

Abstract
Low-frequency vibrational excitations of proteins macromolecules in the terahertz frequency region are suggested to contribute to many biological processes such as enzymatic activity, molecular electron/energy transport, protein folding, and others. Two possible mechanisms of the formation of long-lived vibrational modes in protein were earlier proposed by H. Fröhlich and A.S. Davydov in the form of vibrational modes and solitary waves, respectively, to explain high effectiveness of energy storage and transport in proteins. In this paper, we developed a quantum dynamic model of vibrational mode excitation in alpha-helical protein interacting with environment. In the model we distinguish three coupled subsystems, i.e. (i) hydrogen bond peptide groups (PGs), interacting with (ii) the subsystem of side residuals which in turn interacts with (iii) environment (surrounding water) and is responsible for dissipation and fluctuation processes. It was shown that the equation of motion for phonon variables of the PG chain can be transformed to nonlinear Schrodinger equation for order parameter which admits bifurcation into the solution corresponding to weak damped vibrational modes (Fröhlich-type regime). A bifurcation parameter was shown to determine interaction of a protein with environment and in part, energy pumping to the protein due to its interaction. In the bifurcation region, a solution corresponding to Davydov soliton was shown to exist. The suggested mechanism of emergence of the macroscopic dissipative structures in the form of collective vibrational modes in alpha-helical proteins is discussed in connection with the recent experimental data on the long-lived collective protein excitations in the terahertz frequency region.
1. Introduction
Proteins are the most complex molecules in cells which consist of thousands of different atoms assembling in 3D ordered structures fitted well their biological functions such as enzyme activity, transport, structural, sensor, signalling, and others. These functions are realised through complex intraprotein dynamics including protein folding, conformation transitions, post-translation modification, and protein-protein interaction. This intrinsic dynamics is orchestrated by both intramolecular excitations and protein interactions with their cellular environment. Protein excitations related to their functions and structure include either separate atom groups and bonds or spread over large scale subdomains of proteins. High-frequency vibrations of atom groups and intrapeptide bonds’ (e.g. C=O stretching) excitations lying in IR optical spectrum, while low-frequency collective vibrational modes are ranged at the lowest end of the far IR spectrum in the terahertz frequency range (THz) [1]. Large-scale collective excitation such as normal vibration modes [2], [3], sound waves (phonons) [4], and coherent vibrational states [5], [6] are reported to be excited in protein molecules at the time scale from picosecond to nanosecond. A role of these collective modes in protein structure and functions such as folding, allosteric interaction, enzyme catalysis, intramolecular transport of energy/electron, and photosynthesis are the subject of intensive experimental and theoretical works [2], [3], [7]–[9].
The physical mechanism of collective coherent excitation far from thermal equilibrium in biosystems and mechanism of energy supply of this excitation were proposed and investigated by H. Fröhlich [10], [11]. He has developed a phenomenological nonlinear model of collective longitudinal vibrational-polar modes (phonons) in biological systems such as biological membranes and protein macromolecules and showed that they can be excited in the frequency range of 0.1 GHz - 1 THz. The mechanism of excitation of the long-lived coherent vibrations is defined by a continuous supply of metabolic energy, e.g. ATP hydrolysis. These type of collective vibrations (polarization waves) was suggested to play a key role in biological processes such as enzymatic catalysis and protein-protein interaction [12]. The general idea underlying this mechanism of long-lived coherent vibrations in biological systems lies in the assumption that when the energy is supplied to the system at a rate greater than that of energy loss, vibrational modes can condense into the lowest-frequency vibrational mode like the phenomenon of Bose-Einstein condensation. In contrast to Bose-Einstein condensation occurring in thermal equilibrium, Fröhlich condensation takes place in nonequilibrium condition at the energy supply and dissipation in nonlinear molecular structures [13]. Thus, excitation of Fröhlich mode in the form of coherent quantum dynamic structure can be considered as the emergence of a space-temporal dissipative structure in according to Prigogine’s theory [14] and governed by self-organization principals of Haken’s synergetics in nonlinear systems [15].
Another type of coherent excitation in the form of solitary waves was proposed theoretically by A.S. Davydov in order to unravel one of the central problems in bioenergetics related to a highly effective long-distance transport of energy, electrons and protons within macromolecules [16]. As a results, the theory of soliton transport of energy in the alpha-helical proteins has been developed in his works [16], [17]. It was shown that alpha-helical peptide structure plays a significant role in the formation of soliton travelling long distance with weak decay. The soliton model was applied to describe transport of energy released by ATP localised in the amide-I vibration (C=O bond oscillation) along peptide groups at room temperature [16]. Mechanism of localisation, storage, and transport of energy in the model is defined by nonlinear interactions of the high-frequency amide-I excitation (1667 cm−1) and low-frequency acoustic modes in the 1D protein structure. Alternative to the quantum Davydov model [17], a classical vibrational model of the interpeptide excitation dynamics was suggested by Takeno and soliton stability was studied at room temperature [18]. Many theoretical aspects of soliton dynamics in protein macromolecules including soliton stability, thermalization, solitons’ interaction and others were developed and investigated in various approximations [19]. The unified approaches to the description of both Davydov soliton mode and Fröhlich condensation mode excitations in proteins at the conditions sufficiently away from equilibrium were developed [5], [13], [20], [21].
Interest to the experimental observation of both of these types of the coherent excitations did not abate since the 1970s when these concepts have been developed and remain an area of intense experimental research and source of lively debate [22]–[24]. Searching for these two types of coherent excitations in protein macromolecules was carried out in IR absorption spectrum by Raman spectroscopy and in molecular excitation by THz radiation [25], [26].
In this work, we model collective excitations in alpha-helical peptide macromolecule interacting with environment in the framework of quantum dynamics. Following the Fröhlich approach [12] we distinguished three subsystems in the protein molecule interacting with its environment: (i) hydrogen bond peptide groups (PGs) interacting with (ii) the subsystem of side residuals of the protein which in turn interacts with surrounding water. Here we used the quantum mechanics approach developed earlier to model autolocalized states (polaron) of quantum quasiparticle of intramolecular excitation (valent electron) in a bimolecular chain of PGs interacting with the substrate [19], [27], [28]. In this approach, we investigate conditions at which molecular interaction among the PGs, side residual chain and protein’s environment causes the coherent excitations of vibrational and soliton types in alpha-helical protein structures.
2. Phonons in a one-dimensional chain of hydrogen-bonded peptide groups interacting with side radicals
The secondary protein structure, alpha-helix is formed as a result of folding up of polypeptide chain in a helix due to the interaction of amino acid residues. This interaction determines space periodicity of the secondary structure in proteins and its stability is ensured by hydrogen bonds between NH- and CO- groups of the backbone (Fig. 1a). The alpha-helix has a form of a coil and its inner part consists of tightly twisted backbone with amino acid radicals directed outwards (Fig. 1b). Electrical charge distribution in the peptide group (PG) forms its electrical dipole moment equal to 3.5 D and directed along H-bond [17].

