

ABSTRACT
Soft-tissue sarcomas are rare, heterogeneous and often aggressive mesenchymal cancers. Many of them are associated with poor outcome, in part because biomarkers that can reliably identify high-risk patients are lacking. Studies on sarcomas often are limited by small sample sizes rendering the identification of novel biomarkers difficult when focusing only on individual cohorts. However, the increasing number of publicly available ‘omics’ data opens inroads to overcome this obstacle.
Here, we combine high-throughput transcriptome analyses, immunohistochemistry, and functional assays to show that high adenosine monophosphate deaminase 2 (AMPD2) is a robust prognostic biomarker for worse patient outcome in undifferentiated pleomorphic sarcoma (UPS). Publicly available gene expression and survival data for UPS from two independent studies, The Cancer Genome Atlas (TCGA) and the CINSARC reference dataset, were subjected to survival association testing. Genes, whose high expression was significantly correlated with worse outcome in both cohorts (overall and metastasis-free survival), were considered as prognostic marker candidates. The best candidate, AMPD2, was validated on protein level in an independent tissue microarray. Analysis of DNA copy-number and matched gene expression data indicated that high AMPD2 expression is significantly correlated with copy-number gains at the AMPD2 locus. Gene-set enrichment analyses of AMPD2 co-expressed genes in both UPS gene expression datasets suggested that highly AMPD2 expressing tumors are enriched in gene signatures involved in tumorigenesis. Consistent with this prediction in primary tumors, knockdown of AMPD2 by RNA interference with pooled siRNAs or a doxycycline-inducible shRNA construct in the UPS cell line FPS-1 markedly inhibited proliferation in vitro and tumorigenicity in vivo.
Collectively, these results provide evidence that AMPD2 may serve as a novel biomarker for outcome prediction in UPS. Our study exemplifies how the integration of available ‘omics’ data, immunohistochemical analyses, and functional experiments can identify novel biomarkers even in a rare sarcoma, which may serve as a blueprint for biomarker identification for other rare cancers.
INTRODUCTION
Soft-tissue sarcomas comprise a broad range of mesenchymal tumors1, which are often characterized by high relapse and metastasis rates2,3. For many sarcoma entities there are presently no prognostic biomarkers available that could help in tailoring individualized therapy4. Therefore, most patients are treated with radical and sometimes mutilating surgical resections, combined with various chemotherapeutic and/or irradiation protocols2,3.
Despite the general rarity of sarcomas, The Cancer Genome Atlas (TCGA) consortium provides a multidimensional genetic and clinically annotated dataset for various rare tumor entities including undifferentiated pleomorphic sarcoma (UPS)5, which constitutes a major clinical challenge6.
The diagnosis of UPS is established for high-grade malignant neoplasms characterized by tumor cells with diffuse pleomorphism and the absence of a specific line of differentiation1. Most UPS occur in the extremities at an age of over 60 years7. Although these sarcomas are heterogeneous, they are commonly aggressive. While the 5-year overall survival is around 60%, metastasis-free survival in the same time interval is only around 30%8. In fact, UPS tend to frequently metastasize, especially to the lungs (40-50%)1. The standard treatment for UPS comprises limb sparing resection combined with either neoadjuvant and/or adjuvant radiotherapy9. So far, no clear benefit of other and more aggressive therapy regimens has been proven7,9, possibly due to the lack of prognostic markers that can discriminate high-risk from low-risk patients.
In this study, we combined bioinformatic analyses of the TCGA and an additional gene expression dataset for UPS, immunohistochemical analysis of a tissue microarray (TMA), and in vitro and in vivo experiments to probe for robust prognostic markers in UPS. We found that high expression of the adenosine monophosphate deaminase 2 (AMPD2), which is involved in purine metabolism10, is associated in all independent cohorts with worse patient outcome, possibly by promoting proliferation of UPS cells.
