

Abstract
Hundreds of long non-coding RNAs (lncRNAs) have been identified as potential regulators of gene expression, but their functions remain largely unknown. To study the role of lncRNAs during vertebrate development, we selected 25 zebrafish lncRNAs based on their conservation, expression profile or proximity to developmental regulators, and used CRISPR-Cas9 to generate 32 deletion alleles. We observed altered transcription of neighboring genes in some mutants, but none of the lncRNAs were required for embryogenesis, viability or fertility. Even RNAs with previously proposed non-coding functions (cyrano and squint) and other conserved lncRNAs (gas5 and lnc-setd1ba) were dispensable. In one case (lnc-phox2bb), absence of putative DNA regulatory-elements, but not of the lncRNA transcript itself, resulted in abnormal development. LncRNAs might have redundant, subtle, or context-dependent roles, but extrapolation from our results suggests that the majority of individual zebrafish lncRNAs are dispensable for embryogenesis, viability and fertility.
Long non-coding RNAs (lncRNAs) comprise a heterogeneous group of transcripts longer than 200 nucleotides that do not encode proteins. LncRNAs have been proposed to affect the expression of neighboring or distant genes by acting as signaling, guiding, sequestering or scaffolding molecules1–5. The functions of specific lcnRNAs in dosage compensation (xist6,7, tsix8, jpx9) and imprinting (Airn10,11, MEG312,13, H1914,15) are well established, and mutant studies in mouse have suggested that fendrr, peril, mdget, linc-brn1b, linc-pint16, and upperhand17 are essential for normal development. However, other studies have questioned the developmental relevance of several mouse lncRNAs, including Hotair18, MIAT/Gumafu19, Evx1-as20, upperhand, braveheart and haunt21. In zebrafish, morpholinos targeting the evolutionarily-conserved lncRNAs megamind (TUNA22) and cyrano resulted in embryonic defects23. However, a mutant study found no function for megamind and revealed that a megamind morpholino induced non-specific defects24. These conflicting results have led to a controversy about the importance of lncRNAs for vertebrate development16,21. We therefore decided to mutate a group of selected zebrafish lncRNAs using CRISPR-Cas9, and assay their roles in embryogenesis, viability and fertility.
Transcriptomic studies of early embryonic development23,25 and five adult tissues26 have identified over 2,000 lncRNAs in zebrafish27, of which 727 have been confirmed as non-coding based on ribosome occupancy patterns28. For our knockout study we selected 24 bona fide lncRNAs based on conservation, expression dynamics and proximity to developmental regulatory genes (see Fig 1 for selection criteria for each lncRNA). These criteria were chosen to increase the likelihood of functional requirement. In addition, we selected a protein-coding RNA with a proposed non-coding function (squint). The genomic location, neighbor-relationship, and expression levels of the selected lncRNAs and their neighboring genes are shown in supplementary figures Fig S1, Fig S2, and Fig S3, respectively.

lncRNA names are shown in the first column. lncRNAs were named using the last 4 digits of their corresponding ENSEMBL Transcript ID or their chromosome number if no transcript ID was available (e.g. lnc-1200 is located on chromosome 12). The second column represents ribosomal occupancy pattern along the length of lncRNAs in comparison to the 5’UTR, coding and 3’UTR of typical protein-coding transcripts28. The third column shows the transcript ID for the investigated lncRNA or its genomic coordinate in GRCz10. Column Four represents the expression dynamics of the lncRNAs (see also Figure S3) (log2 (FPKM +1) between 0 and 8)25. The fifth column shows the deletion size. Sixth column represent the percentage decrease in the level of lncRNA in comparison to wild type from 3 biological replicates (qRT-PCR). The seven and eight columns show the presence of embryonic phenotypes, viability and fertility (at least 15 adult pairs per allele) of homozygous mutant fish. Ninth column shows the upstream and downstream neighboring genes in a 200kb window centered around the lncRNA’s TSS. The last column provides the selection criteria for each lncRNA.
