

ABSTRACT
Human papillomavirus (HPV) accounts for hundreds of thousands of new cases of cervical cancer yearly, and half of these women die of this neoplasia. This study investigates the possibility of HPV16 to infect human peripheral blood leukocytes in ex vivo assays. We have developed a leukocyte separation method from heparinized blood samples aiming cellular integrity and viability. We have expressed humanized L1 and L2 viral capsid proteins in HEK293T epithelial human cells, transiently transfecting them with vectors encoding humanized HPVL1 and L2 genes. Recombinant L1/L2 capsid proteins and structured virus-like particles interacted with human peripheral blood mononuclear cells – lymphocytes and monocytes – and were internalised through a pathway involving CD71 transferrin receptors. This was observed, at a percentile of about 54% T-CD4, 47% T-CD8, 48% B-CD20, and 23% for monocytes-CD14. The group of polymorph nuclear cells: neutrophils-eosinophils-basophils group did not internalise any VLPs. Blockage assays with biochemical inhibitors of distinct pathways, like chlorpromazine, rCTB, filipin, nystatin, liquemin, and sodium azide also evidentiated the occurrence of virus-like particles’ indiscriminate entrance via membrane receptor on mononuclear cells. This study shows that HPV16 L1/L2 VLPs can interact with the plasma membrane surface and successfully enter lymphocytes without requiring a specific receptor.
IMPORTANCE Human papillomaviruses (HPVs) belong to the Papillomaviridae family and are classified in Alpha papillomavirus, Beta papillomavirus, Gamma papillomavirus, and Mu papillomavirus genera, based on DNA sequence of the L1 gene. They are associated with the development of benignant skin warts, cell transformation and malignant tumours. L1 is its major capsid protein, and L2 is the minor HPV capsid protein. HPV type 16 is considered a high-risk Alpha papillomavirus due to the association with 50% of cervical cancers worldwide. The most frequent HPV-associated cancer type is cervical cancer, but etiological association has also been demonstrated for carcinomas of the penis, vulva, vagina, anus and oropharynx – including base of the tongue and tonsils – regions. This study indirectly investigates the possibility of HPV16 to infect other cell types in vitro, particularly human peripheral blood leukocytes.
INTRODUCTION
In 1974, zur Hausen’s researches pointed to the HPV as the major aetiological agent in cervical cancer and, later, its DNA was detected in tumours found in other anogenital regions (1–3). Today, there are more than 200 different types of well characterized HPVs, of which approximately 40 types infect the genital tract. The most frequently found are types 6 and 11 that induce benignant skin warts formation, and types 16 and 18 which are associated to cancer development. There are at least 12 HPV types that according to the Agency for Research on Cancer (IARC) are considered oncogenic to humans (4, Papillomavirus Episteme (PaVE); http://pave.niaid.nih.gov/#home). Among them, HPV16 and HPV18 are those responsible for approximately 70% of all cervical cancers (4, 5). In 2025, the projected global estimate of cervical cancer is expected to rise to 720,415 new cases per year, and it is expected that half of these women will die of this neoplasia (5). However, the difficulty to obtain enough viable wild types or recombinant HPV particles has limited researches to distinct aspects of virus biology (6). All viruses enter host cells to survive, replicate and evade the immune system. So far, the entry of HPV and its traffic through the host cells are still not completely elucidate (7). Several studies describe this process like a complex set of interactions among different pathways, receptors, co-receptors and co-factors. In keratinocytes, HPV seems to be internalised via clathrin-dependent endocytic mechanisms, but it might use alternative uptake pathways to enter cells, such as a caveolae-dependent route, among others depending on viral type (6–9).
Currently, HPV is recognised as one of the main causes of infection-related cancer worldwide, as well as the causal factor of other diseases. Infection with high-risk HPV types is the aetiological cause of cervical cancer and is strongly associated with a significate fraction of penile, vulvar, vaginal, anal and oropharyngeal cancers (10, 11). HPVs are responsible for approximately 88% of anal cancer and 95% of anal intraepithelial neoplasia grades 2/3 lesions, and 40%-50% of penis and vulvar cancers (10). In oropharyngeal cancers, HPV DNA was detected in 35%-50% cases (10). In all HPV-positive non-cervical cancers, HPV16 is the most common HPV type detected, followed by HPV types 18, 31, 33 and 45. Among the non-cancerous HPV-associated conditions, genital warts and recurrent respiratory papillomatosis are surely linked to HPV6 and 11 (10). In addition, it was demonstrated recently that HPV16 virus-like particles (VLPs) L1/L2 interact with haematopoietic precursor stem cells, present in the amniotic fluid from healthy pregnant women (12). Whereas new pathologies are increasingly being associated to the HPVs; the responsibility and costs of HPV-associated diseases and cancer remain an important public health issue in all countries, regardless of their economic developmental level (13). Due to the continuous worldwide propagation of HPV it is necessary to investigate the possibility of HPV internalisation by different human cell types. In the present study, we addressed the capacity of HPV16 L1/L2 VLP entrance/uptake of – in peripheral blood leukocytes in vitro.
MATERIAL AND METHODS
Production of HPV16 L1/L2 VLPs
The production of VLPs was carried out throughout the recombinant protein expression HPV16 L1 and L2 in epithelial human cells of the HEK293T lineage, cultured as previously described (14). Transfection of these cells occur in a transient way using pUF3L1h and pUF3L2h vectors, which are regulated by the human cytomegalovirus promoter containing complete sequences of genes L1 and L2 – a method already described in detail (14, 15). In this study, besides being used to produce VLPs containing both capsid proteins, HEK293T cell line was also used as a control factor on interaction assays with human peripheral blood leukocytes. All assays performed in this study were analysed in duplicate, being representative of at least four independent tests.
Blood collection
Blood samples from 10 healthy female volunteers, ages ranging from 35 to 55, were requested based on epidemiological data associated with genital HPV infection (16). The screening of the volunteers was based on recent blood, Pap smears and colposcopy tests and on data filled out by the candidate on a written informed consent form and on a questionnaire, for the purpose of laboratory research. Volunteers who did not present Pap and colposcopy results within reference values were dismissed. Blood samples were collected in sterile tubes containing heparin – for interaction assays with leukocytes – and EDTA – for blocking assays –, and were processed quickly, within a maximum of 2-hours after collection. During this interval, they were stored at 4°C.
Leukocytes’ separation
Whole blood was collected in heparin at a concentration of 0.1 mg/ml, centrifuged at 1,000 rpm for 7 minutes at 5°C in a Sorvall® RT 6000 Refrigerated Centrifuge (Du Pont, Wilmington-DE, USA) with a horizontal angle rotor. The plasma supernatant containing platelets and leukocytes was gently removed and transferred with the aid of a Pasteur pipette to a new tube. An aliquot was collected for total and differential counts, performed in a Neubauer chamber, diluted (ratio 1:1) in Trypan blue and smears were stained by May-Grünwald-Giemsa in order to determine their composition. Leukocytes sedimented were centrifuged again at 1,200 rpm, for 3 minutes, at 5°C, to remove platelets. Leukocytes precipitates were resuspended in 0.85% saline solution for subsequent tests (17). The Neubauer chamber counts and smears containing distinct cell types were analysed by light microscopy, with a Leica DMIL I microscope (Leica Microsystems GmbH, Vienna, AUT).
