

Abstract
This study seeks to better quantify the parameters that drove the evolution of flight from nonvolant winged dinosaurs to modern birds. In order to explore this issue, we used fossil data to model the feathered forelimb of Caudipteryx, the most basal non-volant maniraptoran dinosaur with elongate pennaceous feathers that could be described as forming proto-wings. In order to quantify the limiting flight factors, we created three hypothetical wing profiles for Caudipteryx representing incrementally larger wingspans, which we compared to the actual wing morphology as what revealed through fossils. These four models were analyzed under varying air speed, wing beat amplitude, and wing beat frequency to determine lift, thrust potential and metabolic requirements. We tested these models using theoretical equations in order to mathematically describe the evolutionary changes observed during the evolution of modern birds from a winged terrestrial theropod like Caudipteryx. Caudipteryx could not fly, but this research indicates that with a large enough wing span Caudipteryx-like animal could have flown, the morphology of the shoulder girdle would not actually accommodate the necessary flapping angle and metabolic demands would be much too high to be functional. The results of these analyses mathematically confirm that during the evolution of energetically efficient powered flight in derived maniraptorans, body weight had to decrease and wing area/wing profile needed to increase together with the flapping angle and surface area for the attachment of the flight muscles. This study quantifies the morphological changes that we observe in the pennaraptoran fossil record in the overall decrease in body size in paravians, the increased wing surface area in Archaeopteryx relative to Caudipteryx, and changes observed in the morphology of the thoracic girdle, namely the orientation of the glenoid and the enlargement of the sternum.
Introduction
The origin of birds has still been a theme of considerable scientific debate [1–7]. Currently it is nearly universally accepted that Aves belongs to the derived clade of theropod dinosaurs, the Maniraptora [8,9]. The oviraptorosaur Caudipteryx is a member of this clade and the basal-most maniraptoran with pennaceous feathers [10–12]. The longest of these feathers are located distally on the forelimb, strongly resembling the ‘wings’ of birds. However, the relative length of these feathers compared to those in volant birds and the brevity of the forelimb itself compared to the si/e of the body and length of the hindlimbs all point to a clearly terrestrial animal [13–15]. It is understood that winged forelimbs must have evolved first for some other purpose and were later exapted for flight. Hypotheses regarding their original function range from ornamentation, temperature regulation, or locomotion [16–21]. The feathers on the forelimbs and tail in Caudipteryx are straight with symmetrical vanes similar to the wing feathers in flightless birds. It’s certainly clear that Caudipteryx couldn’t fly [22–24]. However, as a fairly fast cursorial animal, the presence of feathered distal forelimbs must have had some effect on the locomotion of Caudipteryx. The aerodynamic effect of the basal-most known forelimbs with pennaceous feathers has the potential to shed light on the evolution of flying wings in Paraves (see Supplementary Materials for more basic information about the flight kinematics of bird’s wings).
We mathematically modeled Caudipteryx with three hypothetical wing sizes (morphotypes B, C, and D) across a range of input values for flapping angle, wing beat frequency and velocity (the hypothetical wingspans are based on such modern birds listed in S1 Table and the wing profiles are inspired by an eagle [25]). We explore what values would be necessary to allow Caudipteryx to fly. This analysis quantifies the physical constraints that exclude the cursorial Caudipteryx from engaging in volant behavior despite the presence of feathered forelimbs and hint at the evolutionary changes necessary to evolve flight in the maniraptoran lineage from a cursorial animal with small wings like Caudipteryx to the neornithine condition, which is supported through observations from the fossil record of pennaraptorans.

