

Abstract
Gene duplication and subsequent evolutionary divergence have allowed conserved proteins to develop unique roles. The MarR family of transcription factors (TFs) has undergone extensive duplication and diversification in bacteria, where they act as environmentally-responsive repressors of genes encoding efflux pumps that confer resistance to xenobiotics, including many antimicrobial agents. We have performed structural, functional, and genetic analyses of representative members of the SlyA/RovA lineage of MarR TFs, which retain some ancestral functions, including repression of their own expression and that of divergently-transcribed multidrug efflux pumps, as well as allosteric inhibition by aromatic carboxylate compounds. However, SlyA and RovA have acquired the ability to counter-silence horizontally-acquired genes, which has greatly facilitated the evolution of Enterobacteriaceae by horizontal gene transfer. SlyA/RovA TFs in different species have independently evolved novel regulatory circuits to provide the enhanced levels of expression required for their new role. Moreover, in contrast to MarR, SlyA is not responsive to copper. These observations demonstrate the ability of TFs to acquire new functions as a result of evolutionary divergence of both cis-regulatory sequences and in trans interactions with modulatory ligands.
Introduction
As organisms adapt to new or changing environments, their regulatory networks must evolve to ensure that individual genes are appropriately expressed in response to environmental signals (1). An important mechanism for the evolution of conserved essential proteins, including transcription factors (TFs), is gene duplication, which allows the subsequent diversification of gene and protein families and the development of new functions (2). More than 50% of bacterial genes are believed to have descended from original duplication events (3-5), providing a broad foundation from which bacteria can evolve complex and adaptive traits.
The MarR family is an ancient family of TFs, predating the divergence of archaea and bacteria (6). It has undergone extensive gene duplication events, with recent estimates suggesting that bacteria encode an average of seven MarR TFs per genome (7). MarR TFs typically function as environmentally-responsive repressors of genes encoding efflux pumps that export xenobiotics, including many antimicrobial agents, and are defined by the presence of a winged helix-turn-helix (wHTH) DNA-binding domain (8). The prototypical MarR protein of Escherichia coli represses a single operon, marRAB, which encodes a transcriptional activator (MarA) required for the expression of the AcrAB efflux pump, which in turn confers resistance to β-lactams, quinolones, and tetracyclines (9-11). MarR is allosterically regulated by many small molecules, in particular small aromatic carboxylate compounds such as salicylate, which induce a structural change that reduces the affinity of MarR for DNA (10, 12-14) and derepresses the expression of its cognate promoters. A recent study suggests that MarR can also be inhibited by intracellular copper (Cu2+), which oxidizes a conserved cysteine residue at position 80, promoting the formation of disulfide bonds between MarR dimers and causing individual dimers to dissociate from DNA (15). Free copper is thought to be liberated from membrane-bound cytoplasmic proteins during envelope stress induced by antimicrobial agents. Dimerization of MarR TFs is required for DNA binding, as it allows these proteins to recognize palindromic sequences via the α4 recognition helix, which makes sequence-specific contacts with the major groove, while the wing makes sequence-independent contacts via the minor groove (16).
A duplication event producing the SlyA lineage of MarR TFs most likely resulted from an ancient horizontal gene transfer event or from intragenomic recombination of a MarR family TF prior to the divergence of the Enterobactericeae. SlyA has been best characterized in Salmonella enterica serovar Typhimurium, where it serves primarily to upregulate virulence genes (17-19). Although this contrasts with the classical repressive role of MarR TFs, work in our and other laboratories has demonstrated that SlyA positively regulates genes by a counter-silencing mechanism, in which repression of AT-rich promoters by the histone-like nucleoid-associated protein H-NS is relieved by SlyA (17, 20). SlyA cooperatively remodels the H-NS-DNA complex in concert with the response regulator PhoP (20, 21), which is activated by conditions found within phagosomal compartments, including low Mg2+ (22), acidic pH (23), and cationic antimicrobial peptides (24). SlyA orthologs, represented by Hor, Rap, and RovA in Pectobacterium (Erwinia), Serratia, and Yersinia, respectively (25), are conserved in nearly every species of Enterobacteriaceae, even including endosymbionts such as Sodalis glossinidius (26), which have undergone extensive gene loss and degenerative evolution (27). This high degree of conservation suggests that the SlyA lineage occupies an essential role in the regulatory network organization of Enterobacteriaceae. Although conclusive mechanistic evidence to demonstrate that other SlyA orthologs function as counter-silencers has not yet been obtained, existing evidence is strongly suggestive of counter-silencing, as several are known to up-regulate horizontally-acquired traits, which are generally repressed by H-NS, in a number of species, including Yersinia spp. (28-30), Dickeya dadantii (31), Pectobacterium carotovorum (25) and Shigella flexneri (32).
TFs can evolve in two ways: in cis, through their promoters and associated regulatory elements, both transcriptional and post-transcriptional, altering expression patterns to respond to different environmental and physiological stimuli, and in trans, affecting their interactions with cognate binding sites, other proteins, and regulatory ligands. We sought to understand the evolutionary transition of the SlyA/RovA TF lineage from the ancestral function of MarR family TFs as environmentally-responsive and dedicated repressors of small regulons to counter-silencers of extensive networks of horizontally-acquired genes, with a particular focus on in cis changes in gene expression and in trans changes in modulation by inhibitory ligands. Structural and comparative analyses of representative members of the SlyA lineage were performed to identify the evolutionary changes that allowed SlyA to adopt its new role. Here we show that SlyA has retained an ability to undergo conformational changes in response to aromatic carboxylates, regulate gene expression in an environmentally-responsive manner, and repress the expression of a linked drug efflux system. However, SlyA/RovA lineage genes have undergone extensive evolution in cis to support the higher levels of expression that are required for counter-silencing. Finally, we show that linked efflux pumps are not conserved in some Enterobacteriaceae, even though SlyA/RovA TFs have been evolutionarily retained, suggesting that these regulators have been conserved not due to their primordial role in regulating antimicrobial resistance but rather as a consequence of their counter-silencing function, which is essential to maintain the regulated expression of horizontally-acquired genes in Enterobacteriaceae.
Results
Salicylate-mediated inhibition of SlyA activity
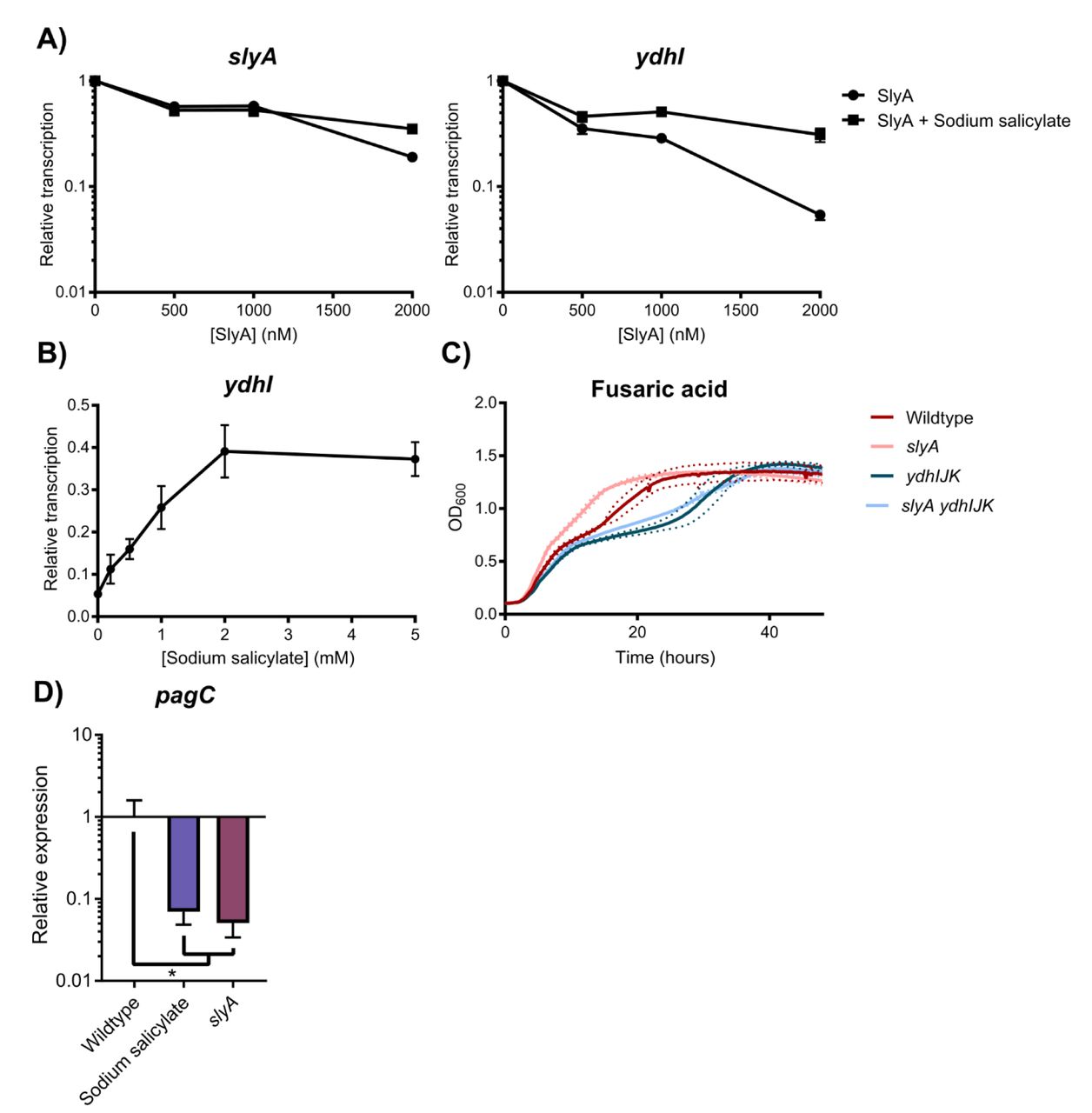
As environmentally-responsive repressors whose conformation and regulatory actions are modulated by small aromatic carboxylates (12, 14), MarR family TFs are inhibited by salicylate in vitro (12). In their structural analyses of SlyA-DNA interactions, Dolan et al, (33) inferred from our structural data (see below) that salicylate might regulate SlyA. Using electrophoretic mobility shift assays, they demonstrated that salicylate inhibits DNA-binding by SlyA. To confirm that this influences the function of SlyA as a transcriptional regulator, we performed in vitro transcription assays (IVTs) of slyA and the divergently transcribed ydhIJK efflux pump operon. Supercoiled plasmid DNA containing the slyA-ydhIJK region was incubated with RNA polymerase (RNAP) and increasing SlyA concentrations in the presence or absence of salicylate. SlyA repressed slyA transcription approximately 5.3-fold, while ydhI transcription was inhibited ∼19-fold (Figure 1A, B). The addition of 2mM sodium salicylate reduced SlyA-mediated repression to 2.8-fold and 3.2-fold, respectively, indicating that the sensitivity to aromatic carboxylates observed in classical MarR TFs has been retained by SlyA.