Using CRISPR-Cas9 (Fig S4) we generated 32 knockout-alleles. 24 alleles removed regions containing transcription start sites (TSS-deletion; 244bp to 736bp), and 8 alleles fully or partially removed the gene (1kb to 203kb) (Fig 1). qRT-PCR analysis demonstrated effective reduction in the levels of the targeted lncRNA transcripts (average reduction of 94 ± 6%; Fig 1), which was further tested and confirmed for a subset of lncRNAs by in situ RNA hybridization (Fig 2B, 3B, 4C, 4D, 5B and 6D).
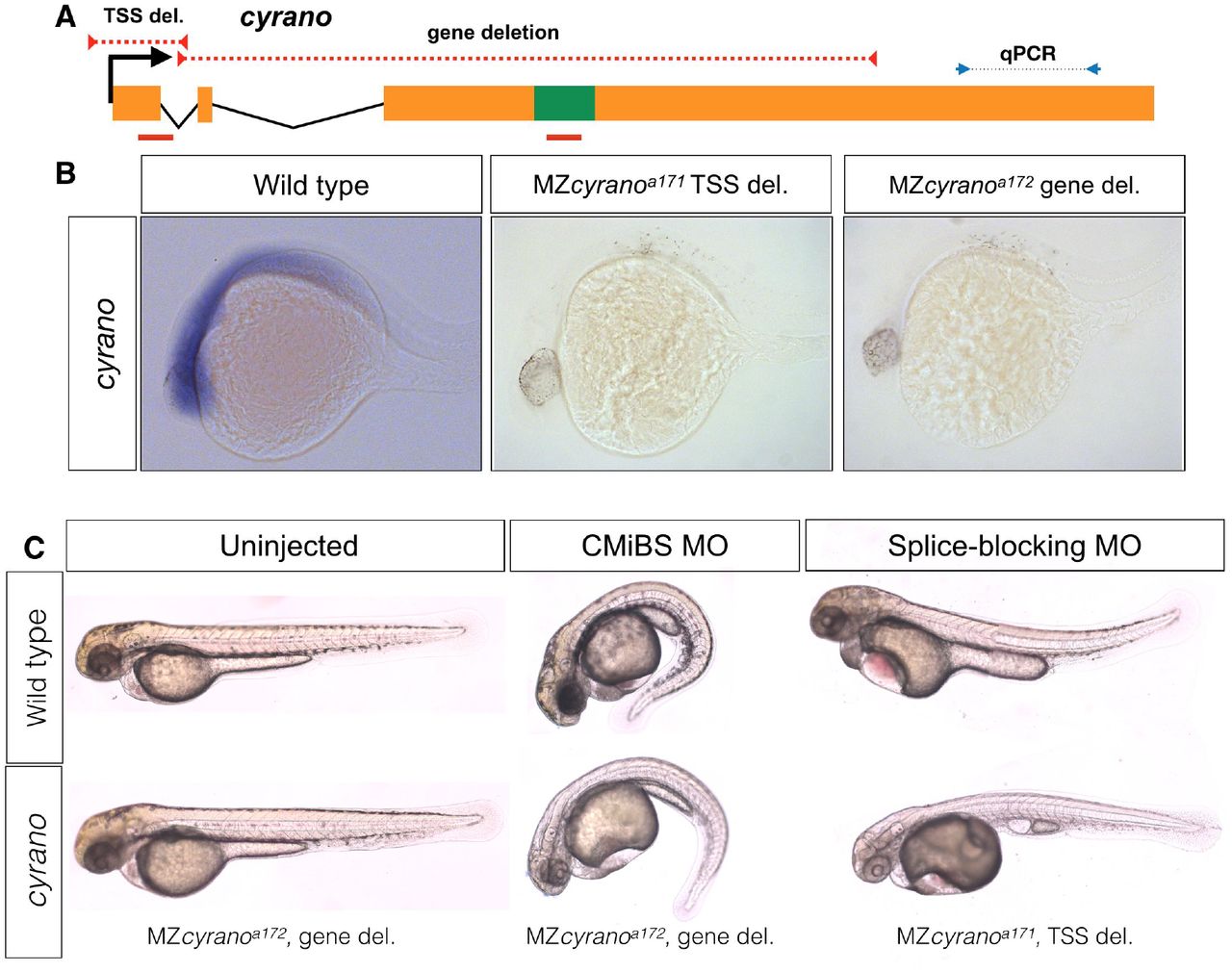
A) The positions of TSS-deletion allele and gene deletion allele are marked by dashed red lines. Green box represents the conserved element in cyrano which is complementary to miR-7. Solid red lines indicate the position of the first exon-intron boundary (e1i1) morpholino and conserved microRNA binding site (CMiBS) morpholinos. Arrows flanking black dotted line mark the primer binging sites for qRT-PCR product. B) Representative images of in situ hybridization for cyrano in wild type (15/15) and both homozygous TSS-deletion (21/22) and gene deletion (18/18). C) At 1-dpf gene deletion mutants (lower-left), (and TSS-deletion mutants, not shown) were not different from the wild type embryos (upper-left). Morpholino injected wild-type embryos (upper-middle and upper-left) reproduced observed phenotype in Ulitsky et. al.24. Morpholino injected deletion-mutants, lacking the corresponding binding sites for morpholinos, (lower-middle and lower-left) were comparable to morpholino injected wild types.
