

Abstract
Basement membranes (BMs) are specialized layers of extracellular matrix (ECM) mainly composed of Laminin, type IV Collagen, Perlecan and Nidogen/entactin (NDG). While the essential and evolutionary conserved functions of Laminin, Collagen and Perlecan are well documented in Drosophila and other species, the proposed role of NDG as the major ECM linker molecule has been challenged by several in vivo studies revealing that NDG is dispensable for viability and BM formation. Here, we report the characterization of the single Ndg gene in Drosophila. Embryonic Ndg expression differed from that of other BM components and was primarily observed in mesodermal tissues and the chordotonal organs, whereas NDG protein localized to all BMs. While loss of Laminin strongly affected BM-localization of NDG, Ndg null mutants exhibited no overt changes in the distribution of BM core components. However, loss of NDG led to ultrastructural BM defects compromising barrier function and stability in vivo. Although Ndg mutants were viable, loss of NDG led to decreased fecundity in flies as well as impaired crawling behavior and reduced response to vibrational stimuli in larvae. Further morphological analysis revealed accompanying defects in the larval peripheral nervous system especially in the chordotonal organs and the neuromuscular junction (NMJ), where Ndg genetically interacted with the Leukocyte-antigen-related-like (Lar) receptor gene to regulate NMJ extension and synaptic differentiation. Taken together, our analysis suggests that NDG is not essential for BM assembly but mediates BM stability and ECM-dependent neural plasticity during Drosophila development.
Summary Statement In this study we characterize Drosophila Nidogen/Entactin (Ndg) mutants revealing that loss of Ndg impairs basement membrane (BM) stability and permeability as well as proper function of the nervous system.
Introduction
Proteome analysis of isolated BMs has identified over 100 associated proteins highlighting the complex nature of these specialized ECM sheets (Uechi et al., 2014). However, BMs mainly assemble from a small subset of ECM proteins referred to as the “BM toolkit” which is a highly conserved feature of most metazoan species (Hynes, 2012). BMs consist of two different mesh-like networks formed by self-assembly of either heterotrimeric Laminin molecules or type IV Collagens which are then linked by their binding partners Nidogen (NDG), the heparan sulfate proteoglycan (HSPG) Perlecan and additional Collagen XV/XVIII homologs (Jayadev and Sherwood, 2017; Yurchenco, 2011). The Drosophila genome harbors a minimal set of nine genes encoding components of the “BM toolkit” (Hynes, 2012). Four genes encode for two Laminin α-, one β- and one γ-subunit which form the only two Laminin heterotrimers in the fly, two adjacent loci encode for type IV Collagens (Col4a1 and viking), one for a ColXV/XVIII homolog named Multiplexin, a single Nidogen/entactin gene, and the terribly reduced optic lobes (trol) locus that encodes for the Perlecan protein core. With the exception of Ndg, all mayor BM components of Drosophila have been functionally characterized (Borchiellini et al., 1996; Datta and Kankel, 1992; Friedrich et al., 2000; Henchcliffe et al., 1993; Lindsley and Zimm, 1992; Martin et al., 1999; Meyer and Moussian, 2009; Urbano et al., 2009; Voigt et al., 2002; Wolfstetter and Holz, 2012; Yarnitzky and Volk, 1995; Yasothornsrikul et al., 1997).
In 1983, Timpl et al (Timpl et al., 1983) reported the identification of an 80 kDA protein from mouse Engelbreth-Holm-Swarm (EHS) sarcoma cell lines which was named Nidogen due to its ability to self-aggregate into “nest-like structures”. Although this protein was later identified as a proteolytic fragment of Entactin, a 150 kDa sulfated glycoprotein described by Carlin et al. (Carlin et al., 1981), the terms Nidogen or Nidogen/Entactin are now commonly used to refer to the non-cleaved protein (Martin and Timpl, 1987). The Nidogen protein consists of three globular domains (G1 to G3), a rod-like segment between G1 and G2 as well as an EGF-like-domain-containing segment that connects G2 and G3 (Durkin et al., 1988; Fox et al., 1991). Nidogen forms stable complexes with Laminin through binding of its globular G3 domain to the Laminin γ-subunit, whereas the G2 domain mediates binding to Collagen IV and Perlecan (Fox et al., 1991; Hopf et al., 2001; Mann et al., 1989; Reinhardt et al., 1993). Therefore it has been suggested that Nidogen supports the formation of ternary complexes within the BM and functions as an “ECM linker molecule” connecting the Laminin and Collagen IV networks (Aumailley et al., 1993; Ho et al., 2008; Mayer et al., 1993; Mayer et al., 1995).
Functional analysis of the two Nidogen family members in mammals reveals that the single knockouts of Nidogen 1 (NID1) and Nidogen 2 (NID2) in mice neither result in lethality nor cause any gross defects in BM formation and morphology. However, NID1−/−. animals exhibit neurological phenotypes like spontaneous seizures and hind limb ataxia (Dong et al., 2002; Murshed et al., 2000; Schymeinsky et al., 2002). Moreover, elevated NID2 levels in BMs of NID1−/−. mice suggest a functional redundancy between the two members of the mammalian Nidogen family (Miosge et al., 2000). Analysis of double mutant NID1−/−. NID2−/−. mice does not reveal an essential function during embryogenesis and embryonic BM formation but complete absence of Nidogen causes perinatal lethality due to impaired lung and heart development accompanied by defects in the organ-associated BMs (Bader et al., 2005). Moreover, a varying degree of syndactyly and occasional twisting of forelimbs is observed in NID1−/−. NID2−/−. animals (Bose et al., 2006). In accordance with these results a non-essential function of Nidogen for invertebrate development is revealed by the analysis of C. elegans nidogen-1 (nid-1, the single Nidogen homolog in the nematode) mutants that are viable and fertile and do not display abnormalities during BM assembly. In addition to a slight reduction in fecundity, nid-1. mutant animals exhibit aberrant guidance and positioning of longitudinal nerves, movement defects in a body bending assay, and altered neuromuscular junction (NMJ) organization which suggests a role in nervous system patterning (Ackley et al., 2003; Hobert and Bulow, 2003; Kang and Kramer, 2000; Kim and Wadsworth, 2000; Kramer, 2005). A recent study by Zu et al. (Zhu et al., 2017) reports body length reduction in Danio rerio upon nid1a gene depletion but also suggests that Ndg-associated phenotypes are likely to be obscured by functional redundancy and genetic compensation (Rossi et al., 2015) between the four predicted Nidogen family members in zebrafish.
In this work, we report the characterization of the single Nidogen/entactin (Ndg) gene in Drosophila. We found that Ndg was highly expressed in the Drosophila embryo and NDG protein was abundant in all basement membranes (BMs) suggesting an essential function during development. In contrast, analysis of Ndg mutants revealed that loss of NDG did not affect viability and fertility in general, arguing for a negligible role during development. This was further confirmed by examination of BM assembly in Ndg mutant embryos revealing no overt changes in overall BM protein distribution. Interestingly, ultrastructural SEM analysis and permeability assays revealed a porous BM surface and severe defects in BM barrier function accelerating the disruption of imaginal disc BMs under osmotic stress conditions. Moreover, Ndg. mutant larvae displayed a range of strong behavioral phenotypes such as reduced responses to external stimuli, impaired motility and climbing performance as well as altered crawling behavior and gravitaxis. Our morphological analysis of the larval nervous system indicated that these phenotypes correlated with improper positioning of neurons, defasciculation of nerves as well as severe defects in chordotonal organ and neuromuscular junction (NMJ) organization. We also found that Ndg genetically interacted with the Leukocyte-antigen-related-like (Lar) receptor gene to promote NMJ differentiation and plasticity. Taken together, our analysis of Drosophila Ndg did not support an essential role of NDG for the assembly of BMs but suggested that NDG essentially contributes to the robustness of the BM barrier and ensures ECM plasticity in response to external cues.