(a) The chain of hydrogen-bonded peptide groups. (b) Structure of alpha-helical protein where one of the three hydrogen-bonded PG chains with side radicals (R) is shown. Hydrogen-bonds are depicted by dots.
Consider a one-dimensional chain of hydrogen-bonded peptide groups with hydrogen bonds between (NH)- and CO-groups (Fig. 1). Assume intrinsic motion of proton and the rest of the PGs (NCO). Define equilibrium positions of the PGs in the l-site z=la (l=0, 1, 2,…) along the alpha-helix (the z-axis), where a= 5.4 Å is chain spacing. Denote displacement of the atoms from equilibrium positions of the PG in the l-site by ξl,1 for hydrogen and ξl,2 for the rest of the PGs. In harmonic approximation, potential energy of interaction between atoms of the nearest PGs and nearest radicals R is expressed by a quadratic form:
 where χ1,2 and χ2,1 are elastic coefficients of hydrogen and valence bonds respectively; χ0,1 and χ0,2 are elastic coefficients of interaction of protons and group NCO with the side chain radicals, respectively. Choose the following cyclic boundary condition for the PG chain
where χ1,2 and χ2,1 are elastic coefficients of hydrogen and valence bonds respectively; χ0,1 and χ0,2 are elastic coefficients of interaction of protons and group NCO with the side chain radicals, respectively. Choose the following cyclic boundary condition for the PG chain
 where N is a number of GPs in the chain.
where N is a number of GPs in the chain.
To write the Hamiltonian of the PG chain, express the operators of atom displacement from equilibrium positions through the operators of the creation  and annihilation bk,s of phonons as
and annihilation bk,s of phonons as
 where the operators of the creation and annihilation satisfy the commutative relationships
where the operators of the creation and annihilation satisfy the commutative relationships

