

Abstract
Background and Purpose: Cystic Fibrosis (CF) is caused by mutations in the CF Transmembrane conductance Regulator (CFTR), the only ABC transporter functioning as a channel. Unique to CFTR are two highly conformationally dynamic regions: the regulatory extension (RE) and regulatory insertion (RI). Removal of the latter rescues the trafficking defect of CFTR with F508del, the most common CF-causing mutation.
We aimed here to assess the impact of RE removal (alone or with RI or genetic revertants) on F508del-CFTR traffic and how CFTR modulator drugs corrector VX-809/lumacaftor and potentiator VX-770/ivacaftor rescue these combined variants so as to gain insight into the mechanism of action (MoA) of these drugs.
Experimental Approach. We generated ∆RE and ∆RI CFTR variants (with and without genetic revertants) by site-directed mutagenesis and used them to stably transfect BHK cell lines. We studied CFTR expression and stability by Western blotting and pulse-chase respectively, plasma membrane levels by cell surface biotinylation and channel activity by the iodide efflux technique.
Key Results. Our data demonstrate that ∆RI significantly enhanced rescue of F508del-CFTR by VX-809. Thus, while the presence of the regulatory insertion seems to be precluding full rescue of F508del-CFTR processing by VX-809, this region appears essential to rescue its function by VX-770, thus suggesting some contradictory role in rescue of F508del-CFTR by these two modulators. Nevertheless, this negative impact of RI removal on VX-770-stimulated currents on F508del-CFTR can be compensated by deletion of the regulatory extension which also leads to the stabilization of this mutant. We thus propose that, despite both these regions being conformationally active, RI precludes F508del-CFTR processing while RE affects mostly its stability and channel opening.
Supporting Information: Additional figures with supplementary data
Introduction
Cystic Fibrosis (CF), a life-threatening recessive disorder affecting ~80,000 individuals worldwide, is caused by mutations in the gene encoding the CF transmembrane conductance regulator (CFTR) protein present at the apical membrane of epithelial cells. This is the only member of the ATP-binding cassette (ABC) transporter family, functioning as a channel, actually a cAMP-dependent chloride (Cl-)/ bicarbonate (HCO3-) channel. It consists of two membrane-spanning domains (MSD1/2), two nucleotide binding domains (NBD1/2) and a cytoplasmic regulatory domain (RD), which is unique to CFTR.1 The MSDs are linked via intra- and extra-cellular loops (ICLs, ECLs, respectively). ATP binding promoting NBD1:NBD2 dimerization and subsequent phosphorylation of RD at multiple sites lead to channel gating2. The NBD1:NBD2 and ICL4:NBD1 interfaces were shown to represent critical folding conformational sites 3–5 important for the gating and maturation of the CFTR protein 5,6.
Although >2,000 CFTR gene mutations were reported (http://www.genet.sickkids.on.ca/cftr/app), one mutation F508del occurs in 85% of CF patients. This NBD1 mutant fails to traffic to the plasma membrane (PM) due to protein misfolding and retention by the endoplasmic reticulum quality control (ERQC) that targets it to premature proteasomal degradation. F508del-CFTR folding is a complex and inefficient process but it can be rescued, at least partially, by several treatments. These include low temperature incubation7, genetic revertants4,5,8–13 or pharmacological agents, like “corrector” VX-809 (lumacaftor)14, one of the first CFTR modulator drugs to receive FDA-approved in combination with potentiator VX-770/ivacaftor1. Determining the additive/ synergistic rescue of F508del-CFTR by small molecules correctors together with other rescuing agents/revertants is very valuable to determine the mechanism of action (MoA) of these CFTR modulators5.
Comparative studies of CFTR with other ABC transporters are very powerful to understand the uniqueness of some of its regions, their influence on CFTR maturation and function as well as how they affect distinctive binding of CFTR to modulator drugs. Two such unique regions are present in NBD1 of CFTR, which are absent in NBDs of other ABC transporters (Fig.S1) - the regulatory extension (RE) and regulatory insertion (RI). Both regions were described as highly conformationally dynamic3,16 following PKA phosphorylation at certain (660Ser, 670Ser; 422Ser)18,19. Importantly, removal the 32-amino acid RI (∆RI) was reported to rescue traffic of F508del-CFTR13.
Our first goal here was to assess the impact of removing the 30-amino acid RE (∆RE) alone or jointly with ∆RI on the traffic of F508del-CFTR. Secondly, we aimed to evaluate how RE and or RI removal from F508del-CFTR influences the rescue of this mutant by genetic revertants. Our third and final goal was to determine how the traffic and function of these combined variants of F508del-CFTR (∆RE, ∆RI plus genetic revertants) are rescued by CFTR modulator drugs VX-809 (corrector) and VX-770 (potentiator) to gain further insight into their MoA.
Our data show that although F508del-CFTR without RE did not traffic to the PM, it showed a dramatic stabilization of its immature form (evidencing a turnover rate ~3 × lower than that of wt-CFTR, being the turnover of F508del-CFTR itself is 2x faster vs. wt-CFTR). Results also show that, while ∆RI further increased processing of F508del-CFTR with revertants to almost wt-CFTR levels, RE removal completely abolished their processing, thus highlighting the different impact of the two dynamic regions on revertant-rescued F508del-CFTR. Most strikingly, although VX-809 rescued ∆RI-F508del-CFTR and ∆RE-∆RI-F508del-CFTR processing to wt-CFTR levels, nevertheless to achieve maximal function of F508del-CFTR, removal of just RI was insufficient, as both RI and RE had to be absent from F508del-CFTR. These data indicate that removal of these two regions has a positive effect on the rescuing efficacy of F508del-CFTR by CFTR modulators.
Methods
CFTR variants, cells and culture conditions
Several CFTR deletion variants were produced by site-directed mutagenesis corresponding to the removal of residues: ΔRIL – 404Gly-Leu435; ΔRIS –412Ala-Leu428; ΔREL –647Cys-Ser678; ΔRES –654Ser-Gly673; ΔH9 –637Gln-Gly646; ΔREL-ΔH9 –637Gln-Ser678; ΔRIS-ΔRES –both 412Ala-Leu428 and 654Ser-Gly673; and ΔRIL– ΔRES – 405Gly-Leu435 and 654Ser-Gly673. All the constructs were produced using full length wt-CFTR and F508del-CFTR.