Results and Figures
Ndg transcript expression and protein localization
Developmental Northern blot analysis revealed high levels of Ndg expression in Drosophila embryos after gastrulation (Sup. Fig. 1). We therefore applied fluorescence in situ hybridization on white1118 (w1118) embryos to analyze Ndg mRNA distribution in comparison to NDG protein localization revealed by antibody staining (Fig. 1). In line with our Northern blot analysis, Ndg expression was apparent at stage 11/12 and could be detected in single cells of the head, especially in the gnathal segments (Fig. 1A, asterisk) as well as in segmentally located patches of cells in the dorsal mesoderm (arrowhead, Fig. 1A). Moreover, the midline-associated, mesodermal dorsal median cells (DMC, Fig. 1B, C) as well as four surrounding somatic myoblasts (sMB, Fig. 1B) exhibited strong Ndg expression whereas we detected only very weak signals in the amnioserosa (AS, Fig. 1C). At this time NDG protein, embedding DMCs and sMBs, formed a thin sheet between ecto-and mesoderm and outlined the region of the prospective anal plate. After germ band retraction (Fig. 1D) mRNA expression had increased and was now visible in the forming dorsal and ventral muscles, the amnioserosa (AS) and the segmentally located chordotonal organs (Cho). This was in agreement with the expression profile obtained from developmental Northern blot analysis which revealed a strong Ndg mRNA increase in older embryos (Sup. Fig. 1). Internal views indicated a decrease of expression in the DMCs (arrow) whereas Ndg was now expressed in the esophageal visceral muscle primordium (eVM, arrowhead) and the joint region between hind-and midgut (Fig. 1E). At this time, accumulation of NDG protein in the forming BMs around the developing brain and ventral nerve cord (VNC), the differentiating tracheal system, the future digestive tract as well as the forming somatic muscles was observed (Figure 1D, E). Embryos at stage 16 displayed strong mRNA expression in somatic and visceral muscles and in the cap cells (CC in Figure 1H) of the chordotonal organs, whereas the protein localized to all embryonic BMs (Figure 1F-I). Interestingly, whereas Ndg mRNA expression was absent from neuronal tissues, the BMs surrounding the chordotonal neurons (CN), the VNC and the brain were highly NDG-positive (Figure 1H, I). In summary, Ndg mRNA expression during embryogenesis followed a dynamic and distinct pattern involving mostly mesodermal cells and tissues but also chordotonal cap cells and the amnioserosa whereas the protein was highly abundant in all embryonic BMs. Interestingly, no transcript expression was observed in hemocytes or fat body cells although these tissues strongly express other BM components like Laminin or type IV Collagen (Le Parco et al., 1986; Pastor-Pareja and Xu, 2011; Rodriguez et al., 1996; Wolfstetter and Holz, 2012).
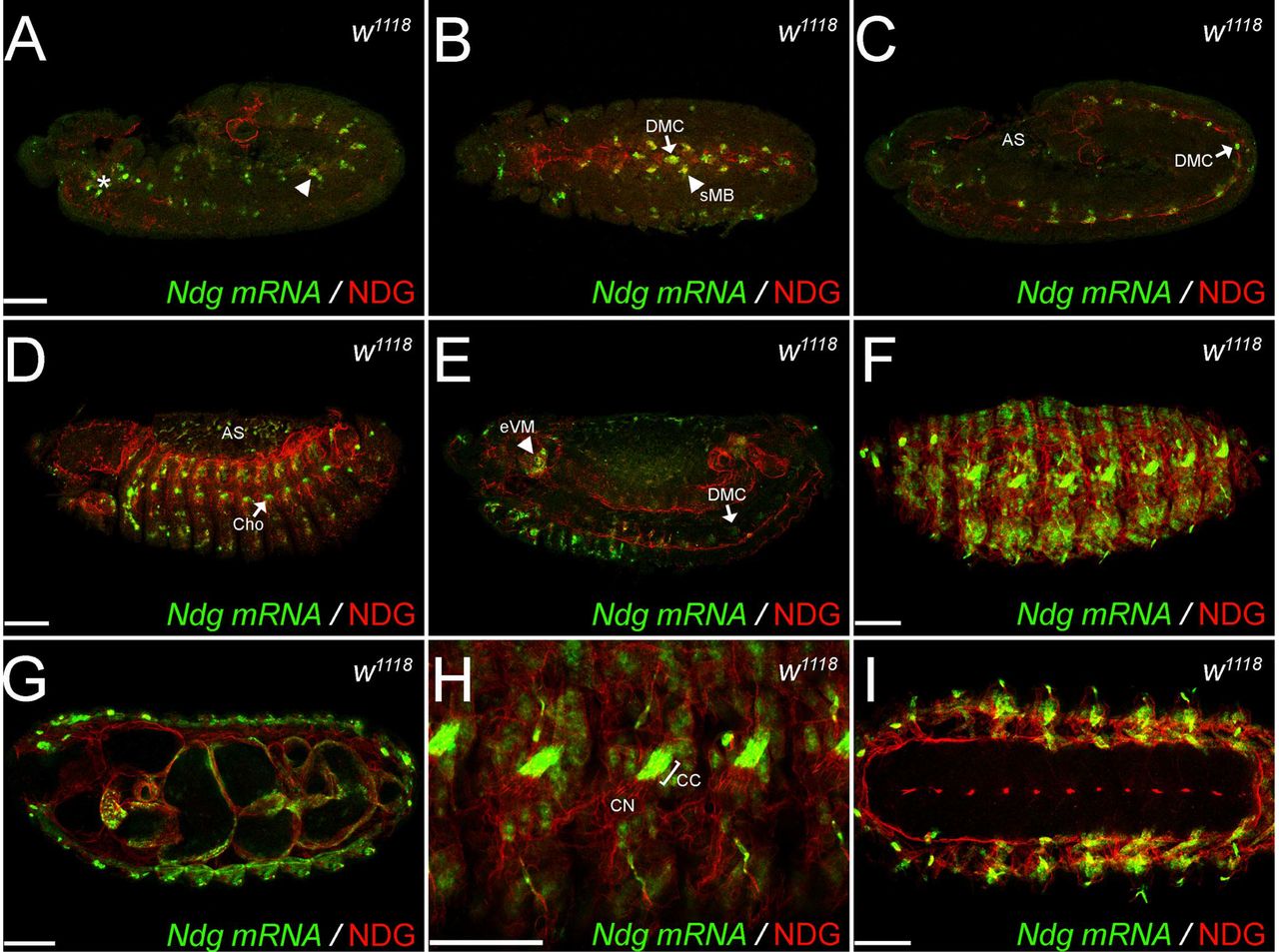
Confocal stacks of Drosophila white (w1118) embryos at developmental stage 12 (A-C), stage 14 (D, E) and stage 16 (F-I). Ndg mRNA (green) was visualized by fluorescence in situ hybridization (FISH), and antibody staining against NDG (red) reveals the secreted protein. Embryos are orientated in lateral views except for B, G and I which represent ventral views. AS: amnioserosa, CC: chordotonal cap cell, Cho: chordotonal organ, DMC: dorsal median cell, eVM: esophageal visceral mesoderm, CN: chordotonal neuron, sMB: somatic myoblast. Asterisk and arrowhead in (A) depict Ndg expression in the head region and the dorsal mesoderm respectively. Scale bars = 50 μm.
Localization of NDG to BMs depends on Laminin
We next asked whether the absence of other major BM components affects the incorporation and localization of NDG within the BM. Therefore we performed NDG antibody staining on late stage 16 embryos lacking either Laminin, type IV Collagen or Perlecan (Figure 2). Control siblings exhibited strong NDG staining of all embryonic BMs and around the chordotonal organs (Figure 2A). To differentiate between the NDG localization properties of both Laminin trimers we employed either LanA or wing blister (wb) loss of function mutations that impair formation and secretion of only one Laminin trimer (Urbano et al., 2009). Interestingly, we found a strongly disrupted and punctuated NDG pattern in transheterozygous LanA9-32/Df LanA embryos lacking the Laminin A-yielding trimer (Figure 2B), whereas we did not detect changes in wbHG10/Df wb embryos in which the Wb-containing trimer is absent (Figure 2C). The loss of all secreted Laminin trimers in embryos lacking the only Laminin y-subunit (LanB2knod/Df LanB2) resulted in a dramatic loss of NDG localization (Figure 2D) indicating a redundant function of both Laminin trimers for the localization of NDG to BMs. On the other hand, embryos deficient for trol (the Perlecan encoding locus) as well as embryos carrying a deficiency for the two Drosophila type VI Collagens (Col4a1 and viking) displayed no changes in NDG localization and distribution (Figure 2E, F) suggesting that Laminin is the primary crucial component for NDG distribution and localization to BMs.

Stage 16 embryos in lateral orientation were stained for NDG protein (green). (A) Control sibling embryo with balancer-associated YFP expression (red) displays proper NDG localization to BMs with strong accumulation at the chordotonal organs. (B) Punctate NDG pattern in a LanA9-32JDf LanA transheterozygous embryo. (C) NDG protein distribution appears unaffected in wbHG10JDf wb embryos. (D) Disruption of the NDG pattern in LanB2knodJDf LanB2 embryos. (E) Df trol (loss of Perlecan) embryos display a segmentation phenotype due to the additional loss of the adjacent giant locus but no changes in NDG localization. (F) Regular NDG distribution in an embryo deficient for the two Drosophila type VI Collagen encoding loci Col4a1 and viking (vkg). Scale bar = 50 μm.
Generation of Ndg deletion mutants
In order to investigate the function of Ndg in Drosophila, we employed imprecise excision of the Minos transposon insertion Mi{ET1}MB04184 to generate deletions in the Ndg locus (Sup. Fig 2A). Two excision lines were obtained in which 0.4 kb respectively 1.4 kb of the Ndg locus including parts of the 5’ UTR were deleted (henceforth referred to as NdgΔ0.4. and NdgΔ1.4, see also Material and Methods). Further analysis employing NDG-specific antibodies revealed that the strong NDG staining observed in control siblings (Sup. Fig 2B) was completely absent in homozygous NdgΔ1.4. embryos (Sup. Fig 2C), whereas faint NDG expression could be detected in the NdgΔ0.4. mutant (Sup. Fig 2D). Therefore, we concluded that the deletion in NdgΔ1.4. resulted in a protein null allele while the smaller molecular lesion in NdgA0A. caused a hypomorphic Ndg allele characterized by strongly reduced NDG expression.