To determine whether SlyA is an autorepressor like other members of the MarR family, in vitro transcription (IVT) assays were performed on supercoiled template DNA containing slyA and the divergently expressed ydhI gene in the presence of increasing SlyA concentrations (A). Reactions were performed with or without 2mM sodium salicylate to determine whether SlyA is inhibited by small aromatic carboxylate compounds like other MarR TFs. (IVT analysis of SlyA-mediated regulation of pagC is shown in Fig. S1.) To determine whether SlyA exhibits a dose-dependent response to salicylate, IVT assays were performed on ydhI in the presence of 2µM SlyA and increasing concentrations of sodium salicylate (B). Wildtype, slyA, ydhIJK, and slyA ydhIJK cultures were grown in the presence of 30 µg/ml fusaric acid and cell density (OD600) measured over time to determine if ydhIJK encodes a functional anti-microbial efflux pump (C). Data represent the mean (solid line) of three independent experiments, each consisting of three replicates. Dashed lines represent the SD. (Growth curves in the absence of fusaric acid is shown in Fig. S2.) To determine whether salicylate also inhibits SlyA counter-silencing activity, pagC transcripts were quantified by qRT-PCR from early stationary phase (OD600≈2.0) cultures in minimal N-medium containing 10 µM MgCl2 in the presence or absence of 2 mM sodium salicylate (D). Transcript levels are normalized to rpoD, and data represent the mean ± SD; n≥3. Asterisk indicates P=0.05.
We then confirmed that the ydhIJK operon encodes a functional antimicrobial efflux pump by growing wildtype, slyA, ydhIJK, and slyA ydhIJK mutant strains in the presence of an aromatic carboxylate with antimicrobial activity, fusaric acid (Figure 1C). Both the ydhIJK and slyA ydhIJK mutant strains exhibited delayed growth in the presence of fusaric acid relative to the wildtype strain, suggesting that ydhIJK confers fusaric acid resistance in wildtype S. Typhimurium. Conversely, slyA mutants exhibit improved growth in the presence of fusaric acid, as predicted when ydhIJK is derepressed. Collectively, these observations indicate that the S. Typhimurium SlyA TF has retained ancestral functions characteristic of the MarR family.
To determine whether salicylate inhibits SlyA-mediated counter-silencing as well as repression, expression of the counter-silenced pagC gene was measured by qRT-PCR in the presence or absence of 2mM sodium salicylate (Figure 1D). Wildtype cultures grown in salicylate phenocopied a slyA mutant strain, with a >14-fold reduction in pagC expression, indicating that salicylate is a general allosteric inhibitor of SlyA function, most likely inducing a structural change that reduces affinity for DNA as described in other MarR TFs (8, 10). Unexpectedly, SlyA retained an ability to interact with DNA upstream of the ydhI promoter even in the presence of salicylate (Figure 2 - figure supplement 1), but this interaction may represent non-specific interactions with the wing domain, as salicylate inhibited SlyA interaction with a 12 bp DNA region that is highly homologous (75% identity) to the consensus high affinity binding site (19, 34, 35), centered near the −35 of the ydhI TSS.

To assess salicylate-induced SlyA structural changes, 1H,15N-TROSY NMR spectroscopy was performed on 0.3mM uniformly-labelled 15N-SlyA in the absence (black) or presence (red) of 2mM sodium salicylate. (Differential DNA Footprinting Analysis, or DDFA, of the SlyA-DNA interaction in the presence or absence of salicylate is shown in Figure 2 – figure supplement 1 Ligand binding analysis is shown in Figure 2 – figure supplement 2.)
To determine whether salicylate-mediated inhibition correlates with structural changes in SlyA, 1H,15N-TROSY NMR spectra of S. Typhimurium SlyA in the presence or absence of salicylate were collected. The apo-SlyA spectrum is well dispersed (Figure 2), with ∼85% of the expected resonances observed. However, wide variation among individual resonances with respect to peak width and intensity may signify [1] weak non-specific interactions between SlyA dimers, [2] varying rates of exchange of amide protons with solvent, or [3] conformational exchange. Previous studies of MarR found that apo-MarR is also highly disordered (8), suggesting that the observed spectral characteristics of apo-SlyA can be ascribed to the presence of multiple conformational states in solution. Addition of salicylate to SlyA induces chemical shift perturbations throughout the spectrum and nearly 90% of expected resonances are now observed. The large-scale changes in chemical shifts show that backbone amides throughout the protein are stabilized in a different environment relative to apo-SlyA. These observations are consistent with ligand binding to SlyA dimers inducing global structural changes, likely stabilizing a single protein conformation in solution, a conformation that is no longer able to interact with specific high-affinity DNA binding sites. The affinity of the SlyA-ligand interaction was determined via the quenching of intrinsic tryptophan fluorescence in the presence of increasing concentrations of benzoate, an analogous small aromatic carboxylate compound (Figure 2 - figure supplement 2). Benzoate induces similar chemical shift perturbations to those observed in response to salicylate but does not cause inner filter effects that interfere with fluorescence measurement, as there is relatively little overlap between the UV spectra of benzoate and SlyA, in contrast to salicylate. Although this assay was not able to differentiate between multiple SlyA-benzoate interactions, the KD of the SlyA-benzoate interaction was determined to be ∼40µM, which is similar to the previously determined affinities for EmrR/MprA and HucR (1-10µM), and significantly stronger than that of the MarR-salicylate interaction (0.5-1mM) (13, 36).
