

ABSTRACT
Potato virus Y (PVY) is a major potato pathogen that causes annual losses of billions of dollars. Control of its transmission requires extensive use of environmentally damaging insecticides. Rysto confers extreme resistance (ER) to PVY and is a valuable trait in resistance breeding programs. We isolated Rysto using Resistance gene enrichment sequencing (RenSeq) and PacBio SMRT (Pacific Biosciences Single-Molecule Real Time Sequencing). Rysto encodes a nucleotide binding-leucine rich repeat (NLR) protein with an N-terminal TIR domain, and is sufficient for PVY perception and extreme resistance in transgenic potato plants. We investigated the requirements for Rysto-dependent extreme resistance, and showed that Rysto function is temperature-independent and requires EDS1 and NRG1 proteins. Rysto may prove valuable for creating PVY-resistant cultivars of potato and other Solanaceae crops.
Author contributions:
M.G-B., K.W and J.H. performed most of the experimental work, data analyses, and writing; A. W., I.W-F., H.J., K.S. and K.M performed the research; J.J. and W.M. edited the article; J.H. supervised the work and assisted with writing.
INTODUCTION
Potato virus Y (PVY), a member of the Potyvirus genus (Potyviridae family), is the most economically important virus infecting potato crops, with tuber yield and quality losses ranging from 10 to 90 % depending on year, cultivar and location (De Bokx and van der Want 1987; Valkonen 2007). PVY can also infect other Solanaceae species such as bell pepper, tomato and tobacco (Aramburu 2006). The virus is easily transmitted mechanically by over 40 aphid species in a non-persistent manner (Brunt 2001). Intracellular pathogens such as viruses are difficult to control chemically; PVY outbreaks are addressed by the destruction of infected plants or pesticide treatment to limit the population of vectors (Kopp et al. 2015). Resistance breeding remains the best method to prevent viral infections. PVY variants are highly variable at the biological, serological, and molecular levels (Scholthof et al. 2011). They can be classified into different strain groups depending on host response. Virus strains PVY0 and PVYC usually induce mild symptoms of infection, e.g. leaf mosaic lesions, crinkling, leaf drop and dwarfing, while leaf symptoms of PVYN and PVYNW infection are barely noticeable. In addition to severe leaf symptoms, PVYNTN infection leads to potato tuber necrotic ringspot disease which greatly affects tuber marketability (Schubert et al. 2007).
Plant defense responses to viruses are multifaceted and depend on several factors including plant age and tissue type. The most durable control of PVY is provided by resistance genes. In potato, there are two main types of resistance against PVY: extreme resistance (ER), conferred by the Ry genes, and hypersensitive resistance (HR), conferred by the Ny genes (Valkonen 2015). The HR to PVY is usually strain-specific and may result in a range of necrotic reactions in both locally and systemically infected leaves (Valkonen et al. 1998). However, under some conditions HR may be ineffective for restriction of PVY in plants (Vidal et al. 2002). In contrast ER genes are broad-spectrum and confer strong and durable resistance, characterized by lack of visible symptoms after inoculation (Flis et al. 2005).
Resistance genes against PVY infection were introduced into potato cultivars from wild or domesticated Solanum species. Ten genes for resistance to PVY and one gene for resistance to PVA (a potyvirus related to PVY) were mapped to four potato genome segments on chromosomes IV, IX, XI and XII (Valkonen et al., 2017, van Eck et al. 2017). Some of these genes, for instance Rysto and Ry-fsto from S. stoloniferum, were introduced into multiple potato cultivars worldwide and shown to confer durable resistance against multiple PVY strains. Nevertheless, no genes conferring effective HR or ER-type of resistance have previously been isolated (Valkonen et al. 2017).
It has been estimated that controlling major diseases through the informed deployment of resistance genes could contribute over 30% towards crop yield whilst reducing the requirements for chemical applications (Gebhardt and Valkonen 2001). This might be achieved by implementation of new breeding tools to select resistant genotypes and deploy them as varieties (Armstrong et al 2018).
Most plant resistance genes encode intracellular nucleotide-binding, leucine-rich repeat (NLR) receptors, some of which carry an N-terminal TIR domain (TIR-NLRs), and others an N-terminal coiled coil domain (CC-NLRs) (Jones et al., 2016). A useful tool for identification and assembly of full length NLR-encoding genes that cosegregate with resistance is SMRT RenSeq (Witek et al. 2016). The methodology is more robust and cost-effective in monitoring NLR sequences than whole-genome sequencing. All currently known NLRs effective against viruses, nematodes and the late blight pathogen Phytophthora infestans can be tracked with SMRT RenSeq in potato and their polymorphisms defined (Armstrong et al 2018).
There is a high demand for strong and durable genes against PVY infection and R genes conferring ER to PVY fulfil this requirement. As the understanding of mechanisms that underlie ER response to viruses is still incomplete it is of key importance to identify the network of factors that contribute to effective resistance against the pathogen.
In this study, we set out to isolate the Rysto gene conditioning the ER trait using SMRT RenSeq. We found multiple TIR-NLR-encoding paralogs that cosegregate with resistance, and demonstrated that paralog Rysto confers resistance to PVY and to the related potyvirus PVA in different Solanaceae backgrounds. We show that PVY and PVA coat proteins are recognized as avr factors for Rysto triggered immunity. Finally, we investigate the functional relationship between ER and HR, and show that extreme resistance is EDS1, NRG1 but not SA or temperature dependent.
RESULTS
Rysto originates from S. stoloniferum
Extreme resistance to PVY was introgressed to commercial potato cultivars from three different wild Solanum spp. In the European cultivars, resistance originates mostly from S. stoloniferum (Rysto) and is not yet overcome (Valkonen et al. 2017). Therefore, we set out to clone Rysto using dihaploid clone (dH) Alicja, which has PVY resistant S. stoloniferum in its ancestry. We generated diploid mapping population consisting of 391 F1 individuals, and evaluated for resistance to PVY by mechanical and graft inoculations as described in Flis et al. (2005). The segregation ratio of resistant versus susceptible progeny in the mapping population deviated from the 1:1 ratio expected for the segregation of a single dominant gene and was distorted towards resistance (149 susceptible and 242 PVY-resistant F1 individuals).