Interaction assays of HEK293T cells and HPV16 L1/L2 VLPs
HEK293T cells (2×104 cells/ml) were plated and maintained under growth conditions, washed with PBS and incubated with 120 µg of VLPs in DMEM without FBS, for 4-hours at 37°C and 5% CO2. After this, cells were washed twice with PBS for 3 minutes each in order to remove non-interactive particles. Cells were then fixed with 2%PFA (Paraformaldehyde, Sigma-Aldrich) in PBS for 1-hour, at 4°C, and washed three times with PBS, 5 minutes each. Plates were kept at 4°C until immunofluorescence assays. Controls were performed in the absence of VLPs and/or the denaturation thereof, by heating at 100°C for 10 minutes (18, adapted).
Interaction assays of human leukocytes and HPV16 L1/L2 VLPs
Leukocytes (2×104 cells/ml) of healthy volunteers were incubated with 120 µg of VLPs produced in this study and RPMI without FBS for 4-hours at 37°C and 5% CO2, under gentle agitation. Cells were centrifuged at 1,000 rpm for 7 minutes and cell pellets washed three times with PBS, 5 minutes each. Then, cells were fixed with 2% PFA in PBS for 1-hour, at 4°C. After this step, the centrifugation and PBS washing processes were repeated. Samples were kept at 4°C until immunofluorescence assays. Controls were the same as described above for HEK293T cells.
Interaction assays of leukocytes, transferrin and HPV VLPs
The same protocol described above was employed, with minor changes described below. Leukocytes were incubated with 120 µg of VLPs in separate samples, as follows: the HPV16 L1/L2 VLPs produced in this study, and L1 VLPs of HPV6, 11, 16 and 18 from Gardasil® vaccine, used as control, kindly provided by Merck Sharp & Dohme. RPMI without FBS, together with transferrin (Tf) conjugated to fluorochrome TexasRed® (Molecular Probes™), were added (ratio 1:60, Tf:RPMI). Samples were kept for 15, 45, 60 and 120 minutes in an incubator at 37°C and 5% CO2, under gentle agitation. The other procedures remained unchanged.
Blocking assays for membrane receptors
Leukocytes of healthy donors were counted and incubated in RPMI medium overnight over coverslips containing poly-L-lysine (Sigma-Aldrich), at 37°C and 5% CO2. Then, cells were washed and a fresh medium was placed together with specific biochemical inhibitors of ligand uptake (Table 1), used isolated or associated, and incubated for 2 hours (18). After incubation, cells were washed and 120 µg of HPV16 L1/L2 VLPs (19) were added and incubated with medium for 4 hours. Cells were washed again and fixed in 2% PFA solution. Immunofluorescence assays were carried out to detect the VLP-PBMC interaction by confocal microscopy, using specific antibodies to recognise L1 and L2 proteins. Z-axis 3D images were obtained to confirm the presence of VLPs within cells.
Biochemical inhibitors used to examine their effects on the entry of HPV VLPs in PBMC.
Immunofluorescence assays for internalisation analysis
Leukocytes were washed and fixed as already described. Samples were incubated in PBS containing 1% BSA for 5 minutes under gentle agitation. Later, cells were incubated with the primary antibodies (suppl. Data – Table 1); diluted in PBS containing 0.01% Tween® 20 and 0.5% BSA – pH 8 – for 2 hours, under gentle agitation, at room temperature. They were then washed three times with PBS for 10 minutes each, followed by incubation with the corresponding secondary antibodies (suppl. Data – Table 2); conjugated with fluorochromes; and diluted in PBS containing 0.01% Tween® 20 and 1.5% BSA for 1 hour, under light stirring, at room temperature. Once more, they were washed three times with PBS for 10 minutes each. Samples of leukocytes with transferrin and VLPs were labelled with Phalloidin conjugated AlexaFluor® 594, incubated for 20 minutes. Immediately after that, they were rinsed twice with PBS for 10 minutes each. Aliquots of cell suspension from both items assayed were adhered over silanized slides, and mounted with 5 µl Mowiol® and coverslips. Samples were kept at 4°C until CLSM analysis, using Confocal Laser Scanning Microscope Zeiss 510 Meta of the Butantan Institute (FAPESP Process No. 2000/11624-5; Carl Zeiss GmbH, Jena, DEU).
Interactions of HPV16 VLPs produced in this study with PMBC (mononuclear cells) from peripheral blood of healthy female volunteers.
RESULTS
Leukocytes’ identification
After isolation of leukocytes from healthy donors, control smears stained by May-Grünwald-Giemsa method showed cell morphology preservation (17), presenting a positive correlation with the control blood smears. Counts indicated approximately 98% lymphocytes; 0.7% monocytes; 1.5% polymorph nuclear cells; 0.1% red blood cells and 0.1% platelets. The ultrastructure was well preserved for all cell types analysed, with well-defined and intact membranes (17). The efficiency and speed of the method for obtaining leukocytes, preserving morphology and cell viability was confirmed.
Analysis of interaction between HEK293T cells and HPV16 L1/L2 VLPs
HEK293T cells interaction with HPV16 L1/L2 VLPs was analysed by immunofluorescence. We emphasize that the genome of this cell line does not contain any HPV DNA sequences, but it contains Adenovirus DNA and SV40 T-Ag. After 4-hours at a 37°C interaction, cells were washed in order to remove any particles or proteins that did not interacted with HEK293T cells. Then, cells were fixed and stained with the primary antibody Camvir-1 (BD Biosciences – suppl. Data – Table 1) for L1 proteins and with L1 conformation-specific anti-VLP antiserum (Biodesign – suppl. data – Table 1). It was therefore possible to observe the internalisation of VLPs in some cells. In these cells VLPs were found in the cytoplasm, near the core region (suppl. data – Img. 1 A-D), stained with anti-VLP. These results were confirmed by the overlap of images (suppl. data – Img. 1 A and C), and by the most thoroughly detailed internalisation display on the Z-axis scanning sections (suppl. data – Img. 1 B and D). The morphological evaluations show that probably the VLPs’ internalisation occurs simultaneously with a large number of particles, similar to the formation of endocytic vesicle structures (suppl. data – Img. 1 C, white arrow). These results suggest that after 4 hours, structured VLPs (suppl. data – Img. 1 A-D), as well as non-structured VLPs – such as pentameric and monomeric forms of L1 stained with Camvir-1 (suppl. data – Img. 1 E) –, were internalised in epithelial cells from the human kidney (HEK293T), across the cell membrane (suppl. data – Img. 1 B and D). The negative controls showed no immunostaining for L1 and VLPs and no changes in cell morphology were detected (suppl. data – Img. 2).