Phenotypic analysis of Ndg mutants
Nidogen mutants generated in C. elegans and mouse are viable and display only mild phenotypes (Bader et al., 2005; Bose et al., 2006; Dong et al., 2002; Kang and Kramer, 2000; Murshed et al., 2000; Schymeinsky et al., 2002), a surprising finding which is in strong contrast to the essential developmental roles demonstrated for the other BM molecules (Arikawa-Hirasawa et al., 1999; Clay and Sherwood, 2015; Poschl et al., 2004; Yao, 2017). In agreement with these former observations, Drosophila NdgΔ mutants were viable and fertile and could be maintained as homozygous stocks. Despite the lack of any obvious developmental function we noticed several peculiarities in NdgΔ homozygous stocks that were more apparent in the NdgΔ1.4. line when compared to NdgΔ0.4. In non-crowded, standard culture conditions, NdgΔ pupal cases were preferentially formed in the lower half of the vial or even directly on the food, indicating potentially impaired larval climbing abilities (Sup. Figure 3A, B). We also noticed a difference in the orientation of the pupal cases which was shifted towards the horizontal axis in NdgΔ vials (Sup. Fig 3B). Moreover, we observed a decrease in fecundity in homozygous NdgΔ animals compared to the w1118. control (Sup. Fig 3C) and NdgΔ1.4. flies additionally exhibited incompletely inflated wing blades at low (~5 %) penetrance rates (Sup. Fig 3D, E).
Distribution of major BM components is not altered in Ndg mutant embryos
To analyze the proposed function of NDG as universal ECM cross linker that connects Laminin and Collagen layers of BMs (Timpl and Brown, 1996), we studied the distribution of known BM components in NdgΔ1.4/Df Ndg embryos. Therefore, we applied antibodies raised against Laminin subunits and trimers, type IV Collagens and Perlecan as well as the Collagen-associated molecule SPARC (Fig 3 and Sup. Fig 4). In addition, we employed GFP-trap insertions in the loci of viking (vkg::GFP), one of the two Drosophila type IV Collagen genes, and terribly reduced optic lobes (trol::GFP), encoding for Drosophila Perlecan (Sup. Figure 4).

(A-D) Distribution of LanA, LanWb, COLL IV and PCAN in control and transheterozygous NdgΔ1.4IDf Ndg embryos (E-H) at stage 16. Embryos appear in in lateral (D), dorsolateral (A, B, E, F, H) or ventrolateral orientation (C, G). Scale bar = 50 μm.
In case of Laminin, all antibodies employed stained embryonic BMs indicating reactivity with secreted Laminin (Figure 3A, B). Laminin A (LanA) outlined the BMs of all internal organs in control and Ndg mutant embryos (Fig 3A, E), whereas antibody staining against the Laminin a1, 2 subunit Wing blister (LanWb) was found mainly in the BMs around the digestive tract and the apodemes of control and Ndg mutant embryos (Fig 3B, E). This was in agreement with antibody staining against secreted K-cell Laminin (LanKc) which appeared similar in Ndg mutants compared to control embryos (Sup. Fig 4A, D). Additionally employed antibodies against the Laminin β- and γ-subunits (referred to as LanB1 and LanB2, Sup. Fig 4B, C, E, F) demonstrate BM staining of control and Ndg mutant embryos and due to the strong reactivity with non-secreted Laminin intermediates also labeling of hemocytes and fat body cells. Further analyses employing antibodies against Collagen IV (COLL IV, Fig 3C, G), the vkg::GFP protein trap (Sup. Fig 4G, J), and the collagen-binding protein SPARC (Sup. Fig 4H, K) revealed that these proteins were present in embryonic BMs of Ndg mutants. Finally, we did not observe obvious differences in Perlecan staining of control and Ndg mutant embryos detected by either PCAN-specific antibodies (Fig 3D, H) or trol::GFP (Sup. Fig 4I, L). Taken together, the complete loss of NDG seems not influence the formation or overall assembly of embryonic BMs.

(A, B) Confocal sections from somatic muscle preparations of control (A) and NdgΔ1.4 (B) larvae incubated with low weight TexasRed-coupled Dextran (TexRed-Dextran, white) to reveal BM leakage. Insets are higher magnifications of the highlighted areas. Confocal images were acquired with identical channel settings. (C-F) Visceral BM ultrastructure of dissected w1118. control (C) 3rd instar larvae and homozygous (D, E) and transheterozygous (F) Ndg mutants revealed by SEM. LVM: Longitudinal visceral muscle. Scale bars = 20 μm (A, B), 5 μm (C-F).
Loss of NDG affects BM ultrastructure and function
The viability of NdgΔ mutant flies and the presence of all BM proteins analyzed so far (Fig 3 and Sup. Figure 4) suggested that BM assembly was not severely affected by the absence of NDG, therefore questioning its proposed role as universal ECM linker molecule. However, the loss of NDG might impair BM stability and function. To address this in more detail we dissected w1118. and NdgΔ1.4. 3rd instar larvae and employed a Dextran-based permeability assay in addition to SEM ultrastructural analysis (Figure 4). Spot-like Texas Red (TexRed)-coupled Dextran plaques were observed across the surface of the somatic muscles of w1118 control larval filets (Figure 4A). In comparison, NdgΔ1.4. larval filets appeared much brighter after dextran staining and we observed a strong leakage of TexRed-Dextran into the muscle proper (Figure 4B) indicating enhanced diffusion through the BM tissue barrier in the absence of NDG.
Further ultrastructural analysis of w1118 control 3rd instar larvae revealed that the visceral BM which covers the surface of the larval midgut appeared as smooth sheet spread across the underlying longitudinal and circular visceral muscles (Figure 4C). In comparison, the surface of the visceral BM in homozygous NdgΔ mutant larvae exhibited holes (Figure 4D-F), which were frequently found in close vicinity to the longitudinal visceral muscles (LVM) but also appeared in other areas. We also observed a variation in phenotypic strength between individual larvae. While most null mutant NdgΔ1.4. animals displayed strong BM phenotypes, a minor fraction also exhibited weaker defects resembling NdgΔ0.4. larvae. On the other hand, BM defects observed in some of the analyzed NdgΔ0.4. larvae appeared similar to the null allele. Interestingly, examination of heterozygous NdgΔ/CyO larvae (Sup. Figure 5B-D) already revealed a weak BM phenotype characterized by few and small surface holes which was never observed in w1118. (Figure 4C) or in additional control larvae from the w*; KrIf-1/CyO balancer stock (Sup. Fig 5A), indicating a dose-dependent effect of NDG on BM morphology.
Loss of Ndg leads to BM disruption under osmotic stress
To further investigate whether the absence of NDG comprises the mechanical stability of the BM, we exposed wing imaginal discs to an osmotic shock by incubating them in deionized water. When water diffused into the hypertonic interior of the imaginal discs it created balloon-like swelling of the disc before eventual bursting. Imaginal discs of larvae resembling the genetic background of balanced and homozygous NdgΔ. animals (w1118 and w*; KrIf-1/CyO) withstand the resulting osmotic pressure for ~430 s respectively ~388 s before bursting. In case of wing discs derived from balanced, heterozygous NdgΔ animals we observed a slight but not significant increase (NdgΔ0.4/CyO = 472 s, NdgΔ0.4./CyO = 520 s; and Df Ndg/CyO= 407 s). In contrast to this NdgΔ0.4. wing discs burst after ~213 s and NdgΔ1.4. or NdgΔ1.4/Df Ndg discs tore after ~240 s indicating decreased BM stability (Figure 5).

Quantification of osmotic stress applied to 3rd. instar larval wing discs of the indicated genotypes. Means and SDs (whiskers) are shown. Compared to w1118. and w*; KrIf-1/CyO balancer controls, imaginal discs from Ndg mutant animals resist osmotic stress for a significantly reduced mean time (NdgΔ0.4/CyO = 472 s vs. NdgΔ0.4. = 213 s, NdgΔ1.4ICyO = 520 s vs. NdgΔ1.4., 239 s; and Df Ndg/CyO = 407 s vs. NdgA1A/Df Ndg, 241 s, n ≥ 20 wing imaginal discs). 1-W anova indicated significant differences among means (F(.7,188) = 11.8, p < 0.001) and unpaired, two-tailed t-tests were applied to compare heterozygous controls and homozygous samples (***p < 0.001,*p = 0.017).