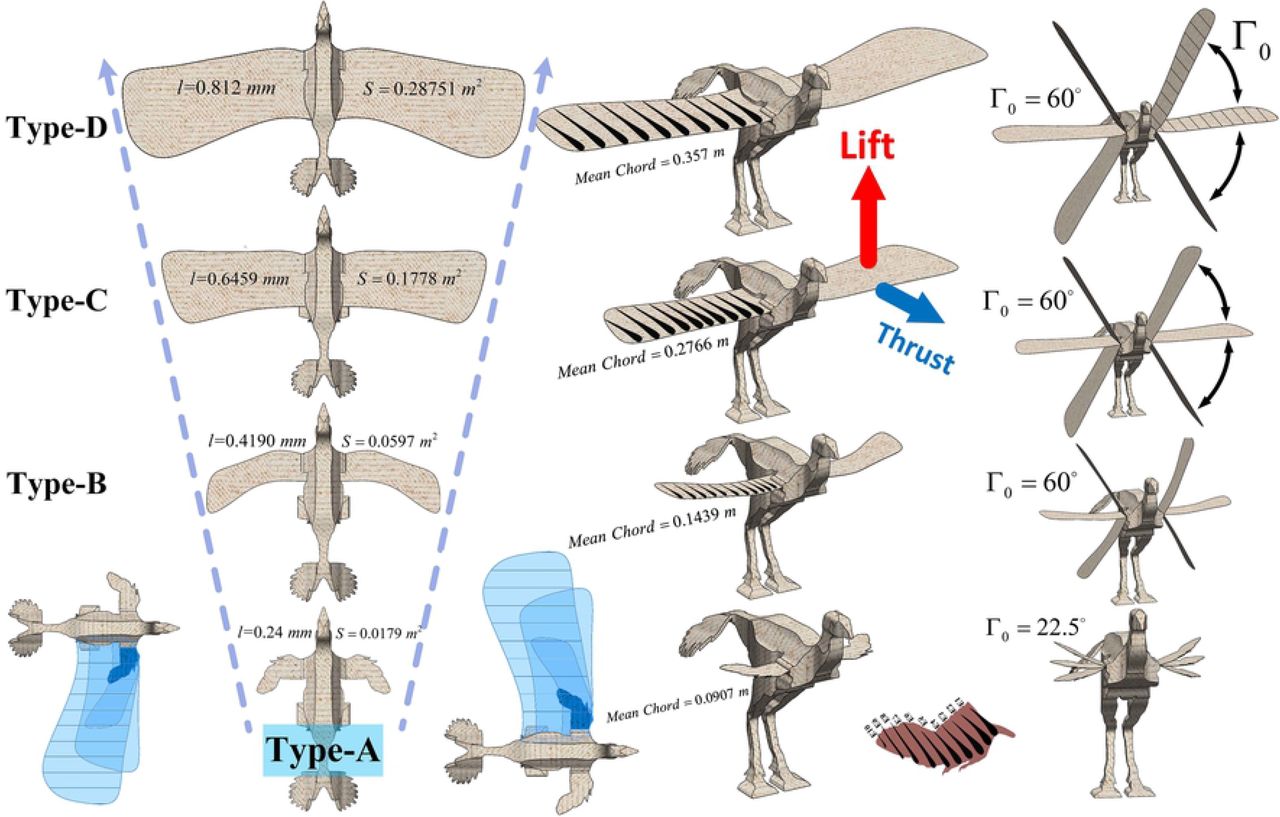
Type A is an actual Caudipteryx’s model and the wings in Types B, C and D are assumed in different sizes changing in linear form.
Materials and methods
In order to achieve a reasonable accuracy in analyses, each wing type (Fig 1) is divided into ten elements, each modeled using unsteady aerodynamics in order to capture lift, thrust/drag and required power (Fig 2). Type-A is based on fossil data and represents a realistic estimate of wing length (l) which we measure to be 0.24 meters. Types-B, C and D are hypothetical models with increasing wing lengths of 0.419, 0.6459 and 0.812 meters, respectively, and for the sake of simplicity, we approximately calculate the areas of the wings with the equivalent rectangular ones. Wings size in terms of area S (m2), mean chord length C (m), aspect ratio (λ), flapping angle (Γ0) and other wing angles describe the state of the wing (Fig 1 and Table 1). The element’s motion includes a plunging velocity ḣ and pitch angle (twist angle) θ. We assumed that the wing’s aspect ratio is large enough to pass flow through each element in the mean stream direction. Hence, the normal force dN of the element’s total attached flow, is equal to the normal force dNa of element’s circular plus apparent mass effect, as an additional normal force contribution which acts at the mid chord. Delaurier and Larijani have given the formulas below based on mathematics, tests and experiments [26–32].
Specifications of Types-A, B, C and D

Kinematics model of flapping flight for each element of the wing of Caudipteryx.