Previous observations in mammalian cell culture systems suggested that lncRNA promoters can affect the expression of nearby genes29. To test if these results hold true in vivo, we measured the changes in the expression of neighboring genes (a 200 kb window centered on each lncRNA) in lncRNA mutants. Several mutants displayed changes in the expression of neighboring genes (Fig S5). In particular, 10 out of 40 neighboring genes showed more than two-fold changes in expression, lending in vivo support to observations in cell culture systems29.
To determine the developmental roles of our selected lncRNAs, we generated maternal-zygotic mutant embryos (lacking both maternal and zygotic lncRNA activity) and analyzed morphology from gastrulation to larval stages, when all major organs have formed. Previous large-scale screens30,31 have shown that the visual assessment of live embryos and larvae is a powerful and efficient approach to identify mutant phenotypes, ranging from gastrulation movements and axis formation to the formation of brain, spinal cord, floor plate, notochord, somites, eyes, ears, heart, blood, pigmentation, vessels, kidney, pharyngeal arches, head skeleton, liver, and gut. No notable abnormalities were detected in 31/32 mutants. Moreover, these 31 mutants survived to adulthood, indicating functional organ physiology, and were fertile (Fig1). In the following section we describe the results for five specific lncRNAs and put them in the context of previous studies.
Cyrano
cyrano is evolutionarily conserved lncRNA and based on morpholino studies, has been suggested to have essential functions during zebrafish embryogenesis23 and brain morphogenesis 32. cyrano has also been suggested to act as a sponge (decoy-factor) for HuR during neuronal proliferation33, regulate miR-7 mediated embryonic stem cell differentiation34, and control the level of miR-7 in the adult mouse brain35. We generated two mutant alleles that removed the TSS (cyranoa171) or the gene (cyranoa172), including the highly conserved miR-7 binding-site (Fig 2A, B). The expression level of the nearby gene (oip5) was not affected in either of these mutants (Fig S5). In contrast to previous morpholino studies in zebrafish23 but in support of recent findings in mouse35, cyrano mutants developed normally and were viable and fertile.
The difference between morphant23 and mutant phenotypes might be caused by compensation in the mutants36,37. To test this possibility, we injected the previously used morpholinos targeting the first exon-intron boundary (e1i1) or the conserved miR-7 binding site (CMiBS) into wild type and homozygous deletion mutants. The TSS-mutant allele lacked the e1i1 morpholino binding site and the gene deletion allele lacked the CMiBS morpholino binding site (Fig 2A). The previously reported phenotypes, including small heads and eyes, heart edema, and kinked tails were found in both wild type and mutants (Fig 2C), demonstrating that the morpholino-induced phenotypes were non-specific. These results reveal that cyrano transcripts or their evolutionarily conserved miR-7 binding site, are not required for embryogenesis, viability or fertility.