MATERIALS AND METHODS
Retrieval and processing of gene expression and clinical data
Normalized mRNA expression levels from RNA sequencing (RNA-seq) data of UPS were retrieved together with corresponding clinical annotations and further normalized genetic readouts (copy number variation) from the TCGA data portal. RNA expression data from the CINSARC (complexity index in sarcomas) microarray analysis of UPS (GSE21050, Affymetrix HG-U133-Plus2.0 microarray)11 were downloaded from the Gene Expression Omnibus (GEO). The corresponding clinical data for GSE21050 were extracted from the attached series matrix file. All CEL files were manually inspected to be generated on the same microarray and simultaneously normalized with Robust Multi-array Average (RMA)12 including background correction, between-array normalization, and signal summarization using a custom brainarray chip description file (CDF; v19 ENTREZG), yielding one optimized probe set per gene13. Patient characteristics for both cohorts are given in Table 1.
Characteristics of the UPS cohorts
Analysis of association with patient outcome in gene expression datasets
For both RNA expression datasets patients were stratified in two subgroups defined by either high or low expression of a given gene (cut-off: median expression level of the given gene). Differential event-free survival in both subgroups was assessed in each cohort independently with the Mantel-Haenszel test, a time-stratified modification of the log-rank test. Events were defined in the TCGA cohort as ‘death’, and in the GSE21050 cohort as ‘occurrence of metastasis’. This analysis was performed automatically for all 17,536 genes represented in both expression datasets with an in-house software. Those genes whose high expression showed a concordant and significant (P < 0.01; without correction for multiple testing) association with worse event-free survival in both cohorts, were manually inspected and displayed using the Kaplan-Meier method in the GraphPad Prism 5 software.
Linear regression analysis for copy-number variations (CNVs) and gene expression
To evaluate if AMPD2 expression is regulated by CNVs in UPS, the log2 transformed segment mean values for the copy-number at the AMPD2 locus and the AMPD2 expression of the TCGA UPS cohort were displayed and subjected to linear regression analysis in GraphPad Prism 5 software.
Gene Set Enrichment Analysis (GSEA)
To identify gene sets that are enriched among AMPD2-co-regulated genes, all genes in both gene expression datasets were ranked by their Pearson correlation coefficient with AMPD2 expression and a pre-ranked GSEA with 1,000 permutations was performed14.
Immunohistochemistry and immunoreactivity scoring
A tissue microarray (TMA) provided by T. Knösel comprising 83 UPS, each represented by two cores (1 mm in diameter), was used to analyze the correlation of AMPD2 expression levels with overall survival. Antigen retrieval was carried out by heat treatment with ProTaqs IV Antigen Enhancer (401602392, Quartett, Berlin, Germany). Next, the TMA slides were stained with a polyclonal anti-AMPD2 antibody raised in rabbit (1:1,300; HPA050590, Atlas Antibodies, Bromma, Sweden) for 60 min at RT, followed by a monoclonal secondary horseradish peroxidase (HRP)-coupled horse-anti-rabbit antibody (ImmPRESS Reagent Kit, MP-7401, Vector Laboratories, Burlingame, USA). AEC-Plus (K3469, Agilent, Santa Clara, USA) was used as chromogen. Samples were counterstained with hematoxylin (H-3401, Vector Laboratories). The specificity of the used anti-AMPD2 antibody was validated by staining of xenografts from an UPS cell line (FPS-1) with an experimentally induced knockdown of AMPD2 (Fig. 4). In the TMA, the AMPD2 immunoreactivity was scored by a consultant pathologist (T. Knösel), who was blinded to the clinical data. Cytosolic AMPD2 immunoreactivity was classified into ‘no’, ‘low’, ‘intermediate’, and ‘strong’. Assignment of the scoring results to the clinical data was performed by an independent statistician (H. Altendorf-Hofmann). Association with overall survival was calculated with the Kaplan-Meier method and Mantel-Haenszel test in SPSS.
