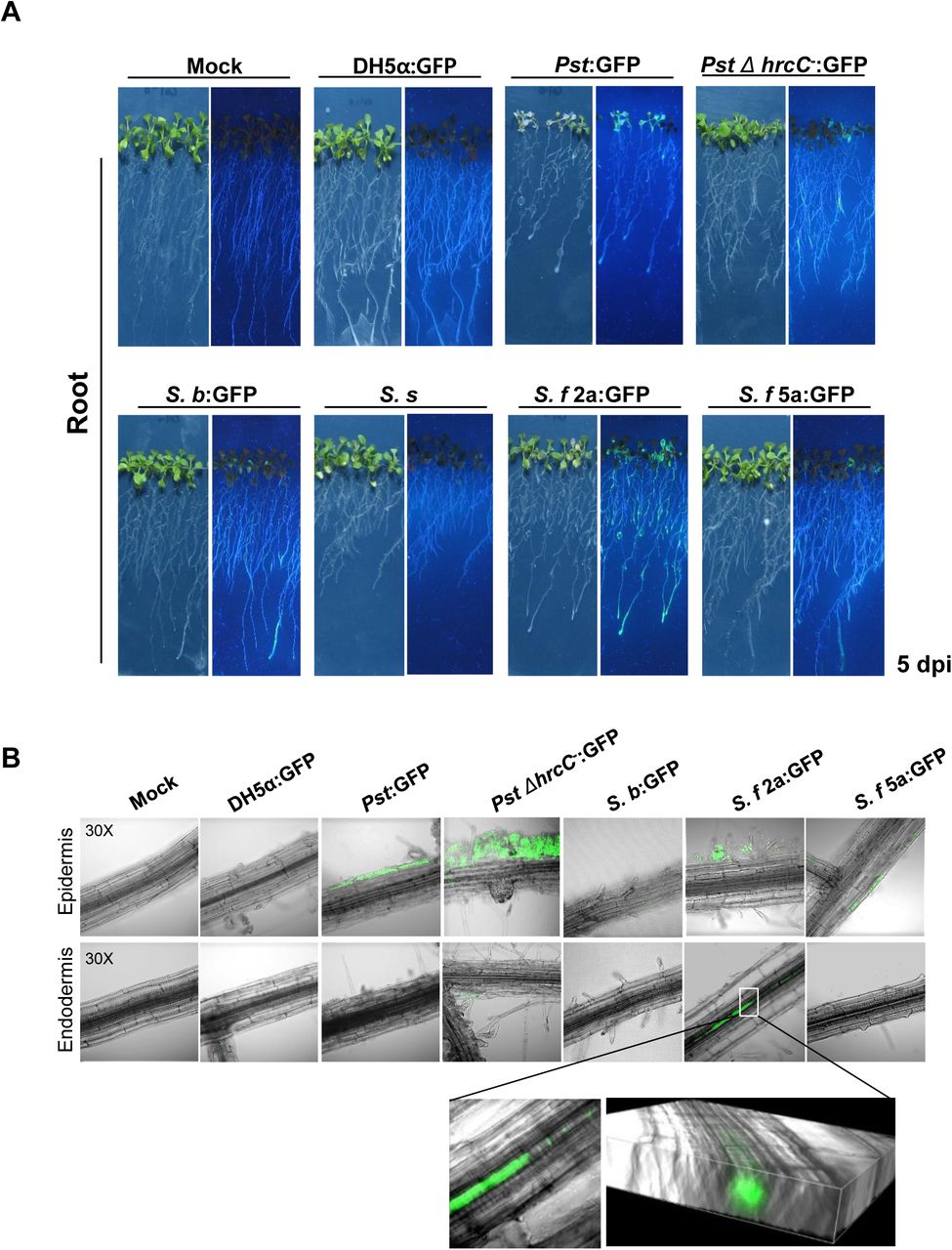
Previously published studies demonstrate that several human pathogenic bacteria, including E. coli O157:H7 and Enterococcus faecalis, invade the leaves and roots of A. thaliana (Jha et al., 2005; Deering et al., 2012). To determine whether Shigella strains invade through A. thaliana roots, we attempted to label the four Shigella strains and DH5a with green fluorescent protein (GFP) and then observe them in live plant tissues. The S. b, S. f 2a, and S. f 5a strains were labeled successfully with GFP (the S. s strain was not), confirming that GFP labeling did not affect their growth in plants (Fig. S1).