RenSeq combined with SMRT sequencing and bulked segregant analysis leads to successful cloning of Rysto
It was previously shown that Rysto maps to the distal end of long arm of potato chromosome XII (Flis et al 2005, Song et al 2005, van Eck et al 2017), which corresponds to the region downstream of 58 Mb in the reference potato genome clone DM (PGSC, 2011). Jupe et al. (2013) showed that this region contains 18 complete and partial NLR immune receptors from both TIR-NLR and CC-NLR classes, organized in four clusters (Figure 1a). We therefore hypothesized that Rysto likely encodes an NLR, and applied RenSeq to predict candidate genes. To assemble the NLR-encoding genes of the resistant parent with high confidence, SMRT-RenSeq was performed. Assembly of PacBio Reads of Interest (ROI) resulted in 1,555 contigs, out of which 1254 were annotated as NLRs using NLR-parser software (Steuernagel et al. 2016). Mapping of short read RenSeq data from resistant (R), susceptible (S) and bulked susceptible (BS) plants, and subsequent polymorphism calling resulted in 10 linked contigs showing presence/absence polymorphism. Expression of candidate genes was confirmed using cDNA-RenSeq data obtained from the resistant parent. These analyses resulted in 11 candidate NLRs. Four candidate genes belonged to CC-NLR and seven to the TIR-NLR class. A phylogenetic tree constructed with full-length amino acid sequences showed that candidate genes belong to three different clades and show only distant relation with cloned functional Solanaceae genes. All TIR-NLRs belonged to the same clade and show amino acid identity of between 75% and 90%. The CC-NLRs genes split into two distinct clades, consistent with their physical separation in the potato genome (Fig. 1, Supplementary Fig. 1).

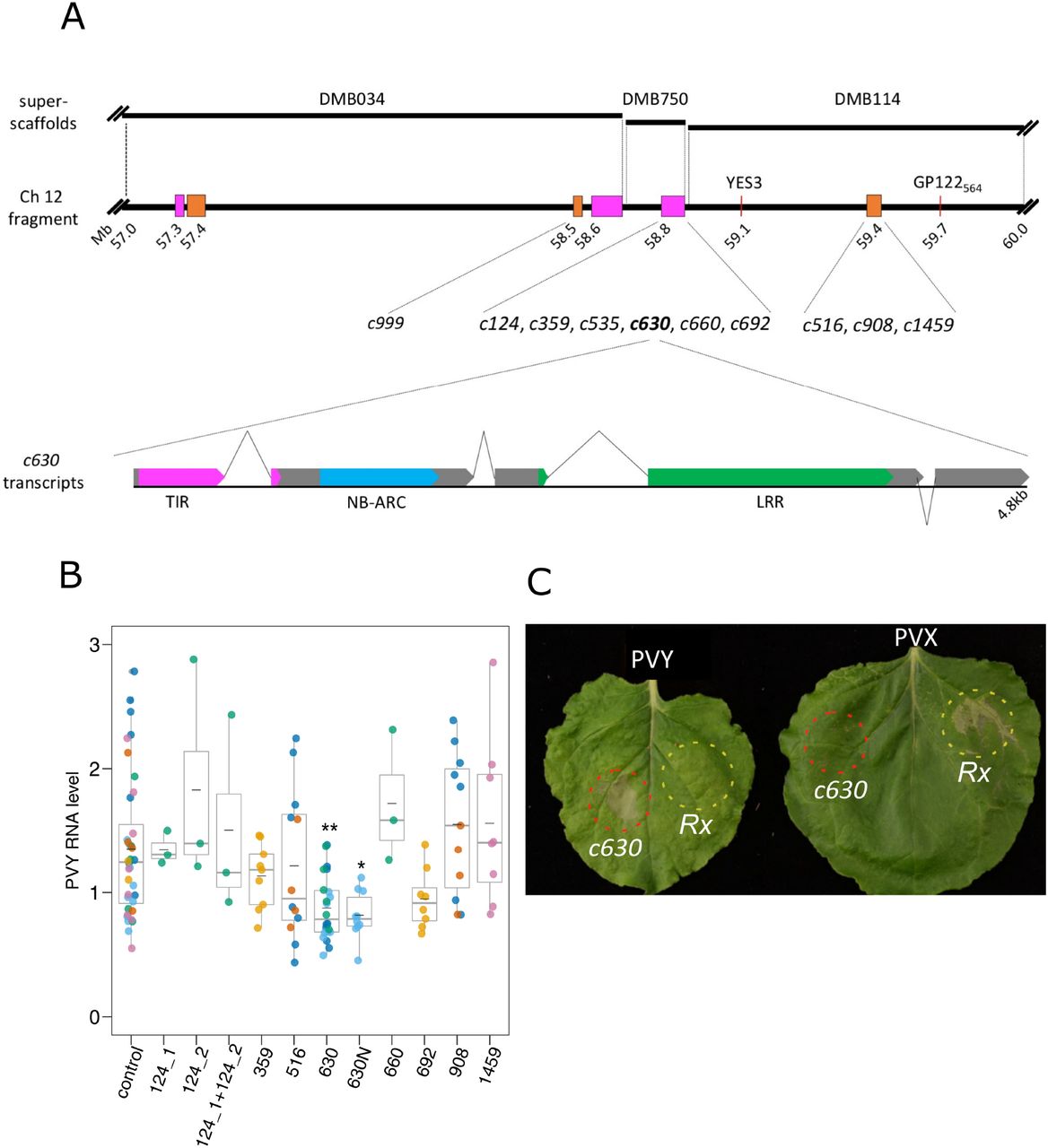
(a) Schematic representation of candidate genes and a fragment of chromosome 12 linked to the Rysto gene.
Potato superscaffolds (DMB034, DMB750 and DMB114; top panel) were aligned to the distal end of longer arm of potato chromosome 12 (57 to 60Mb fragment shown; middle panel), which is linked to the Rysto gene. Annotated NLRs/NLR clusters are depicted in orange (CNL) and magenta (TNL). Red lines indicate known markers linked to Rysto. Distances are given in megabases (Mb) according to the DM potato reference genome (PGSC 2011), numbers indicate proximal position of marked sequences. Candidate contigs for Rysto derived from SMRT RenSeq were aligned to the DM potato genome and best hit position is shown on the map. Map drawn to the scale.
Lower panel presents schematic structure of the c630 transcripts. Most abundant version (supported by ~80% of cDNA-RenSeq reads) consists of 4 exons. Additional intron (depicted with inverted lines) positioned 24 nucleotides upstream of initial stop codon results in fifth exon at 3’ end. Exons drawn in grey, colors indicate canonical NLR domains; TIR (magenta), NB-ARC (blue) and LRR (green). Scheme drawn to the scale.