Here δ is the Kronecker symbol, k is the wave number taking N values in the first Brillouin zone
 and index s points to either acoustic (s=1) or optic (s=2) phonons.
and index s points to either acoustic (s=1) or optic (s=2) phonons.
The Hamiltonian of phonons in the PG chain can be written in the harmonic approximation as
 where two functions Ωs(k) define frequencies of acoustic and optic branches of the vibration in the chain with s = 1 and 2 respectively. The dispersion relationship Ωs(k) for the PG chain with interaction defined by eq. (1) can be obtained in the form
where two functions Ωs(k) define frequencies of acoustic and optic branches of the vibration in the chain with s = 1 and 2 respectively. The dispersion relationship Ωs(k) for the PG chain with interaction defined by eq. (1) can be obtained in the form
 where
where
 and
and

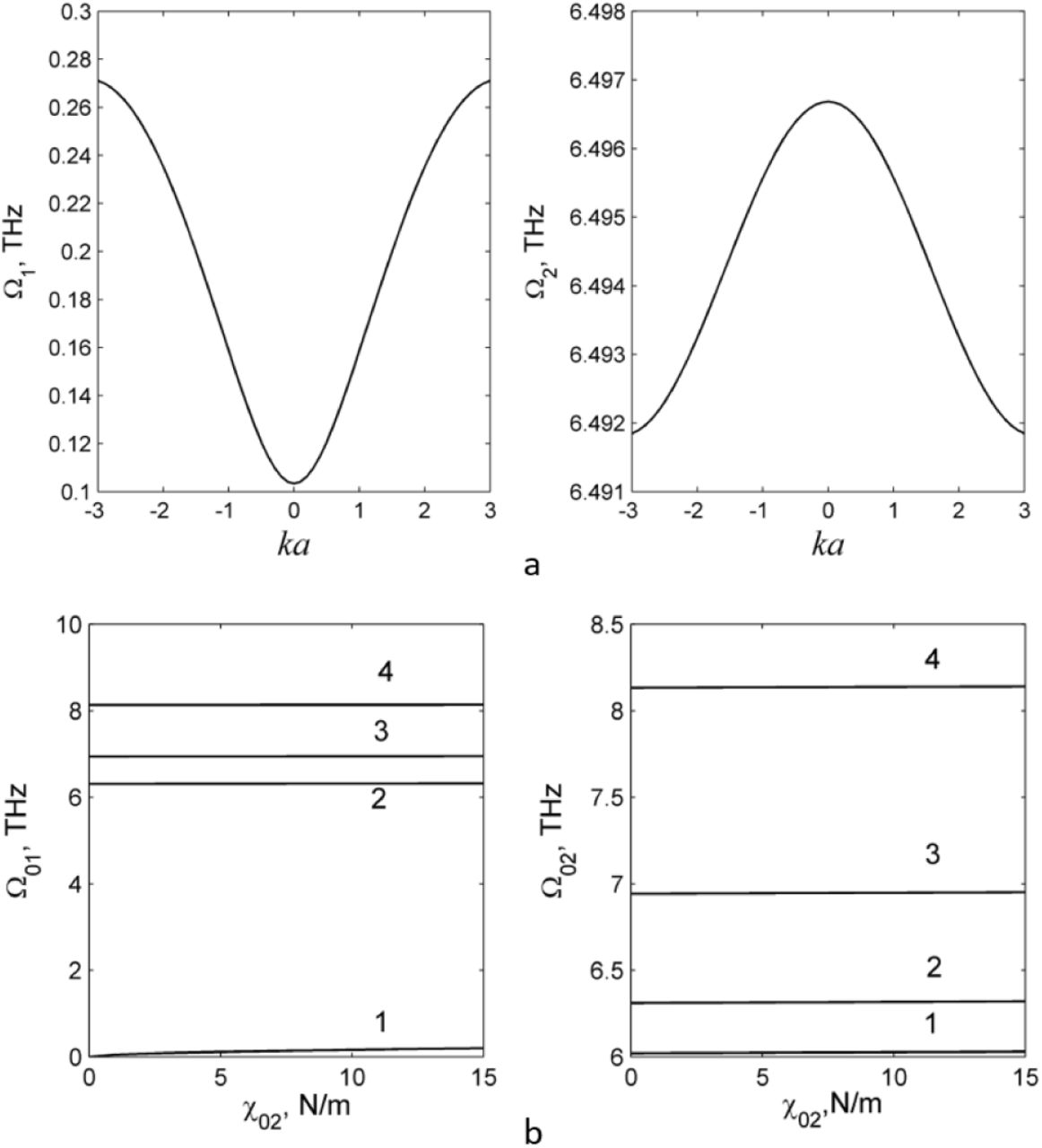
Here m1=mp and m2 = 41.7mp are mass of proton and the PG group respectively. Dispersion curves calculated at the values of elastic constants of the PG chain [19] are shown in Fig. 2. As seen, according to dispersion equation (8), the PG chain with interaction (1) obeys a narrow band of the normal modes of the vibrations Ωs(k) in terahertz frequency range.
{kind=link}
{kind=link}
Dispersion curves for the chain of hydrogen-bounded PGs at the following elastic constants [19]. (a) s=1, χ1,2 = 13.0 Nm−1, χ2,1 = 17.0 Nm−1, χ0,1 = 5.0 Nm−1, χ0,2 = 0 Nm−1.