Next, bacterial suspensions were dropped onto the root tips of Arabidopsis plants that were grown vertically. Five days later, whole plants and root tissues were photographed under ultraviolet (UV) light to observe the distribution of fluorescently labeled bacteria (Fig. 4). In accordance with the disease phenotypes observed, Pst (which is a foliar pathogen) exhibited strong fluorescence throughout the plant, despite the fact that it was applied to root tips (Fig. 4A). To observe Pst in root tissues, inoculated roots were washed with sterile water and observed under a fluorescence microscope. GFP fluorescence was observed only in epidermal cells after application of Pst to root tips (Fig. 4B). This finding indicates that Pst spreads throughout the entire plant along its surface, not by invading root tissues or colonizing the roots. Fluorescence was also observed throughout plants when GFP-labeled S. b, S. f 2a, and S. f 5a were applied to root tips, although the fluorescence intensity differed (Fig. 4A). In root tissues, GFP fluorescence was observed in root endodermal cells only in S. f 2a-treated plants (Fig. 4B). Taken together, these results indicate that Shigella invades plant leaves and roots in a strain-dependent manner, and then moves along the surface and through the internal vascular tissues of the plant.

Shigella S. f 2a invades and colonizes plant roots. (A, B) Root tips of Arabidopsis seedlings were drop-inoculated with GFP-labeled bacterial suspensions (5 × 107 cfu/ml). (A) Bacterial labeled Shigella are localized in the epidermal or endodermal cells of Arabidopsis roots. GFP images were taken using a confocal microscope. Higher magnification micrographs and 3D Raman confocal volume images show internalization of S. f 2a in Arabidopsis roots. All experiments were repeated at least three times, and representative results are shown.
In addition, we dropped bacterial solutions onto root tips and observed symptoms in Arabidopsis plants for 14 d. Severe inhibition of root growth and leaf yellowing were observed in plants inoculated by the S. b, S. s, and S. f 2a strains (Figure S3). Inoculation of S. f 5a caused slight inhibition of root growth, but caused much less damage to the plant than other strains.
Shigella T3S effectors are necessary for attachment and multiplication in Arabidopsis
T3SS is the key determinant of the virulence of many Gram-negative bacteria, including animal and plant pathogens. Thus, we investigated if pathogenic proteins, which are involved in animal host infections, are required for invasion and multiplication of Shigella in plants. To study the biological role of T3SS in the interaction between Shigella and plants, we used noninvasive variants of S. f 2a and S. f 5a (strains Δvp and BS176, respectively) (Sansonetti et al., 1982; Wenneras et al., 2000; Shim et al., 2007). To facilitate observation of bacterial invasion in living plants, strains S. f 2a, S. f 5a, and their variants were labeled with GFP, and bacterial growth and expression of effector proteins were verified (Fig. S4). Bacterial proliferation in plants after inoculation with Δvp or BS176 strains was 10 times lower than that after inoculation by parental Shigella strains; similar results were observed for GFP-labeled Shigella strains and mutants (Fig. 5A).

The virulence plasmid-deficient mutant of Shigella is impaired in the Arabidopsis multiplication. (A, B) Arabidopsis seedlings in 1/2 MS medium were flood-inoculated with DH5a, S. f 2a, Δvp, S. f5a, or BS176, and their GFP-labeled variants (5 × 105 cfu/ml). (A) The bacterial cell numbers in planta were determined at 0 and 3 dpi. Bars represent the mean ± SD of three replicates and the different letters indicate significant differences between samples (P < 0.05, one-way ANOVA). (B) Images of GFP-labeled wild-type Shigella in leaf epidermal cells of Arabidopsis acquired at 24 hpi under a fluorescence confocal microscope. Bar, 10 μ,m. The blue represents auto-fluorescence of chlorophyll. All experiments were repeated at least three times, and representative results are shown.
To examine involvement of T3SS in bacterial invasion of the plant surface, leaves were flood-inoculated with GFP-labeled bacteria and leaf surfaces were observed 24 h later (Fig. 5B). Examination under UV light revealed that the levels of GFP-labeled S. f 2a and S. f 5a on leaf surfaces were higher than those of Δvp and BS176, especially in open stomata (Fig. 5B). We also confirmed that none of the Shigella strains grew in plant culture medium (Fig. S5), and that effector protein expression was maintained in bacteria recovered from plants (Fig. S6).
To determine if other Shigella serotypes interact with plants in a T3SS-dependent manner, we attempted to obtain S. s and S. b mutants that had lost pathogenic plasmids via continuous subculture. A non-pathogenic mutant was successfully obtained for S. s (Fig. S7). When plants were inoculated with this T3SS-deficient S. s strain, disease phenotypes and bacterial growth were greatly reduced (Fig. S7), as observed with the S. f serotype. Altogether, these results suggested that the T3SS of Shigella that operate during an infection in human are also required for interactions with plants.
Expression of Shigella T3S effector genes and modulation of Arabidopsis MAPK signaling after Shigella inoculation
To further investigate involvement of Shigella T3S effectors in plant interactions, we measured expression of key virulence effectors related to human pathogenesis, including ipaB, ipaC, icsA, icsB, virB, and virF, in S. f 2a‐, S. f5a‐, Δvp‐, and BS176-infected Arabidopsis plants (Bando et al., 2010). These effectors play a role in mammalian cell lysis (ipaB, ipaC), intracellular spread (icsA, icsB), and regulation of virulence factor (virF, virB) expression (Ogawa et al., 2008). Total RNAs were isolated from Arabidopsis leaves at 6 and 12 h after Shigella inoculation, and changes in expression of virulence genes were confirmed by quantitative RT-PCR (Fig. 6A). Expression of all Shigella virulence genes examined in WT S. f 2a-or S. f 5a-inoculated Arabidopsis leaves increased.
