

Abstract
Circadian clocks control daily rhythms in physiology. In Drosophila, the small ventral lateral neurons (sLNvs) expressing PIGMENT DISPERSING FACTOR (PDF) are the master pacemaker neurons. Despite the importance of sLNvs and PDF in circadian behavior, little is known about factors that control sLNvs maintenance and PDF accumulation. Here, we identify the Drosophila SWI2/SNF2 protein DOMINO (DOM) as a key regulator of circadian behavior. Depletion of DOM eliminates morning anticipation and impairs rhythmicity. Interestingly, the two splice variants of DOM, DOM-A and DOM-B have distinct circadian functions. DOM-A depletion leads to arrhythmic behavior, while DOM-B knockdown lengthens circadian period. Both DOM-A and DOM-B bind to the promotor regions of key pacemaker genes period and timeless, and regulate their protein expression. Furthermore, we identify that DOM-A is required for the maintenance of sLNvs and transcription of pdf. Lastly, constitutive activation of PDF-receptor signaling rescued the arrhythmia and period lengthening of DOM downregulation. Taken together, our findings reveal that splice variants of DOM play distinct roles in circadian rhythms through regulating abundance of pacemaker proteins and sLNvs maintenance.
Introduction
Circadian clocks allow animals to anticipate daily oscillations in behavior, physiology and metabolism [1]. The core of the molecular clock is a negative transcriptional-translational feedback loop, which is evolutionarily conserved across species [2]. The fruit fly Drosophila melanogaster has been a powerful model in dissecting the molecular and neuronal mechanisms of circadian rhythms. In Drosophila, a heterodimeric complex of CLOCK (CLK) and CYCLE (CYC) activates rhythmic transcription of clock-controlled genes, including the transcriptional repressor period (per) and timeless (tim). PER and TIM dimerize and repress their own transcription by blocking CLK-CYC transactivation [3]. A number of kinases (CK1, SGG, NEMO, etc) and phosphatases (PP1, PP2A) also control the circadian clock at the post-translational level [3].
Circadian locomotor rhythms are controlled by a small set of clock neurons expressing core pacemaker proteins in the brain. In each hemisphere of the fly brain, there are ∼75 clock neurons, which are divided into clusters based on their anatomical locations and functions in circadian behavior [4]. There are three groups of dorsal neurons (DN1s, DN2s, and DN3s), the lateral dorsal neurons (LNds), the lateral posterior neurons (LPNs), the large ventral lateral neurons (lLNvs), and the small ventral lateral neurons (sLNvs). The ILNvs and four sLNvs express the neuropeptide PIGMENT DISPERSING FACTOR (PDF), while the fifth sLNv is PDF negative. The PDF positive sLNvs are the key pacemaker neurons: they control the circadian rhythmicity under constant darkness (DD) [4]. They are also critical for generating morning anticipatory activity before lights on (i.e. dawn) under light-dark cycle (LD) [5,6]. The sLNvs send axonal projections towards the DN1s and DN2s, and the structural plasticity of dorsal projections has been shown to be under circadian control [7-9]. PDF positive sLNvs and their dorsal projections are formed in 4 hrs old first instar larvae after hatching [10].
Despite the importance of sLNvs in the circadian behavior, little is known about the mechanisms that control the maintenance of sLNvs.
PDF are the most prominent neuropeptide in the regulation of circadian behavioral rhythms [11]. PDF plays a critical role in the synchronization of different groups of clock neurons by binding to PDF receptor (PDFR), and activating cAMP signaling [12-15]. Loss of PDF or PDFR abolishes morning anticipation and significantly impairs behavioral rhythmicity in DD. Not only as a critical circadian output signal, PDF has also been shown to control the molecular clock by regulating the stability of pacemaker proteins PER and TIM recently [16,17]. PDF abundance in the sLNvs is regulated by CLK and CYC [18-20]. Another pacemaker protein VRILLE also promotes PDF levels [21]. Despite these studies, mechanisms regulating PDF abundance remain poorly understood.
DOM plays a critical role for incorporation and removal of the only H2A variant-H2A.V in Drosophila [22]. DOM is a chromatin-remodeling protein, which belongs to the SWI/SNF2 DNA-dependent ATPase family [23]. DOM is involved in oogenesis, wing development, cell viability and proliferation, neuroblast maintenance and polarity, as well as in dendrite development [23-26]. In the fly genome, two alternative splicing variants of dom encode DOM-A and DOM-B. Interestingly, a recent study identified that DOM-A and DOM-B play distinct roles in cell-type specific development during Drosophila oogenesis [24].
A few chromatin remodelers have been shown to regulate circadian photo-responses and gene expression [27-29]. However, the role played by chromatin remodeling in the control of Drosophila circadian clocks is still largely unknown. Here, we report the functions of DOM in the regulation of circadian rhythms. Using isoform-specific RNAi and rescue, we demonstrate distinct functions of DOM-A and DOM-B in the regulation of circadian locomotor rhythms. Depletion of DOM-A in circadian neurons leads to arrhythmic behavior and long period, while DOM-B downregulation specifically lengthens the circadian period in flies. Both DOM-A and DOM-B bind to the promoters of per and tim, and regulate the abundance of PER and TIM levels, which is critical for the control of circadian period. However, DOM-A is specifically necessary for maintenance of sLNvs as well as accumulation of PDF in these neurons. Indeed, activation of PDFR signaling restores the locomotor rhythms of DOM downregulation. Together, our results suggest that the two alternative spliced variants of DOM play distinct roles in circadian rhythms.