Crystal structure of salicylate-SlyA
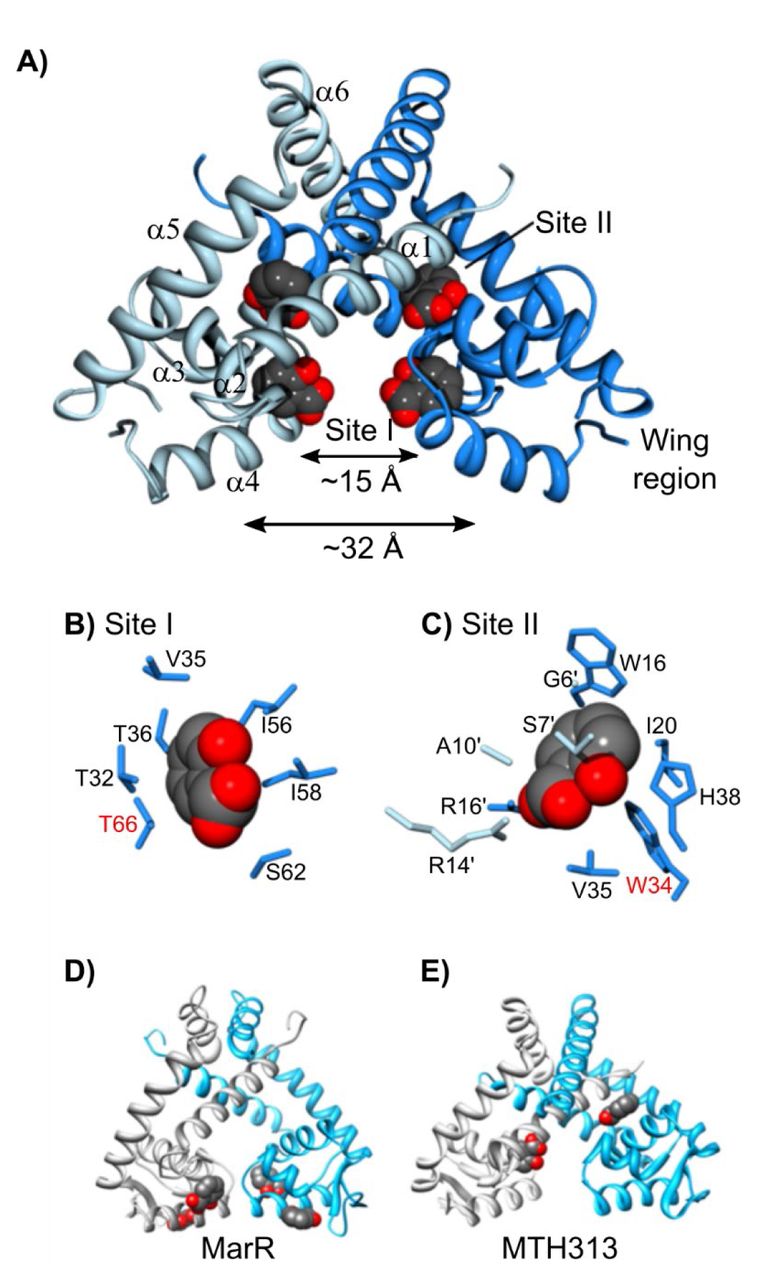
To further analyze the mechanism of allosteric inhibition of SlyA, the structure of the salicylate-SlyA co-crystal was determined. Studies by other groups have previously determined the structure of apo-SlyA and the SlyA bound to DNA (33), demonstrating that SlyA is similar in overall structure to other MarR proteins, consisting of six alpha helices (Figure 3A). Helices α1, α5, and α6 make up the dimerization domain, while α3 and α4, along with the wing region between α4 and α5 comprise the wHTH DNA-binding domain. These two domains are separated by α2. Dolan et al. (33) previously observed that the two recognition helices of the apo-SlyA dimer are only ∼15Å apart, in a closed conformation. During interaction with a high-affinity binding site, the helices move a significant distance to accommodate the 32Å distance between major grooves.

The crystal structure of the SlyA dimer with salicylate bound at sites I and II was determined at a resolution of 2.3Å (A). Residues involved in coordinating bound salicylates are shown for sites I (B) and II (C) Residues required for salicylate-mediated inhibition of SlyA (see Fig. 4) are highlighted in red. The structures of salicylate-bound MarR (D; PDB entry 1JGS) and MTH313 (E; PDB entry 3BPX) dimers are shown for comparison. (An image of the SlyA-salicylate crystal is shown in Figure 3 - figure supplement 1.)
SlyA formed large crystals in the presence of 75mM sodium salicylate (Figure 3 - figure supplement 1). However, we were unable to obtain usable crystals of apo-SlyA. Diffraction data set and refinement statistics are summarized in Tables S1 and S2, respectively. The two SlyA molecules in the asymmetric unit form two different SlyA dimers in this crystal form. Space group symmetry operations generate the other subunit in each dimer. The dimers are very similar in structure, and further discussion will focus on the dimer formed by polypeptide chain A. We were unable to observe electron density for the tips of the wings, so these regions are absent from our structural model. Difference electron density maps (|Fo|-|Fc|) identified two salicylate molecules bound per SlyA monomer at sites referred to as Site I and Site II (Figure 3A-C). Salicylate molecules interact with these sites via hydrophobic interactions with their aromatic rings, while the carboxylate and hydroxyl groups are positioned to interact with the solvent. Site I is composed of residues from α2, α3, and α4, as well as I58 in the loop between α3 and α4, and is well positioned to sterically inhibit DNA binding. It should be noted that the residue numbers in the deposited PDB file are not in register with the residue numbers in this text which are based on alignment of SlyA orthologs. Comparison with apo-SlyA and SlyA-DNA structures (33) indicates that this salicylate molecule causes the α4 recognition helix to rotate by ∼35° around its axis, disrupting specific contacts with the DNA major groove. Site II is formed by residues from both dimer subunits, almost completely sequestering the salicylate molecule from the solvent. The buried polar groups of salicylate interact with S7’ and R14’ in α1 of one subunit and R17 in α1 and H38 in α2 from the other subunit. A third salicylate binding site was observed on the surface of each subunit of the dimer. However, this site is adjacent to SlyA residues involved in crystal packing contacts and may not be biologically relevant.
Mutational analysis of allosteric inhibition of SlyA
We constructed a series of slyA alleles with site-specific mutations of the salicylate-binding pocket in order to test the functional significance of salicylate interactions in vivo. When wildtype slyA is expressed in trans from its native promoter, pagC expression increases over 350-fold in inducing medium containing 10µM MgCl2 compared to cultures grown with salicylate (Figure 4A). We tested 8 different mutant alleles for changes in counter-silencing activity in response to salicylate. One allele substituting an alanine for tyrosine 66 in site I (T66A), resulted in complete abrogation of salicylate-mediated inhibition, suggesting that T66 is essential for salicylate binding. However the T66A mutation also decreased pagC expression in the absence of salicylate over 25-fold, indicating that it is required for the wildtype activity of SlyA. A second mutant, W34A in site II, also exhibited reduced salicylate-mediated repression. The analysis of these mutants indicates that both site I and site II bind salicylate in vivo, and that both sites influence SlyA activity. Notably, both T66 and W34 are absolutely conserved in over 55 enterobacterial genera examined in this study (see below), suggesting that these residues are important for SlyA function throughout the Enterobacteriaceae (Table S3).

Selected residues involved in coordinating SlyA-bound salicylate were mutated in pSL2143, a plasmid expressing slyA from its native promoter, and assayed for their ability to restore counter-silencing in a chromosomal slyA mutant. Cultures were induced in 10µM MgCl2-containing medium in the presence or absence of 2mM sodium salicylate (A). Levels of pagC mRNA were quantified by qRT-PCR and normalized to rpoD. The mutant strain carrying the empty pWSK29 vector was included as a control. To determine if SlyA was directly inhibited by Cu2+ via C81, pagC expression was quantified in wild-type or isogenic slyA and slyA C81S mutant S. Typhimurium strains in inducing medium in the presence or absence of 100µM CuCl2 (B). To confirm that Cu2+-mediated inhibition occurs under these conditions, marA transcription was also quantified in the presence or absence of 100µM CuCl2. Data represent the mean ± SD; n=3. Asterisks indicate P < 0.005.
To determine whether salicylate directly inhibits SlyA in vivo or stimulates the release of intracellular Cu2+ from membrane bound proteins to promote disulfide bond formation, we mutated the single cysteine residue in SlyA, C81. SlyA C81A exhibited a similar salicylate inhibition phenotype to wildtype SlyA, indicating that allosteric inhibition does not occur by disulfide bond formation between cysteine residues in vivo. We subsequently generated a chromosomal C81S mutant to determine whether SlyA is directly inhibited by Cu2+. Transcription of pagC in strains encoding wildtype or C81S slyA was reduced approximately two-fold after the addition of 100µM CuCl2 to the growth medium (Figure 4B). Although this difference was statistically significant (p<0.001), it was also observed in the mutant strain encoding C81S SlyA, indicating that this modest effect is not due to disulfide-dependent tetramerization as proposed for MarR (15). Furthermore, modulation by copper does not appear to be biologically significant in comparison to the 350-fold effect of salicylate. Cu2+-mediated derepression of the marRAB operon was confirmed under comparable conditions by measuring marA expression, which increased approximately 7.5-fold (Figure 4C).
Evolutionary analysis of the SlyA TF lineage
To identify other variables that may have contributed to the evolution of the SlyA lineage, we performed an evolutionary analysis of species representing 60 genera of Enterobacteriaceae, including candidatus organisms for which genomic data was available. SlyA is strongly conserved, with orthologs found in 55 organisms, suggesting that SlyA plays a central role in enterobacterial regulatory circuitry. Genera lacking identifiable SlyA orthologs include Buchnera, Hamiltonella, Samsonia, Thorsellia, and Plesiomonas, which are only distantly related to Salmonella and Escherichia in comparison to other members of the Enterobacteriaceae. Notably, we were also unable to identify an hns ortholog in Plesiomonas. Phylogenetic analysis reveals five clusters of SlyA orthologs (Figure 5), with the general structure of the phylogram resembling the genomic tree for Enterobacteriaceae (37). SlyA orthologs in enteric pathogens including E. coli, S. Typhimurium, and S. flexneri form cluster I, while cluster II is composed of a more heterogeneous group of organisms, including plant pathogens such as D. dadantii, insect endosymbionts like S. glossinidius, and more distantly related pathogens such as Y. pseudotuberculosis. SlyA orthologs in clusters I and II are known to function as pleiotropic regulators, despite significant divergence from the other clusters (28, 31). Clusters III and IV are comprised of environmental organisms such as the plant-associated species Pantoea agglomerans and Phaseolibacter flectens, while cluster V contains hydrogen-sulfide producing bacteria such as Pragia fontium. This degree of conservation suggests that SlyA diverged from the greater MarR family of TFs prior to the divergence of Enterobacteriaceae from other bacteria. Although not strongly conserved outside of Enterobacteriaceae, SlyA orthologs can be detected in selected species throughout the Gammaproteobacteria, indicating that the lineage is ancient. However, the slyA gene typically exhibits an AT-content (51% in S. Typhimurium) marginally higher than that of the chromosomal average (48% in S. Typhimurium), suggesting that it may have been duplicated through an ancient transfer event.