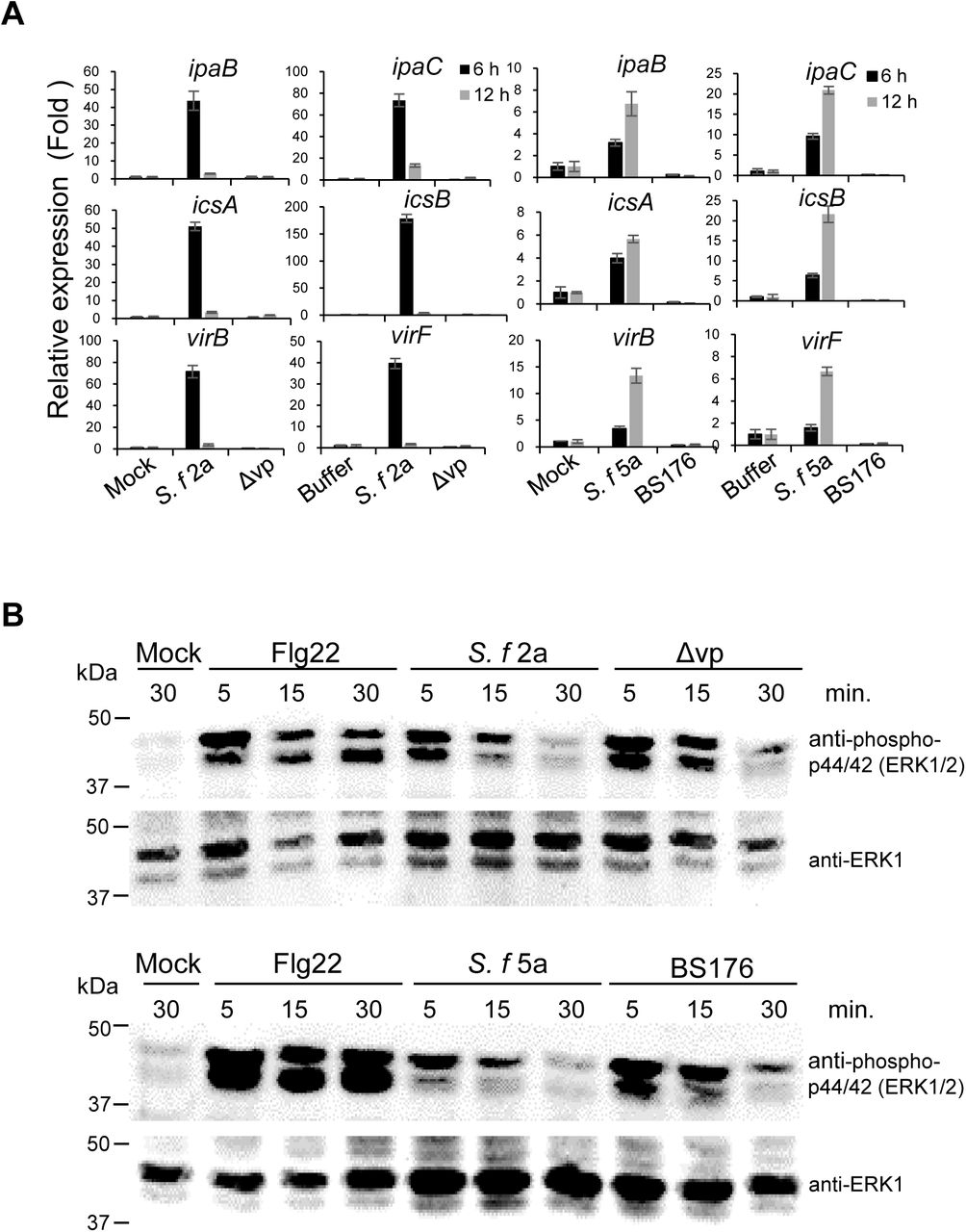
Expression of Shigella effectors and MAPK activation in Arabidopsis plants in response to virulence plasmid-deficient Shigella mutants. (A, B) Arabidopsis leaves were syringe-infiltrated with buffer, 1 μM flg22, or bacterial suspension (5 × 106 cfu/ml), and samples were collected at the indicated times. (A) Transcription of Shigella effectors (ipaB, ipaC, icsA, icsB, virB, and virF) in infected Arabidopsis leaves was analyzed by qRT-PCR. qRT-PCR results were normalized to expression of 16s rRNA. Expression of effectors by plasmid-deficient mutant strains was compared with that in the WT. Data are expressed as the mean ± SD of three replicates. (B) Immunoblotting was performed using either anti-phospho-p44/42 (ERK1/2, upper panels) or anti-ERK1 (lower panels) antibodies. All experiments were repeated at least three times, each with similar results.