(b) Relative level of PVY RNA after infection of N. benthamiana leaves transiently expressing candidate contigs.
Three leaves of N. benthamiana plants were infiltrated with the vector pICSLUS0003::35S overexpressing c124_1, c124_2, c359, c516, c630, c660, c692, c908, and c1459, pICSLUS0001 vector overexpressing c630 under control of native regulatory elements or an empty vector48 hours later leaves were inoculated with PVYNTN strain. Seven days after PVY inoculation mRNA was isolated from upper, non-inoculated leaves and PVY RNA levels were quantified with qPCR. As reference genes EF1 and L23 were used. Error bars represent the standard deviation of the means (-). Additionally median is also presented. Statistical analysis was perform using one-way ANOVA with Dunnett’s test.
Experiments were performed on 3-10 plants for each construct and repeated three times.
(c) HR response of N. benthamiana plants expressing c630 or Rx genes.
Two fully developed leaves of N. benthamiana plants 14 days following PVYNTN (left) or PVX infection (right), were infiltrated with a suspension of A. tumefaciens carrying pICSLUS0003c630 (area marked with red) or pGBT-Rx-GFP (area marked with yellow) Seventy-two hours after infiltration expression of c630 in PVY infected plants resulted in HR similarly to when Rx gene was delivered into plants infected with PVX virus. Experiment was repeated three times.
Homology searches of candidate genes to the reference genome positioned them in three clusters located in the distal end of the long arm of chromosome 12 (58-59Mb of DM reference genome, Fig. 1a), where markers linked to Rysto are also located (GP122564, Witek et al. 2006 and YES3, Nie et al. 2016). This part of chromosome 12 consists of three neighboring superscaffolds DMB034, DMB750 and DMB114 and one of them (DMB114) was suggested to be a likely location of Rysto (van Eck et al. 2017). Thus, all selected candidates NLR genes from SMRT RenSeq data localize to the distal end of chromosome 12 where Rysto was previously positioned.
Functional analysis of candidate contigs in transient expression assay in N. Benthamiana plants
To establish whether the candidate genes are functional in planta, the open reading frames (ORFs) of 11 co-segregating NLRs were PCR-amplified and placed under control of the 35S promoter in a binary vector pICSLUS0003 as described (Witek et al., 2016). Two functional tests were then performed. In the first test, the constructs were transiently expressed in N. benthamiana, followed by PVYNTN inoculation. At 7 dpi, leaf samples were collected and levels of viral RNA were measured using qPCR. One candidate gene (c630) reduced virus multiplication as compared to control N. benthamiana plants (Fig. 1b). The c630 ORF with its native 5’ and 3’ regulatory sequences (2,263 nt upstream from 5’ and 1,013 nt downstream from 3’ end, respectively) was cloned into the pICSLUS0001 binary vector as described (Witek et al., 2016). Transient assays in N. benthamiana also showed a statistically significant (P = 0.011475) inhibition of virus multiplication (Fig. 1b). Candidate gene c999 showed autoactivity, even when infiltrated with low OD (0.1) and another candidate gene, c535 could not be cloned.
In the next step, N. benthamiana plants were systemically infected with PVYNTN or PVX, followed by c630 transient expression by Agro-infiltration. The expression of c630 in plants carrying PVYNTN resulted in a hypersensitive response (HR) (Fig. 1c), similar to when the Rx gene was delivered into plants infected with PVX (Bendahmane et al., 1999)
Expression of c630 triggers HR to different viruses
To test whether c630 elicited an immune response to different of strains of PVY N. benthamiana plants were infected with: 0, N, N-Wilga, NTN isolates of PVY, or cross related Potato Virus A (PVA). At 7 dpi, leaves showing symptoms of viral infection were infiltrated with Agrobacterium strains carrying c630 or an empty vector. Transient expression of c630 resulted in a strong HR in plants carrying all PVY isolates and PVA, but not PVX (Potato virus X) or TMV (Tobacco Mosaic Virus) (Supplementary Tab. 1.). No HR was observed when c630 was not expressed in control plants. These experiments further verified c630 as a candidate for the functional Rysto gene.
Rysto expression restricts systemic spread of PVY and PVA
To further investigate Rysto function, stable transgenic Solanaceae plants (potato and tobacco) carrying Rysto or two other non-functional homologues under the control of 35S promoter or native regulatory elements were created using Agrobacterium-mediated transformation.
Two potato cultivars S. tuberosum cv. Maris Piper (MP) and cv. Russet Burbank (RB), susceptible to PVY, were then chosen to test Rysto functionality. Transgenic plants from both cultivars carrying Rysto under control of 35S promoter or native promoter (for MP only) were inoculated with PVYNTN or mock treated with water. Systemic virus spread was monitored in upper, non-inoculated leaves using enzyme-linked immunosorbent assay (ELISA) at 3-weeks after inoculation. In the MP background expressing Rysto under control of 35S, in 10 out of 12 tested transgenic lines PVY was not detected in the upper part of the plant (Supplementary Tab. 2). To confirm these ELISA results, three randomly chosen transgenic lines (two resistant and one susceptible) were tested using qPCR (Fig. 2b). The inhibition of virus multiplication and spread in two resistant transgenic plants correlated with elevated levels of Rysto gene expression. In the susceptible transgenic line, the expression of Rysto transgene was not detected. In cv. Russet Burbank in all 4 tested independently isolated transgenic lines, PVY was not detected using ELISA or using qPCR (Fig. 2c, Supplementary Tab. 3).

(a) Illustration of ER type response of PVY-inoculated leaves of S. tuberosum cv Russet Burbank plants expressing Rysto.
Four-week-old transgenic potato plants cv. Russet Burbank carrying construct 35S::Rysto-630, and non-transformed plants were inoculated with PVYNTN. Two weeks after infection chlorosis were observed on inoculated leaves of non-transformed plants (right) in contrast Rysto transgenic plants (left) remained symptomless.
(b) Effect of Rysto expression completely blocks PVY spreading in transgenic potato plants. Four-week-old transgenic potato plants cv. Russet Burbank (panel B) or Maris Piper (panel C) carrying construct 35S:: Rysto, Maris Piper plants carrying Rysto construct under control of native 5’- and 3’-regulatory elements (panel D) and non-transformed plants were inoculated with PVYNTN. Three weeks after viral inoculation, mRNA was isolated from upper, non-inoculated leaves. The PVY RNA levels and the expression of Rysto were quantified using qPCR, relative to the EF1 and Sec3 reference genes and expressed as means ± SD calculated from three biological replicates per plant line.