(B) s=2 χ1,2 = 13.0 Nm−1, χ2,1 = 17.0 Nm−1, χ0,1 = 5.0 Nm−1, χ0,2 = 0 Nm−1. (b) Dependence of the frequency Ω01 (C) and Ω02 (D) (k=0) on constant χ02 at the fixed values of elastic constants χ1,2 = 13.0 Nm−1, χ2,1 = 17.0 Nm−1 and different constant χ0,1: line 1 - χ0,1 = 0.0 Nm−1, line 2 - χ0,1 = 3.0 Nm−1, and line 3 - χ0,1 = 25.0 Nm−1. Dispersion curves were calculated in MATLAB package (MathWork Inc).
Similarly introduce the phonon Hamiltonian for the radical system of the PG chain which can be represented as a system of N oscillators of mass M having own frequencies of oscillation Ωt(q)
 where
where  and cq,t are the operators of the creation and annihilation of phonons in the radical chain which corresponds to the displacement of the radicals from their equilibrium positions Çl. Ωt(q) are the phonon frequency of type t in the radical chain.
and cq,t are the operators of the creation and annihilation of phonons in the radical chain which corresponds to the displacement of the radicals from their equilibrium positions Çl. Ωt(q) are the phonon frequency of type t in the radical chain.
Consider interaction of the phonons in the PG and radical chains in the alpha-helical protein structure by adding the following anharmonic operator of interaction to the Hamiltonian of non-interacted chains
 where
where
 Substitution of eqs. (3) and (10) into eq. (9) gives the operator of anharmonic perturbation in the form
Substitution of eqs. (3) and (10) into eq. (9) gives the operator of anharmonic perturbation in the form
 where
where

The matrix elements (13), which are different from zero on the functions of occupation numbers, corresponds to the processes which occur with energy and momentum conservation without consideration of umklapp process