In particular, induction was higher and faster in S. f 2a-treated plants than in S. f 5a-treated plants, which is in agreement with the earlier results showing that S. f 2a was more pathogenic to plants than S. f 5a. Expression of virulence genes was not detected in plants inoculated with non-pathogenic mutants Δvp or BS176, similar to buffer-treated control plants. These results indicate that common virulence factors regulate interactions with both plants and human intestinal cells.
To investigate the plant innate immune responses to Shigella inoculation, MAPK phosphorylation was evaluated. The activation of MAPK by phosphorylation is a conserved response of the earliest microbe-triggered immune signaling in both plants and animals (Zipfel, 2009). The flg22 peptide is a representative microbe-associated molecular pattern in plants (Bethke et al., 2009). In plants treated with flg22, pronounced MAPK phosphorylation was apparent within 5 min of treatment and this response lasted up to 30 min (Fig. 6B). On the other hand, MAPK phosphorylation in plants treated with S. f 2a or S. f 5a was reduced; from 15 min on, it was strongly suppressed and almost completely disappearing after ca. 30 min (Fig. 6B). Meanwhile, in plants treated with the virulence plasmid-deficient mutants, Δvp or BS176, MAPK activation was recovered, in contrast to S. f 2a-or S. f 5a-treated plants, although the degree and duration of the phosphorylation were lower than those elicited by the flg22 treatment (Fig. 6B). These observations indicated that Shigella suppresses the innate immunity of Arabidopsis via its T3S virulence effectors.
Suppression of immune signaling in Arabidopsis plants by Shigella T3S effectors OspF and OspG
The most important role of T3S effector proteins injected into host cells is modulation of the host immune response. Therefore, we chose two Shigella effectors, OspF and OspG, to study suppression of plant immune responses by Shigella effector proteins. Shigella OspF was chosen because of its ability to inhibit MAPK signaling, which is conserved in plants and animals (Arbibe et al., 2007; Li et al., 2007). OspG is an important immunosuppressive effector protein secreted at the later stages of infection; this protein interferes with activation of the NF-ĸB pathway (Kim et al., 2005), which is absent from plants.
To find out whether OspF and OspG have virulence activity in plants, we introduced them into a phytopathogen, Pst, and monitored its pathogenicity. First, we used an AvrRpt2-derived T3SS reporter system to check that the introduced OspF or OspG protein was delivered to plant cells via Pst T3SS (Mudgett, 2005). When Arabidopsis leaves were syringe-infiltrated with various Pst-producing AvrRpt101–255 constructs, we found that Pst producing OspF:AvrRpt2101–255 or OspG:AvrRpt2101–255 induced a cell death response at 1 dpi, while control Pst cells containing the empty vector did not. This indicated that the Shigella effectors OspF and OspG were delivered successfully into Arabidopsis cells via Pst T3SS (Fig. S8A, B). Full-length AvrRpt2 was used as a positive control for cell death response. Finally, we examined whether the virulence of Pst was increased by expression of Shigella OspF or OspG. Plants infected with Pst cells producing OspF: HA or OspG: HA showed more severe symptoms than plants infected with the empty vector control (Fig. 7A; Fig. S8C). In addition, the number of bacterial cells was 10 times higher than that in plants infected with the empty vector control (Fig. 7B). Production of OspF:HA and OspG:HA by Pst was confirmed by immunoblotting with an anti-HA antibody (Fig. S8D). Taken together, these observations suggest that the Shigella effectors OspF and OspG function as virulence factors in plant cells.

The virulence activityies of Shigella effectors OspF or OspG. (A, B) Arabidopsis plants were sprayed with Pst cells carrying an empty vector (pME6012), or with cells producing OspF:HA or OspG:HA (5 × 107 cfu/ml). (A) Disease symptoms were monitored for 4 d after spraying. (B) Proliferation of Pst cells producing Shigella effector proteins in Arabidopsis at 0 and 4 d after spray-inoculation. Bars represent the mean ± SD of six replicates, and different letters indicate significant differences between samples (P < 0.05, two-way ANOVA). All experiments were repeated at least three times. (C) Co-expression of OspF:GFP suppresses tobacco MEK2DD-triggered cell death. GFP, OspF:GFP, or OspG:GFP was produced with HA:MEK2DD in N. benthamiana leaves upon infiltration by Agrobacterium carrying the appropriate expression constructs (OD600 0.4). (D) Production of specific proteins in samples from panel A was analyzed by immunoblotting using anti-GFP, anti-HA, anti-phospho-p44/42 (ERK1/2), and anti-ERKl antibodies. Ponceau S was used to stain the RuBisCo protein (loading control). The asterisk indicates the size of the GFP protein not associated with effector proteins.