BHK cells lines expressing ΔRIL-, ΔRIL-F508del-, ΔRIL-G550E-, ΔRIL-G550E-F508del, ΔRIL-R1070W-, ΔRIL-R1070W-F508del, ΔRIS-, ΔRIS-F508del, ΔRIS -G550E-, ΔRIS-G550E-F508del, ΔRIS-R1070W-, ΔRIS-R1070W-F508del, ΔRES-, ΔRES-F508del, ΔRES-G550E, ΔRES-G550E-F508del, ΔRES-R1070W-, ΔRES-R1070W-F508del, ΔREL-, ΔREL-F508del, ΔRIS-ΔRE-, ΔRIS-ΔRES-F508del, ΔRIL-ΔRE-, ΔRIL-ΔRES-F508del, ΔH9-, ΔH9-F508del, ΔREL-ΔH9-, ΔREL-ΔH9-F508del-CFTR were produced and cultured as previously described 11. Cells were cultured in DMEM/F-12 medium containing 5% (v/v) FBS and 500μM of methotrexate. For some experiments, cells were incubated with 3μM VX-809 or with the equivalent concentration of DMSO (Control) for 48h at  .
.
Western blot
To study the effect of removal of regulatory extension (RE) and or regulatory insertion (RI) in combination with genetic revertants and VX-809, cells were incubated for 48h at  with 3μM VX-809. After incubation, cells were lysed and extracts analysed by Western blot (WB) using the anti-CFTR antibody (Ab) 596 or anti-calnexin Ab as a loading control. Score corresponds to the percentage of band C to total CFTR (bands B+C) as normalized to the same ratio in samples from wt-CFTR expressing cells. Blot images were acquired using BioRad ChemiDoc XRC+ imaging system and band intensities were measured using Image Lab analysis software.
with 3μM VX-809. After incubation, cells were lysed and extracts analysed by Western blot (WB) using the anti-CFTR antibody (Ab) 596 or anti-calnexin Ab as a loading control. Score corresponds to the percentage of band C to total CFTR (bands B+C) as normalized to the same ratio in samples from wt-CFTR expressing cells. Blot images were acquired using BioRad ChemiDoc XRC+ imaging system and band intensities were measured using Image Lab analysis software.
Pulse-chase and immunoprecipitation
BHK cells lines stably expressing CFTR variants were starved for 30 min in methionine-free a-modified Eagle's medium or minimal essential medium and then pulsed for 30min in the same medium supplemented with 100μCi/ml [35S]methionine. After chasing for 0, 0.5, 1, 1.5, 2 and 3h in a-modified Eagle's medium with 8% (v/v) fetal bovine serum and 1mM non-radioactive methionine, cells were lysed in 1ml of RIPA buffer [1% (w/v) deoxycholic acid, 1% (v/v) Triton X-100, 0.1% (w/v) SDS, 50mM Tris, pH7.4, and 150mM NaCl]. The immunoprecipitation (IP) was carried out using the anti-CFTR antibody in independent experiments and Protein G–agarose or Protein A–Sepharose beads. Immunoprecipitated proteins were eluted from the beads with sample buffer for 1h at room temperature and then electrophoretically separated on 7% (w/v) polyacrylamide gels. Gels were pre-fixed in methanol/acetic acid (30:10, v/v), washed in water and, for fluorography, soaked in 1M sodium salicylate for 60 min. After drying at 80°C for 2h, gels were exposed to X-ray films and further analysed and quantified by densitometry.
Iodide efflux
CFTR-mediated iodide effluxes were measured at room temperature using the cAMP agonist forskolin (Fsk 10μM) and the CFTR potentiator genistein (Gen, 50μM) or VX-770 (10μM) and Gen (50μM).
Biochemical determination of the plasma membrane levels of CFTR
To determine plasma membrane levels of CFTR protein, we performed cell surface biotinylation in BHK cells cultured on permeable growth supports or tissue culture plates using cell membrane impermeable EZ-Link™ Sulfo-NHS-SS-Biotin, followed by cell lysis in buffer containing 25 mM HEPES, pH 8.0, 1% (v/v) Triton, 10% glycerol (v/v), and Complete Protease Inhibitor Mixture, as described previously 23,24. Biotinylated proteins were isolated by streptavidin-agarose beads, eluted into SDS-sample buffer, and separated by 7.5% (w/v) SDS-PAGE.
Multiple sequence alignment
Sequences for NBD domains of ABC transporters were obtained from Uniprot 25 (human and mouse CFTR) or the PDB 26 (1B0U, 1L2T, 1G29, 1G6H, 1JJ7). Alignments were performed with Jalview 27 using the T-Coffee 28 algorithm using the default parameters.
Data and Statistical analyses
The data and statistical analyses used in this study comply with the recommendations on experimental design and data analysis in pharmacology (Curtis et al., 2015). Quantitative results are shown as mean±SEM of n observations. To compare two sets of data, Student t-test was used and differences considered to be significant for p-values ≤ 0.05. In Western blotting (Figures 1–3), pulse chase (Figures 4, S3); cell surface biotinylation (Figure 4) and in iodide efflux studies (Table 1, Figure 5, S4 and S5), n represents the number of experiments performed with distinct cell cultures on different days, being thus biological replicates.