The normal force of the element’s circular and the apparent mass effect are defined as below

 where ρ is the density of the airflow, V is the flow’s relative velocity at the ¼ chord,
where ρ is the density of the airflow, V is the flow’s relative velocity at the ¼ chord,  is the angle between flapping axis and mean stream velocity
is the angle between flapping axis and mean stream velocity  is the mean angle between chord and flapping axis, α0 is the angle of the zero lift line the value is fixed for airfoil in each situation along the wing), α’ is the flow’s relative angle of attack at the ¾ chord (Fig 2), it’s given by
is the mean angle between chord and flapping axis, α0 is the angle of the zero lift line the value is fixed for airfoil in each situation along the wing), α’ is the flow’s relative angle of attack at the ¾ chord (Fig 2), it’s given by
 where λ is the aspect ratio of the wing of Caudipteryx, c is the wing element chord length, k = cω / 2U which is the reduced frequency (ω is in radians and ω = 2πf, and f is in hertz). Equation (4) is simplified by formulation of the modified Theodorsen function which was originally presented by Jones [33], c1 = 0.5λ/(2.32 + λ) and c2 = 0.181 + (0.772/λ). α and α̇ are given by
where λ is the aspect ratio of the wing of Caudipteryx, c is the wing element chord length, k = cω / 2U which is the reduced frequency (ω is in radians and ω = 2πf, and f is in hertz). Equation (4) is simplified by formulation of the modified Theodorsen function which was originally presented by Jones [33], c1 = 0.5λ/(2.32 + λ) and c2 = 0.181 + (0.772/λ). α and α̇ are given by

 where α is the relative angle of attack at the ¾ chord. The pitch angle θ is
where α is the relative angle of attack at the ¾ chord. The pitch angle θ is  where δθ is the dynamically varying pitch angle (Fig 2). The θ(y, t) is a function of y and ω. Therefore, δθ(y, t) = −β0y sin(ωt) which prescribes the twist of wing of Caudipteryx where β0 is a constant representing the twist angle per unit distance along the wing span (°/m). Hence, the first and second derivatives of θ(y, t) with respect to time are written as
where δθ is the dynamically varying pitch angle (Fig 2). The θ(y, t) is a function of y and ω. Therefore, δθ(y, t) = −β0y sin(ωt) which prescribes the twist of wing of Caudipteryx where β0 is a constant representing the twist angle per unit distance along the wing span (°/m). Hence, the first and second derivatives of θ(y, t) with respect to time are written as


The plunging displacement of each element is h(t) = (Γ0y) × cos(ωt) (the imposed motion), where Γ0 is the maximum flapping amplitude and y is the distance between flapping axis (wing base) and center of a wing segment. Therefore, the plunging velocity ḣ (t) and acceleration ḧ(t) at the leading edge are given as


Pitch angle (twist angle) changes during flapping and these variations are proportional to flapping angle in a cycle (S2 Fig). The wing motion of Caudipteryx relative to U is included in the flow velocity V given in equation (2) and also term of α’ is taken into account, V is

The element’s circulation distribution generates forces along the chord axis direction (Fig 2), hence the chordwise force due to the camber (dDcamber) and chordwise friction drag due to viscosity are given by
 where (Cd)f = 0.89/ (log(Rechord))2.58 is the friction drag coefficient [34] of the skin that is included in Reynolds’ number of local chord length and
where (Cd)f = 0.89/ (log(Rechord))2.58 is the friction drag coefficient [34] of the skin that is included in Reynolds’ number of local chord length and  is the tangential flow speed to the element. For the two dimensional airfoil, Garrick expressed dTs for the leading edge as below [35,36]
is the tangential flow speed to the element. For the two dimensional airfoil, Garrick expressed dTs for the leading edge as below [35,36]
 where ηs is the efficiency term which presents that in reality because of viscous effects, the efficiency of leading edge of most airfoils are less than 100%. Therefore, the total chordwise force along the chord axis is expressed with
where ηs is the efficiency term which presents that in reality because of viscous effects, the efficiency of leading edge of most airfoils are less than 100%. Therefore, the total chordwise force along the chord axis is expressed with