We attempted to produce the OspF and OspG proteins in plant cells to verify whether their known function in the human host is conserved in the plant cell. To do this, subcellular localization of the two proteins fused with GFP was evaluated in the plant cells. The DNA fragments encoding the OspF:GFP and OspG:GFP fusions were inserted into a binary plant expression vector and transiently expressed in Nicotiana benthamiana leaves using the Agrobacterium system (Fig. S9A, B). The fluorescence signal for both proteins was strong in the plant cell nucleus; OspF:GFP fluorescence was also observed along the cytoplasmic membrane, and punctate OspG:GFP fluorescence was observed in the cytosol (Fig. S9C). In the host human cell, OspF localization is nuclear (Zurawski et al., 2006; Arbibe et al., 2007) and that of OspG is nuclear and cytoplasmic (Kim et al., 2005; Zhou et al., 2013; de Jong and Alto, 2014), i.e., their subcellular localization is similar to that of plant cells.
Previously, in vitro studies demonstrated that OspF exerts a phosphothreonine lyase activity and irreversibly removes phosphate groups from MAPK (Arbibe et al., 2007; Li et al., 2007). We used a phenotypic screening system involving a MEK2 (a tobacco MAP kinase kinase2) mutant, MEK2DD, to investigate whether the virulence associated with Shigella OspF in plants was linked to the same mechanism of action as in animals. MEK2DD, a constitutively active mutant of MEK2, induces cell death when overproduced in plant leaves (Yang et al., 2001; Kim and Zhang, 2004). In this experiment, an HA:MEK2DD clone fused to an HA epitope tag at the N-terminus was used to monitor expression of MEK2DD. As expected, co-production of HA:MEK2DD and GFP (control) resulted in a pronounced cell death (Fig. 7C). Co-production of OspF:GFP, but not OspG:GFP, and MEK2DD completely inhibited the MEK2DD-induced cell death (Fig. 7C). The production of the two effectors fused with GFP was then evaluated in the MEK2DD-producing plant leaves; the production of OspF:GFP was apparent, but that of OspG:GFP was not (Fig. 7D), even though both proteins were stably produced in the absence of MEK2DD (Fig. S9B). The degradation of the OspG protein may have been associated with the activated MAPK. Indeed, as assessed by immunoblotting with specific anti-phosphorylated MAPK antibodies, MAPK phosphorylation was very weak in the OspF:GFP-producing plant samples in comparison with GFP-or OspG:GFP-producing plant samples (Fig. 7D). The production of MEK2DD protein was confirmed in all samples using anti-HA antibodies (Fig. 7D).
These observations strongly suggest that the Shigella effector OspF inhibits plant immune responses by inhibiting activation of MAPK (as in humans), and that OspG induces immunosuppression in plants by targeting distinct MAPK pathways.
OspF or OspG affects Shigella proliferation in plants
To see whether the OspF or OspG proteins play a role in the interaction between Shigella and plants (as in human cells), we inoculated plants with S. f 5a mutants lacking the OspF or OspG proteins and examined their growth. Growth of mutants lacking ospF or ospG was as deficient as that of the virulence plasmid-deficient mutant BS176 (Fig. 8A). Reduced growth of S. f 5a ΔospF or S. f 5a ΔospG mutants was restored completely by complementation of the mutation, indicating that these two effector proteins are indeed important for bacterial growth in plants (Fig. 8A). Next, we monitored activation of MAPKs to determine whether plant immune suppression was affected by deletion of ospF or ospG. As shown in Figure 8B, S. f. 5a ΔospF induced stronger phosphorylation of MAPKs than wild-type S. f 5a; this was offset by complementation with OspF. By contrast, phosphorylation of MAPK in S. f 5a ΔospG-inoculated plants was no different from that in plants inoculated with wild-type S. f 5a, although phosphorylation of MAPKs increased slightly after complementation with OspG (Fig. 8B). Taken together, these results indicate that Shigella effectors OspF or OspG play an important role in increasing bacterial proliferation in both plant and animal hosts.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The Shigella effector proteins OspF and OspG underpin the bacterial virulence in plants. (A) The virulence of the OspF-or OspG-deficient Shigella mutants is impaired in the Arabidopsis model. Arabidopsis seedlings in 1/2 MS medium were flood-inoculated with S. f 5a, ΔospF, ΔospF+pospF, ΔospG, or ΔospG+pospG (5 × 105 cfu/ml). Photographs of representative disease symptoms were taken, and bacterial cell numbers in planta were determined, at 3 dpi. The bars represent the mean ± SD of three replicates. The different letters indicate significant differences between samples (P < 0.05, one-way ANOVA). (B) MAPK activation in Arabidopsis plants in response to infection by ospF or ospG deletion mutants. Arabidopsis leaves were infiltrated with each bacterial strain as described previously and then collected at the indicated times. Anti-phospho-p44/42 (ERK1/2, upper panels) or anti-ERK1 (lower panels) antibodies were used for immunoblotting. All experiments were repeated at least three times, with similar results.
