

ABSTRACT
Tau is a neuronal, microtubule-associated protein responsible for controlling the stabilization of microtubules in neurons. Tau function is regulated by phosphorylation, but in some neurological diseases it becomes aberrantly hyperphosphorylated, thus contributing to the pathogenesis of several neurological diseases such as Alzheimer’s disease. Western Blot (WB) has widely been employed in the assessment of Tau levels and fragmentation in neurological disease models. However, quantification of Tau levels/cleavage by WB should be interpreted with care, as this approach has been recognized as a complex, multi-step technique prone to produce artifactual results if not properly performed. Here, we evaluated the influence of freeze-and-thaw, a common storage procedure preceding WB, to the integrity of Tau in rat brain extracts. We observed that samples submitted to freeze-and-thaw presented a ~25 kDa fragment shown to be derived from Tau. Based on this observation, we strongly recommend that molecular analysis of Tau levels in rodent brain samples in future investigations be performed using fresh extracts. Finally, we discuss the impact of our findings to the translational power of studies involving Tau-induced neurodegeneration.
INTRODUCTION
Tau is a neuronal, microtubule-associated protein (MAP) responsible for controlling the stabilization of microtubules in neurons, thereby impacting the coordination of the axoplasmic transport of organelles, proteins, lipids, synaptic vesicles and other important cargos along the neuron [1, 2]. Tau function is regulated by phosphorylation, but in some neurological diseases it is found aberrantly hyperphosphorylated [2, 3]. Due to the strong correlation between high levels of phosphorylated Tau (pTau), aggregation, microtubule destabilization and neuronal dysfunction/death, elevated pTau levels have been assigned as a hallmark of several neuropathologies, collectively known as tauopathies, such as Alzheimer’s and Pick’s disease [3-6]. In addition, it has also been shown that increased production of Tau protein fragments as a result of enzymatic cleavage also correlates with neurodegeneration in cellular models [7] and, more importantly, with memory deficits in animal models [8].
Western Blotting (WB) has been widely employed in the assessment of Tau protein levels (either total or hyperphosphorylated), as well as in the analysis of Tau fragmentation in a number of studies [8-11]. Quantification of Tau levels/cleavage by WB should be interpreted with care, as this approach has been recognized as a complex, multi-step technique that requires case-to-case standardization and is subjected to produce artifactual results if not performed properly [12, 13]. One critical care is the integrity of the protein of interest in the sample, since unsuitable conditions for collection, storage and/or handling may damage the biological material and lead to conclusions that do not accurately reflect the biological phenomenon under investigation [12, 14]. This issue is particularly relevant in studies comprising the analysis of a large set of samples, in which storage by freezing is common.
The goal of the present study was to demonstrate the influence of freeze-and-thaw prior to WB analysis on the integrity of Tau in rat brain extracts. We observed that samples submitted to freeze-and-thaw presented fragmentation of endogenous Tau protein, as seen by the presence of an extra ~25 kDa band simultaneously to the weakening of the bands corresponding to full-length Tau in extracts of different cerebral regions. Based on this finding, we recommend that molecular analysis of Tau levels in rodent brain extracts should be carried out using fresh extracts, in order to produce results that better reflect the investigated biological phenomenon.
MATERIAL AND METHODS
Animals
All procedures were approved by the Ethics Committee on Animal Use of Ribeirão Preto Medical School (CEUA-FMRP), under the protocol #017/2014-1. All experiments were carried out using 12 month-old animals (Rattus norvegicus), both control Wistar (n=7) and Wistar Audiogenic Rat (WAR) (n=9) strains [15]. Control Wistar rats were obtained from the Central Vivarium of the University of São Paulo (USP) at Ribeirão Preto and kept at 25° C, 12h/12h photoperiod and water and food ad libitum. WAR animals were obtained from the Vivarium of the Department of Physiology of the USP Ribeirão Preto Medical School and kept at the same conditions.