gas5
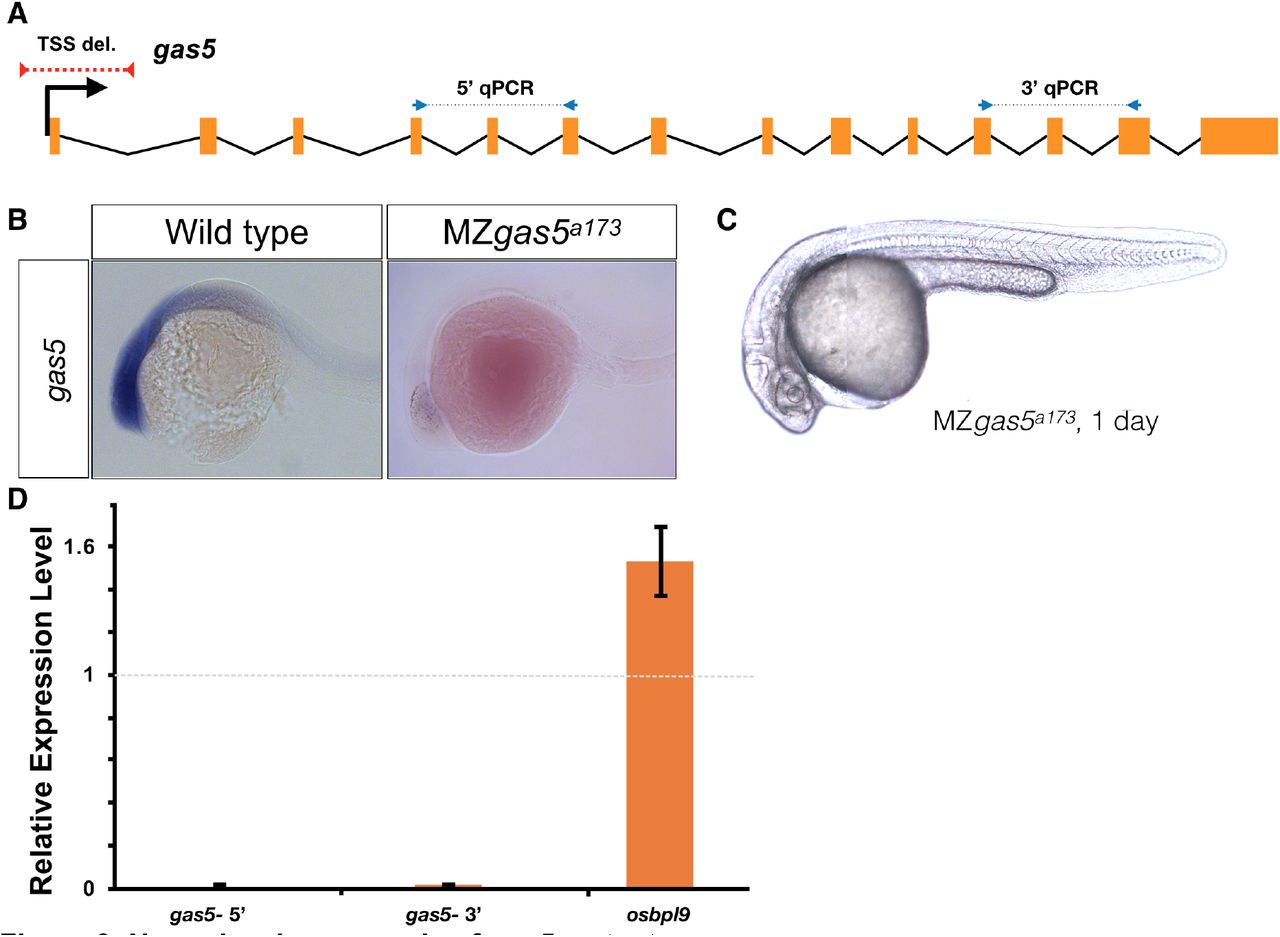
gas5 is an evolutionarily conserved lncRNA (growth-arrest specific 5)38 that is highly expressed in early development (Fig 3B) and hosts several snoRNAs implicated in zebrafish development39. Knockdown and knockout studies in cell culture40 have indicated that gas5 might act as a tumor suppressor41 and exert effects at distant genomic sites42. However, the role of this lncRNA in development has not been studied in any vertebrate. Our gas5a173 mutant allele removed the sequences containing the TSS (−169 to +127) (Fig 3A) and resulted in complete elimination of its expression (Fig 3B, 3D). Expression of the neighboring gene osbpl9, encoding a lipid binding protein, was increased by 50% (Fig 3D), but gas5a173 mutants were indistinguishable from wild type (Fig 3C), reached adulthood and were fertile.