Analysis of interactions between human leukocytes and HPV16 L1/L2 VLPs
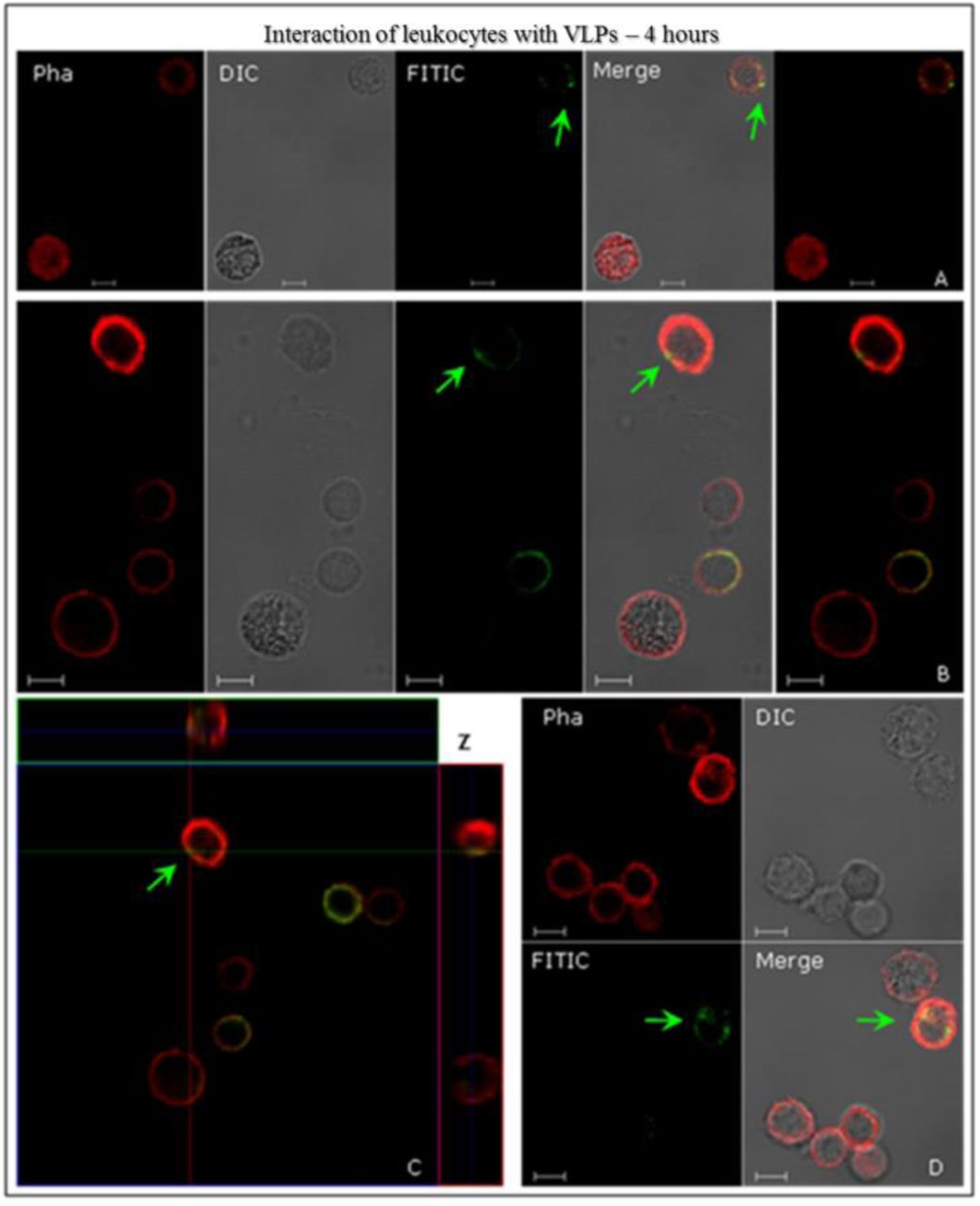
L1/L2 particles were added to human leukocytes, for 4-hours at 37°C, and prepared for indirect CLSM immunofluorescence analysis, in order to investigate the possibility of interaction between the leukocytes and the HPV16 L1/L2 VLPs. Results indicate an interaction of VLPs with peripheral blood mononuclear cells (PBMC) – T and B lymphocytes and monocytes – from healthy women volunteers (Fig. 1). Structured HPV16 L1/L2 VLPs also interacted with and were internalised by leukocytes (Figs. 2 and 3). After 4-hours of interaction, in most leukocytes examined, these particles were found in the cell cytoplasm, as was the case with HEK293T. These results were confirmed by the images’ overlap (Figs. 1 A, B and D; Figs. 2 and 3), and the detailed internalisation, as shown on the Z-axis scanning sections (Fig. 2 C). Fig. 1 C (green arrow) shows VLPs across the cell membrane. The Z-axis’ sweep cuts were able to demonstrate the internalisation of the VLP with a larger number of particles similar to the formation of endocytic vesicles structures (Fig. 2 A, white arrow), like those found in HEK293T cells (suppl. data – Img. 1 C, white arrow), a phenomenon endorsed by morphological evaluations. In Fig. 3, the interaction of leukocytes with the VLPs is illustrated, showing the colocalisation of HPV16 L1/L2 proteins recognised by anti-L1 (green) and anti-L2 (red), respectively. This colocalisation was expected in VLPs that are composed of these two proteins, as it might be seen from the images’ overlap (Fig. 3). Fig. 3 B shows the internalisation of VLPs by leukocytes in a significant amount when compared to other interactions. At least 15 fields were analysed per experiment, containing from 4 to 10 cells per field. The presence of structures similar to vacuoles can also be observed (Fig. 3 B, blue arrow). Control assays showed no immunostaining, neither with the antibodies used to detect L1 and L2 separately, nor with structured HPV16 L1/L2 VLPs (suppl. data – Img. 3).
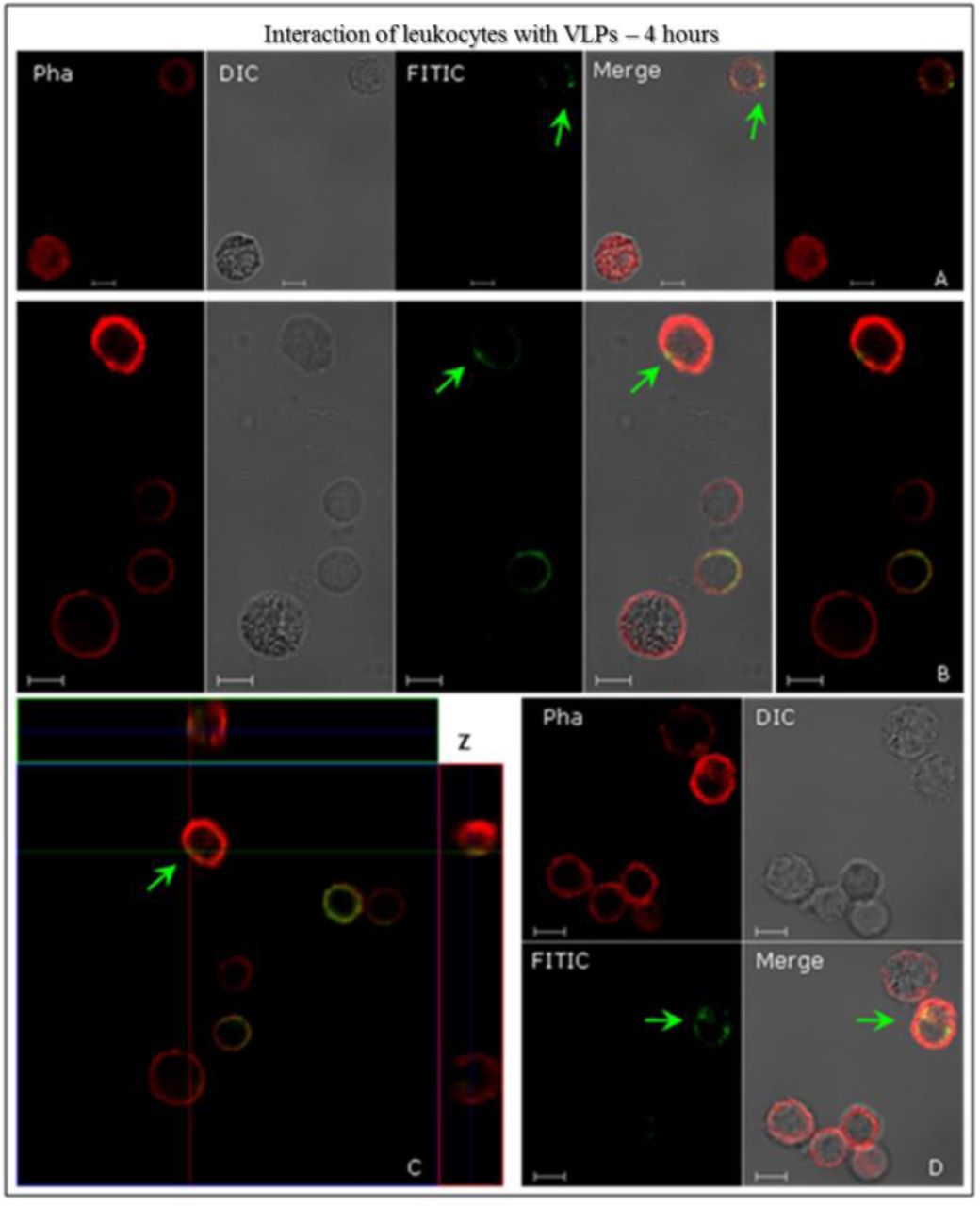
Interactions of human leukocytes with VLPs of HPV16, detected by indirect immunofluorescence. The leukocyte cells were incubated with the VLPs for 4 h. Following fixation, they were immunostained with anti-HPV16 L1 antibody and then stained with a secondary antibody conjugated to FITC (green). In all assays, AlexaFluor® 594-conjugated Phalloidin (red) detected the actin cytoskeleton. The internalisation of VLPs (green arrows) is shown in image C, through the sweep cuts on the Z-axis (C) 9.50 µm thick sections and each section to 1.60 µm. CLSM Zeiss LSM 510 Meta. Magnification: Objective C-Apochromatic 63xs /1.4 oil. Bar = 5 µm.