The evolutionary history of the SlyA TF lineage was inferred using the Maximum Likelihood method and the Le-Gascuel model (85) with Mega X software (86). The tree is drawn to scale with the branch length representing the number of substitutions per site. The slyA genes have evolved in five major phylogenetic clusters: cluster I comprises most enteric species, including S. Typhimurium and E. coli; cluster II is heterogeneous, containing more distantly related pathogenic species from Proteus and Yersinia, as well as endosymbionts like Sodalis; clusters III and IV contain several plant-associated bacteria, such as Pantoea agglomerans and Phaseolibacter flectens; cluster V is comprised of hydrogen sulfide-producing bacteria. (A multiple sequence alignment of selected SlyA orthologs can be found in Figure 5 - figure supplement 1).
To determine if allosteric inhibition is likely to be conserved throughout the SlyA lineage, we aligned the sequences of SlyA orthologs from different bacterial species to analyze the conservation of residues involved in salicylate binding (Table S3). Cluster I exhibited nearly complete conservation of the salicylate-binding residues, with only four of 24 genomes encoding polymorphisms. Cluster II exhibited the greatest variation, with 16 of 23 genomes encoding polymorphisms, including 10 which encoded the polymorphism H38Y. We identified only one polymorphism in site I in six genomes, a T32I substitution, suggesting that site I is particularly important for SlyA function. As further evidence of the importance of site I, a T66A mutation abrogated allosteric inhibition of SlyA. The central residues of site II (R14, W16, R17, and W34) are similarly conserved suggesting that these residues are also important for SlyA function. We were able to mutate two of these residues (Figure 4A), R14 and W34, and found that W34 is also involved in allosteric inhibition. Individual polymorphisms such as T32I and H38Y appear to have evolved independently in multiple clusters, suggesting purifying selection, although their effect on SlyA activity is currently unknown. Alignment of SlyA protein sequences from representative species of each of the five clusters revealed that most sequence variation occurs in the carboxyl-terminus oligomerization domain (Figure 5 - figure supplement 1), suggesting that ligand sensitivity and DNA binding are conserved features of the SlyA lineage.
As the ancestral function of MarR TFs is the negative regulation of genes encoding drug efflux pumps, a function that is conserved in S. Typhimurium, we examined slyA orthologs throughout the Enterobacteriaceae for linkage to flanking genes of known or hypothetical function: slyB, which encodes an putative outer membrane lipoprotein with no observed phenotype, ydhJ which encodes a hemolysin D homolog, and ydhK, encoding the efflux pump (Figure 6A, B). It should be noted that ydhJ also exhibits homology to emrA, which encodes an antimicrobial efflux pump-associated protein linked to and regulated by another MarR family TF called MprA or EmrR (38, 39). We found that cluster I exhibited the strongest linkage, with 100%, 95%, and 100% linkage to ydhJ, ydhK, and slyB respectively, while cluster V, containing the hydrogen sulfide-producing bacteria, exhibited no linkage to slyA for any of the genes examined. Outside of cluster V, slyB was strongly associated with slyA, exhibiting 76% linkage overall. However, the ydh operon was not strongly linked outside of cluster I, exhibiting 26%, 33% and 33% linkage in clusters II, III, and IV respectively. In contrast, although marR was absent from 26 of the enterobacterial species examined, marR was linked to marAB in every marR-carrying species. This suggests that the primary function of MarR is to regulate the marRAB operon, and MarR consequently does not play an important role in enterobacterial regulatory circuitry. In contrast, the loss of the ancestral linkage between slyA and ydhJ outside of cluster I suggests that the SlyA lineage has evolved to serve a distinct function.
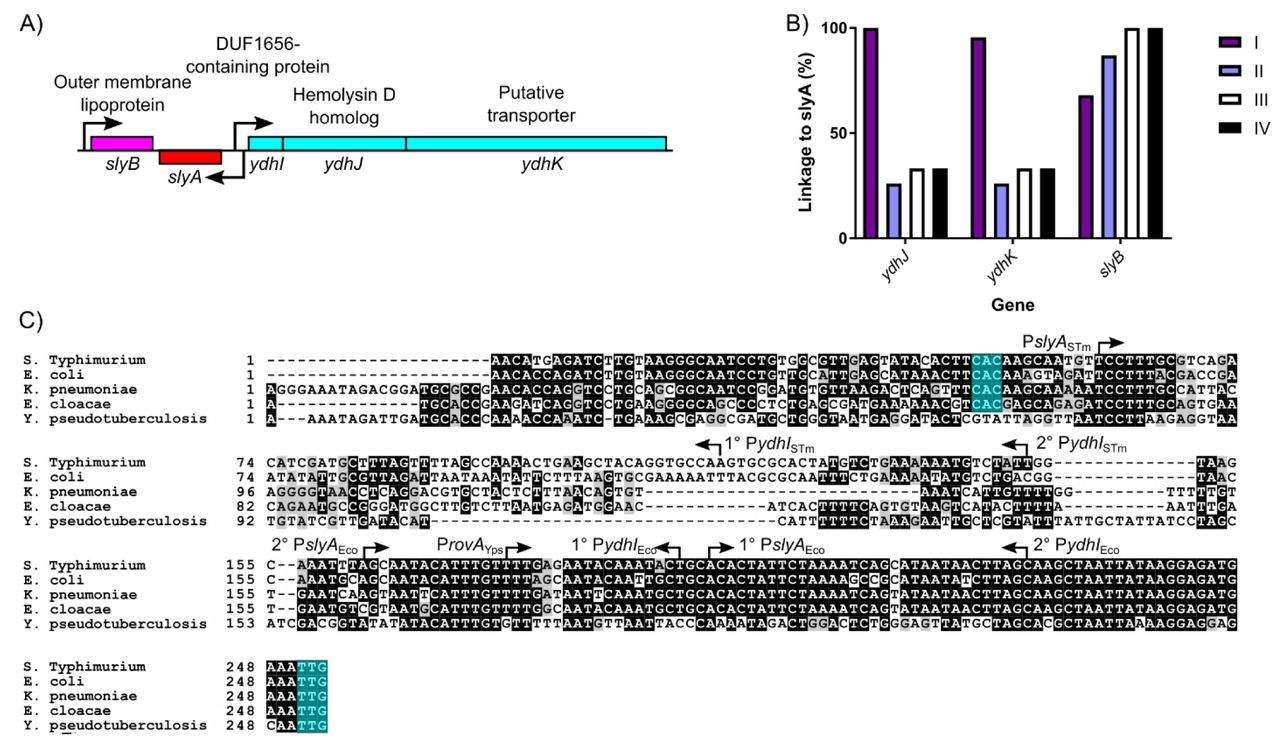
The slyA region of S. Typhimurium and closely related enteric species is diagrammed in (A). Arrows indicate TSSs, and boxes represent protein coding sequences. The ydhIJK efflux pump operon is transcribed divergently from slyA. A protein of unknown function is encoded by ydhI, and ydhJ and ydhK encode a hemolysin D/EmrA homolog and a transporter protein, respectively. slyB encodes a putative outer membrane lipoprotein, transcribed convergently to slyA. The linkage of slyA to slyB, ydhJ, and ydhK, is determined for all of the species from clusters I-IV shown in Fig. 5 (B). Species from cluster V do not exhibit linkage to these genes and are not shown. A multiple sequence alignment of the slyA promoter region from the closely-related species S. Typhimurium, E. coli, K. pneumoniae, and E. cloacae, as well as more distantly-related Y. pseudotuberculosis (C). Arrows indicate the position and orientation of previously characterized TSSs (30, 55, 57). Where multiple TSSs have been identified, the primary (1°) and secondary (2°) start sites are indicated. The slyA (positions 248-250) and ydhI (positions 47-70, dependent on the species) start codons are highlighted in teal when present. (Alignments of this region from representative species of Salmonella, Escherichia, and Yersinia are shown in Figure 6 - figure supplement 1. A phylogenetic analysis of the slyA promoter region of a subset of genera is shown in Figure 6 - figure supplement 2.)