Results
DOM regulates circadian locomotor rhythms
Mass spectrometry (MS) label-free quantitative proteomics approach was previously performed to identify the BRAHMA (BRM) chromatin-remodeling protein complex as interactor of circadian clock proteins [30]. In the same data set, we observed that several core subunits of the ATP-dependent DOM chromatin-remodeling complex, including DOM, NIPPED-A, PONT, REPT, and Mrg15, interact with CLK in the nucleus of Drosophila S2 cells (Table 1). Since DOM is the ATPase subunit of the protein complex and shows significant binding to CLK (especially C-terminal FLAG-tagged CLK) according to SAINT (Significant Analysis of INTeractome) scoring [31], we decided to further investigate its role in circadian regulation. Interestingly, prior studies indicated that dom mRNA levels in the sLNvs are clearly enriched about 4.4-folds compared to other neurons [32].
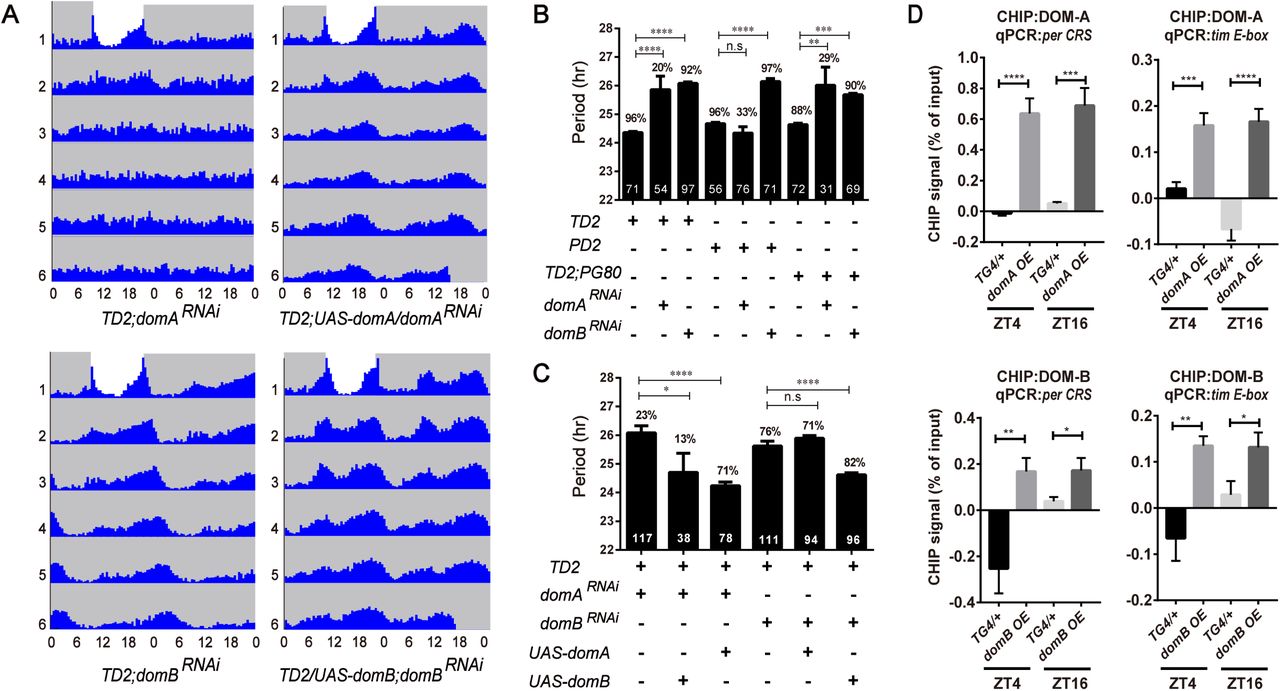
Homozygous dom mutants are lethal, therefore we used RNAi to deplete DOM in circadian neurons. Dicer 2 was co-expressed to enhance RNAi efficiency and has been shown to have no effects on circadian rhythms [33,34]. When we expressed dom dsRNAs using tim-GAL4, a circadian tissue-specific driver [35], most of the flies became arrhythmic, and the amplitude of rhythm was significantly reduced (Fig. 1A and Appendix Table S1). Only 31% of flies with dom downregulation (41% for another independent line dom RNAi#2) were rhythmic, and interestingly the circadian period of these lines was about 1.5 hours longer than the control. The reduced rhythmicity and amplitude were also observed when we only targeted the PDF positive LNvs using pdf-GAL4 [11] (Fig. 1A). However, neither percentage of rhythmicity nor amplitude was affected when we targeted all circadian cells except the PDF positive LNvs, using a combination of tim-GAL4 with the repressor transgene pdf-GAL80 (Fig. 1A). However, a slight period-lengthening of activity rhythm was detected. These results indicated that DOM primarily functions in the LNvs to control circadian behavior, but might be needed in PDF negative circadian neurons to fine-tune circadian period length.

A. Free-running period (top panel) and power (bottom panel) of DOM depleted flies. The percentages of rhythmic flies are shown above each column. The number of tested flies is shown in each column. dom mRNA targeted by the two non-overlapping RNAi lines from the TRiP stock center. domRNAi#1 (JF01502) is a long double-stranded RNAi line, while domRNAi#2 (HMS01855) is a short hairpin RNAi line. TD2=tim-GAL4, UAS-dcr2, PD2= pdf-GAL4, UAS-dcr2, TD2; PG80= TD2, pdf-GAL80. n= 50–90; error bars represent ± SD; n.s., non-significant, **p < 0.01, ***p < 0.001, ****P < 0.0001; one-way ANOVA.
B. Average locomotor activity of flies under 3 days of 12:12 hr LD conditions. Dark activity bars represent the night, and white bars represent the day. The significant differences of the values (shown above bars) indicate morning anticipation is severely disrupted in dom RNAi lines.