Formalin-fixed and paraffin-embedded (FFPE) xenografts of the FPS-1 UPS cell line were stained with hematoxylin and eosin (H&E). AMPD2 staining was carried out as described above. For cleaved caspase-3 staining, antigen retrieval was carried out by heat treatment with Target Retrieval Solution Citrate pH6 (S2369, Agilent). Then, slides were incubated with the polyclonal cleaved caspase-3 primary antibody raised in rabbit (1:100; 9661, Cell Signaling, Frankfurt am Main, Germany) for 60 min at RT followed by ImmPRESS Reagent Kit. DAB+ (K3468, Agilent) was used as chromogen, hematoxylin for counterstaining. Ki-67staining was performed with a VENTANA BenchMark system (Roche, Basel, Switzerland) with an ultraView detection kit (Roche) and a monoclonal mouse anti-Ki-67 antibody (M7240, Agilent).
Cell lines and cell culture conditions
The FPS-1 UPS cell line was established and described previously15. HEK293T cells were obtained from ATCC (Manassas, USA). Both FPS-1 and HEK293T cells were grown in RPMI 1640 medium (Merck, Darmstadt, Germany), supplemented with 15% or 10% tetracycline-free fetal bovine serum (Biochrom, Berlin, Germany), respectively, and 100 U/ml penicillin and 100 µg/ml streptomycin (Merck) in a humidified 5% CO2 atmosphere at 37°C. Cells were subcultured every two to seven days, detaching the cells with trypsin/EDTA (Merck). Cells were only used up to 15 passages, checked routinely for the absence of mycoplasma by nested PCR, and maintenance of cell line identity was verified by STR-profiling.
RNA interference (RNAi)
Transient suppression of AMPD2 expression was achieved by transfection of FPS-1 cells with 10 nM of an siPOOL (siTOOLs, Planegg, Germany) consisting of 30 different short interfering RNAs (siRNAs) targeting AMPD2 (sipAMPD2), which virtually eliminates off-target effects16. A commercial non-targeting siPOOL (siTOOLs) was used as a control (sipControl). The siPOOLs were complexed with the transfection reagent HiPerfect (Qiagen, Venlo, The Netherlands) and target cells were reversely transfected. After 24 h the transfection reagent was diluted by doubling the amount of cell culture medium. Knockdown efficacy was validated by quantitative real-time polymerase chain reaction (qRT-PCR).
Generation of FPS-1 cells with an inducible AMPD2 targeting shRNA
To study the effects of AMPD2 knockdown for a longer period of time, we employed the lentiviral pLKO-TET-ON all-in-one vector system containing a puromycin selection cassette17. Herein, we cloned either a non-targeting short hairpin RNA (shControl; 5’-CAACAAGATGAAGAGCACCAA-3’) or an shRNA against AMPD2 (shAMPD2; target sequence 5’-GGGTATCTGGGAAGTACTTTG-3’). Vectors were expanded in Stellar Competent Cells (Clontech, Kyoto, Japan) and clones with integrated shRNA were sequenced by Sanger sequencing to validate the correct shRNA sequence. Lentivirus was produced in HEK293T cells, which were transfected using the Lipofectamine LTX Plus Reagent system (Life Technologies, Darmstadt, Germany). Target cells were infected with filtered supernatant of the HEK293T cells without Polybrene. Before the cells reached confluence, successfully transduced cells were selected with 1 µg/ml puromycin (InvivoGen, San Diego, USA). For induction of shRNA expression, 0.5 µg/ml doxycycline (dox) (Merck) was added to the culture medium.
Analysis of cell proliferation
For assessment of proliferation, 4 × 105 FPS-1 cells were seeded in wells of 6-well plates and transfected with either sipAMPD2 or sipControl. Medium was doubled after 24 h. 48 h after transfection cells were re-transfected. Cells including their supernatant were counted 120 h after initial transfection using the Trypan-Blue (Sigma-Aldrich, Taufkirchen, Germany) exclusion method and standardized hemocytometers (C-Chip, Biochrom). The same assays were carried out with dox-inducible shRNA FPS-1 infectants, in which the AMPD2 knockdown was induced by addition of 0.5 µg/ml dox after 24 h. Doxycycline was refreshed after 72 h.