Interactions of human leukocytes with VLPs of HPV16, detected by indirect immunofluorescence. The leukocyte cells were incubated with the VLPs for 4 h. Following fixation, they were immunostained with the conformational anti-HPV16 VLPs and stained with a secondary antibody conjugated to FITC (green). AlexaFluor® 594-conjugated Phalloidin (red) detected the actin cytoskeleton. The internalisation of VLPs (green arrows) is shown in image C, through Z-axis sweep cuts (C) 7.80 µm thick sections and each section to 1.30 µm. In (A) we see a structure similar to an endocytic vesicle (white arrow). CLSM Zeiss LSM 510 Meta. Magnification: Objective C-Apochromatic 63xs /1.4 oil. Bar = 5 µm.

Interactions of human leukocytes with VLPs of HPV16, detected by indirect immunofluorescence. After fixation, cells were immunostained with anti-HPV16 L1 antibody and then stained with a secondary antibody conjugated to FITC (green). Then immunostained with anti-HPV16 L2 antibody and revealed by AlexaFluor®555 secondary antibody (red). The internalisation of VLPs (yellow arrows) is shown in confocal images (merge). In (B) structures similar to vacuoles (blue arrow) might be observed. CLSM Zeiss LSM 510 Meta. Magnification: Objective C-Apochromatic 63xs /1.4 oil. Bar = 5 µm.
Identification of human PBMC interactions with VLPs
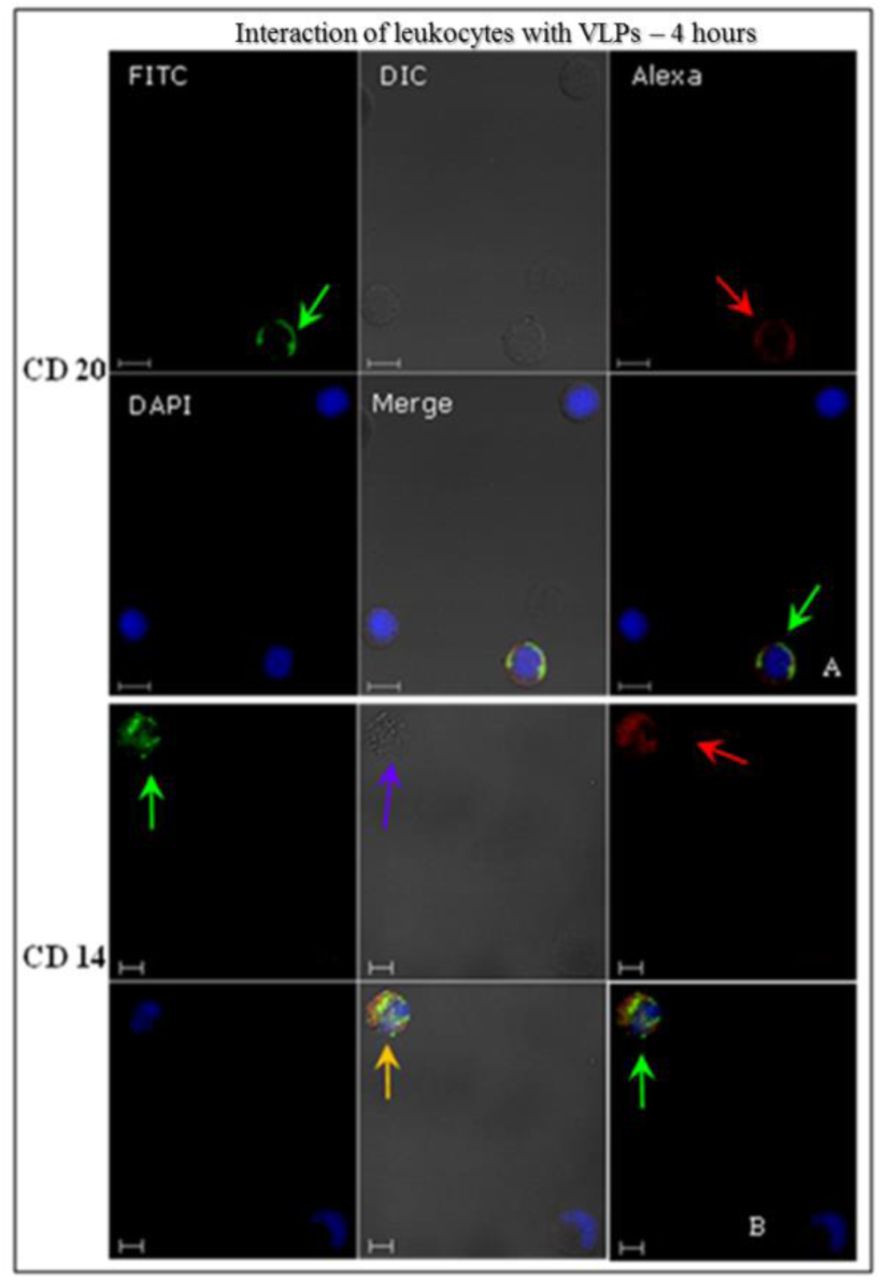
These cells were treated with antibodies in order to recognise specific cell membrane receptors for each type (suppl. data – Table 1), and analysed by indirect immunofluorescence using CLSM to identify PBMC interactions with HPV16 VLPs, after 4-hours incubation at 37°C. HPV16 VLPs interacted with ex vivo PBMC (Figs. 4-6). These results were confirmed by the images’ overlap (Figs. 4 and 5) and displayed detailed internalisation on the Z-axis sections (Fig. 6). T and B lymphocytes showed greater competence to internalise VLPs, at around 47%-52% of cells (Table 2). Through morphological assessments, T-lymphocytes that internalised VLPs and were recognised by anti-CD8 showed endocytic vesicles-like structures (Fig. 4 B, white arrow). Only 23% of monocytes that were identified by anti-CD14 antibody interacted with the VLPs (Table 2).