Therefore, the equations of element’s instantaneous lift and thrust are rewritten as

 where dL and dT are the instantaneous lift and thrust of the element, respectively. To integrate along the wingspan and to get whole wing’s instantaneous lift and thrust, for wings, it is given by
where dL and dT are the instantaneous lift and thrust of the element, respectively. To integrate along the wingspan and to get whole wing’s instantaneous lift and thrust, for wings, it is given by
 where l is the wing span length. The average of the lift and thrust can be obtained via integrating equation (17) over a cycle as φ = ωt.
where l is the wing span length. The average of the lift and thrust can be obtained via integrating equation (17) over a cycle as φ = ωt.
 where
where  and
and  are the averages of the lift and thrust, individually. To calculate the required power against the forces, it is represented for attached flow by
are the averages of the lift and thrust, individually. To calculate the required power against the forces, it is represented for attached flow by
 where dMac is the element’s pitching moment about its aerodynamic center and dMa is composed of apparent chamber and apparent inertia moments are given respectively as follows
where dMac is the element’s pitching moment about its aerodynamic center and dMa is composed of apparent chamber and apparent inertia moments are given respectively as follows

 where Cmac is the coefficient of airfoil moment about its aerodynamic center. The instantaneously required power, Pinput (t), for a whole wing is derived below
where Cmac is the coefficient of airfoil moment about its aerodynamic center. The instantaneously required power, Pinput (t), for a whole wing is derived below

Then the average input power in a cycle is found from


Illustrations of the avian flight apparatus in Archaeopteryx and living birds. A keeled sternum indicates increased capacity for flight based on the increased surface area for the attachment of the flight muscles. The realistic mode and reconstructed wing type (type-A) is based on Caudipteryx fossil [24].
The weight of body is computed about 50 N for the holotype of Caudipteryx dongi-IVPP V12344-Based on equation relating femur length to mass [37], the average of the lengths of the left and right femur are both 149 mm. With the formulas above, we compare the lift, thrust and power of each model with regards to the changes in wing angles (flapping angle (S4 Table),  (S5 Fig) and dynamic twist angle (S6 Fig)), flapping frequency from lower frequency (1 Hertz) (S9 Fig and S10 Fig) to higher frequency (8 Hertz) (Fig 4) and velocity while the range of the air velocity changes from almost zero (U=0.05 m/s) (Fig 4, S8 Fig and S9 Fig) to higher speed (U= 10 m/s) (Fig 4 and S9 Fig). In the analyses on type-A, the wing angles are
(S5 Fig) and dynamic twist angle (S6 Fig)), flapping frequency from lower frequency (1 Hertz) (S9 Fig and S10 Fig) to higher frequency (8 Hertz) (Fig 4) and velocity while the range of the air velocity changes from almost zero (U=0.05 m/s) (Fig 4, S8 Fig and S9 Fig) to higher speed (U= 10 m/s) (Fig 4 and S9 Fig). In the analyses on type-A, the wing angles are  ,
,  , α0 = 30deg, β0 = 180deg deg and Γ0 = 22.5deg and in hypothetical models(types B, C and D), the corresponding parameters are
, α0 = 30deg, β0 = 180deg deg and Γ0 = 22.5deg and in hypothetical models(types B, C and D), the corresponding parameters are  ,
,  , α0 = 1deg, β0 = 10 deg and Γ0 = 60 deg.
, α0 = 1deg, β0 = 10 deg and Γ0 = 60 deg.