Altered crawling behavior and NMJ morphology in Ndg mutant larvae
When placed on agar plates supplemented with yeast paste as food source NdgΔ1.4 1st instar. larvae were often observed outside the food and exhibited overall lethargic and reduced crawling movements (Figure 6A, B). To monitor and compare their behavioral repertoire we recorded age-matched w1118 controls and NdgΔ1.4 larvae that were placed on Agar plates in the absence of food (Fig 6C, D). Crawling of control w1118 larvae was dominated by smooth forward movement with occasional turns and pauses that allowed the larvae to explore the whole experimental arena (Figure 6C, E and Sup. Movie). In contrast, Ndg mutants generally moved within a much smaller area and displayed a variety of crawling defects including excessive head turning as well as exaggerated rolling and bending motions (Figure 6D, F and Sup. Movie). Interestingly, the phenotypic strength exhibited some variation since 70 % of 2nd instar larvae (n > 40) and half of the analyzed 3rd instar larvae (n > 40) displayed rather strong behavioral abnormalities whereas the remaining animals exhibited weaker crawling defects. We further analyzed crawling of NdgΔ 3rd instar larvae in more detail, investigating the velocity and the stride frequency of the undisturbed crawling pattern (Figure 6G, H). Comparison of the mean crawling velocity and the stride frequency showed comparable differences between the analyzed genotypes. The mean crawling velocity of NdgΔ0.4 mutant larvae was slightly but not significantly reduced whereas NdgΔ1.4 larvae showed a significantly reduced mean velocity compared to w1118 larvae (Figure 6G). Also the stride frequency in both NdgΔ strains was significantly lower when compared to w larvae (Figure 6H). However, NdgΔ mutants displayed the more severe phenotype in terms of reduced crawling speed and stride frequency (Figure 6G, H).

(A) Schematic example of the observed distribution of w1118.and NdgΔ1.4. 1st instar larvae (black dots) on agar plates (depicted as circles) outside a central food source (red). (B) Quantification of the larval distribution assay. Mean of larvae outside the food: w1118 = 1.44, NdgΔ0.4 = 2.31, NdgΔ1.4 = 19.1, NdgΔ0.4/NdgΔ1.4 = 8, NdgΔ1.4/Df Ndg = 19.3; 1-W anova p < 0.001, Tuckey’s multiple comparisons test with adj. p-value: w1118 versus NdgΔ0.4 = 0.947 non-significant (n.s.), w versus NdgΔ0.4/NdgΔ1.4. = *p < 0.01, w1118. vs. NdgΔ1.4. ***p < 0.001, w1118. vs. NdgA14/Df Ndg = ***p < 0.001, n = 400 larvae for each genotype. (C, D) Snapshots taken from crawling recordings of w1118. and NdgΔ1.4. 2nd instar larvae. (E, F) Representative tracking patterns of crawling w1118. and NdgΔ1.4. 2nd instar larvae. Individually colored lines represent recordings of 25 s. Time points are denoted if the larva left the monitored area before the end of recording. Time intervals in brackets indicate absence from the recorded area. (G) Quantification of mean crawling velocity measured as 5s of uniform crawling from recordings of w1118. and NdgΔ1.4. L3 larvae. Mean: w1118 = 1. 265, NdgΔ0.4 = 1.07, NdgΔ1.4 = 0.907 mmJs) (K-W anova p < 0.0001, Dunn’s-test with mult. adj. p-value: NdgΔ0.4 = 0.0597, NdgΔ1.4. ***p < 0.0001 versus w 1118. (H) Quantification of the stride frequency calculated from 5 strides in a row from recordings of w1118. and NdgΔ1.4 3rd. instar larvae. Mean: w1118 = 73.32, NdgΔ0.4 = 62.25, NdgΔ1.4 = 47.75 strides/min.; K-W anova p < 0.0001, Dunn’s-test with mult. adj. p-value: NdgΔ0.4.*p = 0.0207, NdgΔ1.4. ***p < 0.0001 versus w1118. Error bars represent SEM. N >23 larvae for each genotype (I, J) Neuromuscular junctions (NMJs, outlined by dashed boxes) innervating muscles 6J7 in the 2 nd larval abdominal segment of w1118. and NdgΔ1.4 3rd. instar larvae are stained with anti-Bruchpilot (BRP, green) and anti-HRP (red). Scale bars = 1 mm (C, E), 50 μm (I).
These crawling phenotypes prompted us to examine neuromuscular junction (NMJ) morphology in wandering 3rd instar larvae. We therefore dissected larval filets and stained them with anti-Bruchpilot (BRP) antibodies to label T-zones in the synaptic boutons and employed anti-HRP staining to reveal the overall NMJ morphology (Figure 6I, J). We focused on the NMJ at muscle 6/7 in the 2nd abdominal segment because it covers a considerably large area, innervates two muscles, and contains many synaptic boutons. While these NMJs in w1118 control larvae appeared to be equally sized, we observed shape and size differences of the corresponding NMJs in NdgΔ mutants, even within the same animal (Fig 6J). We also noticed a higher degree of synaptic branching and increased bouton density (Figure 6J and 8B, C) which appeared to be independent of the NMJ size. Taken together, these results suggest that loss of NDG leads to defects in NMJ organization presumably resulting in the climbing and crawling defects observed in NdgΔ mutant larvae.
Loss of NDG leads to reduced reaction to vibrational stimuli and aberrant chordotonal organ morphology
Our previous analyses revealed that developing chordotonal organs highly expressed Ndg in their cap cells and eventually became embedded in a NDG-positive BM. Notably, some of the larval crawling defects and the altered orientation of pupal cases observed in the absence of NDG could be interpreted as a result of impaired proprioception. Therefore, we wanted to characterize chordotonal function in NdgΔ mutants on a behavioral level and additionally analyze larval PNS morphology with respect to the chordotonal organs (Figure 7). Stimulus-induced, relative body length reduction upon applied vibrational stimuli was compared between 3rd instar w1118 control and NdgΔ larvae (Figure 7A, B). The body length reduction of control larvae in response to vibrational stimuli was on average ~14% whereas homozygous NdgΔ0.4 and NdgΔ1.4. larvae showed significantly reduced retraction values (Fig 7B). NdgA1A mutants again displayed the most severe phenotype with a nearly abolished reaction (~4% body length reduction) to vibrational stimuli (Figure 7B).

(A) Body length recordings of w1117control and NdgΔ0.4 and Ndg mutant larvae exposed to pulsed vibrational stimuli (indicated by black bars). (B) Quantification of body length reduction induced by vibrational stimulation in w1118 control and Ndg mutant larvae. The relative body length reduction (A body length) is calculated from the mean larval length before and during vibrational stimulation. Error bars represent SEM, n = 24, **p < 0.01 ***p < 0.001 versus w1118. K-W anova p < 0.0001 and Dunn’s-test with mult. adj. p-value was applied for statistical analysis. NdgΔ0.4 vs. w1118.p = 0.0061, NdgΔ1.4. p < 0.001 vs. w1118..(C, D) Morphology of the lateral pentascolopidial chordotonal organ (lch5) of control and NdgΔ1.4 larvae revealed by Jupiter::GFP fusion protein (green) expression, NDG-(blue), and HRP-(red) antibody staining. Arrows highlight the sensory cilia whereas neuronal cell bodies are marked by arrowheads. (E, F) HRP labeling was employed to reveal the nervous system in one hemisegment of control and NdgA1′ 4 mutant larvae. Structures of the peripheral nervous system (PNS) such as ddA neurons (surrounded by circles) transmitting nerves (arrow) and neuronal cell bodies (asterisks) are highlighted. Scale bars = 50 μm (C), 100 μm (E).
To reveal potential morphological changes underlying the observed behavioral phenotypes, we examined the chordotonal organs of 3rd instar larvae. Therefore, we utilized the Jupiter::GFP fusion protein which localizes to the microtubule network (Karpova et al., 2006). In addition, we employed anti-NDG and horseradish peroxidase (HRP) staining (Jan and Jan, 1982) to reveal the chordotonal BM and associated neurons respectively (Fig 7C, D). A NDG-positive BM surrounded the whole mechanosensory organ of Jupiter::GFP control larvae including the bulging cell bodies of the chordotonal neurons (Fig 7C, arrowhead) which closely aligned to the scolopale cell. A NDG-positive sheet also surrounded the row of aligned sensory cilia (Fig 7C, arrow). In contrast to this regular alignment, NdgΔ1.4. mutant larvae exhibited detachment of the chordotonal neurons and dislocation of their cell bodies (Fig 7D, arrowhead) from the scolopale cell as well as disrupted alignment of the sensory cilia (Fig 7D arrow). We noticed that the penetrance of these phenotypes varied even between the chordotonal organs of corresponding hemisegments in a single larva (Sup. Figure 6B, C). However, we did not observe chordotonal organs in Ndg mutant larvae that resembled wild-type morphology (Sup. Fig 6A).
In addition to the chordotonal organ defects we noticed aberrations in other parts of the larval peripheral nervous system (Fig 7F). The cell bodies of the dorsal dendritic arborization sensory neurons (ddA neurons) failed to cluster in NdgΔ1.4 mutant larvae (Fig 7F circles). Moreover, transmitting nerves frequently exhibited defasciculation (Fig 7E and F arrow) and positioning of neuronal cell bodies (Fig 7F asterisks) was often altered when compared to the controls (Fig 7E asterisks).