Discussion
In the current study, we investigated the interaction of the human pathogenic bacterium, Shigella, with an alternative host, the Arabidopsis plant. We demonstrated that four Shigella strains, S. b, S. s, S. f 2a, and S. f 5a, invade and colonize Arabidopsis to different extents. It has been clearly demonstrated that, in human, Shigella initially enters the epithelial layer via the M cells through transcytosis, leading to the invasion of the basolateral surfaces of the intestinal epithelial cells. A subsequent gut inflammation leads to the activation of the innate immune response (Phalipon and Sansonetti, 2007). We demonstrated in the current study that, in addition to the human host, Shigella invades Arabidopsis, through vascular tissues and leaf stomata pores (Figs. 2-4). In particular, S. s and S. b formed relatively wide clusters in the surrounding areas, including the guard cells (Fig. 2; Fig. S2). Interestingly, we found that the four studied strains of Shigella associate with the plant cells and induce different plant responses. The bacterial loads of S. s in planta were relatively higher than those of the other strains. By contrast, inoculation of S. f 5a was associated with lower bacterial proliferation and less severe symptoms than observed for other strains (Figs. 1–4; Fig. S2 and Fig. S3). S. f 2a and S. f 5a, which belong to the same serogroup (Lindberg et al., 1991), elicited distinctly different plant responses with respect to disease symptoms. The specific plant responses, which were different for bacterial strains within the same serogroup, suggest that virulent effectors may play a relatively more important role in plant interactions. These observations also indicate that specific plant immune systems may be useful in the search for novel virulence factors expressed by different Shigella strains. Many Gram-negative bacterial pathogens utilize common infection strategies to colonize and invade plant and animal cells, and pathogenicity appears to depend on highly conserved T3SSs, which deliver the effector proteins to host cells (Buttner and Bonas, 2003). By using avirulent mutant strains, we were able to show that effectors that regulate the pathogenesis of shigellosis in humans also play a central role in regulating interactions with Arabidopsis. We showed that secretion of T3S effectors is required to trigger both symptoms and expression of the invasive phenotypes in plants (Fig. 5; Fig. S7). Furthermore, the effector proteins impacted MAPK-dependent innate immune responses in Arabidopsis (Fig. 6–8). Although reduced colonization of T3SS-deficient pathogenic E. coli in plants was previously reported, in contrast with the present study, they suggested that E. coli uses the T3SS apparatus for attachment to leaves, rather than for bacterial growth inside plants (Shaw et al., 2008). The relevance of T3SS for multiplication of Salmonella in plants remains unclear due to the different effects of T3SS function on Salmonella-plant interactions (Iniguez et al., 2005; Schikora et al., 2011; Shirron and Yaron, 2011). By contrast, the T3SS and specific effectors play an important role in proliferation and survival of Pseudomonas syringae pv. syringae B728a on the plant leaf surface (Lee et al., 2012). Until now, the transfer of effector proteins into plant cells by T3SS of animal bacterial pathogens has not been reported. Our results indirectly suggest that Shigella effectors can be transferred into plant cells; however, direct evidence, such as visualization of the delivery of effectors using split fluorescent proteins (Park et al., 2017), is required.
Expression of the T3SS of Shigella is regulated at the transcriptional level and is activated at a permissive temperature (≥ 32°C) (Tobe et al., 1991; Campbell-Valois and Pontier, 2016). We were able to observe expression of the T3SS genes of Shigella under temperatures at which plants grow (22 ± 3°C) (Fig. 6A). A recent study showed that elevation of the temperature increases T3SS-mediated virulence of the phytopathogen Pst in plants, which is in contrast with the negative effect of high temperature on expression of the T3SS of Pst in vitro (Huot et al., 2017). Regardless of the temperature of host cells, it will be interesting to determine whether Shigella regulates T3SS gene expression in vivo and to identify factors that influence T3SS gene expression other than plant temperature.
Salmonella strains capable of proliferating on plant leaves and actively entering plant tissues, root hairs, or trichomes were recently shown to exhibit virulence in animals (Barak et al., 2011; Golberg et al., 2011; Schikora et al., 2011). We demonstrated that Shigella strains actively colonize the surface of and inside Arabidopsis leaves and root tissues (Figs. 1–4) and that bacteria recovered from plants maintain expression of pathogenic proteins (Fig. S6). Collectively, these findings suggest that, similar to Salmonella, Shigella-inoculated plants are a serious risk to food safety and that contamination of plants is another route underlying infection of Shigella, an important human pathogenic bacterium. In this study, we only observed plants artificially inoculated with Shigella in a laboratory environment. Therefore, to confirm the food safety concern surrounding Shigella-inoculated plants, the ability of Shigella to infect a variety of crops grown in the field should be tested. The pathogenicity of plant-isolated Shigella in animals should also be investigated.