C. Actograms showing the average activities on the last day of LD followed by 5 days in DD. Light represents the day and gray darkness. From left to right: (Left panel) Gal4 control; (middle panel) domRNAi#1 flies; and (right panel) domRNAi#2 flies (knockdown of dom in all circadian neurons). Depletion of DOM caused arrhythmia in DD.
Because of the high arrhythmia in DD, we also examined the behavioral phenotype of dom RNAi flies under LD. We found that DOM knockdown abolished the morning peak of anticipatory activity in flies (Fig. 1B). Importantly, two independent dom RNAi lines targeting non-overlapped regions of dom exhibited similar phenotypes in both LD and DD (Fig. 1C and Appendix Fig S1). Thus, it is unlikely that off-target effect of dom RNAi cause these circadian phenotypes. Furthermore, quantitative RT-PCR results showed that dom expression were significantly reduced in fly heads of DOM knockdown as compared to controls (Appendix Fig S1). Together, these results suggest that DOM is important for regulation of circadian locomotor behavior.
DOM regulates the abundance of PER and TIM
Depletion of DOM severely decreased the rhythmicity and lengthened circadian period. To understand how DOM affects circadian rhythms, we first measured the oscillations of mRNA abundance of three core pacemaker genes in fly heads. For clk, the abundance and oscillation of mRNA under LD and DD were not affected in dom RNAi flies (Fig. 2A). However, we found that the abundance of per and tim mRNA was reduced, especially at the time point of peak expression (Fig. 2A). Under DD, depletion of DOM also decreased the level of per and tim mRNA, which indicates that the effects are not due to masking effects of light (Fig. 2A). Next we used PER as a marker of molecular pacemaker and examined the oscillation of PER in pacemaker neurons across 24 hours under DD. Consistent with the changes in mRNA level, the level of PER was also significantly reduced in the sLNvs with DOM depletion (Fig. 2B, 2C). In other groups of circadian neurons, such as the LNds and DN1s, decreased abundance of PER was also observed (Appendix Fig S2). Taken together, our results suggest that DOM controls the abundance of PER and TIM both at mRNA and protein levels.

A. Quantitative RT-PCR showing the expression of clk, per and tim. Flies were collected at the indicated time points (ZT= Zeitgeber Time or CT= circadian time). Downregulation of dom decreased per and tim mRNA levels (middle and right panels), while clk was normal (left panel).
B. Whole mount immunostaining showing the expression pattern of PER in sLNvs. Red represents PER and green is PDF. Flies were entrained for 4 days in LD and transferred to DD and dissected every 4 h on the fifth day. Downregulation of DOM decreased PER levels at CT1-5 and CT17-21. (Scale bar: 50 um.)
C. Quantification of the staining in sLNvs. For each genotype, 16-20 fly brains and 60-80 neurons were used for quantification. White and black bars indicate lights-on and lights-off, respectively. Gray and black bars indicate subjective day and subjective night, respectively. Time (h) is indicated as ZT or CT where CT0 is 12 h after lights-OFF of the last LD day. Two independent experiments were done for each genotype/condition with very similar results. Error bars correspond to SEM. **p < 0.01, ***p < 0.001, ****p < 0.0001 as determined by the t-test.
DOM-A but not DOM-B is regulated by CLK
There are two major splice variants of dom: dom-A and dom-B, which has distinct functions during Drosophila oogenesis [24]. We wondered whether these two isoforms would have different functions in circadian rhythms. We first examined the expression pattern of dom-A and dom-B in the fly head across different times of the day. Interestingly, dom-A exhibited a strong oscillation pattern with a trough around zeitgeber time 9 (ZT9, as ZT0 is light on and ZT12 is light off) and a peak expression near ZT21 (Appendix Fig S3A). Oscillation of dom-A was also observed under DD. Importantly, the oscillation of dom-A expression was abolished in the Clock null mutant Clkout (Appendix Fig S3A) [36]. Together, these data indicate that dom-A is controlled by circadian clock. However, we did not observe a clear oscillation for dom-B expression (Appendix Fig S3B). In order to detect changes at protein level, we took advantage of isoform-specific antibodies to DOM-A or DOM-B (25). We confirmed the specificity of these antibodies by altering the level of DOM-A or DOM-B using RNAi and overexpression (Appendix Fig S3C-3S). Similar to mRNA level, DOM-A protein abundance was dramatically reduced in Clkout, however, no oscillation of DOM-A was observed in wild-type (Appendix Fig S3E,S3G). This loss of DOM-A oscillation at protein level indicates that DOM-A protein might be quite stable. The levels of DOM-B were not affected in Clkout (Appendix Fig S3F,S3H).
DOM-A and DOM-B have distinct roles in circadian regulation
Differential regulations of DOM-A and DOM-B by CLK suggest that they might have different roles in circadian rhythm. We therefore performed isoform-specific downregulation in circadian tissues using small hairpin RNA (shRNA) targeting dom-A or dom-B. These transgenic fly lines have been previously shown to specifically knockdown these alternative isoforms (Appendix Fig S1) [24]. We first quantified the efficiency of dom-A and dom-B downregulation by shRNAs in fly heads using quantitative real-time PCR. Consistent with previous report [24], we observed that each shRNA line specifically downregulated dom-A or dom-B (Appendix Fig S1B). Interestingly, when we depleted DOM-A or DOM-B in all circadian clock neurons, we observed two distinct circadian behavioral phenotypes. Most of the flies expressing dom-A shRNA lost locomotor rhythms, except for ∼20% that were rhythmic and showed a longer period as compared to the control (Fig. 3A, 3B and Appendix Table S1). As with depletion of all dom isoforms, DOM-A knockdown also blunted the morning activity peak under LD (Appendix Fig S4). However, for DOM-B knockdown, flies exhibited a lengthened period of ∼1.5 hrs (Fig. 3A, 3B), but no effect on amplitude of rhythms or morning anticipation was observed (Fig. 3B, and Appendix Fig S4). When we expressed dom-A or dom-B shRNA only in LNvs, or in PDF negative circadian neurons, we found similar effects on rhythmicity or period-lengthening (Fig. 3B). These results suggested that DOM-A and DOM-B are required in both PDF positive and negative circadian neurons for behavior. It is unlikely that the different circadian phenotype is due to greater efficiency of dom-A over dom-B shRNA (Appendix Fig S1B). Actually a previous study has also validated the knockdown efficiency and found that dom-B shRNA is in fact slightly stronger than dom-A in the fly nervous system [24].