Analysis of tumor growth in vivo
2 × 106 FPS-1 cells transduced with inducible shRNA against AMPD2 were injected in the right flank of NOD/Scid/gamma (NSG) mice. Tumor growth was measured every second day with a caliper. When most tumors reached an average diameter of 5 mm, mice were randomized into two groups of which one was treated henceforth with 2 mg/ml dox (bela-pharm, Vechta, Germany) dissolved in drinking water containing 5% sucrose (Sigma-Aldrich), and the other one with 5% sucrose only. Mice were sacrificed when the average tumor diameter exceeded 15 mm (stop criterion). Tumors were quickly extracted, small samples were snap frozen in liquid nitrogen prior to RNA isolation, and the remaining tumor tissue was formalin-fixed (4%) and paraffin-embedded for (immuno)histology. Animal experiments were carried out in accordance with recommendations of the European Community (86/609/EEC), local authorities, and the UKCCCR guidelines (guidelines for the welfare and use of animals in cancer research).
RNA extraction, reverse transcription, and qRT-PCR
Total RNA was extracted using the NucleoSpin RNA kit (Macherey-Nagel, Düren, Germany) according to the manufacturer’s protocol. RNA (1 µg) was reversely transcribed with the High Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific, Rockford, USA), according to the manufacturer’s protocol. For qRT-PCR 1:10 diluted cDNA and 0.5 µM forward and reverse primer for AMPD2 and the housekeeping gene RPLP0 were used in SYBR Select Master Mix (Thermo Fisher Scientific) (total reaction volume: 15 µl) in a Bio-Rad CFX Connect Real-Time PCR Detection System (Bio-Rad, Munich, Germany). All primers were purchased from Eurofins Genomics (Ebersberg, Germany). Primer sequences were as follows:
AMPD2 forward 5’-GGTCTCTGCATGTCTCCATTC-3’;
AMPD2 reverse 5’-CTCAATACCTGGGCCATCAG-3’;
RPLP0 forward 5’-GAAACTCTGCATTCTCGCTTC-3’;
RPLP0 reverse 5’-GGTGTAATCCGTCTCCACAG-3’.