A) Position of the TSS-deletion allele in gas5 is marked by dashed red line. Arrows flanking black dotted lines mark the primer binging sites for 5’-qPCR and 3’-qPCR products. B) Representative in situ hybridization images for gas5 in wild type (11/11) and homozygous TSS-deletion mutants (11/11). C) Maternal and Zygotic gas5 (MZgas5) mutant embryos at 1-dpf were indistinguishable from the wild-type embryos at the same developmental stage (not shown). D) Expression level of gas5 and osbpl9 measured by qRT-PCR. Tor3A, the other neighboring gene, was not expressed at the investigated time-point.
Lnc-setd1ba
Lnc-setd1ba is the zebrafish orthologue of human LIMT 43 (LncRNA Inhibiting Metastasis), which has been implicated in basal-like breast cancers. It is expressed from a shared promoter region that also drives the expression of the histone methyltransferase setd1ba in opposite direction (Fig 4A). Evolutionary conservation in vertebrates and proximity to setd1ba, whose mouse homolog is essential for embryonic development44,45 prompted us to investigate the function of this lncRNA in zebrafish. We removed the gene of lnc-setd1ba downstream of its TSS (3137bp deletion) (lnc-setd1baa174). In situ hybridization and qRT-PCR revealed absence of lncRNA expression (Fig 4C and 4E) and strong upregulation of setd1ba (Fig 4D and 4E) during cleavage stages and slight upregulation of setd1ba and the other neighboring gene rhoF at one-day post fertilization (1-dpf) (Fig 4E). Despite these changes, maternal-zygotic lnc-setd1baa174 mutants were indistinguishable from wild type (Fig 4B), reached adulthood and produced normal progeny.

A) The relative position of lnc-setd1ba and the protein-coding gene setd1ba. The gene deletion region is marked by dashed red line. Arrows flanking black dotted line mark the primer binging sites for qRT-PCR product. B) Maternal and zygotic lnc-setd1ba mutants were not different from wild-type embryos at 1-dpf. C) Representative images of in situ hybridization for lnc-setd1ba at 4-8 cell stage mutant (18/18) and wild-type (25/25) embryos. D) In situ hybridization for the protein-coding mRNA, setd1ba (9/11) in lnc-setd1ba mutants compared to the wild-type embryos (15/15). E) qRT-PCR at 1-cell stage and 1-dpf for the lncRNA and its neighboring genes rhoF and setd1ba.
Squint
Squint encodes a Nodal ligand involved in mesendoderm specification46,47. The previously studied squint insertion mutant alleles (squintHi975Tg 46 and squintcz35 47) lead to delayed mesendoderm specification and partially penetrant cyclopia48. Morpholino and misexpression studies have suggested an additional, non-coding role for maternally provided squint, wherein the squint 3′UTR mediates dorsal localization of squint mRNA, induces the expression of dorsal mesoderm genes, and is required for the development of dorsal structures49,50. This mode of activity assigns squint to the cncRNA family - RNAs with both protein-coding and non-coding roles51. To investigate the non-coding roles of squint mRNA we generated a deletion allele (squint a175) that lacked most of the protein coding region and the 3’UTR, including the Dorsal Localization Element (DLE) implicated in maternal squint RNA localization52 (Fig 5A). In situ hybridization (Fig 5B) and qRT-PCR (Fig 5C) showed that the level of remaining squint transcript was greatly reduced (~90%). MZsquint a175 embryos displayed partially penetrant cyclopia, similar to existing protein-disrupting squint alleles (Fig 5D)46,47,53, but the defects proposed to be caused by interference with squint non-coding activity49 were not detected.