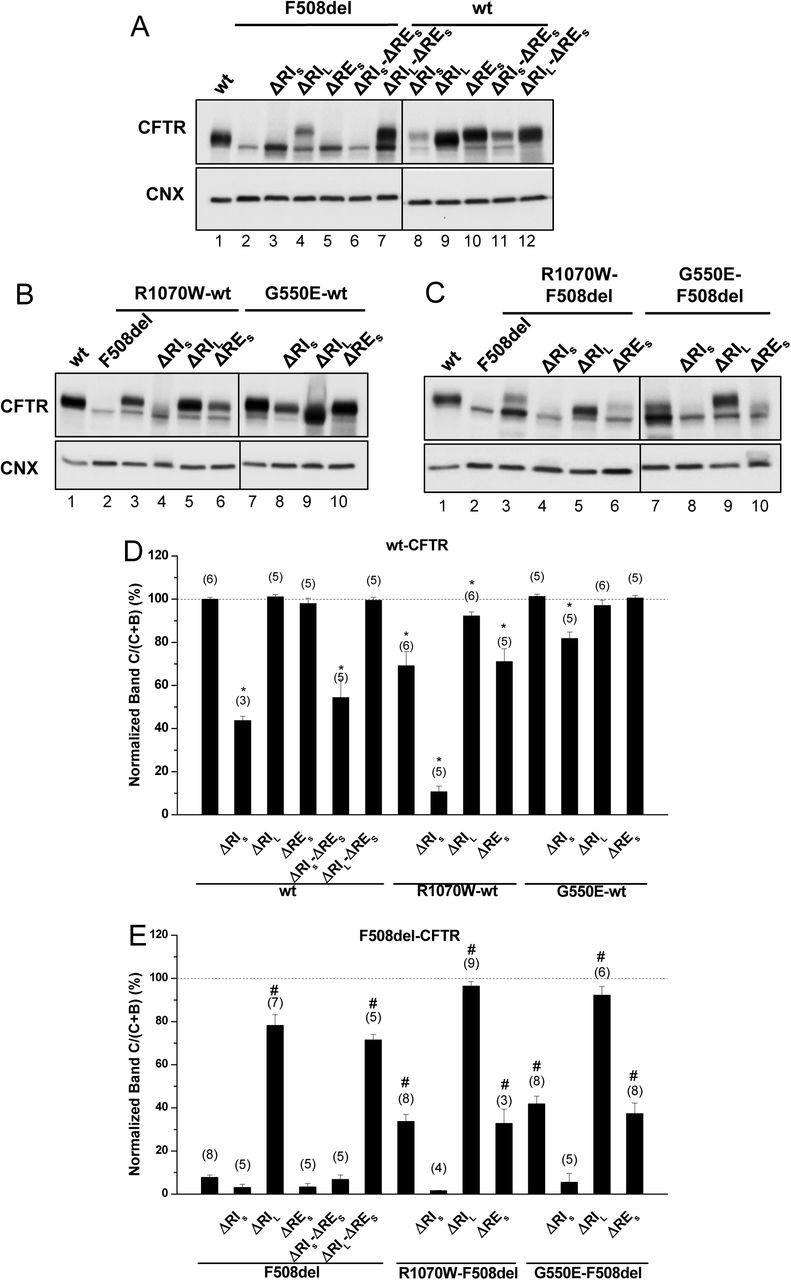
Effect of removal of short regulatory extension (ΔRES) on processing of wt- and F508del-CFTR. (A-C) Samples from BHK cell lines stably expressing different CFTR variants of ΔRES and ΔRI, the latter either in its short “S” or long “L” versions (see Methods) and alone (A) or jointly with (B, C) genetic revertants, as indicated, were analysed for CFTR protein expression by WB with Ab anti-CFTR and also anti-calnexin (CNX) as loading control. All samples were incubated with 3μM DMSO for 48h, as controls for experiments with corrector VX-809 (see Fig.3). Summary of data of CFTR variants on the wt- (D) or F508del- (E) CFTR backgrounds, expressed as normalized ratios (band C/(band C+band B)) and as a percentage to the respective ratio for wt-CFTR and as mean ± standard error of the mean (SEM). (n) indicates nr. of independent experiments. “*” and “#” indicate significantly different (p<0.05) from wt-CFTR and F508del-CFTR, respectively.

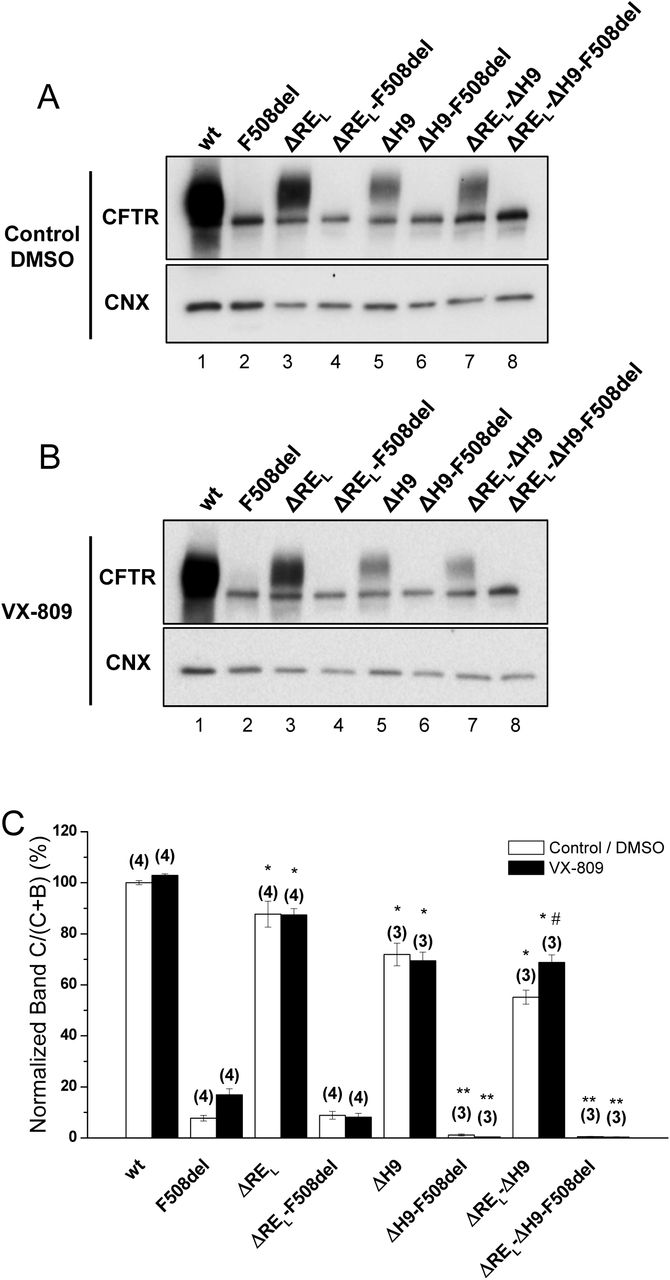
Effect of removal of helix H9 in combination with ΔREL and VX-809 on processing of wt- and F508del-CFTR. (A, B) WB analysis of samples from BHK cell lines stably expressing wt- or F508del-CFTR variants with ΔREL, ΔH9 or ΔREL-ΔH9. Cells were incubated with 3μM VX-809 or DMSO (control) for 48h at  . CFTR protein expression was analysed by WB with the anti-CFTR and anti-CNX Abs. (C) Summary of data expressed as normalized ratios (Band C / (Band C+B)) and as a percentage to the respective ratios on the wt- or F508del-CFTR backgrounds and shown as mean±SEM. (n) indicates
. CFTR protein expression was analysed by WB with the anti-CFTR and anti-CNX Abs. (C) Summary of data expressed as normalized ratios (Band C / (Band C+B)) and as a percentage to the respective ratios on the wt- or F508del-CFTR backgrounds and shown as mean±SEM. (n) indicates  of independent experiments. “#” indicates significantly different from the respective variant treated with DMSO. “*” and “**” indicate significantly different from wt- or F508del-CFTR, respectively.
of independent experiments. “#” indicates significantly different from the respective variant treated with DMSO. “*” and “**” indicate significantly different from wt- or F508del-CFTR, respectively.