Ndg interacts genetically with the Lar receptor at the larval NMJ
The differences in shape and size between corresponding NMJs 6/7 in the 2nd abdominal segment and the higher degree of synaptic branching in NdgΔ1.4 mutants (Figure 7E, F) prompted us to analyze this phenotype in more detail with regard to potential genetic interaction partners. Upon closer examination of the NMJ geometry in NdgΔ mutant larvae we observed that in contrast to wild-type NMJs (Figure 8A) the overall branching was increased and major branching points were shifted towards the center of the junction (Fig 8B). We also observed bouton clustering and a significantly enhanced number of boutons per square at NdgΔ junctions suggesting that NDG actually suppresses NMJ maturation (Fig 8C). Notably, these defects were found to be independent from the variable NMJ size observed in NdgΔ animals. Moreover, heterozygous NdgΔ1.4/CyO NMJs exhibited increased bouton clustering and a tendency to branch prematurely (Figure 8D) suggesting that these phenotypic characteristics could be modified upon genetic interaction.
Homozygous as well as heterozygous null mutants of the receptor protein tyrosine phosphatase (RPTP) Leukocyte-antigen-related-like (Lar) have been described to exhibit a strongly reduced bouton number at the larval NMJ (Kaufmann et al., 2002). In agreement to this we found drastically reduced NMJs in the rarely occurring homozygous Lar.13.2 3rd instar larvae (data not shown), whereas heterozygous Lar13.2/CyO animals (Fig 8E) exhibited reduced branching and bouton numbers. In double-heterozygous Lar13.2, NdgΔ1.4./CyO larvae, we observed mutual repression of the phenotypes caused by one copy of either Lar.13.2 or NdgΔ. leading to NMJs that resembled a more “wild-type” morphology (Fig 8F). These results indicate that Ndg and Lar function together to promote differentiation of the larval NMJ in Drosophila.

NMJs of 3rd instar larva stained with anti-Bruchpilot (BRP, green) and anti-HRP (red). (A) Control NMJ of muscle 6/7 in the 2nd abdominal segment. (B) Ndg. mutant (NdgΔ1.4./Df Ndg) larvae exhibit increased and premature synaptic branching as well as increased bouton number and clustering. (C) Quantification of bouton densities (bouton number per area) of control and NdgΔ larvae. (D) Heterozygous NdgΔ1.4/CyO larvae display mild branching defects and increased bouton clustering. (E) Heterozygous Lar13.2ICyO NMJ with reduced branching and boutons. (F) Lar13.2, NdgA14/CyO larvae exhibit an intermediate phenotype when compared to the single mutations (D and E). Scale bar = 20 μm.
Discussion
Analysis of Ndg expression and NDG protein distribution
In this work we have characterized the single Nidogen/entactin gene in Drosophila melanogaster. In the developing embryo, Ndg expression followed a highly dynamic pattern involving diverse mesodermal cell types like somatic muscle founder cells, mature visceral and somatic muscles as well as dorsal median cells. Ndg expression in a subset of somatic muscle founder cells had been previously reported by (Artero et al., 2003) and detailed analysis of an intronic “Ndg muscle enhancer” revealed a complex interplay of Forkhead and Homeodomain transcription factors and their binding sites that are required to drive Ndg expression in the different mesodermal cell types (Busser et al., 2013; Busser et al., 2012; Philippakis et al., 2006; Zhu et al., 2012). Our analysis further revealed Ndg expression in the extraembryonic amnioserosa and in chordotonal cap cells indicating the presence of additional regulatory elements. We did not detect Ndg expression in the embryonic CNS but in the closely associated dorsal median cells (DMCs), which express other BM components and ECM receptors such as Perlecan, Dystroglycan, Glutactin as well as Laminin and type IV Collagen (Friedrich et al., 2000; Mirre et al., 1992; Montell and Goodman, 1989; Olson et al., 1990). DMCs provide cues for proper axonal pathfinding during transverse nerve outgrowth and regulate bifurcation of the median nerve in Drosophila (Chiang et al., 1994). Therefore, studying a potential role of the secreted ECM in this context would be an interesting topic for future investigations. Interestingly, Ndg expression was absent from mesoderm-derived hemocytes and fat body cells although these cells strongly secrete Laminin, type IV Collagens and other ECM components (Le Parco et al., 1986; Pastor-Pareja and Xu, 2011; Rodriguez et al., 1996; Urbano et al., 2009; Wolfstetter and Holz, 2012) indicating that Ndg expression is regulated independently from other BM genes.
CEL-Seq transcriptome analysis in C.elegans reveals high nid-1 mRNA expression in mesodermal tissues upon gastrulation (Hashimshony et al., 2015) and NDG proteins strongly associate with body wall muscles in Drosophila (this study), C. elegans, Ascidians and mice (Ackley et al., 2003; Fox et al., 2008; Kang and Kramer, 2000; Nakae et al., 1993) suggesting an important, evolutionary conserved function in this particular tissue. Indeed, C. elegans Nid-1 and NID2 in mice are enriched at neuromuscular junctions and, in agreement with our analyses in Drosophila, required for proper NMJ structure and function (Ackley et al., 2003; Fox et al., 2008).
NDG is not required for BM assembly
Our analysis of NDG distribution in the absence of major BM components revealed a dependency of NDG localization from Laminin but not from type IV Collagen or Perlecan. This is in agreement with studies on γ1III4-mice in which the NDG-binding site in the Laminin γ1 chain had been deleted. This analysis revealed a severe reduction of NDG from BMs as well as perinatal lethality and organ defects similar to those observed in NIDI−/−. NID2−/−. double mutant mice which further strengthens the assumption that Laminin plays an essential role in localizing NDG to forming BMs (Bader et al., 2005; Halfter et al., 2002; Willem et al., 2002). The two Laminin heterotrimers in the fly function in a redundant manner to localize NDG to embryonic BMs. In a single loss of function background, however, only absence of the LanA trimer (containing the Drosophila Laminin a3/5 homolog) affected NDG localization, suggesting different binding and polymerization activities of the Drosophila Laminins. In line with these findings, combined reduction of LanA and NDG around the developing gonad was also observed in βPS-integrin mutant embryos (Tanentzapf et al., 2007). No obvious changes in the distribution of BM core components (Laminins, Collagens and Perlecan) were found in our immunohistochemical analysis after complete loss of NDG, reflecting its non-essential role during development. This finding, further supported by similar analyses in other species (Ackley et al., 2003; Bader et al., 2005; Kang and Kramer, 2000; Kim and Wadsworth, 2000; Rossi et al., 2015) contradicts the proposed role for NDG as linker between inner and outer ECM networks during BM formation, suggesting a negligible role for NDG in overall BM assembly (Ho et al., 2008; Hohenester and Yurchenco, 2013; Jayadev and Sherwood, 2017; Kramer, 2005; Timpl and Brown, 1996; Yurchenco, 2011).
Loss of NDG affects BM stability and function
Although BM changes are barely detectable in NdgΔ. mutants by conventional immunohistochemistry, ultrastructural analysis revealed that BM continuity is compromised if NDG is absent or reduced. Our findings that BM barrier function and stability were decreased in NdgΔ larvae suggests that NDG, or NDG-mediated crosslinking of other ECM molecules, plays an important role in sealing the assembled BM. Intriguingly, Matsubayashi et al. (Matsubayashi et al., 2017) observe holes in the forming BM around the embryonic Drosophila CNS which are progressively closed during hemocyte migration and concurrent Col IV deposition. In addition, studies on mice demonstrate that although Laminin is sufficient for the assembly of early BM scaffolds, incorporation of type IV Collagen is essentially needed for BM integrity and stability (Poschl et al., 2004). Therefore, it is tempting to speculate that crosslinking of hemocyte-deposited Col IV might be impaired in Ndg mutants leading to decreased BM stability and barrier function.
The increased number of BM holes in close proximity to the visceral longitudinal musculature in Ndg mutants and the reduced tolerance to osmotic shock further implies that NDG protects BM integrity from mechanical stress therefore separating and maintaining different compartments in the larval body. While our experiments demonstrated reduced BM stability in NdgΔ. larvae, the BM defects observed in the background of different Ndg alleles exhibited some phenotypic variation. An explanation for this could be that, although the genetic background of the larvae determines overall BM stability, individual behavior, physical activity, and motility levels could modulate Ndg loss of function phenotypes. A comparable finding was made by Tsai and colleagues (Tsai et al., 2012) who reported a process in which larval NMJ size changes in response to LanA levels at the NMJ which are influenced by crawling activity as well as nervous responses to environmental cues. Given the NDG-localizing properties of LanA as well as the variations in NMJ size observed in NdgΔ animals, it is intriguing to think of a feedback mechanism that employs spatiotemporal reorganization of the larval BM by NDG to adapt NMJ morphology to altered environmental conditions.