Western Blotting
Animals were anaesthetized by inhalation of isoflurane (BioChimico, Brazil) and euthanized by decapitation. Cerebral tissue was collected, dissected and stored in 1.5 mL microtubes at -80° C. Homogenates from dorsal portion of hippocampus and frontal cortex were prepared according to Petry et al. [16]. Tissue fragments were homogenized in ice-cold RIPA buffer (50 mM Tris, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100 and 0.1% SDS, pH 7.5) supplemented with protease inhibitor (1:100, Sigma) and phosphatase inhibitor (10 mM NaF, 10 mM Na3VO4) cocktails at the ratio of 10 μL per mg of tissue using an electric potter (Kimble Chase) in 3 cycles of 10 seconds each. Extracts were centrifuged for 10 minutes at 10000 x g and 4°C. Supernatant was transferred to new microtubes and aliquots were readily used for analysis of Tau protein by WB (fresh extract). Alternatively, parts of the extracts were stored at -20°C for at least 2 weeks and then thawed just before WB (freeze/thaw extract). Preparation of samples for SDS-PAGE included addition of protein loading buffer (0.0625 M Tris-Cl, 2% SDS, 10% v/v glycerol, 0.1 M dithiothreitol, 0.01% bromophenol blue, pH 6.8) to extracts and boiling for 5 minutes at 100°C. The SDS-PAGE was carried out using 12% acrylamide/bis-acrylamide gels and a constant voltage of 90V. For WB, 0.45 μm nitrocellulose membranes (GE Healthcare Life Sciences) were utilized. Membranes were blocked in 5% nonfat dry milk/T-TBS (Tris-buffered saline: 20 mM Tris pH 7,5, 150 nM NaCl, 0,1% Tween 20) solution for 1 h at 25 °C and subsequently incubated with primary antibody overnight at 4°C. Primary antibodies used for Tau assessment were anti-Tau phospho S396 (Abcam) or anti-Tau [E178] (Abcam), both 1:1000 in 5% BSA/T-TBS solution. After washing, membranes were incubated with secondary antibody ECL anti-rabbit IgG HRP (Amersham) at 1:3000 in 5% BSA/T-TBS for 1 h at 25 °C. The membranes were revealed using ECL Prime Western Blotting Detection Reagent (GE Healthcare Life Sciences) and imaged in ChemiDoc imaging system (Bio-Rad) coupled to a digital system and software ImageQuant™ 3.5 (GE Healthcare Life Sciences). For β-actin probing, membranes were stripped and incubated with anti-β-actin (EMD Millipore) at 1:20000 in 3% BSA/T-TBS for 1 h at 25 βC.
In-gel digestion and LC-MS/MS analysis
Aliquots (250 μg total protein) of brain extracts subjected to freeze/thaw were treated with 5 μL of dithiothreitol (50 μg/μL, diluted in ammonium bicarbonate 100 mM, pH 7.8) for 30 min at 37°C. DTT-treated samples were then diluted in loading buffer, boiled for 5 min at 100°C, and alkylated in the dark for 30 min at 37°C with 1250 μg of iodoacetamide (50 μg/μL diluted in ammonium bicarbonate 100 mM, pH 7.8). After sample separation by SDSPAGE (carried out using 12% acrylamide/bis-acrylamide gels and a constant voltage of 90 V), a piece of ca. 30 mm2 of the unstained gel (per lane) was cut off using as a reference the 25 kDa standard band (pre-stained). Gel pieces corresponding to the fragment were further sliced to 1-4 mm2 pieces, washed and digested with trypsin as described by Grassi et al. (2017). Tryptic peptides were successively extracted with 5% formic acid/50% acetonitrile, and then 90% acetonitrile, and dried in a vacuum concentrator. The sample was re-suspended in 50 μL 50% acetonitrile/5% formic acid and centrifuged at 12000 xg for 15 min at 25 °C. The supernatant was injected into a LC-MS/MS Xevo TQS system (Waters). Chromatographic separation was performed in a UPLC (I-class, Waters) using a C18 column (1.8 μm particle size, 100 Å pore size, 1 mm × 150 mm, Waters) in a linear gradient of 5 to 30% acetonitrile over 15 min at 100 μL/min in a formic acid:acetonitrile:water solvent system. Detection of Tau theoretical tryptic peptides was programmed in a scheduled multiple reaction monitoring method using 3–5 transitions per peptide and monitoring windows of 2 min centered on predicted retention time. Both method development and data analysis were conducted using Skyline software [17].