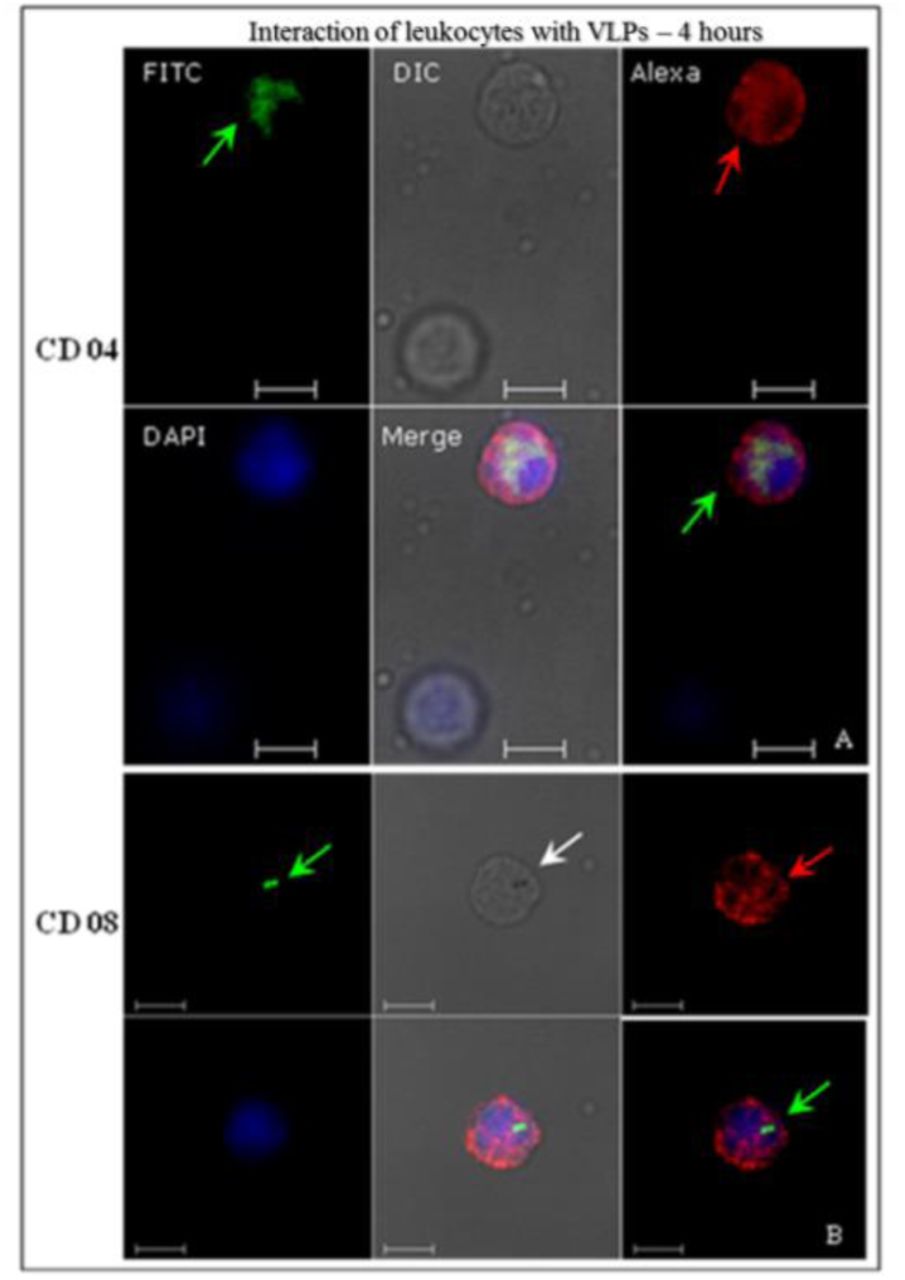
Identification of human leukocyte interactions with the HPV16 VLPs analysed by indirect immunofluorescence. After fixation, cells were immunostained with the conformational anti-HPV16 VLPs and revealed by a secondary antibody conjugated to FITC (green). Then, the cells were treated with anti-CD04 (A) and anti-CD08 (B) and stained with a secondary antibody AlexaFluor®546 (red). VLP internalisation (green arrows) is visible on image overlay (merged images). In (B), a structure similar to an endocytic vesicle is detectable (white arrow). CLSM Zeiss LSM 510 Meta. Magnification: Objective C-Apochromatic 63xs /1.4 oil. Bar = 5 µm.

Identification of human leukocyte interactions with the HPV16 VLPs, analysed by indirect immunofluorescence. After fixation, cells were immunostained with the conformational anti-HPV16 VLPs and revealed by a secondary antibody conjugated to FITC (green). Then, the cells were treated with anti-CD20 and anti-CD14 and stained with a secondary antibody AlexaFluor®546 (red). VLP internalisation (green arrows) is visible on image overlay (merged images). In (B) we can observe the colocalisation of CD14 and VLPs (orange arrow) and vacuole-like structures (blue arrow). CLSM Zeiss LSM 510 Meta. Magnification: Objective C-Apochromatic 63xs /1.4 oil. Bar = 5 µm.

Identification of human leukocyte interactions with the HPV16 VLPs analysed by indirect immunofluorescence. After fixation, cells were immunostained with the conformational anti-HPV16 VLPs and revealed by a secondary antibody conjugated to FITC (green). Then, the cells were treated with anti-CD20 (C), anti-CD04 (A), anti-CD08 (B), anti-CD14 (D) and AlexaFluor®546 revealed by secondary antibody (red). VLP internalisation (green arrows) is shown in the images through Z-axis sweep cuts. (A) 9.25 µm of thick sections and each section of 0.84 µm; (B) 8.56 µm thick sections and each section with 0.95 µm; (C) 9.27 µm thick sections and each section with 1.03 µm; (D) 10,94 µm thick sections and each section having 1.40 µm. CLSM Zeiss LSM 510 Meta. Magnification: Objective C-Apochromatic 63xs /1.4 oil.
Analysis of the colocalisation of VLPs and transferrin (Tf) in human PBMC
After 15 minutes, no colocalisation between L1 VLPs and exogenous Tf was detected (Fig.7). Moreover, it was possible to show the internalisation of VLPs during this period. After 45 minutes, colocalisation of VLPs with exogenous Tf in the cytoplasm of human PBMC (Fig. 8 and 9, yellow arrows), and also with the TfR or CD71, was observed (Fig. 9 A-B, yellow arrows) suggesting that the iron pathway may be used to HPV internalisation. All results were confirmed by the images’ overlap, respectively (Figs. 7-9).

Colocalisation of VLPs of HPV16 and Tf in human leukocytes, detected by indirect immunofluorescence. The leukocyte cells were incubated with the VLPs and the conjugated Tf TexasRed® (red). Leukocytes were treated with anti-HPV16 L1 antibody, revealed by a secondary antibody conjugated to FITC (green). In confocality (merge), the images reveal colocalisation of L1 and Tf (yellow arrows). CLSM Zeiss LSM 510 Meta. Magnification: Objective C-Apochromatic 63xs /1.4 oil. Bar = 5 µm.