Loss of NDG affects behavioral responses and PNS morphology
With the exception of NIDI−/− NID2−/− double mutant mice that die at birth, loss of NDG is generally associated with a range of rather subtle morphological and behavioral phenotypes. Notably, abnormalities described in the absence of NDG are not always fully penetrant or exhibit phenotypic variation (Bader et al., 2005; Bose et al., 2006; Dong et al., 2002; Hobert and Bulow, 2003; Kang and Kramer, 2000; Kim and Wadsworth, 2000; Murshed et al., 2000; Schymeinsky et al., 2002; Zhu et al., 2017). Moreover, subtle behavioral phenotypes can be enhanced by adjusting the experimental conditions (Ackley et al., 2003). In agreement with these analyses, characterization of Drosophila NdgΔ. mutants did not reveal an essential developmental function but a range of less overt, often variable phenotypes. After vibrational disturbance of a normal crawling phase, wild-type Drosophila larvae show a complex sequence of behavioral pattern. The larvae discontinue their forward movement and show a head retraction (“hunch”) followed by a head turning phase (“kink”) (Bharadwaj et al., 2013; Ohyama et al., 2013; Wu et al., 2011). This sequence of behavior is initiated upon mechanical stimuli delivered to the substrate which are detected by segmental chordotonal organs, mediated by stimulus processing in the central nervous system, and executed under neuromuscular control. Our analysis of the vibrational response behavior in NdgΔ. larvae shows a statistically significant reduction in body contraction. This finding points to a crucial role of NDG in vibrational response behavior. However, the precise level of NDG action cannot be inferred from these behavioral data. NDG expression in chordotonal cap cells, the morphological defects observed in larval chordotonal organs as well as the altered geotaxis of NdgΔ larvae suggest a contribution to mechanosensation. Defective wing inflation and the aberrant NMJ morphology observed in NdgΔ animals additionally indicate that loss of NDG affects the neuromuscular system. Indeed, previous research on C. elegans and mouse has uncovered a role of NDG for the structural development of neuromuscular junctions as well as altered locomotor behavior upon loss of NDG (Ackley et al., 2003; Bader et al., 2005; Dong et al., 2002; Fox et al., 2008). Given the high conservation of NDG throughout the animal kingdom (reviewed in (Ho et al., 2008)), this finding could hint at a possible comparable function for NDG in insects.
The receptor protein tyrosine phosphatase (RPTP) Leukocyte-antigen-related-like (Lar) and its cytoplasmic binding partner Liprin-α are required for proper synaptic morphology at the larval NMJ in Drosophila (Kaufmann et al., 2002). Interestingly, a specific splice form of LAR has been shown to bind a Laminin/Nidogen-containing protein complex in Hela cells and LAR-like RPTPs genetically interact with nid-1 to promote presynaptic differentiation in C. elegans (Ackley et al., 2005). Our analysis also reveals that a genetic interaction between Lar and Ndg is conserved at the larval NMJ in Drosophila. However, in contrast to the observations in C. elegans both factors affected overall NMJ morphology and acted antagonistically in this process.
In conclusion, our initial characterization of Ndg mutants in Drosophila neither revealed an essential developmental function nor did it support the proposed role for NDG as universal ECM-linker molecule. However, NDG appears to be important for BM sealing and stability, proper mechanosensation and neuromuscular function. Notably, our analyses suggest that NDG could be an important factor to ensure ECM plasticity in response to environmental cues.
MATERIALS AND METHODS
Fluorescence antibody staining
Antibody staining of Drosophila embryos and larvae was essentially performed as described by Mueller in (Dahmann, 2008). For antibody staining of wandering 3rd instar larvae we adapted the protocol from Klein in (Dahmann, 2008) with the following modifications: animals were relaxed before dissection by briefly dipping them into 60 °C water and a permeabilization step (10 min wash in PBS supplied with 1% Triton-X100) was added before blocking and primary antibody incubation. The following primary antibodies were used in their specified dilutions: mouse anti-Bruchpilot (Brp nc82, 1:100, (Wagh et al., 2006), DSHB), guinea pig anti-Collagen IV (COLLIV, 1:500, (Lunstrum et al., 1988)), sheep anti-Digoxygenin alkaline phosphatase conjugated Fab fragments (DIG-AP, 1:4.000, Roche Applied Science), mouse anti-Green fluorescent protein (GFP, 1:500, Roche Diagnostics), rabbit antiGreen fluorescent protein (GFP, 1:500, abcam), chicken anti-Green fluorescent protein (GFP, 1:500, abcam), goat anti-Horseradish peroxidase, Cy3-conjugated (HRP, 1:200, Jackson ImmunoResearch), guinea pig anti-Laminin A (LanA, 1:500, (Harpaz and Volk, 2012)), mouse anti-Laminin A (LanA, 1:500, (Takagi et al., 1996)), rabbit anti-Laminin B1 (LanB1, 1:400, (Kumagai et al., 1997), abcam), rabbit anti-Laminin B2 (LanB2, 1:400, (Kumagai et al., 1997), abcam), rabbit anti-K-cell Laminin (LanKc, 1:500, (Gutzeit et al., 1991)), rabbit anti-Wing blister (LanWb, 1:100, (Martin et al., 1999)), rabbit anti-Nidogen (NDG, 1:1.000, (Wolfstetter et al., 2009)), rabbit anti-Perlecan (PCAN, 1:1.000, (Friedrich et al., 2000)) and rabbit anti-SPARC (1:500, (Martinek et al., 2008)). Alexa Fluor-, Cy-, Biotin-SP-, and HRP-coupled secondary antibodies were purchased from Dianova and Jackson ImmunoResearch, DAPI from Sigma Aldrich. Embryos and larval tissues were embedded in Fluoromount-G (Southern Biotech) before visualization under Leica TCS SP2 or Zeiss LSM 800 confocal microscopes.
Developmental Northern blot
Northern blot analyses were performed by standard procedures (Maniatis, 1982). RNA was extracted by the guanidine thiocyanate/phenol/chloroform extraction method of Chomczynski and Sacchi (Chomczynski and Sacchi, 1987). Poly(A)-tailed RNA was isolated using a Pharmacia Kit (Pharmacia Biotech). A 2.5 kb fragment of the Ndg cDNA (cNdg5, Stefan Baumgartner unpublished) was radioactively labelled and hybridized to the Northern filter. Exposure time for Northern blots was 1.5 d. The blot was re-probed with a Drosophila 19S probe to evaluate equimolar loading. The Ndg blot shown in this work was identically conducted as those for wing blister (LanWb) and LanA (Martin et al., 1999) allowing scarce comparison of the expression levels of these different ECM genes.
Whole mount in situ hybridization
N-and C-terminal fragments from a full-length Nidogen cDNA clone (DGRC cDNA clone LP19846; GenBank: BT031149.1) were PCR-amplified and sub-cloned into the pCRII-TOPO vector with the TOPO TA Dual Promoter Kit (Invitrogen). Primer sequences for a 571 bp N-terminal fragment were GGACCCATCCATATCCCGCCACAAT and GCAATCAGTGCCACCTGGAAGGTGT while CGTGGCATTGCCGTGGATCCCT and GGTGCATCCTGTGGAGGCGCT were employed to amplify a 549 bp C-terminal fragment. Templates for probe synthesis were generated by PCR using M13 primers supplied with the TOPO TA Dual Promoter Kit. Digoxygenin (DIG)-labeled sense and antisense probes were made by SP6/T7 in vitro transcription with the DIG RNA Labeling Kit (Roche Applied Science). In situ hybridization on Drosophila embryos was performed according to (Lecuyer et al., 2008) with modifications adapted from (Pfeifer et al., 2012). Sheep anti-DIG-AP Fab fragments (1:4.000, Roche Applied Science), biotinylated donkey anti-sheep IgG (1:400, Dianova), the Vectastain ABC Standard Kit (Vector Laboratories) and the TSA Amplification Renaissance Kit (PerkinElmer) were used for fluorescence in situ hybridization (FISH) detection. Specificity of the probes was tested by in situ hybridization on white1118, and Ndg deficient Df(2R)BSC281 embryos, whereupon we detected no differences between the N-and the C-terminal probes (not shown).
Fly stocks and genetics
Flies were grown under standard conditions (Ashburner, 1989) and crosses were performed at room temperature or at 25 °C. Staging of embryos was done according to (Campos-Ortega and Hartenstein, 1997). The following mutations and fly stocks were used in this study: as control or wild-type stocks we employed white1118 (w1118), w*; Krlf-1/CyO P{Dfd-EYFP}2, Oregon-R or balanced sibling embryos. We used Df(2R)BSC281 as a deficiency for Ndg (Df Ndg), Df(2L)Exel7022 which deletes the two adjacent Drosophila type IV Collagen genes viking (vkg) and Col4a1 (previously referred to as Cg25C), Df(3L)Exel8101 as deficiency for Laminin A, Df(2L)TE35B-2 as deficiency for Laminin wing blister (Gubb et al., 1985), Df(3L)Exel6114 as Laminin B2 deficiency (Df LanB2) and Df(1)Exel6230 that removes the trol locus but also the adjacent segmentation gene giant. Mi{ET1}MB04184, P{hsILMiT} and P(A2-3}99B were used to generate the Ndg deletion alleles NdgA04. and NdgΔ1.4. (Sup. Fig 2, see below). The null alleles LanB2knod. (Wolfstetter and Holz, 2012) and wbHG10. were used as well as Lar13.2 that encodes for a truncated form of the Leukocyte-antigen-related-like receptor (Krueger et al., 1996). Protein trap lines were: vkg::GFP (Morin et al., 2001), trol::GFP (Medioni and Noselli, 2005), and Jupiter::GFP (Karpova et al., 2006).