The qRT-PCR was carried out with the following thermal conditions: heat activation at 95°C for 2 min, DNA denaturation at 95°C for 10 sec, and annealing and elongation at 60°C for 20 sec (50 cycles), final denaturation at 95°C for 30 sec. AMPD2 expression before and after knockdown was calculated with the Delta-Delta-Cq method.
RESULTS
High AMPD2 mRNA levels correlate with worse outcome in two independent UPS cohorts
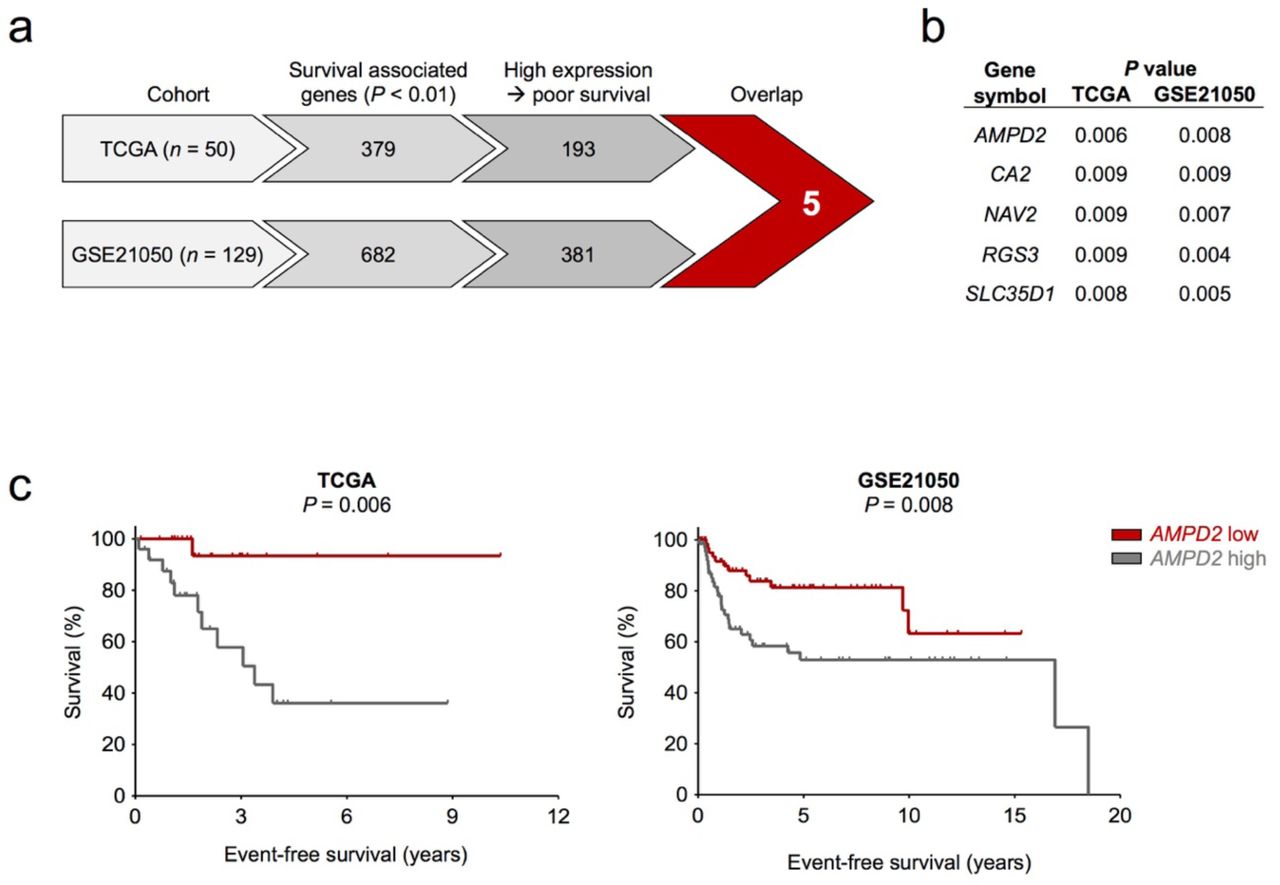
To identify prognostic marker candidates for UPS, we stepwise crossed two independent datasets for which matched clinical data and transcriptome-wide expression profiles were available (Fig. 1a). The first was derived from The Cancer Genome Atlas (TCGA) sarcoma panel (n = 50)5, the second from the CINSARC reference dataset (n = 129; GSE21050)11. We calculated independently for both datasets for each gene (17,536 genes represented in both cohorts) the association with event-free survival (TCGA: event = death; GSE21050: event = occurrence of metastasis) when stratifying each dataset by the median expression of the given gene (see methods section). This yielded 379 genes in the TCGA and 682 genes in the GSE21050 cohort with a P value of < 0.01 (Supplementary Tables 1 and 2). Notably, only five genes were significantly (P < 0.01) and concordantly positively associated with worse event-free survival in both cohorts, namely AMPD2, CA2, NAV2, RGS3, SLC35D1 (Fig. 1b). Interestingly, we noted that for AMPD2 the survival curves in the Kaplan-Meier analyses separated very early, which could indicate a potential biological relevance (Fig. 1c).

a) Workflow of applied filtering steps in the survival analysis and number of genes meeting the criteria.
b) List of the resulting candidates from Fig. 1a and their P values in both cohorts (Mantel-Haenszel test).
c) Kaplan-Meier plots displaying the survival of patients with AMPD2 low versus high expressing tumors in the TCGA and GSE21050 cohort (Mantel-Haenszel test).