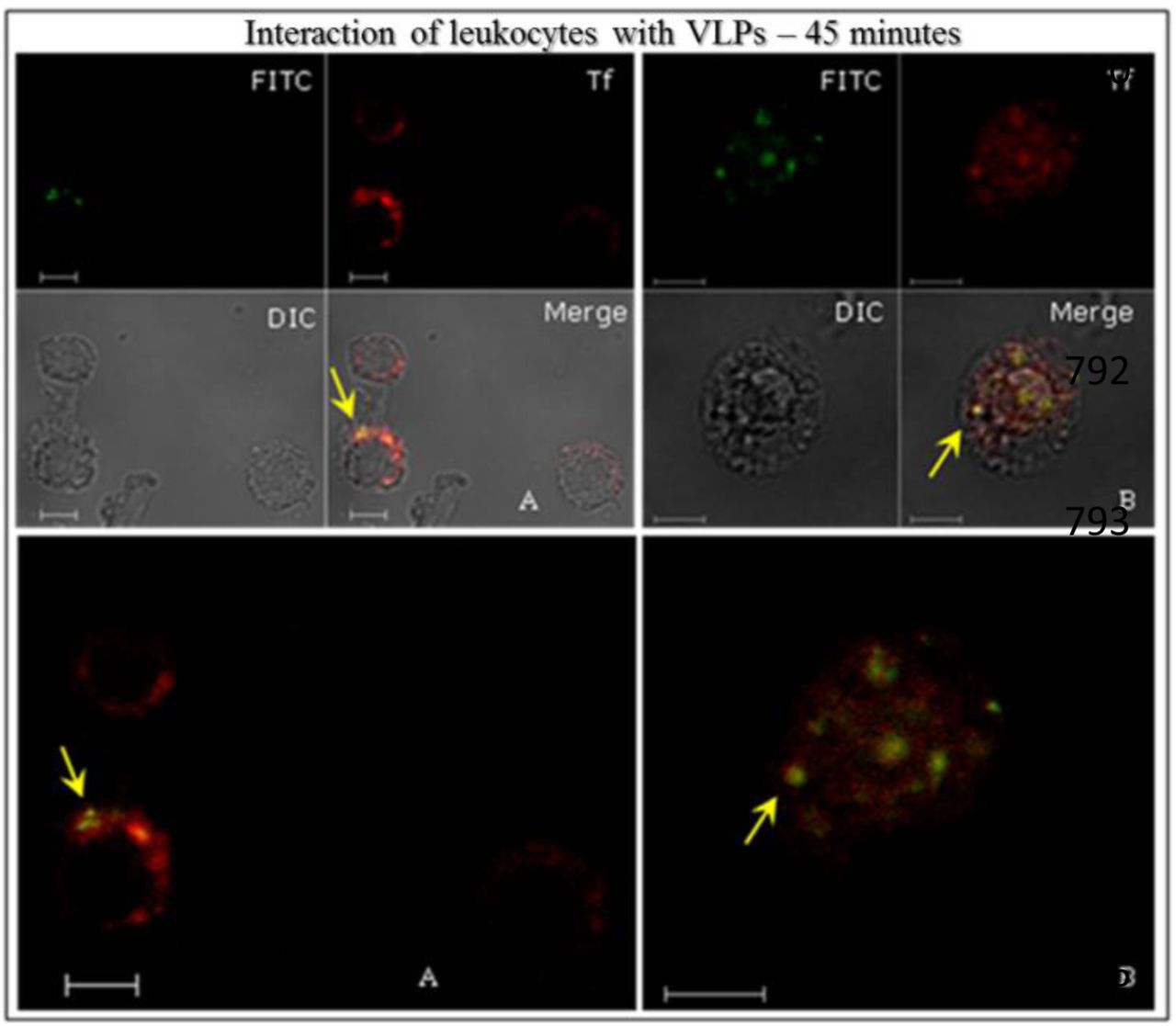
Colocalisation of HPV16 VLPs and Tf in human white blood cells, detected by indirect immunofluorescence. The leukocyte cells were incubated with the VLPs and the conjugated Tf TexasRed® (red). Leukocytes were immunostained with anti-HPV16 VLP antibody and revealed by a secondary antibody conjugated to FITC (green). In confocality (merge), the images reveal colocalisation of VLPs and Tf (yellow arrows). CLSM Zeiss LSM 510 Meta. Magnification: Objective C-Apochromatic 63xs /1.4 oil. Bar = 5 µm.

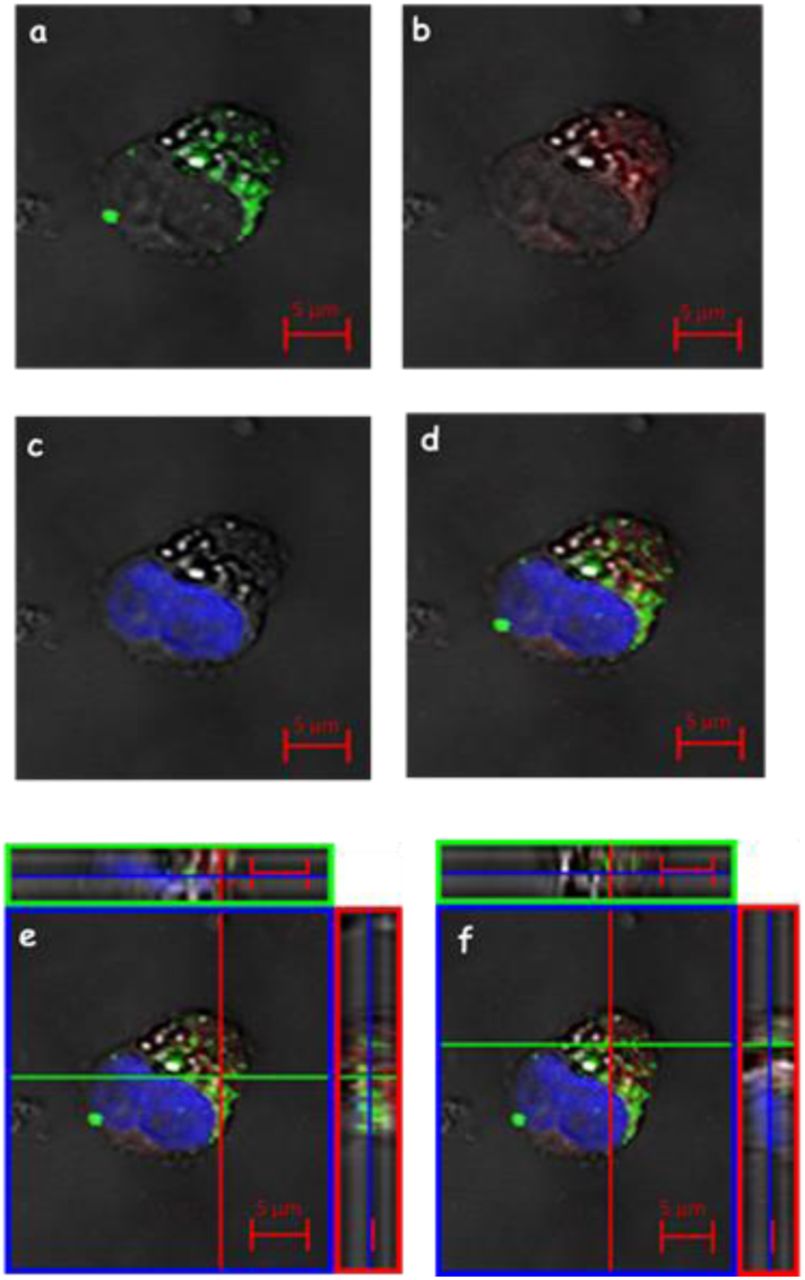
Three-dimensional imaging of leukocyte interactions with the VLPs produced in this study or the VLP vaccine Gardasil, Tf and TfR; detected by indirect immunofluorescence. The leukocyte cells were incubated with the VLPs and the conjugated Tf TexasRed® (red). After fixation, the leukocytes were immunostained with anti-HPV16 VLP antibody and revealed by a secondary antibody, conjugated to FITC (green), then treated with anti-CD71 and revealed by AlexaFluor®633 (blue). (A) and (B) colocalisation Tf and TfR (purple arrow), (C) and (D) colocalisation VLP / Tf / TfR (white arrows), (E) and (F) weak colocalisation VLP / Tf (orange arrows). CLSM Zeiss LSM 510 Meta. Magnification: Objective C-Apochromatic 63xs /1.4 oil.