Effect of VX-809 on processing of wt- and F508del-CFTR variants without regulatory extension. (A-C) WB of samples from BHK cell lines stably expressing different CFTR variants of ΔRE and ΔRI, as indicated, alone (A) or jointly with (B, C) genetic revertants (see Methods). CFTR protein expression was analysed by WB with anti-CFTR and anti-CNX Abs as in Fig.1. All samples were incubated with 3μM VX-809 for 48h. (D, E) Summary of data of variants on the wt- (D) or F508del- (E) CFTR backgrounds, expressed as normalized ratios (band C/(band C+band B)) and as a percentage to the respective ratio for wt-CFTR and as mean±SEM. (n) indicates  of independent experiments. “*” indicates significantly different (p<0.05) from respective variant without VX-809 (as shown in Fig.1 and indicated here by dashed bars).
of independent experiments. “*” indicates significantly different (p<0.05) from respective variant without VX-809 (as shown in Fig.1 and indicated here by dashed bars).

Plasma membrane levels, efficiency of processing and turnover of immature CFTR without Regulatory Extension (REs). (A) BHK cells expressing ΔRIL-F508del and ΔRIL-ΔRES-F508del-CFTR and treated with 3μM VX-809 for 48h (or DMSO control) were subjected to cell surface biotinylation. wt-CFTR samples not treated with biotin were the negative control (NC). After streptavidin pull-down, CFTR was detected by WB. CFTR and CNX are detected in the whole cell lysate (WCL) as controls. (B) Quantification of data in (a) for PM CFTR normalized to total protein and shown as fold change relatively to wt-CFTR cells treated with DMSO. and “#” indicate significantly different from wt-CFTR treated with DMSO and from respective variant without VX-809, respectively (p<0.05). (C) BHK cells expressing wt-, F508del-CFTR alone or jointly with ΔRIL, ΔRES and ΔRIL-ΔRES were subjected to pulse-chase (see Methods) for the indicated times (0, 0.5, 1, 2, 3h) before lysis and IP with the anti-CFTR 596 Ab. After electrophoresis and fluorography, images were analysed by densitometry. (D) Turnover of immature (band B) CFTR for different CFTR variants is shown as the percentage of immature protein at a given time point of chase (Pt) relative to the amount at t=0 (P0). (E) Efficiency of processing of band B into band C is shown as the percentage of band C at a given time of chase relative to the amount of band B at t=0. “*” and “#” indicate statistical significantly different (p<0.05) from wt-CFTR and F508del-CFTR, respectively. Data represent mean ± SEM (n=5).

Functional characterization of the ΔRES and ΔRIL variants of F508del-CFTR with or without VX-809 or VX-770 treatments. (A, B) Summary of data from the iodide efflux peak magnitude generated by BHK cells stably expressing different CFTR variants and treated with 3μM VX-809 (black bars on panel A) or with DMSO as control (white bars on panel a) for 48h and stimulated with 10μM Forskolin (Fsk) and 50μM Genistein (Gen). In (B) cells were not pre-incubated and were stimulated either with 10μM Fsk and 50μM Gen (white bars on panel B) or with 10μM Fsk and 10μM VX-770 (black bars on panel b), as indicated. Data are shown as a percentage of wt-CFTR activity under Fsk/Gen stimulation and as mean±SEM. (n) indicates no. of independent experiments. “#” indicates significantly different (p<0.05) from the same CFTR variant incubated with DMSO (A) or significantly different from the same CFTR variant under Fsk/Gen stimulation (white bars) (B). (C) Summary of iodide efflux data for different F508del-CFTR variants without RES, RIS, RIL alone or jointly with revertants R1070W and G550E.
Reagents
All reagents used here were of the highest purity grade available. Forskolin and genistein were from Sigma-Aldrich (St. Louis, MI, USA); VX-809 and VX-770 were acquired from Selleck Chemicals (Houston, TX, USA).
CFTR was detected with the mouse anti-CFTR monoclonal 596Ab, which recognizes a region of NBD2 (1204 - 1211) from CFFT-Cystic Fibrosis Foundation Therapeutics CFF Therapeutics (Cui et al., 2007) and calnexin with rabbit polyclonal anti-calnexin Ab SPA-860, from Stressgen Biotechnologies Corporation (Victoria, BC, Canada).
Other specific reagents included: X-ray films (Kodak, supplied by Sigma-Aldrich); Complete Protease Inhibitor Mixture from Roche Applied Science (Penzberg, Germany); EZ-Link™ Sulfo-NHS-SS-Biotin from Pierce Chemical Company (Rockford, IL, USA).
RESULTS
Removal of short regulatory extension (RES) alone or with regulatory insertion (RI) has no impact on F508del-CFTR processing
The impact of deleting RES – short RE (Δ654Ser-Gly673, Figure S1) was first assessed, either alone or together with RI in its short and long variants (ΔRIS, ΔRIL), on the in vivo processing of wt- and F508del-CFTR by Western blot (WB) of BHK cells stably expressing such variants (Fig.1A,D,E; Table1). Removal of RES had no impact on processing of either F508del-CFTR or wt-CFTR (Fig.1A, lanes 5,10; Table1), despite that the immature form of ΔRES-F508del-CFTR appeared consistently at increased levels vs. those of F508del-CFTR (Fig.1A, lanes 5,2).
Removal of RES together with RIL - long RI (Δ404Gly-Leu435, Figure S1) led to a maturation increase of F508del-CFTR from 3±2% to 71±3% (vs wt-CFTR levels), similarly to what had been previously reported by Alexandrov et al for ΔRIL alone 13. In contrast, removal of RIS - short RI (Δ412Ala-Leu428, Figure S1) had no impact on ΔRES-F508del-CFTR processing (Fig.1A, lanes 6,7; Fig.1E; Table1). So in summary, removal of RES jointly with RIS or RIL had no effect on F508del-CFTR processing.
Interestingly, when RES and RIS, were jointly removed from wt-CFTR, its processing was significantly reduced to 54±9%, while ΔRES jointly with ΔRIL caused no impact (Fig.1A, lanes11,12, respectively; Fig.1D; Table1). Again, these data were equivalent to removal of ΔRIS or ΔRIL alone on wt-CFTR processing (Fig.1A, lanes8,9; Fig.1E; Table1).
The differential effect caused by removal of RIS vs RIL on F508del- and wt-CFTR emphasize the importance of those 8 N-term (404Gly-Ala412) and 7 C-term (Leu428-Leu435) amino acid residues that differ between those 2 RI regions for the folding and processing of CFTR.