Additionally, when Rysto gene was expressed under control of its native promoter all tested lines (5 lines) were PVY free (Fig. 2d). In all resistant lines non-macroscopic symptoms of HR were observed, regardless of the genotype of potato used (Fig. 2a).
N. tabacum 35S::Rysto transgenic and non-transformed control plants were inoculated with PVYNTN as previously. At 5 dpi, some necrotic lesions were visible only on inoculated leaves of Rysto plants, while no macroscopic symptoms of HR were observed on the non-transformed control plants (Supplementary Fig. 2). In addition, control susceptible plants developed typical systemic symptoms of PVY infection. qPCR on upper, non-inoculated leaves showed that systemic spread of the virus was fully restricted in Rysto expressing lines, while PVY RNA levels in non-transformed plants were high. These results were confirmed in T0 (Supplementary Fig. 3A) and T1 generation plants (Supplementary Fig. 3B). In transgenic tobacco lines expressing Rysto under control of its natural 5’ and 3’ regulatory elements, a local HR similar to that seen in 35S::Rysto plants was also observed at 5 dpi. The absence of viral RNA was confirmed with qPCR (Supplementary Fig. 4) at 7 and 14 dpi and was fully correlated with Rysto transcript expression.
Introduction of Ry genes from Solanum stoloniferum to S. tuberosum may condition resistance not only to PVY but also to close related PVA (Cockerham, 1970). To test whether expression of Rysto determines resistance to PVA, tobacco plants with and without functional Rysto transgene were inoculated with this virus. Western blotting analysis with anti PVA antibodies revealed inhibition of PVA systemic spread in Rysto transgenic ones (Supplementary Fig. 5).
The PVY and PVA coat proteins (CP) are elicitors of Rysto triggered immunity
It was previously reported that there is a single ORF encoding a polyprotein in the PVY genome, which is cleaved by three viral proteases to form functional proteins (Jakab et al., 1997). Thus, to identify the elicitor of the Rysto-triggered immunity the open reading frame from the most promising putative viral proteins (HcPro, NIa, NIb, coat protein [CP]) were cloned and transiently expressed with in transgenic Rysto and non-transformed tobacco plants. Among all tested proteins only the coat protein induced strong HR at 2-3 days post treatment in Rysto plants, whereas no HR was observed in WT control (Fig. 3a). A similar type of analysis was performed for PVA virus. Only PVA coat protein was able to elicit HR when is expressed in transgenic Rysto plants (Fig. 3b).

(a) Rysto recognizes (upper panel) PVY and (lower panel) PVA coat proteins as avirulence factors in transient expression assays.
To identify the elicitor of the Rysto mediated resistance against PVY or PVA viruses the open reading frame of putative viral proteins (HC-Pro, Nia, Nib and CP) was cloned to pBIN53 vector and transiently expressed in N. tabacum. Rysto transgenic and non-transformed plants. As controls (C1, C2) A. tumefaciens or infiltration buffer were used. Three days after treatment HR was observed only for PVY or PVA coat protein (CP).
(b) Comparison of the selected PVY and PVA coat protein amino acid sequences. The comparison is made against PVYNTN CP. Identical residues are shaded in black. The PVA CP shares above 59% identity with the PVY sequence. The alignment was generated using MAFFT-L-INS-I (Katoh and Toh, 2008) and visualized in Jalview 2,10,4b1 (Waterhouse et al. 2009).
Extreme resistance confirmed by Rysto is epistatic to the Ny-1 mediated HR
The functional relationship between two types of defence response to PVY infection, ER and HR, was investigated. Potato cultivar (Rywal) carrying the Ny-1 gene, responsible for HR to PVY infection (Szajko et al., 2008 and Baebler et al., 2014) was transformed with Rysto. Plants of both genotypes (Ny-1 or Rysto /Ny-1) were inoculated with PVYNTN as described previously. Five days after infection HR developed only in parental genotype (Ny-1), while no sign of HR was observed in the Rysto /Ny-1 genotype (Fig. 4b). This indicates that ER conferred by Rysto is epistatic to Ny-1 dependent HR.

(a) Determination of epistatic effect of ER to HR.
Potato Rysto/Ny-1 and Ny-1 plants were inoculated with PVYNTN. Five days after infection symptoms of local cell death occurred only in Ny-1 plants. Experiment was repeated three times.
(b) Rysto dependent PVY multiplication is inhibited in elevated temperature.
Potato Rysto /Ny-1 and Ny-1 plants were inoculated with PVYNTN and divided to two groups; the first group was kept at 20°C, the second at 28°C. Water-treated plants were used as negative controls. Seven days following inoculation, samples of inoculated (lower graphs) and upper non-inoculated leaves (upper graphs) were collected and PVY RNA levels were measured qPCR, relative to the EF1 and Sec3 reference genes and expressed as means ± SD calculated from three biological replicates per plant line. A and C describes names of transgenic Rysto lines used.
(c) Stable transgenic N. tabacum plants carrying Rysto under control of a 35S promoter are resistant to PVY infection in elevated (32°C) temperature.
Seven-week-old N. tabacum 35S::Rysto transgenic and non-transformed plants were inoculated with PVYN205-GFP clone. Typical symptoms of PVY infection were observed after 7 dpi in non-transformed plants in contrast Rysto lines remained symptomless. Picture was taken at 14 dpi.
Extreme resistance mediated by is temperature-independent
To investigate the role of temperature in Rysto-mediated ER, the response to PVY infection was studied in a Ny-1 or Rysto /Ny-1 genotype. PVY mRNA levels were measured 14 dpi in upper non-inoculated leaves following mock and virus treatments in two types of temperature conditions (22°C and 28°C). In both genotypes, no PVY multiplication was observed when plants were kept at 22°C. In contrast, when kept in elevated temperature in Ny-1 genotype HR was inhibited and PVY RNA was detected systemically, whereas in transgenic Rysto /Ny-1 lines PVY spreading was still fully inhibited (Fig. 4a,b). To test whether Rysto hinders systemic infection above 28°C, transgenic and non-transformed tobacco plants were infected with GFP-tagged PVY (PVYN605-GFP) at 32°C. Interestingly, expression of Rysto at higher temperature still prevented systemic spread of the virus (Fig. 4c).