We compared the kinetics of internalization of Tf and TfR (CD71) with the colocalisation of HPV16 L1/L2 VLPs produced in this study and with control HPV6, 11, 16, 18 L1 VLPs derived from the Gardasil® vaccine (Fig. 10). After only 15 minutes of interaction it was possible to observe colocalisation of TfR and Tf (Fig. 10 A-B, purple arrows) with both L1/L2 VLPs (Fig. 10 A) as well as with Gardasil L1 VLPs (Fig. 10 B). By means of morphological analysis, we were able to demonstrate the colocalisation of both VLPs – L1 and L1/L2 – with Tf and TfR in the leukocytes’ cytoplasm, after a 45-minutes interaction period (Fig. 10 C-D, white arrows). This colocalisation among VLPs, Tf and TfR was found in approximately 50% of analysed cells. Within 60 minutes of interaction, we observed a feeble colocalisation between Tf and VLPs in about 30% of the cells (Fig. 10 E-F, orange arrows). No colocalisation was observed in the tests after 120 minutes. The colocalisation was evidentiated in detail by three-dimensional figures stacked in Z-axis (Fig. 10). The negative controls showed no signs of L1 and VLPs. However, it was possible to observe exogenous Tf internalisation (suppl. data – Img. 4). Comparative testing of L1/L2 VLPs produced in this study and of control L1 VLPs from Gardasil® vaccine showed no significant differences in the analysed results.

Immunofluorescence assay detecting the HPV VLPs entry in PBMC, even in presence of specific biochemical inhibitors of ligand uptake: (1) Sodium azide; (2) Chlorpromazine; (3) Liquemin; (4) rCTB; (5) Filipin; (6) Nystatin. (a) shows the overlay (merge) of images detected by stain of L1 protein (green), L2 protein (red), nuclei (blue); (b) Z-axis showing L1 protein within cells; (c) Z-axis showing the entry of L2 protein in cells was not inhibited. CLSM Zeiss LSM 510 Meta. Magnification: Objective C-Apochromatic 63xs /1.4 oil. Bar = 5 µm.
Analysis of the blocking assays of PBMC membrane receptors
After successful identification of VLPs within leukocytes and their colocalisation with Tf and TfR, a variety of biochemical inhibitors, known to inhibit distinct cellular processes, was used to demonstrate the involvement of these and other different pathways of VLPs entry in PBMC.
Chlorpromazine inhibits clathrin-mediated endocytosis of various plasma membrane proteins. Nystatin is a sterol-binding agent that disassembles caveolae in the membrane. rCTB (Clostridium toxin B) acts by shortening the actin filaments’ length, by inhibiting its process in vitro. Liquemin is related with the VLPs obstruction of internalisation through the HSPG (Heparan sulphate proteoglycans) pathway. Sodium azide is an ATPase inhibitor, which blocks particle movement towards the cell body and leads to a diffuse random movement.
The use of these different biochemical inhibitors was not sufficient to block the HPV16 L1/L2 VLPs entry in the PBMC (Figs. 11-12). Chlorpromazine – although known for blocking the clathrin pathway through CD71 receptor of Tf – was not able to inhibit VLPs entrance inside PBMC in a satisfactory manner. The same occurred when rCTB, Filipin and Nystatin, Liquemin and Sodium azide were used to block actin-dependent cell processes, the caveolae pathway, the HSPG pathway, and endocytic pathways, respectively, either assayed separately or associated in an all-inhibitors cocktail. These results suggest that PBMC makes use of multiple internalisation pathways to uptake HPV16 particles.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Assay performed with all inhibitors used in Fig.15 (1–6), in association. (a) Detection of L1 protein (green); (b) L2 protein within cells (red); (c) nuclei (blue); (d) the overlay (merge) of images (a, b and c); Z-axis showing the L1 (e) and L2 (f) proteins within cells. CLSM Zeiss LSM 510 Meta. Magnification: Objective C-Apochromatic 63xs /1.4 oil. Bar = 5 µm.
DISCUSSION
Differently of many Beta and Gamma papillomavirus, Alpha papillomaviruses developed immune system evasion strategies from the host, causing persistent and visible papillomas, which sometimes evolving to cancers (20). Some HPV types including 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58 and 59 have been classified as human carcinogens by the IARC, being responsible for at least 4% of all human malignancies (4, 21, Papillomavirus Episteme (PaVE); http://pave.niaid.nih.gov/#home). Infection by high-risk HPV types is the major factor for the development of cervical cancer, and HPV DNA is also found in tumours affecting other anogenital regions. Besides, they are being correlated with an increasing proportion of oropharyngeal squamous cells carcinomas, and occurrences affecting mainly tonsil and base of tongue regions are currently being documented in particular geographic areas (22). A study describes women who have never experienced sexual intercourse as having genital lesions caused by HPV, showing that, in addition to sexual transmission, could have other forms of infection such as fomites and skin contact, and maternal-foetal vertical transmission is also being considered as a probable infection pathway (23–27).
Cell-virus interaction is the initial step in viral infection. Often, virus adsorption occurs on the plasma membrane of susceptible cells. In general, the most common solution is that the pathogen’s intracellular trafficking occurs through and existing entry mechanism, mediated via clathrin, caveolin, macropinocytosis, among others, moving inside cells to reach the location where viral replication occurs (28). Transcytosis is used to move antigens and protective antibodies across epithelial barriers. Similar to what seems to occur with HIV (29), the transcytosis of HPV through epithelial cells, crossing cellular barriers, should also be dependent on trafficking to the endocytic recycling pathway. The productive life cycle of HPV is directly linked to epithelial differentiation. It is believed that the papillomavirus is capable of maintaining the expression of structural genes, under tight control of the regulatory mechanisms of transcription and translation (30). The L1 capsid protein of HPV, regardless of the presence or absence of L2, has the property of forming structures that mimic morphologically authentic virions, VLPs, which are being used as replacements for HPV infected cell culture in vitro (8, 14, 31, 32). However, the presence of L2 seems to confer greater stability to the virus and VLP, therefore, the viral capsid indeed seems to contribute to the virus’ infectivity (33). Most experimental models explore the process of host cell interaction by molecular or biochemical methods, using cell lines from different tissues and species in interactions with HPV L1 VLPs in their studies.
In the present work, we produced HPV L1/L2 VLPs (14, 15) to investigate the possibility that these particles interact with human leukocytes, in order to understand possible mechanisms involved in these interactions. We have developed a method for the isolation of these cells (17), used in suspension at 37°C, thereby creating an ex vivo system as closely as possible to the natural conditions of HPV16 infection, thus conceiving a novel experimental model.
HPV16 recombinant proteins L1, L2 and VLPs were visualized in the perinuclear region of HEK293T cells, in compliance with data described in the literature (34). The presence of BPV (bovine papillomavirus) in lymphocytes has been discussed (35, 36). Using PCR technique (Polymerase Chain Reaction) in samples taken from humans with anogenital lesions, the presence of HPV16 in peripheral blood samples (37) and in plasma cells (38) was detected. Besides, DNA of other HPV types has been found in PBMC (39, 40). The presence of HPV in lymphocytes of patients treated for anogenital cancers suggests that the virus can remain in these cells even after treatment. However, there are no convincing evidences that support that lymphocytes carry/produce infectious HPV viral particles. Nonetheless, the presence of HPV macromolecules in lymphocytes may play a role in viral immune response.