High AMPD2 protein levels correlate with worse outcome in UPS
As especially AMPD2 appeared as an interesting candidate, we validated our findings based on the mRNA level by immunohistochemical staining for AMPD2 in a third UPS cohort comprising 83 samples (Table 1) on the protein level. To this end, we semi-quantitatively scored the intensity of AMPD2 immunoreactivity. In this cohort, 39 tumors exhibited no, 23 weak, 10 intermediate, and 11 strong AMPD2 immunoreactivity (Fig. 2a). For correlation with overall survival, samples were grouped in AMPD2-negative (corresponding to no immunoreactivity, n = 39), and AMPD2-positive (corresponding to weak to strong immunoreactivity, n = 44). Strikingly, detection of AMPD2 expression was associated with a significantly worse outcome, and early separation of the two groups in Kaplan-Meier analysis (Fig. 2b). Collectively, these findings made in three independent cohorts provide evidence that AMPD2 may constitute a general and robust prognostic marker in UPS.

a) Representative images of UPS tissue cores with no, low, intermediate, and strong cytosolic AMPD2 immunoreactivity. The number of individual UPS samples showing the corresponding staining intensities is reported.
b) Kaplan-Meier plot displaying the overall survival of UPS patients carrying tumors without AMPD2 immunoreactivity (AMPD2-negative) and those with low to strong immunoreactivity (AMPD2-positive) (Mantel-Haenszel test).
Copy number variations (CNVs) promote AMPD2 overexpression in UPS
Since AMPD2 appeared to be heterogeneously expressed in UPS, and since UPS typically feature high rates of genomic aberrations, we reasoned that CNVs at the AMPD2 locus could cause the variable AMPD2 expression. We therefore correlated CNV information and AMPD2 expression data available for the TCGA cohort. In a linear regression analysis, we observed a highly significant correlation between the AMPD2 expression levels and CNVs at the AMPD2 locus in UPS (rPearson = 0.57; P < 0.001) (Fig. 3a).

a) Dot-plot displaying the median AMPD2 expression levels and copy number segment means at the AMPD2 locus for each patient of the TCGA cohort. The red line represents the linear regression of the data, rPearson = 0.57, P < 0.001.
b) Heat-maps for AMPD2 co-and antiregulated genes in the TCGA and GSE21050 cohort with representative enriched gene sets within AMPD2-co-regulated genes. NES, normalized enrichment score.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
a) Bar plots indicating viable cell count of sipControl (grey) and sipAMPD2 (red) transfected FPS-1 cells relative to control on the left, knockdown control on the right; whiskers indicate standard deviation. Statistical comparison by unpaired two-tailed student’s t-test.
b) Left: bar plots indicating viable cell count of shControl and shAMPD2 transduced FPS-1 cells without (grey) and with (red) shRNA induction relative to control; right: knockdown control. Statistical comparison by unpaired two-tailed student’s t-test.
c) Cumulative tumor volume over time for FPS-1 shAMPD2 xenotransplanted mice treated with doxycycline (dox; red) and controls (grey) and corresponding Kaplan-Meier plot below representing the survival time until average tumor diameter exceeded 15 mm (stop criterion) (Mantel-Haenszel test).
d) Representative micrographs of the FPS-1 shAMPD2 tumors from mice without (left) and with (right) AMPD2 knockdown induction with dox (H&E, AMPD2, Ki-67, and cleaved caspase-3 staining; 20X magnification, scale bar = 200 µM).
e) Bar plot for AMPD2 knockdown validation for the FPS-1 shAMPD2 tumors from mice treated with dox in drinking water by qRT-PCR. Whiskers indicate standard deviation. Statistical comparison by unpaired two-tailed student’s t-test.