3. Interaction of alpha-helical protein with environment (heat reservoir)
Consider interaction of the alpha-helical protein with its environment, surrounding water, at non-zero temperature. This protein-water interaction defines dissipation processes in the system and contributes to the structure and functions of proteins. In the model, we represent surrounding water by a large set of harmonic oscillators interacting with the protein molecule and suggest that this system possesses collective excitation modes with amplitude dynamics similar to that of an oscillators’ ensemble. Note that the collective vibrational subpicosecond dynamics (phonons) of the hydrogen bond network was observed in hydration shells of protein molecules in terahertz and IR spectra [29].
Similarly to eqs. (3) and (10), describe oscillators’ dynamics using the operators of the creation  and annihilation Bp,t of phonons with the wave number p and frequencies ωt(p). Then the energy operator of the surrounding water can be written as a sum of energy operators of the single oscillators in the form
and annihilation Bp,t of phonons with the wave number p and frequencies ωt(p). Then the energy operator of the surrounding water can be written as a sum of energy operators of the single oscillators in the form

To calculate energy of joint action of reservoir’s oscillators on the protein macromolecule, WB, we assumed that each oscillator contributes linearly to the energy WB and interacts only with the radical chain of the alpha-helical protein. So, following [18], we suggested that the PG chain interacts with the environment through radicals R (Fig. 1B). Then, interaction operator WB can be written as

Parameter σtt′(g, p) defines a strength of interaction between reservoir’s oscillators of types t′ and the side chain oscillators of type t. Note that operator (15) describes a wide class of relaxation mechanisms related to collective excitation [19].
For simplicity, let drop below indexes which define phonon type, and indexes s, t, and t’ of variables will thereafter mean that the specific variable belongs either to the PG chain or the radicals’ chains or reservoir respectively.
4. Equation of motion
One of the features of a self-organisation behaviour of complex systems is the occurrence of the instability of either one or several variables (dynamic modes) at some critical conditions
[15]. If the rest of the modes damp the different exclusion procedures of the stable variables can be applied at the modelling. It follows that the system behaviour as a whole is defined by the behaviour of few unstable variables which govern all the damped modes. In real systems, a hierarchy of relaxation times takes place most often that allows applying adiabatic approximation for the exclusion of the fast-relaxing variables. In the case of protein molecules, the fast-relaxing variables  and cg relate to the radical subsystem which directly interact with the environment (structured water) surrounding a native protein and supporting stability of its structure.
and cg relate to the radical subsystem which directly interact with the environment (structured water) surrounding a native protein and supporting stability of its structure.
To exclude fast-relaxing variables  and cg, we use Heisenberg equation for these operators
and cg, we use Heisenberg equation for these operators
 where
where  and H is the energy operator of the macromolecule-reservoir system
and H is the energy operator of the macromolecule-reservoir system

Substitution of HP, HC, HB, WP, and WB in to eq. (17) and then to eq. (16) gives the equation of motion for the operator cg
 and the identical equation for the operator
and the identical equation for the operator  . Then let require
. Then let require

The following relationship can be obtained for  and cg from eqs. (18) and (19)
and cg from eqs. (18) and (19)

Using eqs. (20) and commutation relations (4) for the Bose-operators, we get the energy operator in the form
 where the energy operator of protein macromolecule is expressed only through the variables related to the chain of PGs
where the energy operator of protein macromolecule is expressed only through the variables related to the chain of PGs

Here

Hamiltonian operator of the reservoir HB is written in the form
 where
where

The energy of interaction of the alpha-helical protein with the reservoir HPB after exclusion of the variables related to the side chain is defined by
 where
where

Equation of motion for dynamic variables of the PG chain and reservoir can be derived using Heisenberg equation (16) for the operators bk,  , Bp, and
, Bp, and  the Hamiltonian operator (21), and eqs.(22)–(27)
the Hamiltonian operator (21), and eqs.(22)–(27)
 and
and