The ydhIJK operon does not appear to have been exchanged for another drug efflux system. (Figure 6C). Sequences homologous to the proximal portion of the ydhI gene are detectable even in species that have not retained functional coding sequences (e.g., Y. pseudotuberculosis). However, a 100 bp segment of the ydhI-slyA intergenic region, beginning approximately 95 bp upstream of the slyA start codon, has undergone extensive mutation (Figure 6C), suggesting in cis evolutionary adaptation in the slyA lineage of TFs. We examined the regions upstream of slyA in several related species of three genera, Escherichia, Salmonella, and Yersinia, to better understand the recent evolution of this region, and observed that the ydhI-slyA intergenic regions exhibit considerable divergence between genera and conservation within genera (Figure 6 - figure supplement 1). A comparison between the ydhI-slyA intergenic regions of Escherichia coli and Salmonella Typhimurium with those of Klebsiella pneumoniae and Enterobacter cloacae revealed similar degrees of divergence between each genus, suggesting that an ancestral allele can no longer be defined. A phylogenetic analysis of slyA upstream regions in a representative subset of the species described above demonstrated extensive variation throughout the Enterobacteriaceae (Figure 6 - figure supplement 2). However, this variation did not always correlate with SlyA coding region clusters. Although the intergenic regions of cluster I are closely related and may have co-evolved with their respective coding sequences, the other clusters are more variable, and some intergenic regions may have evolved independently of their coding sequences.
Examination of the ydhI-rovA region in Yersinia spp. reveals that rovA in Y. pseudotuberculosis (Figure 6C) and Y. pestis (Figure 6 - figure supplement 1) is not linked to a functional ydhIJK operon, whereas ydhIJK is retained in Y. enterocolitica. This indicates that ydhIJK was not lost by the former species until after the divergence of Yersinia from other Enterobacteriaceae and suggests that other species of Enterobacteriaceae are also likely to have lost ydhIJK recently, as each became adapted to its own specific niche. This represents an example of parallel evolution, with multiple species independently losing ydhIJK to accommodate in cis evolution as their respective SlyA orthologs adapted to new roles.
Functional characterization of a distantly related SlyA ortholog
To understand how the SlyA lineage adapted to its emergent role as an important pleiotropic regulatory protein in the Enterobacteriaceae, we compared the functions of SlyA from S. Typhimurium (SlyASTM) and RovA of Y. pseudotuberculosis and Y. enterocolitica, a relatively divergent ortholog that also functions as a counter-silencer (28-30, 40). RovA of Y. pseudotuberculosis exhibits 76% identity with SlyASTM (Figure 5 - figure supplement 1). RovA is essential for virulence in Y. enterocolitica, and Y. pestis (28, 41) and has been suggested to function both as an activator, interacting with RNAP (42), and as a counter-silencer, alleviating H-NS-mediated repression (29, 30, 40). Notably, direct activation has not been demonstrated for any other member of the SlyA lineage, and the most direct evidence to suggest that RovA functions as an activator is derived from IVT studies using small linear fragments of DNA as a template (42). We have previously shown that small linear fragments are not necessarily representative of physiological regulatory events in intact cells and can generate spurious results in IVT assays (17). Genetic analysis of the inv gene, which is positively regulated by RovA, in an hns mutant strain of E. coli also suggested that RovA functions as both a counter-silencer and an activator. However, these experiments failed to consider the potential contribution of the H-NS paralog, StpA, which is up-regulated in hns mutants and can provide partial complementation (43, 44), which complicates the interpretation of regulatory studies in an hns mutant strain.
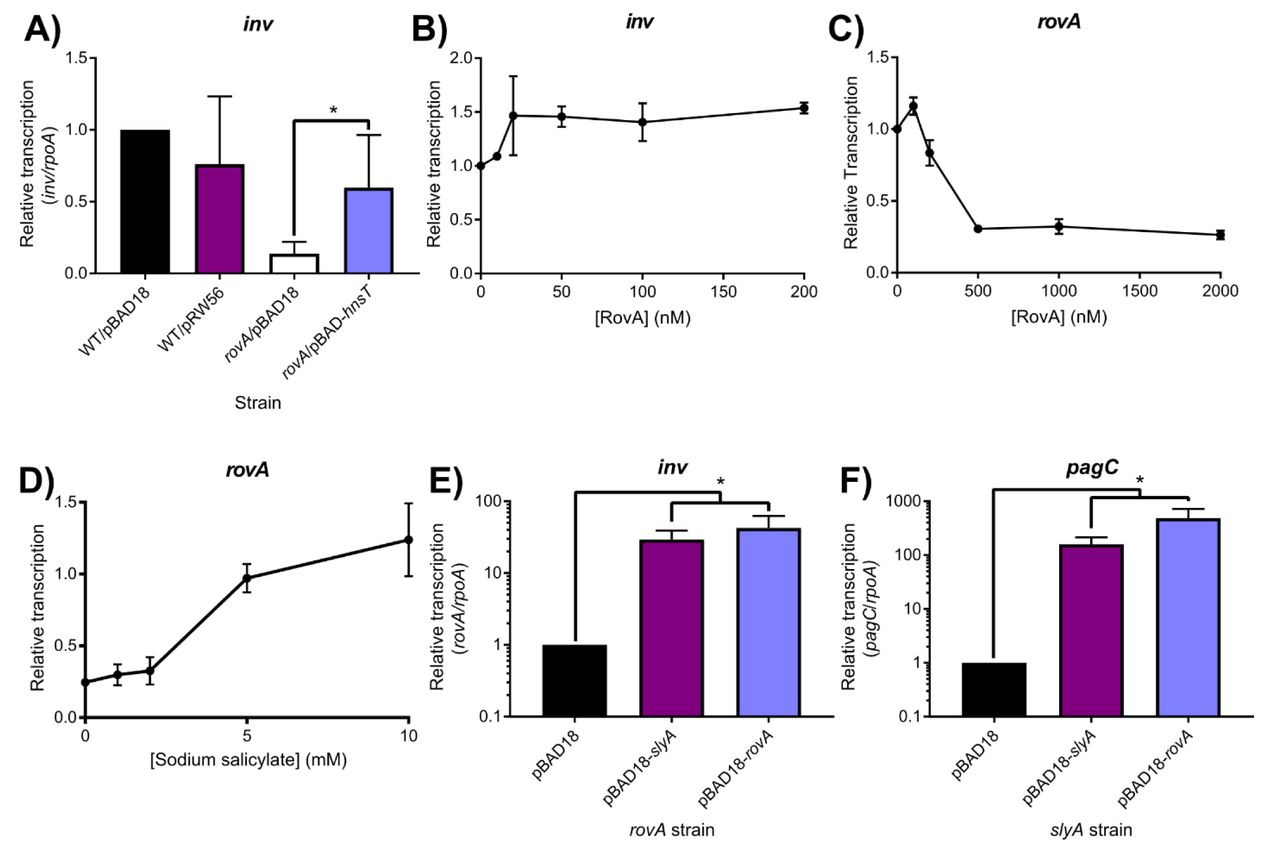
We attempted to corroborate the published findings by performing IVT analysis of inv expression in the presence of RovA. However, expression of inv exhibited only a very modest (∼1.5-fold) increase following the addition of RovA (Figure 7A), which became saturated at a 20 nM concentration; these results are not supportive of direct inv activation by RovA. In contrast, RovA was confirmed to function as an auto-repressor, like other MarR family TFs, as IVT analysis demonstrated a 4-fold decrease in rovA expression following the addition of RovA protein, with the effect reaching saturation at a 500 nM concentration (Figure 7B). RovA has also retained the ability to respond to salicylate (Figure 7C), despite the presence of an H38Y substitution in the second salicylate-binding pocket (Table S3), as 5mM salicylate completely inhibited RovA-mediated repression, similar to our observations with SlyA. It is also notable that RovA has retained salicylate sensitivity despite the loss of the linked YdhIJK efflux pump in Y. pseudotuberculosis. To prove that RovA functions as a counter-silencer in Y. pseudotuberculosis, we measured inv expression in wild-type and rovA mutant strains expressing H-NST from enteropathogenic E. coli (H-NSTEPEC). Mutations in hns cannot be generated in Yersinia as Yersinia spp. carry a single essential hns gene, unlike many other members of the Enterobacteriaceae which encode hns-like genes such as stpA, which are able to partially compensate for the loss of hns. H-NSTEPEC is a truncated hns homolog that has been demonstrated to function as a dominant negative form of H-NS by binding and inhibiting the activity of wild-type H-NS protein (45). The expression of inv in rovA mutant bacteria was approximately four-fold lower than in wild-type cells (Figure 7D). However, inv expression was fully restored upon inhibition of H-NS by hnsTEPEC, demonstrating that RovA functions solely as a counter-silencer of the inv gene. The rovA and slyA genes are capable of complementing each other in both S. Typhimurium and Y. pseudotuberculosis when expressed in trans (Figure 7E, F), up-regulating both inv and pagC, further indicating that RovA functions as a counter-silencer, like SlyA. Together, these observations demonstrate that RovA has retained the ancestral characteristic of environmentally-responsive repression exhibited by other members of the MarR TF family, despite being one of the most divergent members of the SlyA lineage. However, in contrast to MarR TFs outside of the SlyA lineage, it is also able to function as a counter-silencer of horizontally-acquired genes, as exemplified by inv.