These observations suggest that unlike the tobacco N gene, which also encodes a TIR-NLR, high (>28°C) temperature does not compromise Rysto immunity to PVY infection (Samuel 1931).
NRG1 and EDS1 but not SA are crucial for Rysto dependent immunity
For N-dependent resistance of tobacco to TMV, additional genes EDS1 and NRG1 are required (Collier et al.2011, Liu et al., 2002, Peart et al., 2005). Like N, Rysto carries a TIR domain, so we examined whether these two regulator proteins mediate a Rysto dependent response to PVY infection. To test this hypothesis, EDS1 or NRG1 N. benthamiana mutant plants were infected with PVYNTN followed by transient expression of Rysto or GFP, as a control. 72 hpi in WT plants, HR was observed. In contrast, both knockout plants remained symptomless. This suggests that both NRG1 and EDS1 proteins are crucial for the Rysto-mediated response. A similar analysis was performed with N. benthamiana plants expressing bacterial NahG that encodes salicylate hydroxylase (Friedrich et al., 1995). In WT and in NahG plants, symptoms of HR were observed at 72 hpi (Fig. 5). This suggests that elevated levels of SA are not essential for establishing a Rysto dependent resistance.
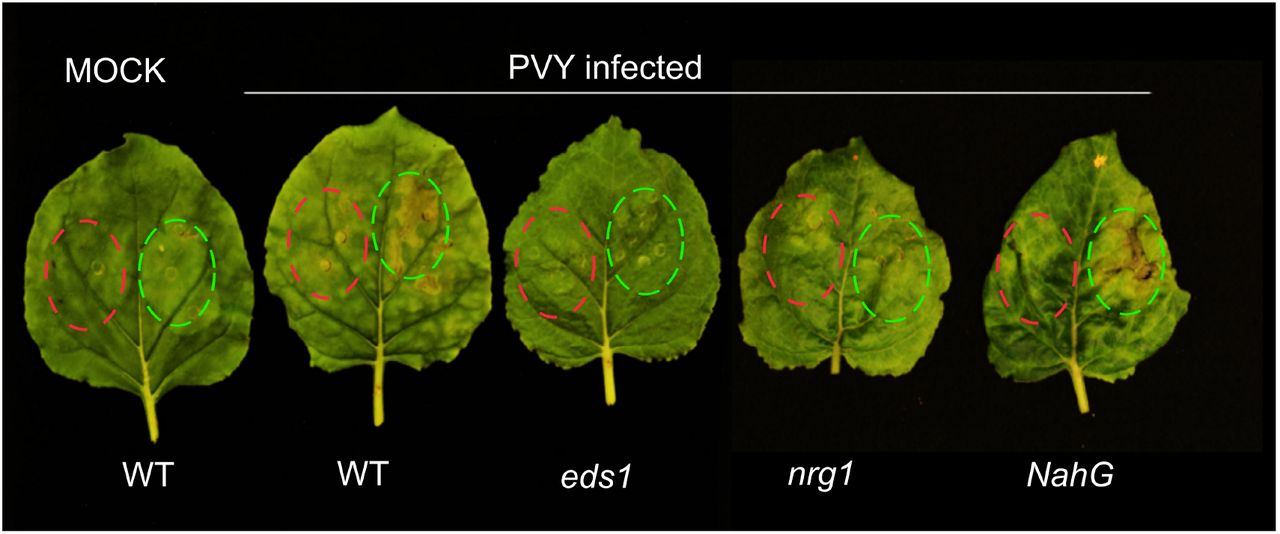
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Fully developed leaves of N. benthamiana EDS1, NRG1 knockout plants or NahG and non-transformed ones were infected with PVYNTN or Mock-treated. Two weeks later leaves with symptoms of PYV infection were infiltrated with suspension of A. tumefaciens carrying pICSLUS0001:c630 (highlighted in green) or pGBT-GFP (highlighted in red) as a control. Three days after infiltration in non-transformed plants and NahG plants (SA free) symptoms of cell death were observed. The experiment was repeated three times with similar results.
DISCUSSION
Genes conferring ER against PVY infection had been introduced into potato cultivars from wild or domesticated species from Solanaceae family, e.g. S. chacoense, S. stoloniferum or S. tuberosum ssp. andigena. Two alleles of Rysto gene derived from S. stoloniferum: Ry-fsto and Rysto, have most often been used (Flis et al. 2005, and Song et al. 2005). In this study, we report the isolation and characterization of a novel, broad-spectrum resistance gene Rysto using RenSeq (Jupe et al. 2013, Witek et al. 2016).
We hypothesized that the underlying PVY resistance gene encodes an NLR protein. Data obtained from SMRT and Illumina sequencing together with SNP calling and presence/absence polymorphism detection revealed 11 transcriptionally active NLR genes, as candidates for the Rysto gene. We mapped these candidates to the DM reference genome (PGSC 2011) at the distal end of chromosome 12, a region shown to carry Rysto-linked markers. Additionally, using graphical genotyping on panel of tetraploid potatoes, van Eck et al. (2017) suggested that Rysto locates on super-scaffold DMB114. This is consistent with our data, as three of 11 candidates were located on this super-scaffold, while remaining on neighboring super-scaffolds, all within 1 Mb distance. Interestingly, all linked NLRs showed presence/absence polymorphism with the same allele ratio, indicating that the whole interval introgressed from S. stoloniferum rarely recombines in cultivated potato.
Functional assays in N. benthamiana and cultivated potato plants confirmed that only one candidate, Rysto, stopped PVY multiplication and spread. The protein encoded by Rysto has motifs and domains typical of TIR-NLR-type resistance proteins and shares less than 34% identity at amino acid level with previously described TNL-type resistance proteins from Solanaceae., including: N, Y-1, Bs4 and Pvr4.
As most potato cultivars lack broad-spectrum resistance to circulating strains of PVY (Valkonen 2015) we showed that Rysto confers resistance against multiple PVY strains (Suppl. Table 1). In contrast, for Rx, Rsv3 or Sw-5 genes mediating ER to other viruses, isolates breaking immunity have appeared (Moreira et al. 1980, Querci et al. 1995, Brommonschenkel et al. 2000, Zhang et al. 2012). Recently two novel genes conferring immunity to some PVY strains were described. Kim et al (2017) isolated, from Capsicum annuum, a dominant gene Pvr4 conferring a broad range of resistance to potyviruses, including some PVY isolates. However, it was tested only in heterologous system (N. benthamiana) and not in any Solanaceae crops. A single recessive gene, called NtTPN1, isolated from tobacco causes veinal HR-like necrosis (Michel et al., 2018). Moreover, Pvr4 was never tested in potato and there is no evidence that it would be functional in that background. Our results suggest that the Rysto-mediated immunity is effective in several potato cultivars and tobacco not only against different strains of PVY but also to the related PVA (Fig. 2a, 4c, Supplementary Fig. 4).