Variation in lift and thrust of Caudipteryx with respect to flapping frequency at any velocity.
Results
When  is 15 deg the realistic Caudipteryx (type-A) is capable of producing maximum thrust force but to have only vertical motion (maximum lift force with a small value of thrust),
is 15 deg the realistic Caudipteryx (type-A) is capable of producing maximum thrust force but to have only vertical motion (maximum lift force with a small value of thrust),  must be increased to 45 deg while U is 0.05 m/s (velocity is almost zero) (S5 Fig). In other hypothetical models (types B, C and D) the situation is almost similar to the realistic one (i.e. with the angle of 20 deg for maximum thrust and 45 deg for maximum lift). Hence, lift increases and thrust decreases as the angle of
must be increased to 45 deg while U is 0.05 m/s (velocity is almost zero) (S5 Fig). In other hypothetical models (types B, C and D) the situation is almost similar to the realistic one (i.e. with the angle of 20 deg for maximum thrust and 45 deg for maximum lift). Hence, lift increases and thrust decreases as the angle of  increases. In all models consumed power is proportional to the thrust force (S5 Fig) and the maximum thrust force requires maximum power.
increases. In all models consumed power is proportional to the thrust force (S5 Fig) and the maximum thrust force requires maximum power.
The feathers in type-A were modeled as softer than those in modern birds (being primitive and symmetrical), and as the dynamic twist angle β0 expresses the stiffness of feathers along the wing span, we presumed β0 = 180° when lift and thrust are at their maximum values. The best and optimum value of this angle for three hypothetical models (types B, C and D) is 10 deg when the velocity is almost zero in one cycle (f=1Hz) (S6 Fig). In all models, as the flapping angle (wing beat amplitude) Γ0 increases, lift, thrust and the required energy also increase simultaneously (S4 Fig).
When birds supinate their wings, drag forces help the birds to brake. Larger wings moving at higher velocities produce greater braking forces (S7 Fig). Based on our analysis, the wings of Caudipteryx (type-A) utilizing both lower and higher wing beat frequencies at any velocity, are always capable of generating positive lift, but thrust force is only positive when the velocity is near zero and is negative during movement at any larger velocity (Fig 4). Hence in type-A, when flapping frequency and velocity increase, lift also increases but thrust converts to the drag force (negative thrust) causing a corresponding increase in metabolic demands (S3 Fig). In all hypothetical wings (types-B, C and D) lift is positive; an increase in flapping frequency at any velocity generates an increase in lift (Fig 4 and S9 Fig).
Although hypothetical wing type-B cannot provide sufficient lift force to overcome gravity (Fig 4 and S8 Fig), type-C is theoretically capable of supporting flight at lower velocity and higher wing beat frequency (namely U=1~2 m/s and frequency=6~8 Hz) but cannot generate large enough lift at higher velocities (Fig 4 and S9 Fig). The larger wings in type-D are capable of sustaining faster flight (8 m/s) with higher flapping frequency (f=4~8 Hz). Heavier birds flying at higher velocities require greater wing span. The obtained lift and thrust forces from Type-A deduced by experiments (S11 Fig) support our theoretical calculations (S2 Table).
Discussion
Powered flight is the most physically demanding form of locomotion. Thus, during the course of evolution of flight from non-volant dinosaurs such as Caudipteryx to Aves, major skeletal adaptations needed to evolve in order to allow the necessary range of motion, accommodate the necessary musculature, and produce these adaptations in a light weight framework. These adaptations include modification of the glenoid to facilitate greater flapping angles and enlargement of the sternum to support the increased musculature necessary to generate significant thrust forces. This had to be paired with the ability to meet the metabolic demands of sustaining flight (the neornithine digestive system is highly specialized) [38–40]. We can infer these changes based on observations of the fossil record. However, the aerodynamic limitations that drove these evolutionary changes have never been explored.
In order to explore this idea, we tested the impact of increasing wing size in Caudipteryx within its actual skeletal framework. We considered three hypothetical wing morphologies (types B, C, and D) that vary in wing span, wing profile, wing chord, and aspect ratio (Fig 1). The goal in this study is to assess the effects of increasing wing size in a terrestrial form in order to identify which variables are limiting volant behavior. We focused on flight kinematic parameters during take off. The flight kinematics formulas utilized here to calculate lift, thrust and power allow us to compare all states in varying frequencies, velocities and flapping angles while ranging air stream velocity changes from almost zero to high speeds. This provides a comprehensive understanding of all important flight parameters.