A. Actograms showing the average activities on the last day of LD followed by 5 days in DD. Light represents the day and gray darkness. From top to bottom: domARNAi (Top left panel); domARNAi; UAS-domA rescue(Top right panel); and domBRNAi (Bottom left panel); domBRNAi; UAS-domB rescue(Bottom right panel). Depletion of domA caused arrhythmia in DD, while knocking down domB lengthened circadian period. domARNAi and domBRNAi phenotypes can be rescued by restoring domA and domB expressing in all circadian neurons respectively.
B. Free-running period and percentage of rhythmicity of domA and domB depleted flies. The percentages of rhythmic flies are shown above the error bar. The numbers of tested flies are shown in each column.
C. Free-running period and percentage of rhythmicity of flies with restoring DOM-A or DOM-B in domA or domB depletion.
D. ChIP assays detecting DOM-A and DOM-B binding more at the per CRS and tim E-box in flies expressing domA (BL64261) and domB (BL64263) in tim-expressing cells as compared to control TG4 flies. Non-specific DOM binding was detected by amplifying an intergenic region (FBgn0003638) of the Drosophila genome and subtracted from the signal from the per CRS and tim E-box 1 signals. Results shown are from at least three biological ChIP replicates, with technical triplicates performed during qPCR for each biological replicate. Error bars represent ± SEM; n.s., non significant,*P < 0.05,**p < 0.01, ***p < 0.001, ****P < 0.0001; one-way ANOVA.
Our isoform-specific knockdown indicated that DOM-A and DOM-B has distinct functions in circadian rhythms. Although the shRNA lines have been shown here and in a previous study [24] to specifically knockdown the dom-A or dom-B isoforms, off-target effects could still contribute to the observed phenotypes. Thus, we performed rescue experiments with UAS transgenic flies expressing either dom-A or dom-B cDNA. Circadian behavior defects were rescued when we overexpressed corresponding UAS lines in the DOM-A or DOM-B knockdown (Fig. 3A, 3C), which indicated that the isoform specific phenotype we observed was not due to off-target effects. Remarkably, we were not able to rescue the long period phenotype of DOM-B knockdown with overexpression of dom-A (Fig. 3C). Similarly, neither was overexpression of dom-B able to rescue the arrhythmic phenotype of DOM-A knockdown, although we did notice that the period-lengthening effect was partially rescued (Fig. 3C).
Since DOM is a chromatin-remodeling protein, and since we observed lower abundance of both per and tim mRNA, we therefore examined whether DOM might be bound to the promoters of per and tim. We used the DOM-A- and DOM-B-specific antibodies to perform chromatin immunoprecipitation (ChIP) at two time points: ZT4 and ZT16, close to the trough and peak time of CLK binding [37]. We observed significant enrichments of DOM-A and DOM-B binding on both per and tim promoters, compared to an intergenic region control (Fig. 3D). However, we did not observe significant difference in binding between ZT4 and ZT16 (Fig. 3D), which indicates that the binding of DOM-A and DOM-B might not be time-dependent.
DOM-A, but not DOM-B, is required for the maintenance of sLNvs and PDF abundance
The blunted morning activity peak and arrhythmia in DOM and DOM-A knockdown flies suggest that there might be defects in the sLNvs or in PDF signaling. There are three major projections of PDF positive LNvs. The lLNvs send projections to the optic lobes and the contralateral brain hemisphere, while the sLNvs send projections to the DN1s and DN2s region [38]. Based on whole mount immunohistochemistry in fly brains, we did not observe obvious defects in the brain structure or projections of the lLNvs, however, PDF levels in the dorsal projections of the sLNvs were barely detectable in DOM and DOM-A knockdown flies (Fig. 4A). Absence of the dorsal projection or low PDF expression in the sLNvs could lead to decrease of PDF in the dorsal projection [19,39]. Our data are consistent with both possibilities. Using pdf-GAL4 and a membrane tethered GFP (CD8-GFP) to label axons of sLNvs, we observed that the dorsal projections of sLNvs were clearly shortened in DOM and DOM-A but not in DOM-B downregulation (Appendix Fig S5). Furthermore, close observation of the PDF positive sLNvs revealed that both the number of sLNvs and PDF levels were reduced (Fig. 4B, 4C, and Appendix Fig S5). For each hemisphere, the average number of PDF positive sLNvs in DOM and DOM-A knockdown flies was approximately 2, compared to 4 in the control flies (Fig. 4C). However, with DOM-B knockdown, we did not observe any obvious loss of sLNvs or reduction in PDF levels, which indicates that this process is specifically controlled by DOM-A (Fig. 4B, 4C). Consistent with the ChIP results, the levels of PER and TIM were found to be significantly decreased in the sLNvs with knockdown of DOM-A or DOM-B (Fig. 4B, 4C). This may also explain why depletion of DOM-A or DOM-B had an effect on period-lengthening.