To determine whether RovA functions as a counter-silencer in Y. pseudotuberculosis, transcription of the Rov-regulated inv gene was quantified by qRT-PCR in wildtype or rovA mutant strains expressing hnsTEPEC, which inhibits H-NS (A). Although inv expression is decreased in a rovA mutant strain, expression is fully restored by the inhibition of H-NS, indicating that RovA functions as a counter-silencer. IVT assays of the inv regulatory region in the presence of increasing RovA concentrations detected only a minimal impact on transcription levels, suggesting that RovA is unlikely to function as a classical activator (B). IVT assays of rovA in the presence of increasing RovA concentrations demonstrate that RovA functions as an autorepressor (C). RovA remains sensitive to inhibition by small aromatic carboxylate molecules, as increasing concentrations of sodium salicylate inhibited RovA-mediated rovA repression in IVT assays. Reciprocal complementation studies were performed by providing either rovA or slyA in trans expressed from the arabinose-inducible pBAD18 vector in rovA or slyA mutant Y. pseudotuberculosis and S. Typhimurium strains, respectively, and measuring transcription of inv and pagC via qRT-PCR. Transcript levels are normalized to rpoA in both species, and data represent the mean ± SD; n=3. Asterisks indicate P < 0.05.
SlyA counter-silencing requires high expression levels
It is interesting to note that SlyA does not play a major regulatory role in Escherichia coli, the most well-studied member of the Enterobacteriaceae. SlyA is only known to regulate two E. coli genes, hlyE and fimB (46-48), despite exhibiting a high degree of homology (91% identity) to SlyA in S. Typhimurium. To better understand the different roles of SlyA in the regulatory hierarchy of E. coli and S. Typhimurium, we performed a comparative genetic analysis of the S. Typhimurium (slyASTm) and E. coli (slyAEco) alleles. In allelic exchange experiments, the slyASTm coding sequence was swapped with slyAEco to determine the effect on the expression of the counter-silenced S. Typhimurium pagC gene (Figure 8A). We observed that SlyAEco is able to counter-silence the expression of pagC similarly to SlyASTm, suggesting that the diminished role of SlyA in E. coli is not attributable to differences in protein sequence. This suggested that the importance of different SlyA lineage proteins within their respective regulatory networks may be the result of the different in cis evolutionary pathways identified in our phylogenetic analysis (Figure 6 and Figure 6 - figure supplement 1), resulting in differences in levels of expression. To test this hypothesis, slyA intergenic region-ORF chimeras were constructed and assessed for their ability to counter-silence pagC. To avoid potentially confounding results due to multiple transcriptional start sites (TSSs), we exchanged the intergenic regions beginning immediately upstream of each start codon. Counter-silencing was assessed in a slyA mutant carrying pKM05, a plasmid with the slyASTM ORF transcribed by the E. coli promoter (PslyAEco), or pKM07, a plasmid with the slyASTM ORF transcribed by its native promoter (PslyASTM). Although pagC expression was similar with either construct under non-inducing conditions, expression was approximately 10-fold lower with slyA expressed from PslyAEco under inducing conditions (Figure 8B), suggesting that the diminished role of SlyA in E. coli is at least partially attributable to differences in slyA expression levels in S. Typhimurium and E. coli. Although slyA expression driven by PslyAEco is only slightly lower under non-inducing conditions, it is approximately 3-fold greater when driven by PslyASTM under inducing conditions (Figure 8C), suggesting that PslyAEco is less responsive to environmental signals associated with virulence gene expression in Salmonella, and thus is unable to regulate slyA expression in a manner appropriate for counter-silencing. Parallel experiments in Y. pseudotuberculosis revealed that rovA is also more strongly expressed from its native promoter (ProvAYps) than from a chimera expressing rovA from PslyAEco (Figure 8 - figure supplement 1), despite the fact that ProvAYps has diverged much more significantly from both PslyAEco and PslyASTm than PslyAEco and PslyASTm have diverged from each other (Figure 6 - figure supplement 1).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Expression of the SlyA counter-silenced pagC gene was measured by qRT-PCR in S. Typhimurium strains encoding wildtype slyA, slyAEco, or mutant slyA under either inducing (10 µM MgCl2) or non-inducing (10mM MgCl2) conditions (A). Expression of pagC (B) and slyA (C) was compared in S. Typhimurium strains in which slyA transcription was driven by either the wildtype (WT) or E. coli (PslyAEco) slyA promoter. Transcript levels are normalized to rpoD and data represent the mean ± SD; n=3. Asterisks indicate P ≤ 0.05. (A comparative analysis of a PslyAEco-rovA chimera is shown in Figure 8 - figure supplement 1.)
Discussion
The ancient MarR family of transcription factors is represented throughout the bacterial kingdom and in many archaeal species (36). This study sought to understand how the SlyA/RovA lineage of MarR TFs in the Enterobacteriaceae evolved to acquire the novel function of counter-silencing. Our observations demonstrate that SlyA proteins have retained vestiges of their ancestral functions, e.g., the environmentally-responsive repression of small molecule efflux systems, while acquiring an ability to act as pleiotropic counter-silencers of horizontally-acquired genes. The latter role has been facilitated by the parallel evolution of cis-regulatory elements that support higher levels of gene expression.
SlyA, like other MarR family TFs, is subject to allosteric inhibition by small aromatic carboxylate compounds such as salicylate, which bind and stabilize the SlyA dimer in a conformation unfavorable for DNA binding (Figure 2 and Figure 2 - figure supplement 1). However, the specific structure and arrangement of the effector binding sites varies significantly among the MarR family. Our structural data indicate that SlyA binds a total of six salicylate molecules per dimer, although two of these are not likely to be biologically relevant (Figure 3). Although the general structure and architecture of the salicylate-SlyA complex is similar to complexes formed by other MarR TFs, including MarR (8) and MTH313 from Methanobacterium thermautrophicum (49), the various TFs differ significantly in their specific interactions with salicylate. MarR binds a total of four salicylate molecules per dimer, whereas MTH313 binds only two (Figure 3D, E). All four salicylate-binding sites in MarR flank the wHTH domain and are partially exposed to solvent, whereas none of these sites corresponds to the salicylate binding sites of SlyA, potentially accounting for the significant differences in affinity for aromatic carboxylates between the two proteins. MTH313 binds salicylate asymmetrically at sites similar to sites I and II of SlyA, binding only one salicylate molecule per monomer. It is possible that the structural variability in the MarR family with respect to salicylate binding reflects an inherent evolutionary flexibility and ligand promiscuity. The MarR family sensor region may have evolved to interact with a variety of small molecules, and salicylate may simply represent a promiscuous probe for potential interactions. Recent studies have suggested that the true ligand of MarR may be copper, liberated from membrane-associated proteins during oxidative stress induced by xenobiotic agents. Copper reportedly oxidizes a cysteine residue (C80) to promote disulfide bond formation between MarR dimers, resulting in subsequent tetramerization and inhibition of DNA binding (15). As almost all characterized MarR TFs have a reactive cysteine residue corresponding to a region near Site II of SlyA, we considered that cysteine oxidation, which has also been described in the MarR family TF OhrR (50), might occur in the SlyA lineage as well. However, mutation of C81, the lone cysteine residue in SlyASTm, had a negligible effect on SlyA activity in both the presence and absence of salicylate. The addition of copper to the growth medium had a minimal impact on SlyA activity and was unaffected by mutation of C81, indicating that cysteine oxidation is not a general mechanism for the allosteric inhibition of MarR family TFs. Furthermore, the high affinity of SlyA for aromatic carboxylates (Figure 2 - figure supplement 2) and the conservation of binding pocket residues (Table S3) reported in this study suggest that environmental sensitivity and allosteric inhibition are a conserved feature of SlyA activity in their new regulatory role.
A phylogenetic analysis of SlyA orthologs in Enterobacteriaceae demonstrates that the SlyA lineage is strongly conserved, even in endosymbiotic species exhibiting significant genome loss (Figure 5 and Figure 5 - figure supplement 1). This suggests that SlyA has a central and essential role in the transcriptional regulatory networks of these species. For example, a recent analysis of the endosymbiont W. glossinidia found that slyA is subject to significant evolutionary constraints (51). This conservation does not appear to be due to its role as a regulator of antimicrobial resistance, as slyA does not exhibit significant linkage to the efflux pump operon ydhIJK, outside of the enteric pathogens in cluster I (Figure 6). Rather, SlyA proteins appear to function predominantly as pleiotropic counter-silencers, facilitating the integration of horizontally-acquired genes, including virulence genes, into existing regulatory networks. A general counter-silencing role has been suggested in multiple enterobacterial species including E. coli (19, 48), Salmonella (17, 20, 21) and Shigella (32) in cluster I, and Yersinia (31, 52), Serratia, and Pectobacterium (Erwinia) (25) in cluster II. Although similar evidence does not currently exist for the endosymbionts of cluster II, this may simply reflect the limited genetic analyses that have been performed in these species.