In type A, the feathers are assumed to be weaker than those in types B, C and D and the flapping angle is much lower, which reflects the basal position of these non-aerodynamic feathers and the morphology of the glenoid. Increasing frequency and flapping angle from ±22.5 deg to ±60 deg (in types B, C and D) generates higher lift force (see S3 Table and S4 Table in Supplementary Materials for more information about the values of flapping beats and flapping angles). However, the theoretical requirements for metabolic power and muscle performance are unrealistic. This quantifies the observed changes in the scapulocoracoid observed during the evolution of birds from non-volant dinosaurs. Our analyses express that during low flapping frequency (f=3 Hz) and when the wind velocity is almost zero in wing types B and C (near type-C) the lift force generated by the wings is nearly enough to overcome gravity for take off. At low velocity and with a higher hindlimb frequency, type-C is able to meet the lift requirements. In this type, although the wings would not be able to provide enough force to fly faster, type C can supply the capability of flying at lower speed. The wings in type D are larger allowing the model Caudipteryx to fly faster than 8 m/s with higher wing beat frequency. Not surprisingly, the largest wings (type D) generate the greatest lift, thrust forces, and drag force during braking. However, as flapping frequency increases, so do the required thrust forces and metabolic requirements.
The aerodynamic behavior of the realistic Caudipteryx model, reconstructed from fossil material (type A), reveals that the wings could generate lift but at any velocity produced more drag than thrust, and thus probably did not evolve in this taxon for aerodynamic function. The analyses indicate that for a given lift or thrust force, all hypothetical types of Caudipteryx would have to increase wing beat frequency to increase their velocities. It depends on wingspan that produces capability of flying, those generations of the new birds whose wing’s profile is similar to those of types B and C, they need to decrease their self-weight during evolution to obtain enough lift and thrust for flight in accordance with the weight. Heavy modern birds (such as Black-browed albatross or Wandering albatross-see Table S1 in Supplementary Materials) shall have large enough wingspan more than that of type C to keep balance between lift force and their body weight.
In order to take off to overcome gravity, sustain flapping flight and maintain lift forces to oppose body-weight, flapping frequency, and flapping angle (as the most significant parameter) and wing span had to increase. At small flapping angles, like those present in Caudipteryx and other non-volant maniraptorans, the wing beat cannot generate large enough lift and thrust. This verifies the observed changes in shoulder architecture in the lineage to Aves from a more ventrolateral (non-volants) or laterally facing (Archaeopteryx) glenoid to the dorsolaterally facing glenoid in birds–clearly the wings of Caudipteryx are the smallest among the wings of known Mesozoic pennaraptorans and the birds, and the wings of volant dromaeosaurid Microraptor are much larger. Heavier flying birds need more lift to become airborne and thus require larger wingspans with large flapping angles and strong flight muscles.
Depending on the wing profile, the flapping angle should be more than 60 degrees (i.e. with a total range of motion of 120 degrees). To accommodate this range of motion, it required the dorsal expansion of the glenoid and an increase in wing musculature to manipulate the larger wings against greater aerodynamic resistance. This would drive up the metabolic cost of flight so high as to make this form of locomotion inefficient. However, this can be obviated by reducing the cost of flapping flight with an overall reduction in body weight.
Observations from fossils suggest that this was achieved both through a reduction in overall body size and by the evolution of additional pneumaticity and thinner bone cortices. Morphological changes of the skeleton such as the reduction of tail, and modification of numerous biological systems such as the loss of the right ovary and evolution of a highly efficient digestive system. This ultimately produced a body shape that was well adapted to spindle shaped, generating less resistance (drag force) during flight (Fig 3) [25,40,41]. Therefore, it is important for efficient flight to decrease body mass and to increase musculature, wing profile, aspect ratio, flapping angle in the evolution from non-volant maniraptoran dinosaur to modern birds.
Author Contributions
Author contributions: Y. S. deduced formulas and prepared programs, simulations, tables and Figures and wrote the first draft of the manuscript; Y. S. and Y. F. accomplished the experiments and completed the 3D reconstruction and video of the Caudipteryx robot; J.-S. supervised the project and proposed the experiment principle; Y.-S., Y. F., J.-S. and J. K. completed the Cladogram of Caudipteryx and investigation of the feathers of the dinosaurs; J. K. provided the fossil and analysis of dinosaurs and provided the major suggestions in revision; All authors discussed the results and commented on the manuscript and contributed ideas to manuscript development and data analysis.
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.
Competing interests:
The authors have declared that no competing interests exist.
Acknowledgements
The authors acknowledge the kind suggestions from Prof. Dr. Corwin Sullivan from the Department of Biological Sciences, University of Alberta, Canada. Prof. Dr. Zhong-He Zhou and Prof. Dr. Min Wang from the Key Laboratory of Vertebrate Evolution and Human Origins, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, 100044, P. R. China. This project was supported by the National Natural Science Foundation of China under grant 51575291, the National Major Science and Technology Project of China under grant 2015ZX04002101, State Key Laboratory of Tribology, Tsinghua University, and the 221 program of Tsinghua University.





{kind=link}
{kind=link}
{kind=link}
{kind=link}