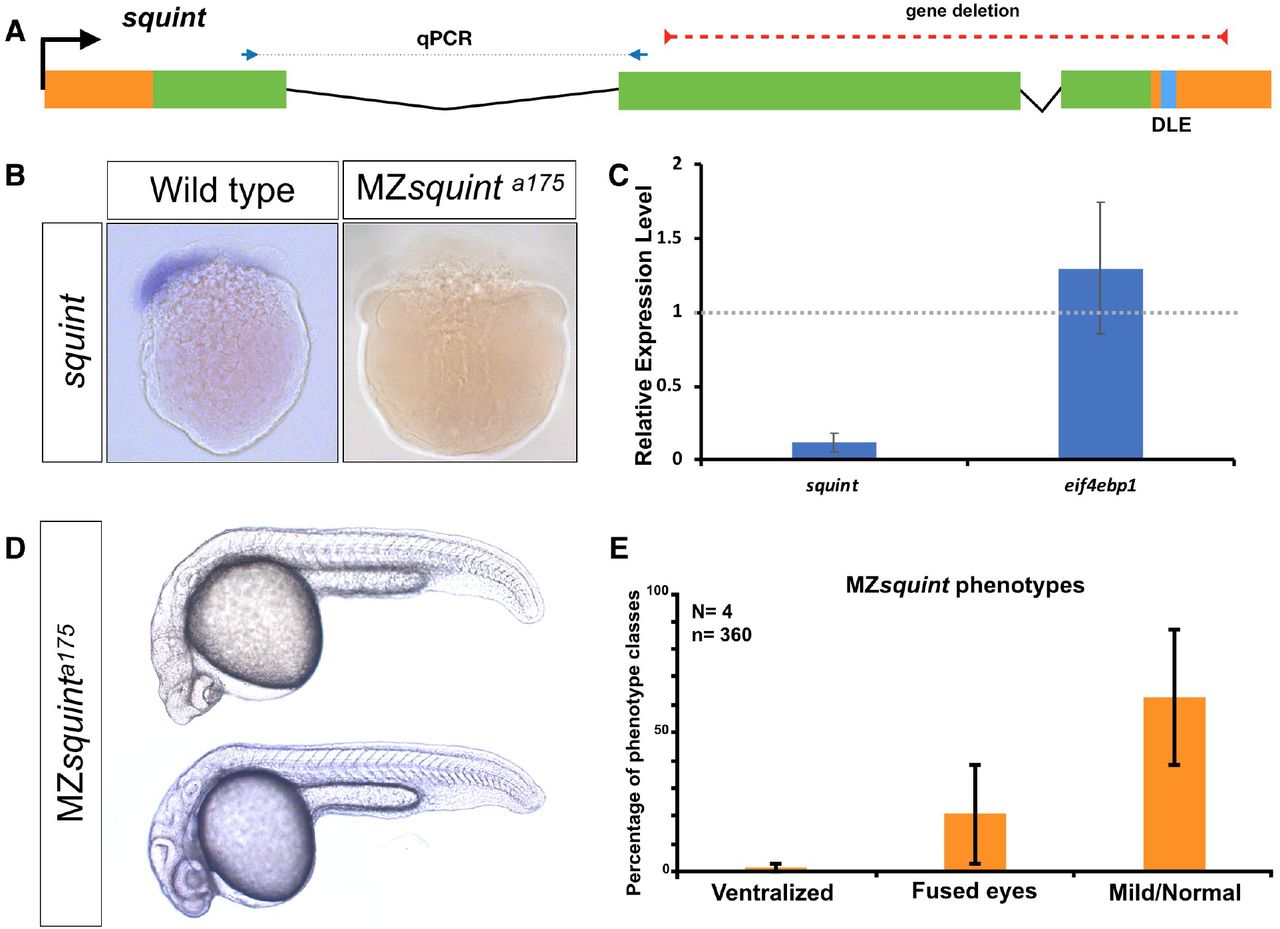
A) The position of untranslated regions (brown), coding region (green), putative Dorsal Localization Element-DLE (blue) and the gene deletion (red dashed line) in the squint genomic locus. Arrows flanking black dotted line mark the primer binging sites for qRT-PCR product. B) In situ hybridization for squint at 8-cell stage on wild-type (18/20) and MZsquinta175(17/17) embryos. C) MZsquinta175 embryonic phenotype (N=4 independent crosses, n=360 embryos). D) Two representative MZsquinta175 embryos. E) qRT-PCR for squint and eif4ebp1 on wild-type and MZsquinta175 embryos.