We tested multiple PVY reading frames and found that viral coat protein (CP) is the recognized factor in Rysto-mediated immunity against PVY and PVA (Fig. 3a). PVY and PVA CPs share ~59% identity (Fig. 3b) and have nucleocytoplasmic localization in infected cells (Supplementary Fig. 6). Interaction of Rysto with CP, directly or indirectly, initiates a response leading to ER by preventing virus accumulation at an early stage of infection. Resistance may be due to inhibition of viral cell-to-cell movement via callose deposition as was reported in soybean-SMV pathosystem (Seo et al., 2014) or by translational arrest of viral transcripts as was described for N gene-mediated resistance to tobacco mosaic virus (TMV) in a process that involves Argonaute 4 (Bhattacharjee et al., 2009). The CP might be recognized Rysto either by recognition of specific amino acid sequence or of a conserved element of CP structure as was reported for Rx protein (Baurès et al., 2000, Candresse et al., 2010).
Rx can trigger HR when co-expressed with PVX coat protein (Bendahmane et al. 1999). Using N. benthamiana we found that Rysto behaves similarly. When plants were infected with PVY followed by Rysto transient expression a strong HR was observed (Fig. 1c). We speculated that the nature of the Rysto response, as either ER or HR, might be determined by the elicitor expression levels or genetic background used. When CP was expressed under control of constitutive promoter (35S) in transgenic Rysto tobacco plants it elicited an HR, whereas CP produced during PVY infection led to ER. Moreover, when Rysto was introduced into S. tuberosum plants no HR symptoms were observed but in N. tabacum, necrotic lesions on inoculated leaves were noticed upon PVY infection (Supplementary Fig. 2). This was also observed for Rx gene where the specificity of recognition was modulated by host factors differing between Rx-expressing potato and tobacco plants (Baurès et al., 2008).
For different resistance genes, ER or HR formation and control of viral spread might be determined by the dose of the R gene. High level HRT gene expression in susceptible Arabidopsis thaliana leads to restriction of virus replication or movement without any visible symptoms of HR, while lower levels of HRT transcript in resistant Arabidopsis lines leads to micro-HR or HR with systemic virus movement, respectively (Cooley et al., 2000). In contrast, our results suggest that ER is not affected by Rysto expression levels. Relative expression levels of Rysto differ between lines depending on the promoter used. Transgenic potato lines under control of native regulatory elements exhibited much lower transcript expression in comparison to those under control of 35S. Nevertheless, these differences did not influence the ER phenotype which was observed in all experiments (Fig. 2b, 2d).
Not only artificial manipulation of R gene expression and dose but also zygosity can affect the mode of immunity. HR induction was also observed for the Tm-2 gene of tomato and its allelic gene, Tm-22, conferring ER to tomato mosaic virus (ToMV). The Tm-2 and Tm-22 confer extreme resistance with little or no induction of HR to ToMV in a homozygous state but induce moderate resistance with accompanying HR-like cell death in a heterozygous state or even in a homozygous state at high temperatures (Hall 1980) suggesting that Tm-2 and Tm-22 are involved in cell death induction.
We also showed that Rysto’s mode of action, unusually for a TIR-NLR, is temperature independent (Fig. 4b, c). Many R genes are non-functional at 28°C or above. This was described for the tobacco N gene, the Mi-1 gene in tomato conferring resistance to root-knot nematodes (Jablonska et al., 2007), the Ny-1 gene conferring resistance to PVY (Szajko et al., 2014), the NLR gene SNC1 in Arabidopsis (Yang and Hua 2004) and many others. All these genes induce defense in permissive temperature but when shifted to elevated temperature the HR is arrested. In contrast, we have shown that Rysto- mediated immunity is still effective in different Solanaceae species at or above 28°C (Fig. 4b, 4c). The exact mechanism of temperature sensitivity remains elusive. It might be dependent on the transcription factor PIF4 which acts as a negative regulator of plant immunity and modulation of its function, which alters the balance between growth and defense (Gangappa et al., 2016). It may also relay on Enhanced Disease Susceptibility1 (EDS1) and Phytoalexin Deficient 4 (PAD4)-transcript accumulation which together promote basal resistance and plant growth inhibition in immune-activated backgrounds (Rietz et al., 2011).
A detailed understanding of the functional relationship between HR and ER has not been yet proposed. In principle, the genes for ER act earlier and more efficiently than the genes for HR, which was shown in comparative studies on the potato Rx and N genes (Bendahmane et al. 1999). Rysto-mediated ER is epistatic to Ny-1 mediated HR against PVY in potato (Fig. 4). We also provide evidence that Rysto/Ny-1 plants are PVY free at elevated temperature (28°C) in contrast to Ny-1 plants (Baebler et al., 2014). Collectively, Rysto-mediated ER is epistatic to HR. Nevertheless, there is a possibility that ER and HR pathways might be interdependent. For instance, Rysto might have been competitive with Ny-1 in recruitment of some signaling components. As Rysto may have higher affinity to selected components than Ny-1, Rysto- mediated immunity is stronger and goes quicker. These components might act as helper NLRs especially since R proteins also engage in complex genetic and biochemical interactions (Kamoun et al., 2018).
To date 314 R genes have been characterized (Kourelis et al., 2018). Only 128 have a proposed mode of action, based on an even lower number of detailed studies of molecular mechanisms. The underlying mechanisms of the remaining 186 R genes are yet unknown (Kourelis et al., 2018). In the Solanaceae CNLs outnumber TNLs by 4.7: 1 (Witek et al., 2016). Tobacoo N mediates recognition of the helicase domain p50 via interaction with NRIP1 protein and activates a resistance response requiring several known general cofactors of disease resistance: EDS1, SGT1, RAR1, HSP90, members of the COP9 signalosome (Liu et al. 2002, 2003, 2004) and NRG1, the CNL helper protein (Peart et al., 2005). Since Rysto contains a TIR domain we decided to search for a mode of action similar to N.