The scientific literature discloses that papillomavirus exhibits specific tropism for species and tissues. However, BPV1 particles, isolated from bovine warts, and HPV16 VLPs interacted successfully with 14 cell lines derived from different species and tissues, as demonstrated in two different studies (32, 41). Using our model to investigate this, HPV16 L1/L2 VLPs interacted in vitro and ex vivo with two very distinct human cells, epithelial kidney cells and PBMC, respectively. After 4 hours of interaction, these particles’ internalisation occurred, showing a very similar distribution pattern on both cell types. In both, L1 VLPs were located in the cytoplasm near the nucleus, suggesting that tissue differences do not affect the ability of HPV16 VLPs to entry in different cell types, and that pentameric and monomeric forms of L1 may also be internalised. The interaction between VLPs and monocytes observed in this study showed colocalisation with CD14. As macrophage precursors, apparently these cells are able to internalise particles at a much larger amount if compared with lymphocytes. The unexpected colocalisation of CD14 membrane receptor and VLPs, as well as the morphological changes observed, suggest that monocytes respond to this interaction as immune cells, in order to eliminate invading microorganisms. CD14 is a 53-kDa molecule, expressed mainly in monocytes, macrophages and granulocytes. It is also found as soluble protein in the serum (42). One known function of this receptor is to bind the complex of LPS (bacterial lipopolysaccharide). Macrophages expressing CD14 can respond to very small amounts of LPS, and this interaction is required for activation of these cells (43). Some studies suggest that the receptor CD16 (FcγRIII), present on the plasma membrane surface of neutrophils, can bind to VLPs at mice splenocytes’ surface, indicating that this receptor has a role to play in immune response against viral capsid proteins (43, 44). It is necessary to clarify the actual mechanisms involved in the interactions between CD14, CD16 and VLPs, in order to better understand how the immune system can properly eliminate the HPV from the host. Tf is a beta globulin responsible for the transport of iron (Fe), through plasma, to the interior of cells. Besides hepatocytes, other cells in the body are able to synthesize it. The TfR is a membrane protein expressed primarily in T and B lymphocytes, macrophages, and proliferating cells such as homodimers with N-terminal cytoplasmic tails (42). TfR is constantly being endocytosed by plasma membrane through clathrin-mediated endocytosis, with the purpose of carrying ferric iron bound to Tf to the inside of cells. This Tf-TfR complex is delivered to the early endosome into a lower pH medium, where iron is released and TfR is recycled to the cell’s surface (45). When evaluating the interactions between HPV16 L1/L2 VLPs, HPV6, 11, 16, 18 L1 VLPs (Gardasil® vaccine) and unstructured HPV16 L1 in PBMCs and in the presence of exogenous Tf, we determined the colocalisation of L1-VLP-Tf and free Tf in these cells, as well as colocalisation with TfR, after 45 minutes of interaction. This colocalisation between VLP, Tf and TfR was found in approximately 50% of analysed cells. These results suggest that HPV16 L1/L2 VLPs, HPV6, 11, 16, 18 L1 VLPs (Gardasil® vaccine) and unstructured HPV16 L1 can enter cells by endocytosis through clathrin-dependent, via TfR. It is probable that, along viral infection processes, interactions between HPV16 (about 55 nm size) and Tf molecules (80-kDa) might indeed happen, through fusion with Fe-Tf-TfR complexes, mimicking its entry into the cell via low pH endosomes, thus enabling transcytosis (29, 46–49).
There have been no reports in the literature showing the internalisation of HPV VLPs in human PBMC. So far, this study has showed that PBMCs of healthy female volunteers were able to internalise HPV16 VLPs. Also, T and B Lymphocytes were the main cell types that further interacted with VLPs. It is important to emphasize that the lymphocytes are cells capable of dividing themselves and it is estimated that the half-life of these inactive cells in humans is of some years. Furthermore, inactive lymphocytes circulate continuously through the bloodstream and lymphatic vessels. Using different cell lineages with VLPs or PsV (pseudovirion), scientists have come to different conclusions. Our findings agree with the data obtained by other researchers who – through the use of PsV-infected HeLa and HaCaT cells – successfully demonstrated that the HPV entry is clathrin-caveolin-cholesterol-and-dynamin-independent (7). The entry of HPV VLPs in leukocytes by independent endocytosis pathways may be due to their ability to recognise, capture and remove foreign substances from the body. The use of different biochemical inhibitors, isolated or associated, does not seem to be enough to block the internalisation of HPV16 L1/L2 VLPs in the PBMC, suggesting that a specific receptor is not required to the uptake and HPV16 entry into the host cells. A recent study demonstrates that intrinsic risk factors are responsible of around 10%-30% of cancer development risk, and that the rates of endogenous mutation accumulation by intrinsic processes are not sufficient to account for the high cancer risks observed. Cancer risk in general is heavily influenced by extrinsic factors like HPV infection (50). Unravelling the molecular steps leading to virus entry into the cells may be important for the development of new strategies for the prevention of infection by HPV and foster immune response against important human pathogen.
CONCLUSIONS
HPV16 L1/L2 VLPs produced in this study interacted with in vitro HEK293T cells and ex vivo PBMC from healthy women volunteers, and were internalised. This study evinced that HPV16 L1/L2 VLPs can interact with the plasma membrane surface, and been uptake by lymphocytes. We also demonstrated that HPV16 L1/L2 VLPs might utilize the endocytic pathway of iron-mediated clathrin as a receptor, which involves multiple membrane receptors simultaneously and also the classical endocytic pathway of iron – the most widespread among vertebrates and the animal kingdom as a whole – we have demonstrated that HPV16 L1/L2 VLPs does not require a specific receptor in order to be internalised by leukocytes. Complementary studies are required to accurately demonstrate the probable mechanisms involved.
Ethical Committee
This work was duly evaluated and approved by the Sao Joaquim Hospital, Real and Meritorious Portuguese Beneficence Association Ethical Committee on Human Research. Protocol number: 369-08.
This work is in full compliance with the National Biosafety Law (CTNBio) guidelines and the present study was approved by the Butantan Institute Institutional Biosafety Committee (CIBio) and CTNBio – Process Nr. 01200.004893/1997-93 – published in the Brazilian Official Gazette (DOU) on August 16, 2011.
ACKNOWLEDGEMENTS
The authors would like to express their deepest gratitude to the CAPES Master’s Degree Fellowship to V. Szulczewski and E.A. Kavati, from the Biotechnology Interunits Postgraduate Programme USP-IBU-IPT; PAP-SES-FUNDAP Fellowships to T.M. Hosoda; Financial Support: FAPESP – Process Number 2004/15122-5; Butantan Foundation and Butantan Institute; to the Prof Dr Richard B.S. Roden, from the Department of Pathology, Johns Hopkins University, Baltimore, MD, USA, which kindly provided us anti-L2 antibody, and for useful comments on the manuscript.
Footnotes
Grant support: This work was supported by grants from the Foundation for Research Support of the State of Sao Paulo – FAPESP (Process number 2004/15122-5), Butantan Institute and Butantan Foundation, Sao Paulo, SP, Brazil.
Conflicts of interest: The authors disclose no potential conflict of interest with the publication of this article.
Authors email: aurora.cianciarullo{at}butantan.gov.br, emefjmlisboa{at}prefeitura.sp.gov.br, erica.kavati{at}butantan.gov.br, hosodatania{at}yahoo.com.br, eleao{at}ibest.com.br, borelli{at}usp.br, eboccardo{at}usp.br, martin.mueller{at}dkfz.de, bkaranam{at}mytu.tuskegee.edu, wbecak{at}yahoo.com
References