Simultaneous removal of long regulatory extension (REL) and helix H9 significantly reduces wt-CFTR processing but increases levels of F508del-CFTR immature form
Next, we assessed the impact of removing the long version of RE - REL (Δ647Cys-Ser678, Fig.2), which significantly decreased wt-CFTR processing to 88±5% (Fig.2A, lane 3; Fig.2C) but had no impact on F508del-CFTR (Fig.2A, lane4; Fig.2C). As helix H9 (Δ637Gln-Gly646, Figure S1) can be considered to be part of this longer RE region 22 since it interacts with RE (Figure S2), we also tested the effect of removing helix H9 jointly with ΔREL. Deletion of both H9 (ΔH9) and REL further decreased the residual processing of ΔREL-F508del-CFTR from 9±2 to 0±0% and in fact the same happened for ΔH9 alone (Fig.2a, lanes 8,6, respectively). Curiously, however, levels of immature F508del-CFTR were significantly increased when both REL and H9 were removed (Fig.2A, lane8), similarly to ΔRES-F508del-CFTR (Fig.1A) and in contrast to ΔREL-F508del-CFTR.
For wt-CFTR, ΔREL-ΔH9 also led to a decrease in processing (55±3%), which was more pronounced than for ΔREL alone, but interestingly removal of H9 helix alone only reduced processing to 72±4% (Fig.2A, lanes 7,5, respectively).
ΔRIS, but not ΔRES, abolishes the plasma membrane rescue of F508del-CFTR by revertants
In order to test whether the stabilized immature form of ΔRES-F508del-CFTR could be rescued to mature form, we next tested the effects of removing the RES from F508del-CFTR with R1070W and G550E revertants. Indeed, both the R1070W and G550E revertants rescued ΔRES-F508del-CFTR to 33±7% and 37±5%, respectively (Fig.1C, lanes 6,10; Table1).
However, removal of RIS from F508del-CFTR with R1070W and G550E revertants, virtually abolished the rescue of F508del-CFTR by both revertants (Fig.1C, lanes 4,8; Table1). Moreover, processing of ΔRIL-F508del-CFTR was further increased by either R1070W or G550E up to 96±2% and 92±4%, respectively (Fig.1C, lanes 5,9; Table1). Alone, R1070W and G550E rescued F508del-CFTR from 8±1% to 34±3% and 42±4% respectively (Fig.1C, lanes 3,7; Table1), as we previously reported 5.
Interestingly, the impaired processing of ΔRIS-wt-CFTR (44±2%) was significantly rescued by G550E (to 82±3%), but curiously it was further reduced by R1070W (to 11%) i.e., close to levels of F508del-CFTR processing (Fig.1B, lanes8,4; Table1). Of note that R1070W alone also reduced wt-CFTR processing to 69±7%, but this reduction could be compensated by the removal of RIL (92±2%) (Fig.1B, lanes 3,5; Table1), while G550E alone caused no effect on wt-CFTR processing (Fig.1B, lane7).
ΔRIL synergises with VX-809, but not with revertants to rescue ΔRES-F508del-CFTR processing
To further test how the stabilized immature form of ΔRES-F508del-CFTR could be pharmacologically rescued, we then assessed the impact of corrector VX-809 (which per se promotes maturation of the F508del-CFTR) on the processing of this and of the other variants, to obtain structural insight on the effects of this novel drug. Although ΔRES-F508del-CFTR could not be rescued by VX-809, data show that this small molecule was able to rescue ΔRIS-ΔRES-F508del-CFTR (from 7±2% to 20±4%) and further increased processing of ΔRIL-ΔRES-F508del-CFTR to wt-CFTR levels (from 71±3% to 96±2%) (Fig.3A, lanes 6,7; Fig.3E; Table1). These data suggest a strong synergistic effect between VX-809 and ΔRIL (and with ΔRIS, albeit to a lesser extent) to rescue ΔRES-F508del-CFTR processing. This is particularly interesting because VX-809 did not rescue processing of the ΔRES-, nor ΔRIS-F508del-CFTR variants (Fig.3A, lanes 5,3; Table1) while it recovered the processing of ΔRIL-F508del-CFTR to wt-CFTR levels (Fig.3A, lane4; Table1). Notably, VX-809 was able to almost completely revert processing impairment of ΔRIS-wt-CFTR and ΔRIS-ΔRES-CFTR (from 44±2% and 54±9%) to 90±3% and 93±3%, respectively (Fig.3A, lanes8,11; Fig.3D; Table1).
Strikingly, VX-809 had no effect on processing of ΔRES-F508del-CFTR with R1070W or G550E (Fig.3C, lanes 6,110; Fig.3E; Table 1), while it further rescued processing of F508del-CFTR variants with R1070W or G550E alone to 46±4% and 73±6%, respectively (Fig.3C, lanes 3,7; Fig.3E; Table 1), as reported 5. Similarly, VX-809 caused no significant further rescue on the processing of ΔRIL-F508del-CFTR with those revertants, but these variants already had processing levels close to those of wt-CFTR (Fig.3C, lanes 5, 9; Fig.3E; Table 1). Interestingly, the impaired processing of ΔRES-R1070W-wt-CFTR (71±6%) was also reverted by VX-809 to 90±3% (Fig.3B, lane 6; Table 1).
Corrector VX-809 failed to rescue any of the ΔREL-, ΔΗ9- and ΔREL-ΔH9-F508del-CFTR variants (Fig.2B, C), while significantly increasing the processing of the ΔREL-ΔH9-wt-CFTR from 55±3% to 69±3% (Fig.2B, lane 7; Fig.2C).
ΔRIL-ΔRES-F508del-CFTR levels at the plasma membrane are equivalent to those of wt-CFTR
To determine the fraction of the above CFTR variants that localize to the PM, we used quantitative cell surface biotinylation. These data showed that PM levels of ΔRIL-ΔRES-F508del-CFTR were equivalent to those of wt-CFTR, while those of ΔRIL-F508del-CFTR were significantly lower (Fig.4A, lanes 5, 4; Fig.4B). Data also confirmed that ΔRES did not induce appearance of F508del-CFTR at the cell surface (data not shown). Corrector VX-809 further increased the PM expression of ΔRIL-ΔRES-F508del-CFTR to levels that are significantly higher than those of wt-CFTR (Fig.4A, lanes 2, 9, Fig.4B). This compound also significantly increased PM levels of ΔRIL-F508del-CFTR to similar levels of wt-CFTR (Fig.4A, lanes 2, 8; Fig.4B).