In EDS1 and NRG1 mutant N. benthamiana plants, the HR after PVY infection and Ry transient expression is lost compared to HR observed in non-transformed control plants. This clearly shows that Rysto’s mode of action is EDS1- and NRG1- dependent (Fig. 5), while Pvr-4 mediated ER to PVY infection in N. benthamiana is EDS1-independent (Kim et al. 2018).
In our investigation, HR occurred after PVY infection and Rysto transient expression in SA-depleted NahG plants (Fig. 5). This indicates that increase of SA is not crucial for Rysto- mediated immunity (Fig 5). Interestingly EDS1 is considered to be a positive SA regulator (Gangappa et al. 2016). Moreover, the EDS1/PAD4-controlled regulatory feedback loop involving SA contributes to the suppression of plant immune response at elevated temperature (Wang et al., 2009). This is in line with a reported lowering of EDS1 protein levels under high temperature conditions (Stuttman et al., 2009). As resistance after PVY infection in Rysto plants was still observed both in 28°C and 32°C we concluded that neither EDS1 nor NRG1 are involved in temperature sensitivity. Moreover, EDS1 may also be involved in a signaling pathway unrelated to SA.
Not only NRG1 but also ADR1 activity may contribute to TNL-mediated immunity (Dong et al. 2016), so additional studies on ADR1 involvement in Rysto’s mode of action are required.
We propose Rysto-mediated immunity to ER against PVY results from Rysto recognition of PVY coat protein and recruitment of such downstream signaling components as NRG1 and EDS1. Conceivably, components of the co-chaperone complex: SGT1, RAR1, HSP90 are also involved in Rysto mediated immunity as factors supporting correct folding of resistance proteins (Peart et al. 2005). Rysto might also be negatively regulated through miRNA activity either by promoting mRNA degradation, inhibiting translation, or suppressing transcription by epigenetic modification.
In summary, our results demonstrate that Rysto plays an important role in defense, and may prove valuable for breeding PVY-resistant cultivars of potato and other Solanaceae crops. Why some NLRs trigger ER and some HR remains obscure, but nevertheless, this type of recognition leads to durable and efficient resistance.
MATERIAL AND METHODS
Plant material
All potato cultivars (Solanum tuberosum ssp. tuberosum) and viruses: PVY0, PVYN, PVYN-Wi and PVX, PVA were obtained from Laboratory for Potato Gene Resources and In Vitro Cultures at the Institute of Plant Breeding and Acclimatisation - National Research Institute, Bonin (Poland). Plants were grown for four weeks in soil under controlled environmental conditions (22°C; 16 h light, and 18°C 8 h dark) as described previously (Szajko et al. 2008). Tobacco plants Nicotiana tabacum cv. Xanthi-nc and Nicotiana benthamiana were grown for six weeks in soil under controlled environmental (22°C, 16h light, 8h dark) as described previously (Hoser et al. 2013). Transgenic potato and tobacco plants were regenerated followed Agrobacterium leaf disc transformation as described by Mac et al. (2004).
Mapping population
Diploid potato (2n = 2x = 24) mapping population was developed by a cross between PVY-resistant heterozygous dihaploid clone dH Alicja, and susceptible clone 83 −3121. dH Alicja was obtained from potato cultivar Alicja via parthenogenesis. The ER to PVY in dH Alicja was conferred by the gene Rysto which was derived from clone MPI 55.957/54. MPI 55.957/54 had in its pedigree Solanum stoloniferum from wild Solanum spp. collection of Max Planck Institute (Ross H. (1958) Handbuch der Pflanzenzüchtung, 2nd, Vol. III:106- 125).
Short- and long-read RenSeq and bioinformatic analysis of the sequencing data
Short-read RenSeq libraries and enrichment was carried out on susceptible (S, 83-3121 clone), resistant (R, dhAlicja) and 149 bulked susceptible plants (BS) as described (Jupe,. 2013 and Witek et al., 2016). Enriched libraries were sequenced with Illumina MiSeq platform using 500 cycles. Additionally, RenSeq was performed on cDNA from resistant parent as described previously (Witek et al., 2016) and sequenced on Illumina HiSeq platform with 300 cycles. Long-read PacBio RenSeq was performed as described in Witek et al. (2016) and PacBio was sequenced on a single RSII SMRT cells what resulted in 69,737 Reads of Interest (ROI) with minimum three passes and accuracy above 90%. and are available under accession number PRJEB28862. Reads obtained from SMRT RenSeq were assembled using Geneious 8.1.2 software as described before (Witek et al.,2016) resulting in 1,555 contigs. R, S and BS reads from Illumina 250PE were mapped to contigs derived from SMRT RenSeq assembly using BWA (Li and Durbin, 2009), with default settings. SNP calling and candidate prediction was performed as described previously (Jupe et al.,2013, Witek et al.,2016). Contigs showing >95% of susceptible allele in mapped BS data were considered as linked. Additionally, candidate NLRs showing presence/absence polymorphism between R and S parent, and linkage to resistance based on BS samples were called. Briefly, numbers of pair-end mapped reads to contigs derived from SMRT RenSeq assembly were calculated using TSL Galaxy built-in scripts (MacLean and Kamoun, 2012) for each R, S and BS samples. Resulting data were sorted and visualized in Microsoft Excel software. As a presence/absence polymorphism, contigs with at least 250 mapped reads from R parent, and less than 10% and 18% of that number for S and BS samples, respectively, were considered. Expression of the candidate genes was determined using cDNA-RenSeq data from R parent as described previously (Andolfo et al., 2014, Witek et al., 2016).
DNA extraction
RenSeq experiments and PCR reactions were conducted on gDNA freshly extracted from young leaves (both MiSeq and PacBio protocols) using the DNeasy Plant Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol.
RNA extraction for RenSeq experiments
For the cDNA RenSeq experiment, RNA was extracted using TRI-Reagent (Sigma-Aldrich, MO, USA) and Direct-zol RNA MiniPrep Kit (Zymo Research, CA, USA), following manufacturer’s recommendations. First-strand cDNA was made using a mix of oligo-dT and random hexamer primers and SuperScript II First-Strand Synthesis System (Sigma-Aldrich, MO, USA). The second strand was made as described (Rallapalli et al., 2014).