A. Representative confocal images showing PDF expression in whole brain and dorsal axonal projection of sLNvs. Brains were dissected at ZT1 for anti-PDF (green) and anti-PER antibodies (red). Comparing to the control (Top left panel), dorsal projection of sLNvs was disrupted in dom (Top right panel) and domA downregulation (Bottom left panel) no effects were observed in the presence of domB RNAi (Bottom right panel) (Scale bar: left, 500 um; right, 100 um).
B. Representative images of sLNvs of associated lines. Brains were dissected at ZT1 for anti-PDF (green) and anti-PER antibodies (red) or dissected at ZT23 for anti-PDF (green) and anti-TIM antibodies (red). Cell number of sLNvs (marked as square brackets), PDF, PER and TIM levels in sLNvs were decreased in dom and domA downregulation flies brains (Top right and bottom left panels), while only affected PER and TIM levels in domB downregulation flies (Bottom right panel). (Scale bar: 50 um).
C. Quantification of sLNvs numbers in each brain hemisphere, as well as PDF, PER and TIM levels in sLNvs. n=32 hemispheres was used for quantification.
D. Pdf transcriptional level in lLNvs and sLNvs using TOMATO fluorescence signal. Pdf transcriptional levels in sLNvs (marked as circle) were decreased in dom and domA downregulation fly brains (middle two panels), while sLNv numbers and pdf transcriptional levels in domB downregulation flies were normal (right panel) (Scale bar: 100 um.).
E. Quantification of sLNvs numbers and TOMATO fluorescence signal in sLNvs and lLNvs. n=24 hemispheres were used. Error bars correspond to SEM. **p < 0.01,***p < 0.001,****p < 0.0001 as determined by t test.
Next we examined which step of regulation causes the reduction of PDF levels in DOM and DOM-A knockdown. DOM regulates gene expression at the transcription level [22]. We therefore examined the expression of pdf using a transcriptional reporter line, pdfTomato [20]. Consistent with PDF staining, we observed significant reduction of pdf transcription in sLNvs of both DOM and DOM-A knockdown (Fig. 4D, 4E). Interestingly, the decrease in neuron number and reduction of PDF level were unique to sLNvs, which were not detected in the lLNvs (Fig. 4D, 4E).
In summary, consistent with differences in circadian behavior, DOM-A and DOM-B knockdown also exhibit differences at the molecular level. Knockdown of DOM-A causes loss of sLNvs and decrease in sLNv PDF levels, which is not observed in DOM-B downregulation.
DOM is required during development and adulthood for circadian rhythms
Decrease of sLNvs numbers and dorsal projections in adult flies might be due to abnormal development or maintenance defects. Therefore, we performed brain dissections at larval stage and pupal stage. The number of sLNvs precursor in the DOM knockdown condition was unchanged compared to the control at 3rd instar larvae (Fig. 5A, 5B), which suggests that DOM does not affect sLNvs development. However, same as adulthood, even at early pupal stage (3 days after pupation), there were only 1-2 sLNvs detected in dom downregulation (Fig. 5C, 5D). These data indicate that DOM is required for the maintenance but not for the development of sLNvs. DOM probably affects sLNvs during larval-pupal metamorphosis. Consistent with adult flies, both PDF and PER levels of dom downregulation were reduced in sLNvs compared to the control in larvae or pupae (Fig. 5B, 5D).

A. Representative larva brain confocal images. L3 stage larvae collected at ZT1 were immunostained with PDF and PER antibodies. Silence the expression of dom in larvae stage affected PDF and PER levels in sLNvs (marked as circle), while number of sLNvs were not affected (Scale bar: left, 200 um; right, 50 um.).
B. Quantification of larva sLNvs, as well as PDF and PER levels. For each genotype, ∼ 22 flies brains and ∼85 neurons were used for quantification.
C. Representative pupal brain confocal images. Flies pupa (3 days after pupation) were collected at ZT1 were immunostained with PDF and PER antibodies. Silence the expression of dom in pupal stage affected number of sLNvs, PDF and PER levels in sLNvs (marked as circle) (Scale bar: left, 500 um; right, 50 um).
D. Quantification of pupal sLNvs, as well as PDF and PER levels. For each genotype, ∼ 25 flies brains and ∼100 neurons were used for quantification.
E. Free-running period and percentage of rhythmicity of DOM, DOM-A and DOM-B depleted flies in adulthood (left part) or during development (right). Stage specific silencing the was done by using the conditional tim-Gal4;tub-Gal80ts driver system.
F. Representative confocal images of fly brain showing projections of PDF positive LNvs. Flies were grown at 18°C until eclosion, and adult flies were entrained for 5 days in LD 30°C. Brains were dissected at ZT1 for anti-PDF (green) and anti-PER antibodies (red) (Scale bar: left, 500 um; right, 50 um).
G. Quantification of the numbers of sLNvs and PDF, PER levels in sLNvs of associated lines. Error bars correspond to SEM. *P < 0.05,***p < 0.001, ****p < 0.0001 as determined by the t-test.
The decrease of axonal projections and number of sLNvs we observed in DOM knockdown flies (Fig. 4A, 4B) suggested that DOM might play a role during development to properly regulate circadian rhythms. Thus, we used tim-GAL4 and GAL80ts TARGET system to temporally express dom dsRNA during development or after eclosion [40]. 18 °C prevents dsRNA production, while 30°C allows expression of dsRNA thus downregulation of DOM. We observed strong rhythmicity but around 1-2 hr period-lengthening with adult-specific DOM or DOM-A knockdown, compared to high arrhythmic activity for knockdown during development (Fig. 5E and Appendix Table S1). Thus, developmental expression of dom or dom-A dsRNA appears to affect the rhythmicity, while adult specific depletion appears to be sufficient for period-lengthening.