To further test whether squint mRNA might have non-coding roles, we injected wild-type and MZsquint a175 embryos with either control RNA, full-length squint mRNA, a non-coding version of squint mRNA, or the putative transcript produced in squint a175. We found that in contrast to wild-type squint mRNA, control RNA, non-protein coding squint RNA or squint a175 RNA did not cause any phenotypes and did not rescue MZsquint a175 mutants. These results indicate that squint does not have the previously proposed non-coding functions and is not a member of the cncRNA family.
Transcript-independent phenotype at lnc-phox2bb locus
Lnc-phox2bb neighbors phox2bb and smtnl1. Phox2bb is a transcription factor implicated in the development of the sympathetic nervous system54,55,56, while smtnl1 has been implicated in smooth muscle contraction 57. Whole-gene deletion of lnc-phox2bb (lnc-phox2bba177) (Fig 6A) led to jaw deformation and failure to inflate the swim-bladder (Fig 6B), and no homozygous mutant fish survived to adulthood. Like the whole-gene deletion allele, the TSS-deletion allele (lnc-phox2bba176) lacked lnc-phox2bb RNA (Fig S5 and Fig 6E), but in contrast to the whole-gene deletion mutants, TSS-deletion mutants developed normally and gave rise to fertile adults. To determine the cause of this difference, we analyzed the expression level and pattern of neighboring genes. We found that the anterior expression domain of phox2bb in the hindbrain was absent in the whole-gene deletion allele (Fig 6D). This finding is consistent with the observation that the deleted region contains enhancer elements for phox2bb58, conserved nongenic elements (CNEs)59, and histone marks related to enhancer regions (H3K4me1 and H3K27Ac)60. We also found that the expression level of smtnl1 increased in gene deletion mutants relative to the TSS-deletion mutant and wild type (Fig 6E). These results indicate that lnc-phox2bb RNA is not required for normal development but that the lnc-phox2bb overlaps with regulatory elements required for proper expression of phox2bb and smtnl1 (Fig 6E).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A) The red dashed lines depict the respective positions of the lnc-phox2bb TSS and gene deletion. Arrows flanking black dotted line mark the primer binging sites for qRT-PCR product. B) Homozygous gene deletion mutants but not the TSS-deletion mutants show embryonic defects in jaw formation (arrow head) and swim bladder inflation (asterisk) by 4-dpf. C) Histone marks (H3K4me1 and H3K27ac) associated with enhancer activity60 and conserved non-genic elements CNEs59 overlap with gene deletion. D) phox2bb expression pattern in the TSS and gene deletions. E) qRT-PCR analysis on MZ TSS-deletion and gene deletion mutants.
In summary, our systematic mutant studies indicate that none of the 25 lncRNAs analyzed here are essential for embryogenesis, viability or fertility, including the prominent lncRNAs cyrano, gas5, and lnc-setd1ba. Additionally, they refute the proposed non-coding function of squint RNA. This mutant collection can now be analyzed for potentially context specific, redundant or subtle functions, but extrapolation suggests that most individual zebrafish lncRNAs are not required for embryogenesis, viability or fertility.
Materials and Methods
Animal care
TL/AB zebrafish (Danio rerio) were used as wild-type fish in this study. Fish were maintained on daily 14hr (light): 10hr (dark) cycle at 28°C. All animal work was performed at the facilities of Harvard University, Faculty of Arts & Sciences (HU/FAS). This study was approved by the Harvard University/Faculty of Arts & Sciences Standing Committee on the Use of Animals in Research & Teaching (IACUC; Protocol #25-08)
Cas9 mediated mutagenesis
Guide RNAs (gRNAs) were designed using CHOPCHOP61 and synthesized in pool for each candidate as previously described62. (See supplementary file 1 for the gRNA sequences). gRNAs were combined with Cas9 protein (50 μM) and co-injected (~1 nL) into the one-cell stage TL/AB wild-type embryos. Genomic DNA from 10 injected and 10 un-injected siblings was extracted63 and screened for the difference in amplified band pattern from the targeted region (See supplementary file 1 for the genotyping primer sequences). The rest of injected embryos were raised to adulthood, crossed to wild-type fish and screened for passing the mutant allele to the next generation. Founder fish with desirable mutations were selected and confirmed by Sanger sequencing of the amplified mutant allele. Heterozygous mutants were crossed together to generate homozygous mutants. At least 15 adult homozygous mutant pairs per allele were crossed to test fertility of mutants and to generate maternal and zygotic mutants (MZ) devoid of maternally and zygotic lncRNA activity.