At the next step, we use the obtained eqs. (28) and (29) to derive the motion equation for the modes growing up to macroscopic values and define system dynamics near unstable stationary points. Moreover, we describe dissipation in the system and express fluctuating forces which are coursed by the interaction of the protein macromolecule with environment and enable the systems to overcome an unstable threshold and reach new dynamic states.
5. The Langevin equation for generalized coordinates of the macromolecule
As known, all basic (microscopic) equations of motion are invariant with respect to time reversal, that is the motion is entirely reversal. Although dissipative forces, violating this invariance, cannot be expressed in the original equations, under certain assumptions the Langevin equations can be derived from the Heisenberg equation for a system interacting with a reservoir represented by a set of harmonic oscillators [30].
So far, we considered systems of quantum oscillators as the model of alpha-helical protein molecule interacting with the reservoir. However, these systems can be considered as the classical ones. This can be justified by that a number of phonons in strong exciting modes defining protein dynamics and a number of oscillators in the reservoir system are significantly larger than unit. This allows us to represent phonon amplitudes by c-numbers and substitute operators bk,  , Bk, and
, Bk, and  in eqs. (28) and (29) for c-numbers βk,
in eqs. (28) and (29) for c-numbers βk, ,Bk, and
,Bk, and  respectively. Amplitude βk(t) can be considered as the generalized coordinates with corresponding generalized momentum
respectively. Amplitude βk(t) can be considered as the generalized coordinates with corresponding generalized momentum  . Moreover, coefficients, defining intensity of phonon interaction of the different subsystems in the model, are assumed to weakly depend on the phonon momentum. Then, eqs. (28) and (29) can be integrated as the classical ones that gives the solution of eq. (29) in the form:
. Moreover, coefficients, defining intensity of phonon interaction of the different subsystems in the model, are assumed to weakly depend on the phonon momentum. Then, eqs. (28) and (29) can be integrated as the classical ones that gives the solution of eq. (29) in the form:
 where Bp(0) is the initial value of amplitude Bp(t) at t=0.
where Bp(0) is the initial value of amplitude Bp(t) at t=0.
Introduce new variable 
 and use thereafter the previous notation βk(t). Then apply adiabatic approximation commonly used in the modelling of cooperative systems, i.e. the relaxation times of the strong exciting phonon modes become longer in comparison with the typical relaxation times for reservoir variables. This allows factoring out the preexponential term in eq. (30) and obtain it in the form
and use thereafter the previous notation βk(t). Then apply adiabatic approximation commonly used in the modelling of cooperative systems, i.e. the relaxation times of the strong exciting phonon modes become longer in comparison with the typical relaxation times for reservoir variables. This allows factoring out the preexponential term in eq. (30) and obtain it in the form
 Integrals in eq. (32) gives
Integrals in eq. (32) gives
 where P is a symbol of principal value. Finally, Bp(t) is obtained in the form
where P is a symbol of principal value. Finally, Bp(t) is obtained in the form

Substitution of eq. (34) into eq. (28) gives us equation for βk(t)

Eq. (35) is the Langevin equation for phonon amplitudes βk(t)
 where the following variables are introduced
where the following variables are introduced

 and
and

Function Fk(t) (39) can be considered as a random force with the correlator:

Assume that at the initial time, amplitudes β(0) and B(0) are not correlated, i.e.
 and
and
 where
where
 and
and

Here θ = 1/kBT, where T is environment (reservoir) temperature.
Substitute eqs. (41)–(44) into eq. (40) and take into consideration that the main contribution in a sum in eq. (40) is given by terms Λ(k,p) = 0 at not too small values of a difference (t – τ). Then finally obtain

It can be shown that after averaging in eq. (38) for γk with consideration of eq. (44) and substituting this result into eq. (45), the value Dk can be written in the form

6. Vibrational dynamics of alpha-helical protein in a long wave approximation
Consider vibrational modes in the alpha-helical protein at the various parameters characterising its environment. We applied a long wave approximation to derive the equation for undamped modes which grow up to macroscopic values and investigate system dynamics in the vicinity of an unstable point. In a long wave approximation, for small values ka << 1 and according to eq. (8), phonon frequencies can be written in the form
 where
where

Here Ω0s and B(0) are defined by eq. (8) at k= 0. Note, that the value
 can be found from the dependence of frequencies Ω0s on the elastic constants shown in Fig. 2.
can be found from the dependence of frequencies Ω0s on the elastic constants shown in Fig. 2.
As in the long-wave approximation the value Λ(k,p) = Λ(p) does not depend on k, we turn to continuum limit in eq. (36) after Fourie transformation by multiplication of all terms in eq. (36) by N−1/2eikz and summing up over k according to eq. (47). In continuum limit, eq. (36) for photon modes with dispersion relation (47) takes the form
 where
where
 and
and