The current study provides new insights into host invasion mechanisms utilized by Shigella to interact with an alternative host, the plant Arabidopsis. Studying trans-kingdom pathogenesis involving human-adapted pathogens, such as Shigella, may uncover novel pathogenic mechanisms uniquely activated in response to specific hosts. When we isolated the two Shigella effectors OspF and OspG, and produced them in plant cells, their localization coincided with that in the animal cells (Fig. S9C), and it was apparent that the production of both proteins increased the virulence of plant pathogens (Fig. 7). In addition, we confirmed that OspF inhibits the innate immune response of plants via the same enzymatic activity as in animals (Fig. 7 and Fig. 8). In animals, OspG inhibits the host immune response by inhibiting the activity of NF-ĸB by blocking degradation of IĸB (Ashida et al., 2015). Plants possess an IĸB-like protein called NIM1 (Ryals et al., 1997); however, no other published studies have investigated whether a NF-ĸB-induced immune response exists in plants. In the current study, we demonstrated the ability of OspG to increase the phytopathogenicity of non-Shigella bacteria, and also observed degradation of OspG:GFP upon constitutive activation of MAPK signaling (Fig. 7). The existence of a plant immune signaling pathway similar to that of animal NF-ĸB, which would also be the target of OspG, may hence be assumed. Characterization of the previously unrecognized stress-activated mediators of the innate immunity in plants upon infection with food-borne pathogens would help better define the defensive functions of plants. Finally, the characterization of plants as an alternative host for food-borne pathogens will be critical in developing effective means to prevent their transmission and disease.
Experimental procedures
Plant materials and growth
Arabidopsis thaliana accession Columbia (Col-0) was used for Shigella infection. Briefly, Arabidopsis seeds were surface-sterilized for 2 min in 70% (v/v) ethanol, incubated in 50% household bleach for 10 min, washed extensively with sterile deionized water, and planted on 1/2 Murashige and Skoog (MS) medium (Duchefa Biochemie, Haarlem, Netherlands) supplemented with 1% sucrose and solidified with 0.6% (w/v) agar (Murashige and Skoog, 1962). Nicotiana benthamiana plants were germinated and grown at 22 ± 3°C under a 16 h light/8 h dark cycle in plastic pots containing steam-sterilized mixed soil (2:1:1, v/v/v, soil/vermiculite/perlite) (Moon et al., 2016). To measure the plant immune response in terms of MAPK activity, 1 μM flg22 peptide (#FLG22-P-1; Alpha Diagnostics, Inc., San Antonio, TX, USA) was used as a positive control (Bethke et al., 2009).
Bacterial strains, growth conditions, and plasmids
The bacterial strains and plasmids used in the study are described in S1 Table. Shigella and Pseudomonas strains harboring plasmid pDSK-GFPuv were generated by electroporation, as described previously (Wang et al., 2007; Hong et al., 2016).
Shigella spp. were grown at 37°C in Luria-Bertani (LB) medium or tryptic soy agar containing 0.003% (w/v) Congo red dye (Sigma-Aldrich, St. Louis, MO, USA) (Runyen-Janecky and Payne, 2002). Pseudomonas syringae strains were grown at 28°C (with shaking at 200 rpm) in King’s B liquid medium (Sigma-Aldrich) containing appropriate antibiotics (King et al., 1954). Escherichia coli DH5a was grown in LB medium at 37°C with shaking (Kennedy, 1971) and used as a non-pathogenic control. Agrobacterium tumefaciens GV2260 was grown at 28°C in LB broth with shaking at 200 rpm (Shamloul et al., 2014).
The coding region of ospF or ospG was PCR-amplified using attB-containing PCR primers (S2 Table). The PCR fragments were cloned into the pDONR™207 vector by BP recombination using the Gateway® BP Clonase™ II kit (Invitrogen, Carlsbad, CA, USA). The products were then transferred to pBAV178 (for AvrRpt2 fusion) or pBAV179 (for HA fusion) vectors by LR recombination (Gateway® LR Clonase™ II, Invitrogen). pBAV178, pBAV179, and pME6012 (empty vector control) were kindly provided by Dr. Jean T. Greenberg (University of Chicago) (Vinatzer et al., 2005). The products were also transferred to pK7FWG2 (obtained from Ghent University, Belgium) by LR recombination to produce the GFP-fused Shigella effectors OspF:GFP and OspG:GFP (Karimi et al., 2002).
Bacterial inoculation assay in planta
Arabidopsis seedlings (2 weeks old) grown in 1/2 MS medium were used for flood-inoculation (Ishiga et al., 2011). Briefly, 10 Arabidopsis seedlings in one container were incubated for 3 min with 35 ml of each bacterial strain suspension (5 × 106 or 5 × 105 cfu/ml) containing 0.02% Silwet L-77 (Lehle Seeds, Round Rock, TX, USA) or buffer. After the bacterial suspensions were removed by decantation, plates containing inoculated plants were incubated in a growth room (23 ± 2°C, 16 h light/8 h dark). Bacterial cell counts from inoculated plants were monitored as described previously (Ishiga et al., 2011). Three inoculated seedlings in one container were harvested by cutting the hypocotyls, and total fresh weight was measured. The cfu were normalized to cfu/mg using sample weight. The cfu of seedlings in three separate containers (as biological replicates) were measured. In addition, the bacterial population was evaluated in more than three independent experiments conducted successively under the same conditions.