Generation of Nidogen deletions
The pMiET1 transposon insertion Mi{ET1}MB04184 (Metaxakis et al., 2005) was used in an imprecise excision screen. Lethality reported for this line was not associated with the Mi{ET1} insertion and homozygous viable, isogenic stocks were established after ~5 generations of free recombination over a wild-type chromosome. The position of the Mi{ET1} insertion, initially revealed by flanking sequence recovery ((Bellen et al., 2011), GenBank: ET201740.1), was confirmed in the isogenic stocks by inverse PCR (Ochman et al., 1988). Thereby, we detected a 35 bp deletion 141 bp upstream of the insertion site that was present in the flanking sequence recovery data (ET201740.1) and in w1118. DNA samples but not in the 6th GenBank release of the ,D. melanogaster, genome annotation (Hoskins et al., 2015) or the corresponding sequence derived from Oregon-R genomic DNA. Therefore, we considered this small deletion as a naturally occurring sequence variation in the Ndg locus. Before serving as Minos transposase source in the screen, the P{hsILMiT} insertion (Metaxakis et al., 2005) was remobilized from the SM6a balancer and inserted onto a w1118. X-chromosome employing P(A2-3}99B as transposase source. pMiET1 remobilization was induced in the germ line of 2nd and 3rd instar larvae by daily 1 h heat shocks at 37 °C in the presence of the w1118, P{hsILMiT} helper chromosome. A total of 301 single excision events were identified due to the absence of the Mi{ET1}-associated Mmus/Pax6-GFP expression (Berghammer et al., 1999) and screened for deletions by single fly PCR analysis with the following primer combination: G C C AAG G AAT GGGAGTGCTCTGGAT and GGAGCCATCCTCGAACTCGTACAATT. Deletions were detected in two viable Mi{ET1}MB04184 excision lines (henceforth referred to as NdgΔ0.4. and NdgΔ1.4). Sequencing the molecular lesions uncovered a 1401 bp deletion (2R: 1031278010314180) and a P-Element remnant of 17 nucleotides (TGCCACGTAGCCGGAAT) in NdgΔ1.4 In the case of NdgΔ0.4. we detected a 417 bp (2R:10313549-10313965) deletion and an insertion of 19 nucleotides (CGAGCAAAATACAAAATAC) at the former P-Element insertion site (Sup. Fig 2).
Dextran permeability assay
Wandering 3rd instar larvae were picked and relaxed by briefly dipping them into 60 °C water. Larval filets were dissected and incubated in 50 Ml of 25 Mg/ml anionic Texas Red-coupled Dextran (10.000 MW, Thermo Fischer Scientific) in PBS for 10 minutes. The dextran solution was removed and the larval filets were rinsed once with 50 Ml PBS. Larval filets were fixed for 10 min in a drop of 4% formaldehyde in PBS and rinsed with PBS before mounting in Fluoromount-G and analysis under a Zeiss LSM 800 confocal microscope.
Larval climbing and gravity assay
Matching numbers of flies were placed on food-filled vials and allowed to lay eggs for three days. Pupae were examined at the pharate adult stage (7-9 days after egg laying at 25 °C). To assay larval climbing performance, the distance between the food surface and the cotton plug was divided into four zones. Pupae formed in each zone were counted. Pupae positioned on the border between two zones were assigned to the next higher zone. Pupae were further assigned into three categories (1. Upright = 0° “head up” or 180° “head down” +/− 22.5°, 2. Tilted = 45° or 225° +/− 22.5°, and 3. Flat = 90° or 270° +/− 22.5°) according to their orientation along the gravity axis.
Scanning electron microscopy (SEM)
Wandering 3rd instar larvae were dissected in PBS. After 3 h fixation in 2.5 % glutaraldehyde in PBS, samples were rinsed several times in PBS and dehydrated in an ascending ethanol series (50 %, 70 %, 80 %, 90 %, and 2x 100 % ethanol for 10 min each). After critical-point-drying in a Balzers CPD 030 at 40 °C, samples were mounted on stubs using double-sided adhesive tape and sputter-coated with a thin layer of gold (Balzers SCD 004). Samples were analyzed under a Zeiss DSM982 scanning electron microscope. Acceleration voltage was set to 3 kV. Settings for tilt angle, spot size, scanning mode and magnifications were kept constant throughout image acquisition.
Osmotic stress assay
Wandering 3rd instar larvae of indicated genotypes were picked and rinsed with PBS. The larvae were cut in half and the anterior part was inverted in order to expose the wing imaginal discs. After removing the anterior part of the gut and fat body, the samples were transferred to distilled water and the time until bursting of the first wing imaginal disc was measured. Measurements were stopped after 10 min and wing imaginal discs that were found intact after this period were assigned to the 10 min group.
Lethality tests
Parental flies (3-7 days old) of the indicated genotype were shifted on grape juice agar plates for 8 days. Agar plates were changed every 2nd day and defined numbers of eggs were transferred to fresh agar plates supplemented with dried yeast for further comparative analysis. Undeveloped eggs, hatched larvae, pupal cases and enclosure were counted daily. All plates were incubated at 25 °C and constantly humidified.
Larval feeding assay
Fifty freshly hatched 1st instar larvae were transferred to apple juice agar plates with a central spot of food-dye-supplemented yeast paste. Larvae moving outside the food were counted 1 and 2 h after the transfer. Fresh yeast paste was added every day and individual larvae were checked for the uptake of the dyed food after 72 h. The assay was performed at 25 °C and 60 % humidity conditions.
Larval locomotion assay
Flies were allowed to lay eggs for 2 h on apple juice agar plates supplemented with yeast paste. Fresh yeast paste was added every day and individual 2nd instar larvae were picked after 72 h (at 25 °C and 60 % humidity) and rinsed with distilled water. A single larva was placed in the middle of a 0 52.5 mm Petri dish prepared with a 0 35 mm central arena (1 % agar in PBS) surrounded by a ring of high salt agar (3 M NaCl in PBS) as locomotion barrier. For every recording a fresh agar plate was used. Larvae were allowed to move freely in the central arena for 2-5 min respectively and locomotion was recorded in a square of 12,5×9,5 mm2 using a Zeiss AxioZoom.V16 stereo zoom microscope equipped with LED ring light and an Axiocam 503 color camera. Time series were recorded with 4 frames/second employing ZEN Blue software and 25 s subsets were further analyzed and edited with the manual tracking plugin MTrackJ and the Fiji distribution of ImageJ (Schindelin et al., 2012).
Larval crawling assay
Larval crawling analyses were performed as described by (Eschbach et al., 2011) for responses to vibrational stimulation and measurement of body length and stride frequency. For the trials, 3rd instar larvae were placed on 1 % agarose covered Petri dishes (60 mm diameter) as a crawling stage. This stage was placed in a silencing foam covered box (57×57×47 cm3.) and illuminated by a red darkroom lamp (Pf712em, Philips, 15 W and 7 lm). Video recordings of the trials were performed via an infrared CMOS-Color-Camera (CCD-651, Conrad Electronics). The analogue video signals were grabbed by using an analogue/digital video converter (700-USB, Pinnacle Systems) and recorded via a computer for further analysis (25 fps). Measurements of crawling speed, stride frequency and body length of the larvae were performed by using the video tracking software Cabrillo Tracker (v4.91, Douglas Brown, Open Source Physics, 2015). To quantify mean velocity and stride frequency for five larvae per trial, the terminal ends were video-tracked for 10-20 s. Only periods of unrestricted and constant crawling behavior were used for analysis. The mean velocity was calculated from 5 s of crawling and the stride frequency from the duration of 5 complete strides in a row. The maximum crawling speed of a single stride was defined as the starting point for each measurement. To determine body length reduction due to vibrational stimuli, apical and terminal ends of four larvae per trial were video-tracked. The vibrational stimuli were generated via the audio edition software Audacity (v2.05, Audacity-Team, GNU GPLv2+, 2015) at a frequency of 100 Hz and delivered via a full-range speaker (BPSL 100/7, Isophon/Gauder Akustik, 60-20.000 Hz and 7 W) to the crawling stage. For each trial, three stimuli with the duration of 1 s and an acceleration of 12 m/s2 were applied (interpulse-interval 9 s).
Statistical analyses
Prism v6.0 and v7.02 (GraphPad Software Inc.) was used for all statistical analyses.
Funding
This work was supported by:
SB: Vetenskapsrådet (621-2003-3408) and Cancerfonden (4714-B03-02XBB).
RHP: Vetenskapsrådet (621-2015-04466), Cancerfonden (2015/391), Barncancerfonden (2015/0096), and Göran Gustafsson Stiftelser.
AH: Deutsche Forschungsgemeinschaft/DFG (Ho-2559/3-3 and 2559/5-1).
Author contributions
GW: Investigation, methodology, conceptualization, visualization, data curation, formal analysis, supervision, validation, writing original draft, writing review and editing.
ID: Investigation, visualization, formal analysis, validation and writing original draft.
KP: Investigation, visualization, formal analysis, validation and writing original draft.
JA: Investigation, visualization, formal analysis, validation and writing original draft.
UT: Investigation, visualization, formal analysis, validation and writing original draft.
DP: Investigation, visualization and writing original draft.
RL-H: Supervision.
SB: Investigation, writing original draft and funding acquisition, writing review and editing.
RHP: Supervision, funding acquisition, writing review and editing.