Phenotype scoring procedure
Visual assessment of live embryos and larvae performed30,31 to identify mutant phenotypes, ranging from gastrulation movements and axis formation to the formation of brain, spinal cord, floor plate, notochord, somites, eyes, ears, heart, blood, pigmentation, vessels, kidney, pharyngeal arches, head skeleton, liver, and gut. At day five formation of swim bladder and overall appearance of the embryos were checked again (at any stage 60-100 embryos were scored). Sixty to hundred fish from heterozygous mutant crosses were grown to adulthood and genotyped to identify the viability of adult homozygous fish. Validated homozygous mutant fish were further crossed together to test for potential fertility phenotypes or putative maternal functions of candidate lncRNAs.
Antisense RNA synthesis and in situ hybridization
Antisense probes for in situ hybridization were transcribed using the DIG RNA labeling kit (Roche). All RNAs were purified using EZNA Total RNA kits (Omega Biotek). Embryos were fixed in 4% formaldehyde overnight at 4°C (embryos younger than 50% epiboly fixed for 2 days). In situ hybridizations were performed according to standard protocols 64. NBT/BCIP/Alkaline phosphatase-stained embryos were dehydrated in methanol and imaged in benzyl benzoate:benzyl alcohol (BBBA) using a Zeiss Axio Imager.Z1 microscope.
qRT-PCR
Total RNA was isolated from 3 individuals or sets of 10-20 embryos per condition using EZNA Total RNA kits (Omega Biotek). cDNA was generated using iScript cDNA Synthesis kit (Bio-Rad). qPCR was conducted using iTaq Universal SYBR Green Supermix (Bio-Rad) on a CFX96 (Bio-Rad). Gene expression levels were calculated relative to a reference gene, ef1a. The mean and standard deviation was plotted for each condition. Three technical replicates were used per condition. The qPCR primer sequences are listed in supplementary file 1.
Bright field Imaging
Embryos were anesthetized in Tricaine (Sigma) and mounted in 1% low melting temperature agarose (Sigma) with Tricaine, then imaged using a Zeiss SteREO Discovery.V12 microscope or Zeiss Axio Imager.Z1 microscope. Images were processed in FIJI/ImageJ 65. Brightness, contrast and color balance was applied uniformly to images.
Sense RNA synthesis and injection
The sequences for the wild-type squint mRNA, non-protein coding squint transcript (One Adenine base was added after 8 in-frame ATG codons, and the 3’UTR sequence kept unchanged) and the squinta175 transcript were synthesized as gBlocks (IDT) containing 5’ XhoI cut site and 3’ NotI site. Fragments were digested and inserted the pCS2 plasmid. Positive colonies were selected, and sanger sequenced to assure the accuracy of the gene synthesis process. Sequences of the constructs are provided in supplementary file 1. mRNA was in vitro transcribed by mMessage mMachine (Ambion) and purified by EZNA Total RNA kits (Omega Biotek). H2B-GFP was used as control mRNA. Each injection mix contained 30ng/ul of squint or control mRNA). 1nl of mRNA mix was injected into the yolk of one-cell stage embryos.
Author Contributions
M.G and A.F.S. designed the study, interpreted the data and wrote the manuscript. M.G. generated all data with support from K.B. and L.P.
Acknowledgments
We thank current and former members of the Schier laboratory, particularly Andrea Pauli and Guo-Liang Chew for their helpful suggestions and support during the early phases of this project, Jeffrey Farrell, Nathan Lord and Maxwell Shafer for their critical comments on the manuscript, and the Harvard zebrafish facility staff for technical support. This work was supported by Leopoldina postdoctoral fellowship LPDS2014-01 to M.G. and NIH grant R01HD076708 to A.F.S.
References