Cloning of the candidate ORFs
For each of 12 selected PacBio contigs PCR primers flanking predicted ORFs were designed. All primers were supplemented with specific 5′ and 3′ extensions to make them compatible with custom USER expression vectors used in this study as described previously (Witek et al., 2016) (see Supplementary Primers Table2). Candidate genes were PCR-amplified from R parent gDNA in 25-μl PCR reactions (35 cycles with annealing at 62 °C and 7 min extension at 72 °C) using Kappa HiFI HotStart Uracil+ Fidelity Polymerase (Manufacturing, R&D Cape Town, South Africa). 30 ng of purified PCR product was hybridized with 30 ng pICSLUS0003 vector in the presence of 1 μl of USER enzyme mix (New England Biolabs, Inc., MA, USA) as described (Witek et al., 2016). All constructs were verified by DNA sequencing. Plasmids containing the candidate genes were transformed into Agrobacterium strain GV3101 for transient complementation assays or LBA404 for a stable potato using the method described (Mac et al.,2004). To create a genomic construct of Rysto (c630) under its native regulatory elements, we PCR-amplified the whole contig (7.5 kB) that was assembled from PacBio reads. PCR amplicons were hybridized into USER - vector pICSLUS0001 lacking 35S promoter and OCS terminator.
Transient expression assay
Infiltrations of Agrobacterium GV3101 carrying the plasmids containing candidate genes were performed in N. benthamiana. Bacteria were grown in YEB medium (5 g of beef extract, 5 g of bacteriological peptone, 5 g of sucrose, 1 g of yeast extract, and 2 ml of 1 M MgSO4 in 1 litre of milli-Q water) supplemented with gentamicin, rifampicin and kanamycin for all constructs. After 1 day, overnight cells were centrifuged at 3,500 rpm and resuspended in infiltration buffer containing 10 mM MgCl2, 10 mM MES (pH 6.5), 100 μM acetosyringone to a final OD600 of 1 and infiltrated into leaves of 4-week-old plants with a 3-ml syringe. HR reaction was observed between 2 and 3 day after infiltration. The same method was used for bacteria carrying a binary plasmid pBIN53 with PVY sequences. Each experiment was repeated and included at least three independent biological replicates.
Resistance assay
In all experiments related to monitoring of PVY infection the isolate NIB-NTN (GenBank: AJ585342.1) was used. The infection was performed in each time on a seven-week-old N. tabacum plants or 4-week-old potato plants under conditions as described (Baebler et al. 2014). In experiments related to different PVY isolates plants were infected under the same experimental conditions with following PVY isolates: 0 (AJ890349), N (FJ666337), N-Wilga (EF558545), or unrelated viruses PVA, PVX and TMV(U1).
RNA extraction and gene expression analysis (qPCR)
Total RNA was extracted using a modified guanidinium-phenol-chloroform protocol (Chomczynski and Sacchi, 1987), treated with DNase1 (ThermoFisher Scientific), and subjected to reverse transcription using mix of random hexamer and oligo primer and RevertAid First-Strand cDNA Synthesis Kit (ThermoFisher Scientific). Gene expression analysis via RT-qPCR was performed using the LightCycler®480 instrument and LightCycler®480 SYBR Green I Master Kit (Roche). Relative gene expression levels were determined using a standard curve method and the value for each target gene was then normalized against the mean of expression values of two reference genes: EF1 and L23 or EF1 and Sec3 for N tabacum and N. benthamiana or potato cultivars respectively, as described (Liu et al., 2012, Tang et al. 2017). Measurements for each sample was analysed in four technical replicates and two dilutions. All primers used for qPCR are described in Supplementary Table 4.
PVY proteins cloning
A full-length infectious clone PVY-N605 (Jakub et al. 1997) was used to create all PVY constructs. Each putative PVY protein was PCR amplified and cloned to pBIN53 vector which carries the CaMV 35S promoter, the PVY 5′ and 3′ UTRs separated by a polylinker, and a poly(A) tail vector as described previously (Mestre et al., 2000).
Western blotting
Agro-infiltrated leaves were collected and ground in liquid nitrogen. Total protein was extracted by incubating the ground leaf samples in extraction buffer containing 100 mM Tris HCl pH 8.0, 1 mM EDTA, 150 mM NaCl, 7.2 mM β-mercaptoethanol, 0.5 mM 4-(2- aminoethyl) benzenesulfonyl fluoride hydrochloride (AEBSF) and 0.03 l M PMSF (phenylmethylsulfonyl fluoride) for 15 min. The homogenate was centrifuged at 21,000×g for 10 min and the supernatant was collected. Samples were fractionated by 12.5% SDS–PAGE and subjected to immunoblot analysis using appropriate alkaline phosphatase-conjugated anti PVA antibodies (Bioreba, Switzerland). Immunoblots were developed using the NBT/BCIP colorimetric detection kit from BioShop Canada Inc.
Statistical analysis
Statistical analyses were conducted using R 3.2.2 within R Studio 0.99.483 Technical replicates consist of readings from the same plant in the same experiment, whereas biological replicates consist of independent plants. Data were analysed using the following pipeline: data were assessed for their suitability to be analysed using parametric tests by testing for the normal distribution of the residuals and Shapiro–Wilk test. If the data were suitable for conducting parametric tests, then analysis of variance (ANOVA) were used. As post-hoc the Dunnett’s or Tuckey’s HSD test was used.
COMPETING INTERESTS
KW, MGB, JH, JDGJ, WM and KS have filed a US patent application 62/538,020 based on this work.
ACKNOWLEDGMENTS
We would like to thank our co-workers, in particular Dr. Magdalena Krzymowska, for critical comments on the manuscript, Prof Ewa Zimnoch-Guzowska for discussions and help in the preliminary phase of the project. We would like to acknowledge Danuta Strzelczyk-Żyta and Izabella Barymow for the technical support, Baptiste Castel for sharing seeds of nrg1 N. benthamiana knockout plants and Brian Staskawicz for sharing seeds of eds1 N. benthamiana knockout plants.
Footnotes
Funding information: Research was supported by OPUS 8 (2014/15/B/NZ9/04384) and OPUS 14 (2017/27/B/NZ9/01803) grants from the National Science Centre, Poland, to Jacek Hennig. Kamil Witek was supported by BBSRC grants BB/L009293/1 and BB/P021646/1 to Jonathan Jones.
Author contributions:
M.G-B., K.W and J.H. performed most of the experimental work, data analyses, and writing; A. W., I.W-F., H.J., K.S. and K.M performed the research; J.J. and W.M. edited the article; J.H. supervised the work and assisted with writing.
REFERENCES