To understand the evolutionary adaptations to accommodate counter-silencing by the SlyA/RovA lineage, we compared three orthologs, slyASTM, slyAEco and rovA. The slyASTM and rovA genes are essential for Salmonella and Yersinia virulence, whereas slyAEco plays a negligible role in E. coli, despite exhibiting a much higher degree of similarity to slyASTM than does rovA. These differing roles are attributable to differences in expression, as SlyAEco is able to function as a counter-silencer in S. Typhimurium when its expression is driven by the S. Typhimurium promoter (Fig. 6). We also observed that slyAEco transcription is significantly reduced in medium with low Mg2+ concentrations, a condition associated with the Salmonella-containing vacuole (SCV) of macrophages, in which SlyA cooperates with the response regulator PhoP to counter-silence virulence genes necessary for intracellular survival (17). This suggests that the diminished role of slyAEco in E. coli may result from its inability to respond to appropriate environmental cues. This is further reinforced by the observation that RovA of Yersinia spp. is able to function as a counter-silencer in S. Typhimurium when expressed in trans from an inducible promoter, and that the rovA promoter of Y. pseudotuberculosis drives rovA transcription more strongly than PslyAEco in a chimeric strain (Fig. S7). This suggests that evolution of the SlyA-RovA lineage in trans, particularly DNA binding specificity, has played only a minor role since the divergence of the Enterobacteriaceae. This is further supported by as comparison of distantly-related SlyA orthologs, which exhibit most sequence divergence in the C-terminal oligomerization domain and not the N-terminal region containing the wHTH (Figure 5 - figure supplement 1). We conclude that the ability of a given SlyA ortholog to serve as a counter-silencer is contingent on its level and pattern of expression, which may be the product of both transcriptional and post-transcriptional activity, as mutations altering TSS position will subsequently alter the 5’ untranslated region of the slyA transcript. Notably, another group recently demonstrated a significant expansion of the SlyA regulon in E. coli when slyAEco is overexpressed, with 30 operons exhibiting regulation by SlyA, 24 of which are also repressed by H-NS (34), indicating that SlyAEco is capable of functioning as a counter-silencer but is not expressed under the appropriate conditions. In S. Typhimurium, the appropriate conditions are those associated with the SCV. However, SlyA orthologs in other species such as the plant pathogens and endosymbionts of cluster II are likely to require expression under vastly different conditions corresponding to their environmental niches. The loss of the divergently-transcribed ydhIJK operon in Y. pseudotuberculosis and other enteric species may be a consequence of genetic alterations to enhance rovA expression as well as the redundancy of drug efflux pumps. An additional possibility is that enhanced slyA/rovA expression might result in hyper-repression of ydhIJK, which would negate the usefulness of the pump to the cell. Even the plant pathogens such as D. dadantii and P. carotovorum, which are most likely to encounter small phenolic compounds in the plant environment (53), have failed to retain ydhIJK. It is also notable that the slyA orthologs in the four species (E. coli, S. Typhimurium, Y. enterocolitica, Y. pseudotuberculosis) in which the transcriptional start sites have been characterized initiate transcription at different positions (30, 54-57), indicating that each species has evolved its cis-regulatory circuit independently (Figure 6).
Previous studies have demonstrated that regulatory evolution can promote adaptation to new niches (58). The SlyA/RovA TF lineage provides a unique example of parallel regulatory evolution to achieve a common functional objective. Throughout the Enterobacteriaceae, the associated ydhIJK pump genes have been repeatedly lost, yet their regulators have been retained, presumably to facilitate the evolution of an appropriately responsive regulatory circuit to enable counter-silencing. This suggests that SlyA/RovA proteins possess intrinsic features that predispose them for a counter-silencing role, perhaps the abilities to respond to environmental stimuli and to recognize a variety of AT-rich target DNA sequences. Studies are underway to characterize these features and to determine their contribution to the evolutionary capacity of Enterobacteriaceae.
Materials and methods
Bacterial strains and general reagents
All oligonucleotides and plasmids used in this study are described in Table S4 and Table S5, respectively. Unless otherwise indicated, bacteria were grown in Luria Bertani (LB) broth with agitation. Salmonella enterica serovar Typhimurium strains were constructed in the ATCC 14028s background and grown at 37°C, unless otherwise indicated. The 14028s slyA mutant strain was described previously (17). Yersinia pseudotuberculosis YPIII (59) and YP107 (60) (a gift from P. Dersch, Helmholtz Centre for Infection Research) were used as the wildtype and rovA strains, respectively, and grown at 24°C. A ydhIJK deletion mutant was constructed using the λ-Red recombinase system (61) and oligonucleotides WNp318 and WNp319. A slyA ydhIJK strain was generated by introducing the slyA::Cm cassette from 14028s slyA::Cm (17) to 14028s ydhIJK via P22HTint-mediated transduction. To exchange the wild-type S. Typhimurium slyA coding sequence with the E. coli allele, S. Typhimurium slyA was replaced with a thyA cassette, via FRUIT (62), using the oligonucleotides STM-slyA-targ-F and STM-slyA-targ-R. The slyA coding sequence from E. coli K-12 was then amplified using the primers STM-Eco-slyA-F and STM-Eco-slyA-R, which include 40 and 41 bases, respectively, from the regions flanking the S. Typhimurium slyA coding sequence. This fragment was electroporated into the slyA::thyA mutant strain and the resulting transformants were plated on minimal media containing trimethoprim, as described for the FRUIT method (62), to select for replacement with the E. coli slyA allele.
The chromosomal C81S slyA mutation (TGC→AGC) was generated by cloning a 1 kb fragment encoding the C81S mutation into the suicide vector pRDH10 (63) using Gibson Assembly (New England Biolabs, Ipswich MA). The Gibson assembly reaction included PCR products generated with primers JKP736/JKP737 and JKP738/JKP739 and genomic S. Typhimurium DNA as well as BamHI-digested pRDH10 to generate pJK723. For integration of slyA C81S into the chromosome, S17-1Δλpir (64)/pJK723 was mated with S. Typhimurium 14028s/pSW172 (65) and plated onto LB+20 µg ml-1 chloramphenicol and incubated at 30°C overnight. Chloramphenicol- and carbenicillin-resistant (Cmr Carbr) colonies that were isolated represented a single crossover event of pJK723 (Cmr) plasmid into the slyA region of S. Typhimurium 14028s (Carbr). Selection for the second crossover event to replace wild-type slyA with a slyA C81S mutation was performed by plating 0.1ml of an overnight culture of Cmr Carbr colony onto LB+5% sucrose plates and incubating at 30°C overnight. Colonies were then streaked onto LB plates at 37°C and putative 14028s slyA C81S colonies confirmed by DNA sequencing.
Cloning
pSL2143 was generated by amplifying the slyASTm region, including its native promoter, with primers slyAcomp-F and slyAcomp-R, and ligating into pWSK29 (66) digested with EcoRV. The G6A, S7A, A10P, R14A, W34A, H38A, T66A and C81A mutants were generated with their respective mutagenic primer pairs (Table S4) using the QuikChange XL Site-Directed Mutagenesis Kit (Agilent Technologies, Santa Clara CA) according to the manufacturer’s protocol. A 2249 bp region containing both slyA and ydhI was amplified by PCR using the primers SlyAreg-F and SlyAreg-R. The resulting fragment was digested with BamHI and HindIII and ligated into the low-copy IVT scaffold vector pRW20 (17) to generate pRW39. An IVT target containing the 2379 bp rovA region from Y. pseudotuberculosis was generated using the primers BamHI-rovA-F and EcoRI-rovA-R. The resulting fragment and pRW20 were both digested with BamHI and EcoRI and ligated together to construct pRW54. An IVT target containing the 2708 bp inv region from Y. pseudotuberculosis was generated via PCR using the primers BamHI-inv-F and EcoRI-inv-R. The resulting fragment and pRW20 were both digested with BamHI and EcoRI and ligated together to construct pRW55. To perform rovA and slyA complementation studies, both genes were cloned into the arabinose-inducible expression vector pBAD18 (67). The rovA gene was amplified using the oligonucleotides EcoRI-rovA-F and KpnI-rovA-R. The slyA gene was amplified using the oligonucleotides EcoRI-slyA-F and KpnI-slyA-R. Both fragments were digested with EcoRI and KpnI, and ligated into pBAD18, generating pRW58 (slyA) and pRW59 (rovA). pRW60 was constructed by cloning an N-terminal 6×His-tagged copy of rovA into pTRC99a (68). The rovA gene was amplified from YPIII genomic DNA by PCR using the oligonucleotides 6HisRovA-F and 6HisRovA-R. The resulting fragment and pTRC99a were digested with NcoI and BamHI, and ligated together. An E. coli PslyA-S. Typhimurium slyA coding sequence chimera was constructed using overlapping PCR. The slyA-ydhIJK intergenic region was amplified from E. coli K12 genomic DNA using primers KMp178 and KMp206, and from S. Typhimurium 14028s using oligonucleotides KMp177 and KMp206. The corresponding slyA ORF from S. Typhimurium was amplified with primers KMp207 and KMp181. The ORF and promoter segments were amplified along with 40bp of complementary overlapping sequence. Products from the ORF and promoter reactions were mixed 1:1 and amplified using primers KMp178 (E. coli) or KMp177 (S. Typhimurium) and KMp181 to amplify the promoter/ORF chimeras. The resulting fragment and pTH19Kr (69) were digested with BamHI and HindIII and ligated together, generating pKM05 (E. coli promoter) and pKM07 (S. Typhimurium promoter). Plasmids were transformed into S. Typhimurium 14028s slyA for gene expression analysis. The hnsT coding sequence was amplified from E2348/69 genomic DNA using the oligonucleotides EcoRI-hnsT-F and HindIII-hnsT-R. Both pBAD18 and the resulting PCR fragment were digested with EcoRI and HindIII, agarose gel-purified, and ligated together to generate pRW57.