To assess root invasion, 10-d-old Arabidopsis seedlings grown vertically in 1/2 MS medium were inoculated by dropping 2.0 μl of bacterial suspension (5 × 107 cfu/ml) onto the root tips. Symptoms were observed under white light, and bacterial proliferation was monitored at 5 dpi by observation of GFP-expressing bacteria under UV light. Three biological replicates were generated in separate plates, and three independent experiments were conducted under the same conditions.
Microscopy
For SEM, flood-inoculated Arabidopsis leaves were fixed in 4% (w/v) paraformaldehyde and dehydrated in an ethanol series (30%, 50%, 70%, 96%, and 100%). The fixed leaves were then dried, coated with gold-palladium, and visualized using a scanning electron microscope (LEO 1455VP, Oberkochen, Germany) (Plotnikova et al., 2000). For TEM, flood-inoculated Arabidopsis leaves were cut off, fixed overnight in 2.5% (w/v) glutaraldehyde, post-fixed in 2% (w/v) osmium tetroxide, dehydrated in ethanol, and embedded in the resin. After staining in 2% (w/v) uranyl acetate and lead citrate, samples were observed under an electron microscope (Bio-TEM; Tecnai G2 Spirit Twin; FEI, USA) (Chae and An, 2016).
For fluorescence confocal microscopy, inoculated Arabidopsis leaves or roots were washed with sterile water. GFP-labeled bacteria or GFP-tagged Shigella effector proteins in plants was observed under a Nikon laser scanning confocal microscope C2 (Nikon, Tokyo, Japan) using filter sets for GFP (λex, 488 nm; λem, 505‒530 nm) or RFP (λex, 561 nm; λem, 570‒620 nm). For each microscopic method, three leaves were used per treatment and at least three microscopic fields were observed for each leaf, including the control.
Expression of Shigella virulence genes in Arabidopsis plants
Total RNA was extracted from Shigella-infected leaves (from three plants) using RNAiso plus (#9108; TaKaRa, Otsu, Japan), according to the manufacturer’s protocol. RT-PCR was performed using M-MLV reverse transcriptase (Invitrogen), according to the manufacturer’s instructions. Quantitative RT-PCR was carried out in a CFX Connect™ Real Time System (BioRad, Hercules, CA, USA) using iQ™ SYBR® Green Supermix (BioRad) and primers specific for target genes (ipaB, ipaC, icsA, icsB, virB, and virF; S3 Table) (Bando et al., 2010). The qRT-PCR results were normalized to expression of 16s rRNA.
Immunoblotting
Total protein was extracted from Shigella- or Agrobacterium-infected leaves (from three plants) using denaturing extraction buffer [150 mM NaCl, 50 mM Tris-HCl (pH 7.5), 5 mM EDTA, 0.1% Triton-X, 1× protease inhibitor cocktail (Roche, Basel, Switzerland), 0.4 M DTT, 1 M NaF, and 1 M Na3VO3]. The extracted proteins were separated on 12% SDS-PAGE gels and transferred to a PVDF membrane (Pierce, Rockford, IL, USA). Antibodies specific for phospho-p44/p42 MAPK ERK1/2 (#4377; Cell Signaling Technology, Danvers, MA, USA), ERK1 (#sc-94; Santa Cruz, Dallas, TX, USA), hemagglutinin (HA) (#S2930; Clontech Laboratories, Mountain View, CA, USA), or GFP (#sc-9996, Santa Cruz) were used for immunoblot analyses. Target proteins were detected using ECL plus reagent (GE Healthcare, Wauwatosa, WI, USA) and visualized using an Alliance 9.7 chemiluminescence imaging system (UVITEC, Cambridge, UK).
Agrobacterium-mediated transient gene expression
A. tumefaciens strain GV2260 harboring the GFP, OspF:GFP, or OspG:GFP genes driven by the 35S promoter was prepared as described previously (Lee et al., 2013). The inoculum (OD600 = 0.4) was infiltrated into 4-week-old N. benthamiana leaves using a 1 ml needleless syringe. To observe MEK2DD-triggered cell death suppression by Shigella effectors, Agrobacterium (OD600 = 0.4) expressing HA:MEK2DD was mixed with Agrobacterium containing GFP, OspF:GFP, or OspG:GFP at a 1:1 ratio and infiltrated into the leaves of N. benthamiana. Each experiment was repeated using at least three leaves from the plant, and each experiment included at least three different plants.
Statistical analysis
All data are expressed as the mean ± SD. The statistical significance of bacterial cell growth in infected plants was examined using Student’s t-test (Microsoft Office Excel) and ANOVA (SPSS v.18; IBM, Armonk, NY, USA) (Moon et al., 2016). Asterisks and letters indicate significant differences between samples (P < 0.05).
Acknowledgments
We thank Dr. Myung Hee Kim for initial support with the preparation of pathogenic Shigella strains, Dr. Jean T. Greenberg for providing the pBAV178, pBAV179, and pME6012 plasmids, and Dr. Cha Young Kim for providing the HA:MEK2DD clone.
The authors declare no conflict of interest.
References