RESULTS
Presence of an anti-Tau-immunoreactive ~25 kDa fragment in brain extracts submitted to freeze-and-thaw
Tau hyperphosphorylation is known to be associated with neurodegeneration and dementia [2, 4]. With the purpose of investigating a possible involvement of Tau hyperphosphorylation in a cohort of aged animals presenting early cognitive decline (data to be published elsewhere), we initially prepared hippocampal extracts from 12-month old rats (both Wistar and WAR strains) and evaluated hyperphosphorylated and total Tau protein levels after storing these extracts at -20°C. Unexpectedly, in addition to the typical profile of three major bands migrating between 50 and 75 kDa (Fig. 1A), which are known to correspond to different isoforms of Tau expressed in rodent brains [18], we found an extra band of ~25 kDa in all samples tested in both phospho and total Tau-revealed membranes. The ~25 kDa band was also observed in freeze-and-thaw extracts from frontal cortex (Fig. 1B). Recently, Zhao et al. [8] reported the presence of a ~35 kDa (TCP35) band in transgenic mice brains expressing human Tau, which was correlated with cognitive decline and identified as a proteolytic product of Tau. Motivated by those findings, and considering that freeze-and-thawing may impact the stability of several proteins [19-22], we wondered whether this ~25 kDa fragment observed under our conditions was endogenously produced or a product of freeze-and-thaw-induced degradation. Interestingly, when the WB was conducted using fresh samples, the ~25 kDa band was significantly weaker or even completely absent in both hippocampal (Fig. 1C) and frontal cortex (Fig. 1D) extracts. Importantly, the bands corresponding to full-length Tau were weaker in the samples in which the ~25 kDa band was present, supporting the notion that it indeed represents a degradation product of Tau generated by freezing-and-thawing the extracts. This phenomenon seems to be specific for Tau, since no differences between fresh and freeze/thaw extracts regarding integrity of β-actin in the same samples were observed (Fig. 1E). The freeze-and-thaw-induced Tau fragmentation was detected at similar degrees in brain extracts of both control Wistar and WAR strains, indicating that this phenomenon was strain-independent.

Representative images of WB probing for both phosphorylated (pTau) and total Tau protein, as indicated. Either freeze-and-thaw (F/T; A, B) or fresh (C, D) extracts from dorsal hippocampus (A, C) or frontal cortex (B, D) were analyzed. Some extracts were additionally probed for β-actin (E). The arrow-head indicates the presence of a band near 25 kDa in both brain regions, when either phosphorylated or total Tau were probed, which is faint or completely absent in fresh samples from the same brain regions of the same animals. Some samples (A: lanes 1-3, 9, 10 and 12; B: lanes 2-5; C: lanes 3 and 6) were obtained from animals of the WAR strain. All the remaining samples were obtained from control Wistar rats.
Identification of the ~25 kDa species associated to freeze-and-thaw as a degradation product of Tau
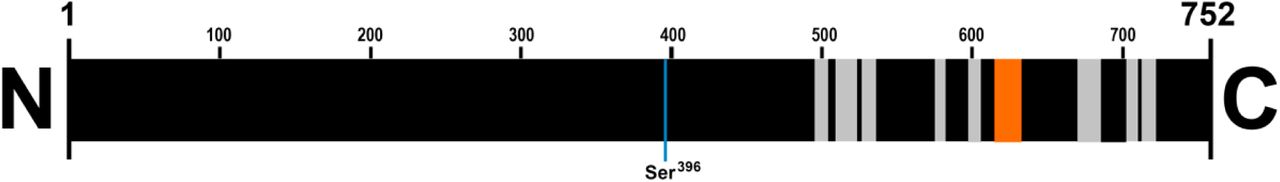
In order to verify whether freeze-and-thawing caused Tau fragmentation, we aimed to biochemically identify the freeze-and-thaw-induced ~25 kDa species by mass spectrometry (MS). For this sake, proteins from hippocampal extract submitted to freeze-and-thaw were separated by SDS-PAGE, a piece of the gel in the range of 25 kDa was cutoff, subjected to trypsin digestion, and eluted peptides were analyzed by target protein MS. Nine peptides in full-length rat Tau sequence were identified, one of those being a unique peptide (Table I). This finding confirms that the ~25 kDa band showing immunoreactivity to anti-Tau antibodies is indeed a degradation product of Tau. Since all of the Tau-associated peptides identified were located near the C-terminus (Fig. 2), it is possible to conclude that cleavage occurred between the N-terminus and residue Ser396, considering that the ~25 kDa fragment is immunodetected by both anti-Tau phospho S396 and anti-Tau E178 (binds to an epitope localized near C-terminus of protein) antibodies. Assuming that the cleavage occurs near to Ser396, the predicted molecular weight of the identified fragment would be ~35 kDa. The precise reason for the ~10 kDa discrepancy between apparent and predicted molecular weight is not yet clear, although inaccuracies in MW determination by SDS-PAGE are often reported (e.g. [23]). It is also important to consider that Tau mRNA alternative splicing generates several isoforms of rat Tau (uniprot.org/uniprot/P19332) and this may implicate in MW prediction errors [24]. Further analyses are necessary in order to determine which isoform has been cleaved in freeze-and-thaw process to generate the ~25 kDa fragment we found.