To get first clues on the potential biological relevance of CNV-mediated AMPD2 overexpression in UPS, we performed gene-set enrichment analysis (GSEA) on AMPD2 co-expressed genes in the TCGA and GSE21050 cohort. The gene-sets with highest normalized enrichment score (NES) were highly concordant in both cohorts (Supplementary Tables 3 and 4) and indicated that high AMPD2 expression in UPS is associated with gene signatures involved in hypoxia and tumor growth (Fig. 3b).
AMPD2 may promote proliferation and tumor growth of UPS
As our GSEA in primary tumors suggested that AMPD2 may have a functional role in growth of UPS, we aimed at validating this prediction in a cell line model. To this end, we took advantage of an established UPS cell line (FPS-1), which recapitulates key tumor features of UPS in vivo15 and performed knockdown experiments. In a first step, we serially transfected the FPS-1 cells with an siPOOL consisting of 30 specific siRNAs against AMPD2 (sipAMPD2) and compared their proliferation with that of cells transfected with a non-targeting control siPOOL (sipControl). Transfection with the sipAMPD2 reduced AMPD2 expression levels onto around 20% after 120 h, which was accompanied by a significant reduction of viable cells by around 30% (Fig. 4a). To validate this finding in vivo, we transduced FPS-1 cells with lentivirus containing a doxycycline (dox)-inducible short hairpin RNA (shRNA) against AMPD2 (shAMPD2; pLKO-TET-ON all-in-one system17) or a non-targeting control shRNA (shControl). While dox-treatment had no effect on AMPD2 expression and proliferation in FPS-1 shControl cells, treatment of FPS-1 shAMPD2 cells with dox significantly reduced AMPD2 expression under 40% and efficiently impaired cell proliferation in vitro (Fig. 4b). We next injected FPS-1 shAMPD2 cells subcutaneously into the flanks of immunocompromised NSG mice and monitored tumor growth over time. Once most tumors reached an average diameter of 5 mm, we randomized the mice and henceforth treated half of them with dox in the drinking water (2 mg/dl). Strikingly, AMPD2 knockdown significantly delayed tumor growth (Fig. 4c), which was accompanied by a significant reduction in proliferating cells as indicated by IHC stain for Ki-67, while it did not induce apoptotic cell death as verified by staining for cleaved caspase-3 (Fig. 4d). The knockdown of AMPD2 was confirmed in each xenograft ex vivo by IHC and by qRT-PCR (Fig. 4d,e), which underscores the specificity of the used anti-AMPD2-antibody. Collectively, these results suggest that AMPD2 may promote proliferation and tumor growth of UPS.
DISCUSSION
For many sarcoma subtypes including UPS, standard treatment regimens are often insufficient to achieve long-term disease control, resulting in local recurrence and/or metastasis, to which many patients will ultimately succumb2,18–20. To identify high-risk patients early on and to assign them upfront to adequate therapy regimens, novel prognostic and predictive biomarkers are urgently required. Although the diversity and rarity of sarcomas pose general obstacles for conducting statistically reliable biomarker studies in these cancers21, the integration of publicly available ‘omics’ data and functional assays might help overcoming this problem.
In the current study, we took a functional genomics approach by combining public gene expression datasets, immunohistochemistry, and cell-based in vitro and in vivo assays to identify AMPD2 as a new prognostic candidate marker for UPS.
The relative small sample size of 50 in the TCGA cohort harbors the risk to miss genuine prognostic markers in survival analysis when correcting hundreds of potential candidates for multiple testing. We therefore consciously relinquished such statistical correction but chose instead, to only accept those genes as prognostic marker candidates with overlapping results in a second independent cohort. In these analyses, we focused on genes whose high expression correlated with worse outcome as they might also represent new therapeutic targets. While clinical TCGA data comprised the time of overall survival, the microarray dataset (GSE21050) contained information on metastasis-free survival. As metastasis is the predominant cause for disease specific death22, both datasets compared complimentary aspects of patient outcome. These analyses highlighted AMPD2 as the most promising biomarker candidate, which was fully validated in a third independent cohort on the protein level.