Given the very significant stabilization of immature ΔRES-F508del-CFTR (Fig.1A), next, we determined how removal of ΔRES affected the processing efficiency and the turnover of the F508del- and ΔRIL-F508del-CFTR variants. To this end, we performed pulse-chase experiments (Fig.4C, D) and indeed our results revealed that ΔRES very significantly stabilized immature F508del-CFTR not just relatively to F508del-CFTR but to levels even significantly higher than those of wt-CFTR (Fig.4C, D). Interestingly, this stabilizing effect was no longer significant for ΔRIL-ΔRES-F508del-CFTR nor for ΔRIL-F508del-CFTR, the latter being equivalent to wt-CFTR (Fig.4C, D). Removal of RIS from F508del-CFTR did not stabilize its turnover (data not shown). As to removal of either RES or RIS from wt-CFTR, it did not affect the processing efficiency or the turnover vs wt-CFTR (Figure S3).
VX-809 jointly with RE and RI removal completely restored F508del-CFTR function as chloride channel
To investigate the channel function of the ΔRES- and ΔRIL-F508del-CFTR variants, we used the iodide efflux technique. Removal of RES alone had no impact on F508del-CFTR function (Fig.5A; black dash line in Figure S4B) as expected from the lack of processed form for this variant (Fig.1A). However, when RES and RIL were removed jointly from F508del-CFTR, functional levels reached 78±7% of wt-CFTR (Fig.5A, C; black dotted line in Figure S4B). As to removal of RIL alone from F508del-CFTR, it restored function to 92±7% of wt-CFTR (Fig.5A, C; black line in Figure S4B), as described 13. Also, the delay in peak response observed for F508del-CFTR (min=4) vs wt-CFTR (min=2), was partially corrected (to min=3) both by removal of RIL alone from F508del-CFTR or together with RES (Fig.5C; Figure S4B, black line and dotted line).
Introduction of G550E and R1070W revertants into the ΔRIL-F508del-CFTR variant slightly but not significantly decreased its function from 92±7% to 71±8% and 76±8%, respectively (Fig.5A, C; black lines in Figure S4C, D), in parallel to the respective processing levels (Fig.1). In fact, these variants showed higher activity levels than the revertants containing RIL (29±3% and 56±6%, respectively). Interestingly however, both G550E- and R1070W-ΔRIL-F508del-CFTR fully corrected (to min=2) the delay in peak response of F508del-CFTR, which was still partially present in ΔRIL- and ΔRIL-RES-F508del-CFTR (min=3).
The effect of VX-809 (3μM, 48h at  ) was also examined at functional level on the ΔRES-, ΔRIL-ΔRES- and ΔRIL-F508del-CFTR variants. Strikingly VX-809 caused an increase in ΔRES-ΔRIL-F508del-CFTR function (84±8% of wt-CFTR) which was parallel to the observed increase in PM expression (Fig.4; Fig.5A, C; black dotted line in Figure S4F). Also, ΔRES-ΔRIL-F508del-CFTR fully corrected (to min=2) the delay in peak response of F508del-CFTR which was still partially present in ΔRIL- and ΔRIL-ΔRES-F508del-CFTR (min=3).
) was also examined at functional level on the ΔRES-, ΔRIL-ΔRES- and ΔRIL-F508del-CFTR variants. Strikingly VX-809 caused an increase in ΔRES-ΔRIL-F508del-CFTR function (84±8% of wt-CFTR) which was parallel to the observed increase in PM expression (Fig.4; Fig.5A, C; black dotted line in Figure S4F). Also, ΔRES-ΔRIL-F508del-CFTR fully corrected (to min=2) the delay in peak response of F508del-CFTR which was still partially present in ΔRIL- and ΔRIL-ΔRES-F508del-CFTR (min=3).
In contrast to the observed increase in processing, VX-809 caused no significant increase in the function of ΔRIL-F508del-CFTR and actually a slight, but not significant decrease was observed and there was still the same delay in peak response (Fig.5A, C; black line in Figure S4F).
A similar slight decrease in function was observed for VX-809 on the ΔRIL-R1070W-F508del-CFTR variant (Fig.5A, C; black line in Figure S4H), but the most striking result was observed for the significant decrease caused by VX-809 on function of ΔRIL-G550E-F508del from 76±8 to 41±3% (Fig.5A, C; black line in Figure S4G). Nevertheless both revertant variants of ΔRIL-F508del-CFTR showed no change in peak response which was still the same as wt-CFTR (min=2).
VX-770-stimulated currents of CFTR variants are dramatically decreased by ΔRIL but not by ΔRIL-ΔRES
Given the interesting and diverse results observed for the ΔRES and ΔRIL under VX-809, next we tested the effects of potentiator VX-770 (Ivacaftor), an approved drug for CF patients with gating mutations (Fig.5B) on these variants and in combination with VX-809, for F508del/F508del patents. Most strikingly, our data show that VX-770/Fsk significantly decreased the function of ΔRIL-F508del-CFTR vs potentiation by Gen/Fsk (Fig.5B, C) from 92±7% to 58±7% of wt-CFTR (Fig.5B, C; black line in Figure S4J). As ΔRES-F508del-CFTR was not processed, we did not test the effect of VX-770 on this variant, but when the ΔRES-ΔRIL-F508del-CFTR variant was assessed for its function after acute application of VX-770 with Fsk, an increase in function (to 87±3% of wt-CFTR) was observed vs its function under Gen/Fsk (78±7% of wt-CFTR) (Fig.5B, C; black dotted line in Figure S4J).
The most dramatic result, however, was observed for the ΔRIL-G550E-F508del-CFTR variant which under VX-770/Fsk only had 4±1% function of wt-CFTR, while under Gen/Fsk it had 76±8% (Fig.5B, C; black line in Figure S4K). For ΔRIL-R1070W-F508del-CFTR this decrease was not observed (Fig.5B, C; black line in Figure S4L), and in fact this variant had a slightly higher function under VX-770 stimulation (72±4%) than ΔRIL-F508del-CFTR (58±7%) (Fig.5B, C, black line in Figure S4H).
The delay in peak response of F508del- vs. wt-CFTR was not corrected for ΔRIL-F508del-CFTR (min=3), but it was corrected for ΔRIL-ΔRES-F508del-CFTR (min=2) (black and dotted black lines in Figure S4J, respectively). Curiously, on the other hand ΔRIL-R1070W-F508del-CFTR showed the quickest peak response (at min=1), in contrast to either R1070W-F508del-CFTR or ΔRIL-F508del-CFTR, both at min=3.
Similarly, VX-770 also significantly decreased the function of ΔRIL-wt-CFTR from 127±15% under Gen to 57±6% (Figure S5).