AH: Investigation, conceptualization, methodology, visualization, supervision, validation, funding acquisition, project administration, writing original draft, writing review and editing.
Competing interests
The authors declare that they have no competing interests.
Supplementary Figure Legends

Total mRNA from adult flies (lane 1), embryos from 0-6 h, 6-12 h, 12-18 h (lanes 2-4), 3rd instar larvae (L3, lane 5) and pupae was extracted and detected by an Ndg-specific probe. No signal was detectable up to 6 h of embryonic development as well as in L3 larvae and adult flies. Later embryonic stages and pupae exhibit Ndg mRNA expression with the strongest intensity around 12 to 18 h of embryonic development. 19S rRNA was used as loading control.>

(A) Schematic representation (created with (Hulo et al., 2008;Rambaldi and Ciccarelli, 2009)) of the NDG protein domain organization (top) and the Ndg locus (below) including the positions of the mobilized P-element Mi{ET1}MB04184 and the obtained genomic deletions NdgA1′ 4 and NdgΔ0.4; 4. Protein domains and parts of the locus that encode for these domains are highlighted in matching colors. The position of the epitope employed to generate the NDG antibody (Wolfstetter et al., 2009) is indicated by a red line. NIDO: NIDO domain, EGF: EGF-like domain, Nidogen G2: Nidogen G2 p-barrel domain, LY:.LDL-receptor class B (LDLRB)/YWTD repeat. (B-D) Confocal stacks from dorsal views of stage 16 Drosophila embryos stained for NDG (red). GFP (green) depicts balancer-associated reporter gene expression in a sibling control. DAPI and auto fluorescence of yolk droplets appears in one channel (white). (B) Strong NDG staining of BMs in control embryos. (C) Absence of detectable NDG protein in embryos homozygous for the NdgΔ1.4. deletion. (D) Decreased NDG protein levels in a homozygous NdgΔ0.4. embryo. Confocal images were acquired with identical settings for the Cy3-detecting channel. Scale bar = 50 μm.
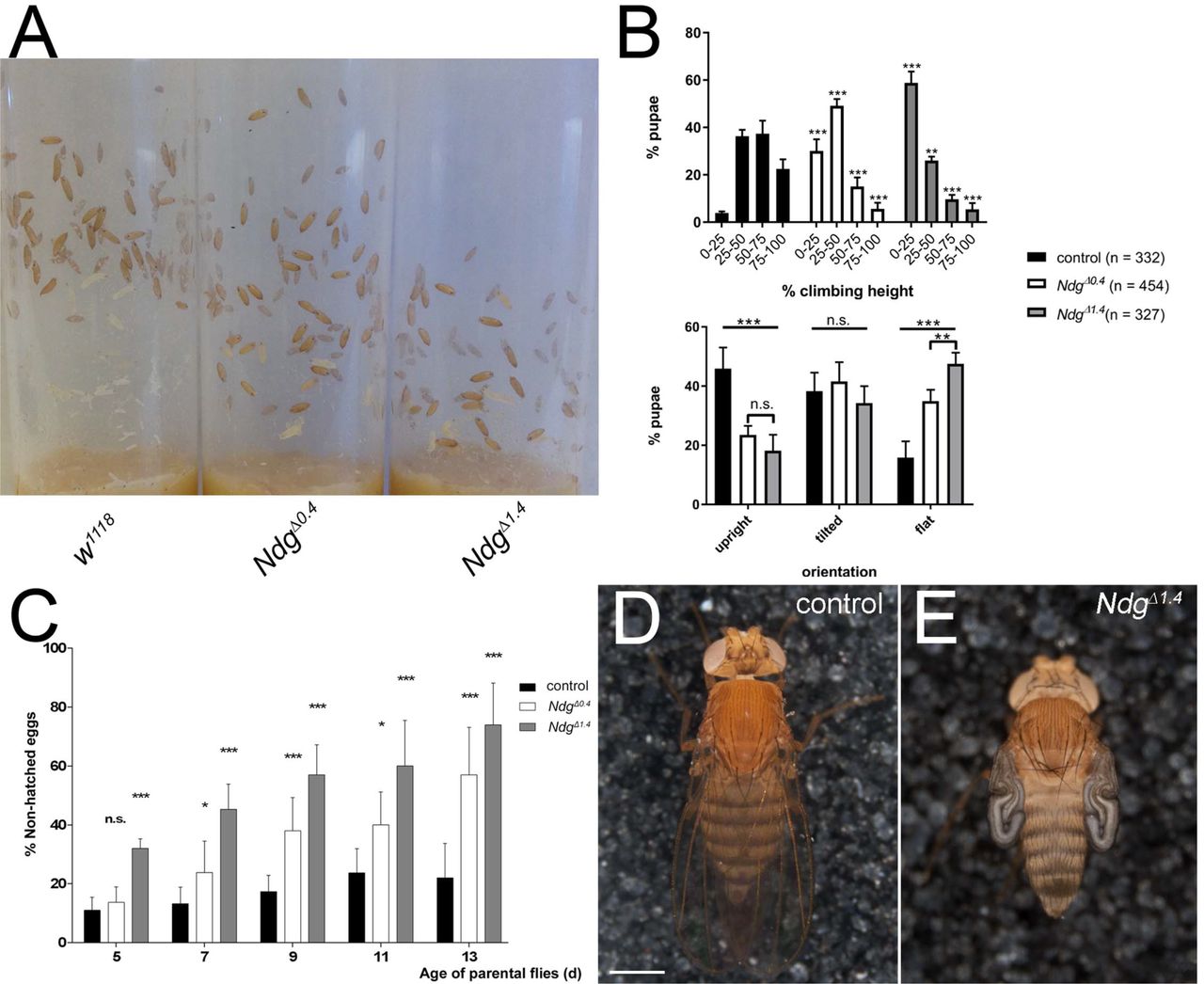
(A) Formation of pupal cases in vials of white1110 control and homozygous NdgΔ. stocks (matching numbers of parental flies and breeding days at 25 °C). (B) Quantification of the climbing and orientation phenotypes. In comparison to the control, NdgΔ pupal cases are preferentially found in the lower half of the vials indicating impaired climbing performance (2-W anova and Tuckey’s multiple comparisons test with adjusted p-values, *** = p < 0.001, **p = 0.005, non-significant (n.s.) = p > 0.05). In NdgΔ vials, the orientation of the pupal cases is significantly shifted towards the horizontal axis (2-W anova and Tuckey’s multiple comparisons test with adjusted p-values, ***p < 0.001, **p = 0.008, non-significant (n.s.) = p > 0.05). (C) Percentage of non-hatched eggs produced by 5 to 13 days old control and NdgΔ flies incubated at 25 °C. NdgΔ produce a significantly increasing number of non-hatched eggs over time, (2-W anova and Tuckey’s multiple comparisons test with adjusted p-values, *** = p < 0.001, ** = p < 0.01, * = p < 0.05 non-significant (n.s.) = p > 0.05). (D) Image of a control fly exhibiting properly inflated wings (n = 3492 animals). (E) Impaired wing inflation is observed at low penetrance in NdgΔ1.4. flies (140 of n = 2839 animals). Scale bar = 500 μm.

Antibody staining against LanKc, LanB1 or LanB2 labels BMs, and in the case of LanB1 and LanB2 additionally fat body and hemocytes of control and transheterozygous NdgΔ1.4IDf Ndg (D, E, F) embryos at stage 16. (G, H, J, K) A similar localization pattern was observed for vkg::GFP and SPARC in control and NdgΔ1.4IDf Ndg embryos. (I, L) trol::GFP localization in control and NdgAA4/Df Ndg embryos. Embryos appear in lateral (B, C, E, F, G, I, K, L) or ventrolateral (A, D, H, J) orientation. Scale bar = 50 μm.

BM ultrastructure of (A) w*; Kμ/CyO control 3rd. instar larvae and (B-D) heterozygous Ndg mutants with indicated genotype revealed by SEM. LVM: Longitudinal visceral muscle. Scale bars = 5 μm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Jupiter::GFP fusion protein (green) localizes to the microtubule network of the chordotonal organ in a Jupiter-GFP control larva. The chordotonal organ is surrounded by a NDG containing BM (blue) that also covers HRP-labelled neurons (red, arrowhead) and sensory cilia (arrows). (B, C) Two hemisegments of a Jupiter-GFP; NdgA14. larva exhibit neuron detachment (arrowheads) and alignment defects of sensory cilia (arrow) in the lateral pentascolopidial chordotonal organ. Scale bar = 50 μm.
Supplementary Movie: Recordings of crawling patterns from white1118 control and NdgA14. 1st instar larvae.
Acknowledgements
Bloomington Drosophila Stock Center (NIH P40OD018537), Lynn Cooley, the Drosophila Genomics Resource Center (DGRC, NIH 2P40OD010949-10A1), the Developmental Studies Hybridoma Bank (created by the NICHD of the NIH and maintained at the University of Iowa), Liselotte Fessler, the KYOTO Stock Center at the Kyoto Institute of Technology, Maurice Ringuette, John Roote, Talila Volk and Gerd Vorbrüggen for sending materials and fly stocks.
References