Furthermore, knockdown of DOM-B only in adulthood caused a ∼1 hr increase in period length, suggesting that unlike DOM-A, the DOM-B splice form is mainly necessary for the regulation of circadian clocks in adult stage (Fig. 5E and Appendix Table S1). We observed a slight but significant lengthening of period for DOM-B developmental depletion, which indicates that DOM-B might also play a role during development.
To exclude anatomical defects in the circadian circuitry, we performed brain dissections in the flies with adult-specific DOM downregulation. As expected, the dorsal projections and number of sLNvs were normal, and PDF level was not affected (Fig. 5F, 5G). These data further suggest that the arrhythmia we observed in DOM knockdown was mainly due to the function of DOM during development. However, the abundance of PER in sLNvs was still reduced, which could explain the lengthened period phenotype (Fig. 5G). Taken together, these data indicate that DOM-A is required during development for the rhythmic activity, while both DOM-A and DOM-B are necessary for period determination post development.
Activation of PDFR signaling in circadian neurons rescues the behavioral phenotype of DOM depletion
Since we observed low PDF levels in the sLNvs with DOM knockdown, we wondered whether the decrease of PDF signaling was the cause of arrhythmia. To test this hypothesis, we determined whether hyperactivation of PDFR could rescue the phenotypes that accompany reduction in DOM expression. To increase PDFR signaling, we expressed a membrane-tethered PDF (t-PDF), which has been shown to mimic high PDF levels [41]. A scrambled peptide sequence (t-PDF SCR) with similar length as PDF was used as a negative control. Strikingly, coexpression of t-PDF restored the rhythmicity to 89% for dom RNAi (71% for another independent dom RNAi line, Fig. 6A, 6B), whereas the t-PDF SCR control was unable to restore arrhythmicity (Fig. 6A, 6B). Interestingly, not only the rhythmicity, but also the period of DOM knockdown was rescued with coexpression of t-PDF (Fig. 6B). We then stained fly brains with PDF and pacemaker proteins PER and TIM (Fig. 6C and Appendix Fig S6). With expression of t-PDF in DOM knockdown flies, we made two interesting observations. First, PDF level as well as the dorsal projection of sLNvs were restored, while the number of sLNvs was still lower (Fig. 6C, 6D). Second, the abundance of PER and TIM in sLNvs was also rescued (Fig. 6C, 6D and Appendix Fig S6). This might be due to the fact that PDF stabilizes PER and TIM [16,17].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A. Actograms showing the average activities on the last day of LD and during 5 days in DD. Light represents the day and gray darkness. From left to right: (Left panel) flies expressing the membrane-tethered scrambled PDF; (middle panel) domRNAi#1 flies expressing a membrane-tethered scrambled PDF (negative control); and (right panel) domRNAi#1 flies expressing the membrane-tethered PDF. Arrhythmia and long period of dom downregulation are rescued with tethered PDF.
B. Free-running period of dom RNAi flies expressing the membrane-tethered PDF.
C. Representative confocal images of brains of dom RNAi flies expressing the membrane-tethered PDF or scrambled PDF. Flies were entrained for 4 days in LD 25°C, and brains were dissected at ZT1 for anti-PDF antibody (green) and anti-PER antibody (red). From top to bottom: (Top panel) fly brain expressing the membrane-tethered scrambled PDF; (middle panel) domRNAi#1 flies expressing a membrane-tethered scrambled PDF; and (bottom panel) domRNAi#1 flies expressing the membrane-tethered PDF. Confocal images are whole brain, dorsal projection and soma of sLNvs from left to the right (Scale bar: whole brain, 500 um; projection, 100 um; sLNvs, 50 um).
D. Quantification of the number and relative PDF and PER levels of sLNvs. For each genotype, totally, 20-25 flies brains and 50-80 neurons were used for quantification of the staining. Error bars correspond to SEM. **p < 0.01, ****p < 0.0001 as determined by t-test.
Discussion
Here, we identify that two alternatively spliced variants of DOM play distinct roles in circadian rhythms. DOM-A is critical for maintenance of sLNvs and PDF abundance during development thus controls morning anticipatory activity and circadian rhythmicity; while DOM-B specifically determines circadian period of locomotor activity by regulating core pacemaker protein PER/TIM abundance.
The abolished morning anticipatory activity in light dark cycle and low rhythmicity under constant darkness in DOM/DOM-A knockdown flies is reminiscent of the phenotypes seen in pdf mutant (Fig.1 and Fig.3) [42]. Based on these observations, we hypothesized that PDF signaling is disrupted with dom or dom-A RNAi. Indeed, we found that PDF abundance was decreased both in the soma and dorsal axonal projections of sLNvs (Fig. 4). Furthermore, by restricting RNAi to adulthood, we rescued the PDF expression and projection, which also restored the rhythms (Fig. 5). These results suggest that the arrhythmic phenotype is due to the decrease of PDF levels in dom or dom-A downregulation. Despite the importance of sLNvs in circadian rhythms, the mechanism underlying sLNvs maintenance is still unclear. Here we find that DOM-A plays important roles in the maintenance of sLNvs. PDF positive sLNvs and dorsal projections are formed in first instar larvae [10]. During development, both DOM-A and DOM-B start to be expressed in embryos [23]. Our data suggest that DOM-A does not affect sLNvs development in larvae, but rather regulates sLNvs maintenance at later stages (Fig. 5). Knockdown of DOM-A may trigger apoptosis or necrosis pathways and cause programmed cell death in sLNvs. In fact, chromatin-modifying pathway has been shown to regulate neuronal necrosis in flies [43]. In the future, it will be interesting to examine whether DOM-A is involved these programmed cell death pathways and blocking necrosis or apoptosis pathway can restore the sLNvs in DOM downregulation.