Discussion
The main goal of this study was to understand how the removal of the regulatory extension (ΔRE) alone or with the regulatory insertion (ΔRI)-two highly conformationally dynamic regions - impacted on the rescue of F508del-CFTR processing and function by two compounds - VX-809 and VX-770 - which in combination were recently approved o clinically treat F508del-homozygous patients. Indeed, CFTR is the sole ABC that functions as a channel and thus, these highly dynamic RE and RI regions which are absent in other ABC transporters, may be of high relevance to understand how CFTR differs from other ABCs, namely in its function as a channel.
These regions RE and RI were originally suggested to be positioned to impede formation of the NBD1-NBD2 dimer required for channel gating 16,17. Indeed, both RI and RE were shown to be mobile elements in solution that bind transiently to the core of NBD in the ß-sheet and α/ß subdomains of NBD13. Similarly to what demonstrated for RI13, is plausible to posit that the dynamic flexibility of the RE may also result in exposure of hydrophobic surfaces thus contributing to the dynamic instability of NBD1 and thus contribute to the low folding efficiency of F508del-CFTR. The RE is a ~30-residue segment at NBD1 C-terminus, so-called because it goes beyond canonical ABC NBDs 16. Although this region was absent from the solved CFTR-NBD1 crystal structure 17, it was described as a helix packing against NBD1 at the NBD1:NBD2 interface and NMR data showed it has significant conformational flexibility 3,20. Notwithstanding, there is some controversy regarding the RE boundaries. The RE (previously called H9c) was defined as 655Ala-Ser670 16,17, 639Asp-Ser670,21 or 654Ser-Gly673,3. According to the crystal structure (PDB code 2PZE), NBD1 extends only to 646Gly, where RD begins, so RE could be proposed to start at 647Cys 3,22, being thus considered as part of RD, a domain absent in other ABC transporters 1. As for the RI (previously called S1-S2 loop) its limits are also variable, being firstly described as a ~35-residue segment (405Phe-436Leu) consisting of a-helices H1b and H1c 16 (Fig.S1). Later, however, these limits were proposed to be 404Gly-Leu435 13,21 or 405Phe-Leu436 22.
Impact of RE and RI on CFTR processing and function
Our data shown here on CFTR variants depleted of different versions of the RE dynamic region demonstrate that unlike RI deletion, removal of short RE - ΔRES (Δ654Ser-Gly673) did not per se rescue F508del-CFTR processing. Nevertheless, ΔRES dramatically stabilized the immature form of F508del-CFTR (see Fig.6). In fact, our pulse-chase experiments show that the immature form of ΔRES-F508del-CFTR exhibits a turnover rate which is ~2.7 fold lower than that of wt-CFTR. Of note, when Aleksandrov et al removed the dynamic region RI from F508del-CFTR they found a dramatic increase in the channel thermostability, even augmented for higher temperatures 13. Interestingly however, removal of the RES from ΔRIL-F508del-CFTR (ΔRIL: Δ404Gly-Leu435) while not affect processing, it further reduced its function (from 92% to 78%).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Summary of the most relevant results observed in the present study. CFTR structure used is the one by Liu F et al 33 and putative bind sites of VX-809 and VX-770 shown are the NBD1:MSD2 (ICL4) interface and to the MSDs (although it is not defined the exact binding site), as described by Farinha et al 41 and by Jih & Hwang 42, respectively.
The latter findings on function of ΔRES-wt-CFTR and ΔRES-ΔRIL-F508del-CFTR were somewhat surprising since the RE was previously described to impede putative NBD1:NBD2 dimerization that is required for channel gating 21. Accordingly, an increase in CFTR activity would be expected upon ΔRES removal, which is actually the opposite of what we observe for wt-CFTR. Besides the fact that those authors studied a different RE version (639Asp-Ser670), this discrepancy could derive from structural constraints and lack of flexibility of the 'single polypeptide' protein that we used, vs their 'split' CFTR channels (2 'halves' of 633 and 668 aa) for which channel function was indistinguishable from wt-CFTR 21. Moreover, since RD phosphorylation is required for channel gating 29,30, the reduced function of ΔRES-wt-CFTR may also result from absence of 660Ser and 670Ser 19,20. Although the RD contains more than ten PKA phospho-sites and no individual one is essential, phosphorylation of increasing numbers of sites enables progressively greater channel activity 31.
Removal of a longer RE version (ΔREL: Δ647Cys-Ser678) was without effect on F508del-CFTR processing and significantly reduced that of wt-CFTR to 88%. Such differential impact on wt- and F508del-CFTR is consistent with the conformational heterogeneity between these two proteins lacking both RI and RE 32.
Removal of RI short version, RIs (A412Ala-Leu428) significantly reduced wt-CFTR processing to less than half of its normal levels, while not rescuing F508del-CFTR processing, thus being essential for CFTR proper folding. This is in contrast to removal of long RI (ΔRIL) which led to 78% processing F508del-CFTR, as reported 13 but without impact on wt-CFTR. These data indicate that those 8/ 7 amino acid residues at the N-term/ C-term of RIS (and absent in RIL) impair the folding efficiency and processing of both wt- and F508del-CFTR.
The recently published cryo-EM structure of human CFTR 33 indicates that these regions are structurally disordered. Consistently, the RI loop in NBD1 is described in the of zebrafish CFTR cryo-structure to contribute to the amorphous density that is observed between NBD1 and the elbow helix of TMD2 34. From a structural point of view, models of the full-length CFTR protein suggest that the two regions (RI and RE) behave differently. The distances between the extremities of the deleted parts are indeed different (ΔRIS = 18Å, ΔRIL= 10Å versus ΔRES = 27Å, ΔREL = 30Å). The longer distances observed for ΔRE imply a substantial reorganization of the C-terminal parts of NBD1 and NBD2, which precludes a clear understanding of what might happen upon these deletions. In contrast, the lower distances observed for ΔRI (near a possible minimum of 6Å) allows a better simulation of the possible structural behaviour of these constructs. Indeed, preliminary molecular dynamics simulations of the ΔRIL-F508del-CFTR suggested that in this case, part of the RD is reorganized and partially takes space left by the ΔRI deletion, thereby substituting it for tight contacts with NBD2. Meanwhile, following a probable allosteric effect 13,22,35, contact of NBD1-F508 with ICL4 residue L1077 (a likely essential contact for channel opening), is nearly completely restored by I507. Such a feature is not observed in the F508del-CFTR model (B. Hoffmann, I. Callebaut and J.-P. Mornon, personal communication).