Fusaric acid resistance assays
Cultures were grown overnight in LB broth at 37°C, then diluted 1:100. Thirty microliters of this dilution were added to 270 µl of LB containing 30 µg/ml freshly prepared fusaric acid in 100-well BioScreen plates (GrowthCurves, Piscataway, NJ). Cultures were grown with continuous maximum shaking at 37°C and regular OD600 measurements were taken on a BioScreen C MBR. Fresh 20 mg/ml stock solutions of fusaric acid were prepared in DMF (dimethyl formamide).
NMR analysis of SlyA-salicylate
SlyA protein for NMR analysis was prepared as an N-terminal 6×His-tagged protein from cells grown in M9 minimal medium supplemented with 15N-ammonium chloride (Cambridge Isotope Labs, Tewksbury MA). SlyA protein expression was induced by the addition of 2 mM IPTG (isopropyl-ß-D-thiogalactopyranoside) and the protein purified to homogeneity using Ni-affinity chromatography as previously described (21), except that samples were dialyzed in 50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 0.1mM EDTA, and 3 mM DTT following purification. Protein and ligand solutions for NMR experiments were prepared in 25 mM sodium phosphate, 150 mM NaCl buffer at pH 7.0 containing 10% D2O. NMR spectra were collected on a Bruker DMX 500hHz spectrometer (Bruker, Billerica MA) on samples equilibrated at 35°C and consisting of 250-350 µM 15N-labeled His-SlyA in the absence or presence of 2mM sodium salicylate or 0.5mM sodium benzoate. Spectra were processed using NMR-Pipe (70) and analyzed using NMR-View (71).
SlyA-salicylate crystallization
SlyA was over-expressed and purified for crystallization as previously described (21), except that samples were dialyzed in 50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 0.1 mM EDTA, and 3 mM DTT following purification. Cryo I and II sparse matrix crystallization screens (Emerald Biosystems, Bainbridge Island, WA) were used to determine initial conditions for His-SlyA crystal formation by sitting-drop vapor diffusion in 24-well crystal trays. Equal volumes (4µL) of SlyA and crystallization solutions were mixed before plates were sealed and kept at room temperature. Crystals appeared within 4-7 days in 20% PEG 300, 10% glycerol, 0.1 M phosphate/citrate buffer, pH 4.2, 0.2M ammonium sulfate (condition #14). Subsequent crystal growth was performed using lab-made 20% PEG 400, 10% glycerol, 0.1 M phosphate/citrate buffer, pH 4.2, and 0.2 M ammonium sulfate. Crystallization of SlyA with sodium salicylate was achieved by adding dilutions of a 2M sodium salicylate aqueous stock solution to the protein-crystallization solution mixture. SlyA-salicylate crystals appeared within 7-10 days. A single crystal appeared at a concentration of 75 mM sodium salicylate and grew to a maximum size of 500µ × 300µ × 150µ (Fig. S4). This crystal was used for X-ray diffraction experiments.
The crystal was frozen at 100°K in its crystallization solution for diffraction data collection on GM/CA-CAT beamline 23-ID-D at the Advanced Photon Source. The space group for the crystals is P21212 with two SlyA molecules in the asymmetric unit. The diffraction data were processed with HKL2000 (72). Dataset statistics are shown in Table S1. The crystal structure of the salicylate complex of SlyA was solved using a model of a previously investigated structure of apo-SlyA (unpublished data). That apo-structure was solved using the molecular replacement program, MOLREP (73) with a search model generated by applying Swiss-Model (74) and the SlyA amino acid sequence to PDB entry 2FBH (75). The structural model for the salicylate complex was refined using REFMAC-5 (76) in the CCP4 suite (77). Rfree (78) was calculated using 5% of the data in the test set. A high-resolution limit of 2.3Å was applied for the refinement, consistent with standards appropriate when the structure was solved. This is the resolution at which Rmerge for the data set drops below 0.40. XtalView (79) and Coot (80) were used to examine sigma A weighted |Fo|-|Fc| and 2|Fo|-|Fc| electron density maps (81). MOLSCRIPT(82), and Raster3D (83) were used to produce structural figures for this paper. Table S2 contains refinement statistics for the structure. Coordinates and structure factors have been deposited in the Protein Data Bank with identifier 3DEU.
6×His-RovA purification
RovA was purified using the same protocol as described previously for SlyA (17). However, overexpression cultures were grown at 24°C to OD600=0.5. IPTG was added to a final concentration of 1mM, and cultures were incubated at 24°C for an additional four h before the cells were harvested by centrifugation and cell pellets stored at −80°C for the subsequent purification of RovA.
In vitro transcription
IVT assays were performed essentially as previously described (17) with the following modifications. All SlyA IVT assays were performed at 37°C. All RovA IVT assays were performed at 24°C. Where indicated, sodium salicylate was added to IVT reactions prior to the addition of SlyA or RovA. All oligonucleotides and templates used in IVT reactions are indicated in Table S6.
RNA isolation and qRT-PCR
RNA was purified using Trizol (Life Technologies, Carlsbad CA) according to the manufacturer’s protocols. cDNA was generated using the QuantiTect Reverse Transcription Kit (Qiagen, Hilden, Germany), and quantified in a BioRad CFX96 (BioRad, Hercules CA), using SYBR Green Master Mix (84).
For the analysis of pagC expression under inducing conditions, S. Typhimurium cultures were grown to early stationary phase (OD600≈2.0) at 37°C in LB broth, then washed three times in N-minimal medium containing either 10 µM (inducing) or 10 mM MgCl2 (non-inducing). Cultures were re-suspended in the appropriate N-minimal medium with 2mM sodium salicylate where indicated and incubated for an additional 30 min at 37°C before cells were harvested for RNA purification studies. For the complementation of slyA with slyA or rovA in trans, overnight cultures were diluted to 0.05 OD600 and grown for two h at 24°C before adding arabinose to a final concentration of 0.02% w/v. Cultures were grown for an additional six h before harvesting RNA.
Y. pseudotuberculosis strains YPIII and YP107 were diluted from overnight cultures to 0.05 OD600 in LB and grown with shaking at 24°C. For the inhibition of H-NS via H-NSTEPEC over-expression, and complementation of rovA with rovA or slyA in trans, YPIII and YP107 cultures carrying either pBAD18 or pRW57 were grown for two hrs before arabinose was added to a final concentration of 0.02% w/v. Cultures were grown for an additional six hrs before harvesting RNA. For slyA/rovA complementation studies, rpoAYS primers targeting rpoA were used as loading controls, as rpoA is sufficiently conserved between the two species as to allow use of the same primers.
Acknowledgments
The National Institutes of Health provided support to FCF (AI39557, AI44486, AI118962, AI112640) and SJL (A148622). GM/CA@APS has been funded in whole or in part with Federal funds from the National Cancer Institute (ACB-12002) and the National Institute of General Medical Sciences (AGM-12006). This research used resources of the Advanced Photon Source, a U.S. Department of Energy (DOE) Office of Science User Facility operated for the DOE Office of Science by Argonne National Laboratory under Contract No. DE-AC02-06CH11357. We thank Eric Larson for helpful discussions and suggestions. We also thank Catherine Eakin and Ponni Rajagopal for assistance with protein purification and crystal growth, Petra Dersch for kindly providing Y. pseudotuberculosis YPIII and YP107, and Ralph Isberg for providing Y. pseudotuberculosis IP32953. Finally, we thank Dr. Rachel Klevit for use of her NMR facilities to collect spectra of SlyA-salicylate complexes.
Footnotes
Signifcant additional data (Figure 2 - figure supplement 2, Figures 4-6) and discussion. Authors updated.
References
- 1.↵
- 2.↵
- 3.↵
- 4.
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵
- 81.↵
- 82.↵
- 83.↵
- 84.↵
- 85.↵
- 86.↵