Tryptic peptides identified by targeted mass spectrometry.
{kind=link}
{kind=link}
All the peptides identified by mass spectrometry (in gray, a unique peptide is highlighted in orange) are located near to the C-terminus of full length Tau from rat. Residue numbers are indicated.
Interestingly, the putative cleavage region identified in this study is close to a proteolysis site for caspase, mapped in human Tau isoform 0N4R (amino acid sequence: HVLGGGSVQIVYKPVD) by Zhao et al. [8]. It is thus possible that this region, which is partially conserved and present in all isoforms of rodent Tau (uniprot.org/uniprot/P19332), represents a proteolysis-prone site in Tau. Indeed, the conformational flexibility of the region surrounding residue Ser396 has been recently demonstrated by the Ser396 phosphorylationinduced α-helix to β-sheet conversion, with possible implications to microtubule binding and aggregation [25].
DISCUSSION
Taken together, our findings clearly indicate that the ~25 kDa Tau fragment is an artifact generated by freezing-and-thawing brains extracts prior to Western Blot, thus highlighting the importance of proper sample handling/storage in the quantitative analysis of Tau (total or phosphorylated) by WB. Unfortunately, in the majority of the studies reporting Tau level analysis by WB, no detailed description on sample preparation and handling prior to WB is depicted. Therefore, it is not possible to rule out that, at least in some of these studies, the reported quantification was not accurate. This is particularly relevant when Tau fragments are correlated with a biological response such as cognitive decline in mice, as in Zhao et al. [8]. In that article, although no description on sample storage is depicted, the authors identified caspase as the enzyme responsible for Tau cleavage in vivo. In contrast, in some studies reporting, for instance, the presence of several Tau fragments in postmortem AD brain extracts (e.g. [26-29]), no detailed description on the sample handling prior to WB is depicted and/or no investigation on the molecular origin of those Tau fragments has been presented. Therefore, it remains to be determined whether those fragments are indeed relevant to neurodegeneration, or in fact correspond to artifacts introduced during the pre-analytical phase.
The huge production of data in contemporary biological sciences has been criticized as a source of unreliable, poorly-translational findings [30, 31]. It is likely that this is in part due to inadequate sample collection, handling and/or storage. The present study intends to emphasize the need of using fresh extracts from rodent brains when Tau levels are to be determined by WB. Considering the wide use of WB to evaluate neurodegeneration in AD and tauopathies, we anticipate that following this recommendation should lead to more accurate interpretations in future studies on Tau-related disorders.
CONCLUSION
Here we report that freezing-and-thawing rat brain extracts before Western Blot led to Tau protein degradation, detected by the presence of an extra band at ~25 kDa in WB analysis with simultaneous weakening of the three major bands corresponding to the fulllength Tau. Based on this observation, we strongly recommend performing molecular analysis of Tau levels in rodent brain samples using fresh extracts.
ACKNOWLEDGEMENTS
This work was supported by grants from Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP), FAEPA and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). We thank Jose Antonio C. de Oliveira and Silvana El-Chedraoui Silva for excellent technical assistance. NGC and VMF hold CNPq Researcher fellowships.
REFERENCES