Effect of VX-809 on F508del-CFTR variants lacking RE and RI
As for rescue of CFTR variants lacking RE and RI by CFTR modulators, VX-809 restored the processing of both ΔRES-ΔRIL- and ΔRIL-F508del-CFTR variants equally well and to wt-CFTR levels (from 71-78% to 92-96%. These data suggest a strong synergistic effect between VX-809 and ΔRIL to rescue ΔRES-F508del-CFTR processing and thus some possible interference of the regulatory insertion with VX-809 binding to F508del-CFTR (see Fig.6). Indeed, although previous studies 5,36,37 suggested putative binding of VX-809 to NBD1:MSD2 (ICL4) interface (see Fig.6), they also suggested scope for further F508del-CFTR correction at distinct conformational sites, so data shown here suggest that VX-809 may also bind to the regulatory insertion.
Interestingly, the processing defect of ΔRIS-wt-CFTR (but not of ΔRIS-F508del-CFTR) was rescuable by VX-809 to 90%, indicating that the amino acid stretches of the RIL that remain present in RIS do not affect the rescue of ΔRIS-wt-CFTR but preclude rescue of F508del-CFTR by VX-809.
We also tested here the impact of removing helix 9 (H9) which precedes the RE (635Gln-Gly646), just after H8 (630Phe-Leu634), both helices proposed to interact with the NBD1:NBD2 heterodimer interface by folding onto the NBD1 ß-subdomain 20,32 as well as to bind ICL4 near Phe508. When H9 is present, ΔREL-wt-CFTR processing appears to be favoured by VX-809, suggesting some synergy of this small molecule with H9 helix to correct the conformational defect(s) caused by ΔREL on wt-CFTR. These data are in contrast to ΔH9-F508del-CFTR which exhibits 0% processing with or without ΔREL and no rescue by VX-809.
Very strikingly, and in contrast to its effect on processing, was the effect of ΔRIL on the VX-770 stimulated currents which were decreased by almost half vs those stimulated by Gen/Fsk (92% to 58%). These data seem to indicate that the absence of the regulatory insertion could impair (and its presence favour) binding of VX-770 to CFTR (see Fig.6). Surprisingly, the removal of REs from ΔRIL-F508del-CFTR could correct this defect and restore the maximal function by VX-770 (see Fig.6). Indeed, under VX-770, ΔRES-ΔRIL-F508del-CFTR exhibited significantly higher activity (87%) than ΔRIL-F508del-CFTR (58%). In contrast, removal of RES from wt-CFTR significantly reduced its function vs that of wt-CFTR (down to 70%) but had no effect on processing.
Impact of F508del-revertants on CFTR variants lacking RE and RI
Another goal of the present study was to assess how presence of the F508del-CFTR revertants G550E and R1070W 5,10,11 influence variants without RE and RI. Remarkably, our data show that the presence of either of these revertants did not affect ΔRES-F508del-CFTR processing, but both of them further increased processing (but not function) of ΔRIL-F508del-CFTR to almost levels of wt-CFTR: 92-96% (G550E) and 71-76% (R1070W).
In contrast, removal of RIS from either of these F508del-CFTR revertants completely abolished their processing, emphasizing how important the different residues between RIS and RIL are for F508del-CFTR conformers partially rescued by the revertants.
Regarding wt-CFTR processing, G550E (at the NBD1:NBD2 dimer interface) also partially recovered the negative effect caused by RIS removal. In contrast, R1070W (at the NBD1:ICL4 interface), negatively affected processing of wt-CFTR (to 69%) and of ΔRIS-wt-CFTR (from 44% to 11%), while not affecting ΔRIL-wt-CFTR. R1070W rescues F508del-CFTR because Trp1070 fills the gap created by deletion of residue Phe508 38. It is not surprising that it perturbs CFTR folding due to clashing of 508Phe and 1070Trp residues. It is, nevertheless curious that R1070W affect more the processing ΔRIS-wt-CFTR than wt-CFTR, to levels of F508del-CFTR, suggesting that both changes affect the same region of the molecule. Rescue of both R1070W-wt-CFTR and R1070W-ΔRIS-wt-CFTR by VX-809 further supports this concept.
The most striking effect of ΔRIL however, was the almost complete abolition of VX-770-stimulated current of G550E-F508del-CFTR to levels even lower than those observed for ΔRIL-F508del-CFTR (see above). It was suggested that VX-770 binds directly to CFTR to the MSDs (although it is not defined the exact binding site), in phosphorylation-dependent but ATP-independent manner and away from the canonical catalytic site 39,40. Both RD and RI were suggested by Eckford and colleagues as putative binding regions for VX-770 39. Nevertheless, the pharmacological effect of VX-770 remains robust in the absence of the RD 40 and here we demonstrate that it does indeed require RI since VX-770 is unable to stimulate either ΔRIL-F508del-CFTR or ΔRIL-G550E-F508del-CFTR (see Fig.6).
Overall, our data show that while the presence of the regulatory insertion seems to be precluding full rescue of F508del-CFTR processing by VX-809, this region appears essential to rescue its function by VX-770, thus suggesting some contradictory role in rescue of F508del-CFTR by these two modulators. Nevertheless, this negative impact of RIL removal on VX-770-stimulated currents on F508del-CFTR can be compensated by deletion of the regulatory extension which also leads to the stabilization of this mutant. We thus propose that, despite both these regions being conformationally active, RI precludes F508del-CFTR processing while RE affects mostly its stability and channel opening.
Conflict of interest
The authors declare no conflict of interest related to this paper.
Author Contributions
IU, ACDP, SA, MJL, VF and CMF preformed the experiments; MDA designed the study and obtained funding; IU, ACDP, HB and MDA interpreted the data, wrote the manuscript.
Acknowledgements
Work supported by grants PTDC/SAU-GMG/122299/2010 (to MDA) and UID/MULTI/04046/2013 centre grant (to BioISI), both from FCT/MCTES/PIDDAC, Portugal. IU, ACDP and VF are/were recipients of SFRH/BD/69180/2010, SFRH/BD/17475/2004, SFRH/BD/87478/2012 PhD fellowships and HMB of SFRH/BPD/93017/2013 post-doctoral fellowship (FCT, Portugal), respectively. Authors acknowledge I Callebaut, B Hoffmann and J-P Mornon for the revision of the manuscript and for the helpful comments.
Footnotes
↵1 Current addresses: Department of Physiology, Anatomy and Genetics, University of Oxford, UK (IU and MJL); Novartis Pharmaceuticals, Montreal, Quebec, Canada (ACDP); Department of Physiology, University of Regensburg, Germany (SA).
↵1 www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm453565.htm
REFERENCES