Here we found that depletion of DOM-A seems to specifically affect the maintenance of PDF positive sLNvs and PDF accumulation in sLNvs, while the PDF positive lLNvs is not affected (Fig. 4). This suggests that as master pacemaker neurons, sLNvs may have some unique regulatory program for maintenance and gene expression, which requires DOM-A. Interestingly, a phosphatase LAR has recently been found to specifically regulate PDF expression in the dorsal projection of sLNvs during development [39]. The pacemaker protein VRI also played important roles in controlling PDF abundance and dorsal arborization rhythms of sLNvs [21]. Future efforts to identify specific regulatory mechanisms in sLNvs will help us understand the maintenance and function of circadian neuronal network.
Why does DOM-A, but not DOM-B, regulate sLNv maintenance and pdf transcription? One possibility might be that DOM-A and DOM-B are associated with different protein complexes. Compared to DOM-B, DOM-A has two unique domains in the C-terminus: a SANT domain and a poly-Q domain. Both of these two domains are known to mediate protein interactions. Interestingly, only DOM-A is identified to bind the Tip60 complex when Tip60 was purified by a tagged protein subunit from Drosophila S2 cells [22]. A previous study finds that depletion of Tip60 or overexpression of a histone acetyltransferase-defective Tip60 decreases axonal growth of the sLNvs in the fly model of Alzheimer’s disease [44]. These results suggest that DOM-A might interact with Tip60 to control the axonal projection and PDF expression of sLNvs.
Lastly, how does DOM control the length of circadian period? This is probably through regulation of PER and TIM abundance. Interestingly, compared to the decrease of per and tim mRNA in DOM downregulation, the master circadian transcription factor clk mRNA is not affected. Does DOM control PER and TIM accumulation through transcriptional regulation? Indeed, we found that from Drosophila S2 cell affinity pull-down, CLK interacts with DOM complex (Table 1). Furthermore, DOM-A and DOM-B binding is enriched in the E-box region of per and tim promoters. Even though no binding differences were observed in DOM-A and DOM-B at ZT4 and ZT16 (Fig. 3), it is possible that other subunits in the DOM complex bind rhythmically, leading to rhythmic activity of the complex. Given that the conserved functions of DOM and human SRCAP/p400 in Notch signaling and histone variant exchange, it is possible that similar mechanisms are leveraged to control circadian rhythms in mammals.
Materials and Methods
Fly Stocks
All the flies were raised on cornmeal/agar medium at 25°C under a LD cycle. The following strains were used: yw, w1118, yw; tim-GAL4/CyO [35], yw;Pdf-GAL4/CyO [11]; yw;tim-GAL4/CyO;Pdf-GAL80/TM6B [45], tublin-GAL80ts, UAS-cd8-GFP, clkout. The following stocks were obtained from the Bloomington Drosophila Stock Center (http://flystocks.bio.indiana.edu/): UAS-domRNAi#1 (BL31054), UAS-domRNAi#2 (BL38941), UAS-domA (BL64261) and UAS-domB (BL64263). DomARNAi (sh-domA) and domBRNAi (sh-domB) fly lines were generous gifts from Dr. Peter B. Becker. PdfTomato line was generated by Dr. Sebastian Kadener.
Behavioral Experiments and Analysis
For behavioral experiments, adult male flies (2-4 days old) were used for testing locomotor activity rhythms. Flies were entrained for 4 full days LD cycle at 25°C, using about 500 lux light intensities, and then released into DD at 25°C for at least 6 days. Locomotor activity was measured with TriKinetics Activity Monitors in I36-LL Percival Incubators. Locomotor activity was averaged over the 4 days entrainment for LD and 6 days for DD. Analyses of period and power were carried out using FaasX software as previously described [46]. Actograms were generated using a signal-processing toolbox implemented in MATLAB (MathWorks) [47]. For GAL80ts experiments, flies were raised at 18°C and tested at 30°C. They were entrained for 5 days and then released in DD for at least 6 days. Morning anticipation was calculated by the ratio of activity counts between 2 hrs before light on and 6 hrs before light on. We first measured the single fly activity counts obtained in twelve 30-min bins between Zeitgeber Time (ZT) 17.5 and ZT24 (6 hr before lights on) and six 30-min bins between ZT22.5 and ZT24 (2 hrs before lights on). The first value is divided by the second to obtain the morning activity index. Morning anticipations of individual flies were then averaged and plotted on the graphs.
Whole-Mount Immunohistochemistry
Whole-mount immunohistochemistry for fly brains were done as previously described [48]. Adult fly (3-6 days old) or L3 stage larval brains were dissected in chilled PBT (PBS with 0.1% Triton X-100) at the indicated time points and fixed in 4% formaldehyde diluted in PBS for 30 min at room temperature. For pupal brains dissection, Drosophila were fully developed and hatched 7 days after pupation in the same cross, pupa were collected 3 days after pupation using a small wet paintbrush and transfer to a dissection dish. After 2-3 times PBS wash, the pupal brains were dissected and fixed as previously described method. The brains were rinsed and washed with PBT three times (10 min each). Then, brains were incubated with 10% normal Goat serum diluted in PBT to block for 60 min at room temperature and incubated with primary antibodies at 4°C overnight. For PER, TIM and CLK staining, we used 1:1,500 rabbit anti-PER, 1:2,000 rat anti-TIM (gift from Dr. Rosbash) and guinea pig 1:2,500 anti-CLK (gift from Dr. Hardin), respectively. We used a 1:200 dilutions for mouse anti-PDF and 1:200 for rabbit anti-GFP (DSHB). After six washes with PBT (20 min each), brains were incubated with relative secondary antibody at 4°C overnight, followed by another six washes with PBT. For PdfTomato staining, flies were directly dissected in chilled PBT at the indicated time points and moutained in the medium (VECTOR). All samples were imaged on a Leica Confocal SP8 system, with laser settings kept constant within each experiment. 10 to 12 fly brains for each genotype were dissected for imaging. Representative images are shown. Image-J software (National Institutes of Health [NIH]) was used for PER, PDF, TIM and CLK quantification in 15-30 sLNvs from at least seven brains. For quantification, signal intensity in each sLNv and average signals in three neighboring non-circadian neurons were measured, and the ratio between signals in sLNvs and non-circadian neurons was calculated.