AMPD2 is the liver isozyme of three known AMP deaminases, converting AMP to IMP, which is crucial for purine metabolism10,23. So far, the protein was reported to be linked with liver pathology24,25, but not with cancer26. In our FPS-1 UPS model, silencing of AMPD2 by RNA interference markedly reduced proliferation and tumor growth in vitro and in vivo without inducing apoptotic cell death. These findings suggest a functional relevance of AMPD2 in UPS, possibly through deregulation of the purine metabolism. Conversely, the frequently observed copy-number gains at the AMPD2 locus and subsequent overexpression of AMPD2 in UPS primary tumors may facilitate tumor growth, and thus contribute to worse patient outcome. Future studies have to further characterize the functional role of AMPD2 in UPS, and to test whether it may additionally serve as a drug target for therapeutic blockage of increased purine metabolism. Also, additional work is necessary to validate the potential prognostic value of the other biomarker candidates that were identified in our transcriptome analyses (Fig. 1b).
Taken together, our data suggest that AMPD2 is a promising novel prognostic biomarker for UPS. We therefore recommend its validation in prospective studies. Furthermore, our study highlights the importance of including even rare entities such as UPS in ongoing cancer genomics projects such as TCGA, and our functional genomics approach may serve as a blueprint for identification and validation of additional biomarkers for other rare cancers.
Author contributions
M.F.O. and T.P.G.P. conceived the study, wrote the paper and designed the figures. M.F.O. performed bioinformatic analyses and experiments. J.S.G. programmed the in-house survival analysis tool. T.Kn. scored IHC staining of TMAs and A.A.H. correlated the results with clinical data. J.M., R.A.R., S.S., G.S., S.O., J.L, M.D., M.C.B., and A.M. supported the experimental procedures. M.H. provided the FPS-1 cell line. T.Ki. provided lab infrastructure and histological guidance. E.B., T.D., and T.P.G.P. supervised the project and provided scientific advice. All authors read and approved the final version of this manuscript.
FUNDING
M.F.O. and M.C.B were supported by a scholarship of the German National Academic Foundation, J.M. by a scholarship of the Kind-Philipp-Foundation, and M.D. by a scholarship of the Deutsche Stiftung für junge Erwachsene mit Krebs. The laboratory of T.G.P.G. is supported by grants from the ‘Verein zur Förderung von Wissenschaft und Forschung an der Medizinischen Fakultät der LMU München’ (WiFoMed; to T.G.P.G.)’, by LMU Munich’s Institutional Strategy LMUexcellent within the framework of the German Excellence Initiative (to T.G.P.G.), the ‘Mehr LEBEN für krebskranke Kinder – Bettina-Bräu-Stiftung’ (to T.G.P.G.), the Matthias-Lackas-Stiftung (to T.G.P.G.), the Dr. Leopold und Carmen Ellinger Stiftung (to T.G.P.G.), the Wilhelm-Sander-Foundation (2016.167.1 to T.G.P.G.), the German Cancer Aid (DKH-111886 and DKH-70112257 to T.G.P.G.), and the Deutsche Forschungsgemeinschaft (DFG 391665916 to S.O. and T.G.P.G.).
CONFLICT OF INTEREST
The authors declare no conflict of interest.
ACKNOWLEDGEMENTS
We thank Mrs. Andrea Sendelhofert and Mrs. Anja Heier for excellent technical assistance.
Abbreviations
- CDF
- chip description file
- dox
- doxycycline
- FTPE
- formalin fixed paraffin embedded
- GEO
- gene expression omnibus
- GSEA
- gene set enrichment analysis
- H&E
- hematoxylin and eosin
- NES
- normalized enrichment score
- NSG
- NOD/Scid/gamma
- qRT-PCR
- quantitative real time polymerase chain reaction
- TCGA
- The Cancer Genome Atlas
- TMA
- tissue microarray
- UPS
- undifferentiated pleomorphic sarcoma
REFERENCES