Chromatin immunoprecipitation (ChIP)
Chromatin immunoprecipitation (ChIP) was performed based on published protocols [49]. Flies entrained in 12 hr light:12 hr dark (LD) conditions at 25°C for four days were collected at two time-points (ZT) on the fifth day. Briefly, chromatin was isolated from 500 μl of fly heads homogenized with a dounce homogenizer (Kimble Chase) for 20 strokes using the loose “A” pestle. Homogenate was sieved by a 70 μm cell strainer (Falcon) then centrifuged to remove cell debris. Pellets were cross-linked using formaldehyde. Samples were sonicated using a focused-ultrasonicator (Covaris M220) on setting for 400-500 bp cDNA and then centrifuged at 10,000 rpm for 10 minutes. Supernatant was collected in two 130 μl aliquots for IP and 26 μl was collected for input and frozen at −80C for analysis. Sonicated chromatin was roughly 500 bp in length (<1000 bp). For each IP, 30 μl of a Protein G Dynabead slurry (Life Technologies) was washed then incubated along with the appropriate antibodies for 4 hours at 4°C with rotation. Amount of antibodies used for ChIP is as follows: anti-DOM-A (20 μg/ml, rabbit, GenScript) which was generated in our lab (antigen protein sequence designed according to 2008-2349 aa of DOM-A), anti-rabbit-IgG (20 μg/ml, Life Technologies), anti-DOM-B (10 μg/ml, mouse, from Dr. Peter B. Becker), anti-mouse-IgG (10 μg/ml, Life Technologies). Following incubation, beads were collected and incubated with chromatin overnight at 4°C with rotation. DNA was eluted using the Qiagen PCR purification kit and subjected to qPCR. At least three technical replicates of qPCR were performed for each biological ChIP replicate and three biological replicates were performed for DOM-A and DOM-B assays. Background binding to a nonspecific antibody (anti-IgG; Life Technologies) bound to Dynabeads was subtracted from input samples and results are presented as the percentage of the input samples. For each assay, at least three biological replicates were performed. The specific primers used for qPCR are described in Appendix Table S2. The technical qPCR triplicates were averaged for each biological replicate as no significant differences were found between the technical replicates, and the error bars represent SEM calculated from variance between biological replicates. Two-tailed t-tests were used to determine statistical differences between control and experimental treatment at each ZT.
Real time quantitative reverse transcription PCR
Flies were collected at the indicated time points and isolated heads were stored at −80°C. Total RNA was extracted from 25–30 heads with TRIzol based on the manufactures protocol (Life Technologies, USA). A 2-µg quantity of RNA was reverse-transcribed with reverse transcription reagents (Invitrogen). For real-time PCR (qRT-PCR) of per, tim, clk, dom, domA, domB and actin, we used a qPCR detection kit (SYBR Select Master Mix For CFX) (Life Technologies, USA). The specific primers used for qPCR are described in Appendix Table S2. All the experiments were performed in the CFX96 Real-Time System (BIO-RAD).
Western blot analysis
Fly heads were collected at the indicated time points and homogenized with pestles, protein extracts were prepared with HEPES-Triton lysis buffer (1X HEPES-Triton buffer, 1 mM DTT, 0.4%NP-40, 0.1% SDS, 10% glycerol, 1X tablet protease inhibitor). Proteins were quantified using BSA assay. For immunoblot analysis, proteins were transferred to PVDF membranes (Genesee Scientific) and incubated with anti-DOM-A (1:300, GenScript), anti-DOM-B (1:5, from Dr. Peter B. Becker) and anti-ACTIN (1:100,DSHB) in blocking solution. Band intensity was calculated and analysed with Image J.
Statistics analysis
Statistical analysis of two data points was performed with Student’s t-test. Statistical analysis of multiple data points was performed with one-way analysis of variance with Tukey post-hoc tests using GraphPad software.
Author Contributions
YZ, JCC and ZL formulated the ideas and designed the experiments. ZL and VHL performed the experiments. YZ, JCC and ZL wrote the manuscript.
Conflict of interest
There is no competing financial interest for the authors.
Acknowledgements
We would like to thank Drs. Luoying Zhang, Patrick Emery, Pedro Miura, and Alexander M. van der Linden for comments and discussions on this manuscript. We thank all the members in the Zhang lab for technical support and helpful discussions. We thank Dr. Becker for the fly strains of domA and domB shRNA, as well as the DOM antibodies. We thank the Bloomington stock center for various fly stocks. We are grateful for TIM and PER antibodies kindly given by Dr. Rosbash and Dr. Stanewsky. We also thank the Developmental Studies Hybridoma Bank for PDF and GFP antibodies. This work was initiated in Dr. Patrick Emery’s lab. This work is supported by the National Institute of General Medical Sciences of the National Institutes of Health under grant numbers P20 GM103650, GM103554, and GM103440, and R01 GM102225 to JCC.
References