Solution of eq. (50) can be represented in the form
 where ρ = z – z0 – Vs, Vs is the velocity of excitation motion along the PG chain, and the real amplitude Φs(ρ) satisfies the following normalization condition
where ρ = z – z0 – Vs, Vs is the velocity of excitation motion along the PG chain, and the real amplitude Φs(ρ) satisfies the following normalization condition

Consider solutions of eq. (50) at weak damping, i.e. when γs ≈ 0. In this condition, according to eq. (46) and fluctuation-dissipation theorem, fluctuations are small and can be neglected. Then eq. (50) takes the form:
 where Ξs is a spectral parameter connected with phonon energy by the equation:
where Ξs is a spectral parameter connected with phonon energy by the equation:

Eq. (54) has solution ϕs(ρ) = const:
 and
and
 where parameter λs defines interaction of the PGs with its environment. At changing λs the oscillation modes become unstable and their amplitudes ϕs(ρ) play a role of the order parameters. Solutions (56) and (57) obtained under conditions of the smallness of dissipation in the system and absence of fluctuations. Thus, living time τs of the dynamic modes of the PG chain corresponding to nontrivial solutions is less than the inverse-time of relaxation
where parameter λs defines interaction of the PGs with its environment. At changing λs the oscillation modes become unstable and their amplitudes ϕs(ρ) play a role of the order parameters. Solutions (56) and (57) obtained under conditions of the smallness of dissipation in the system and absence of fluctuations. Thus, living time τs of the dynamic modes of the PG chain corresponding to nontrivial solutions is less than the inverse-time of relaxation

So, dynamics of this system is defined by weak damped (long-living) phonon modes. More detailed analysis of the dynamic equation of the type (50) is given in [31], where it was shown in part that the right hand side of this equation can be obtained from the potential

This allows writing and solving the corresponding Fokker-Planck equation and then finding a distribution function for the phonons in coherent excitation state
 where parameter Ds defines an intensity of fluctuating force according to eqs. (45) and (46) at k = 0 for both phonon branches. From eq. (60) follows that fluctuations enable the system to switch to a new state. The role of fluctuations is much significant at the transition of the system to an unstable mode at λs ≤ 0 when, as known, fluctuations sharply increase [15]. At the sign change of parameter λs, solution Φs(ρ) = 0 remains one of the solutions of eq. (54). Transition of the system to the new states, corresponding to nontrivial solutions Φs(ρ) ≠ 0, is possible as a result of an action of external factors including fluctuations.
where parameter Ds defines an intensity of fluctuating force according to eqs. (45) and (46) at k = 0 for both phonon branches. From eq. (60) follows that fluctuations enable the system to switch to a new state. The role of fluctuations is much significant at the transition of the system to an unstable mode at λs ≤ 0 when, as known, fluctuations sharply increase [15]. At the sign change of parameter λs, solution Φs(ρ) = 0 remains one of the solutions of eq. (54). Transition of the system to the new states, corresponding to nontrivial solutions Φs(ρ) ≠ 0, is possible as a result of an action of external factors including fluctuations.
Eq. (54), being nonlinear Schrodinger equation, besides the solutions considered above has a solution in the form of solitary wave (soliton) travelling along the z-axis and satisfying normalization condition (53). For any positive Qs, eq. (54) has a normalized partial solution in the form
 where z0 is the soliton centre, Vsol is soliton velocity, and
where z0 is the soliton centre, Vsol is soliton velocity, and

In the presence of dissipation, when γs ≠ 0, a solution of eq. (50) in view of eqs. (51) and (52) is written in the form:
 where
where

The region occupied by soliton, soliton’s width, is defined by equation

7. Discussion: Vibrational modes and self-organization in alpha-helical protein structures
The theoretical investigation of the dynamics of the alpha-helical protein interacting with environment showed that the equation for phonon dynamics in the protein is essentially defined as the equation for the order parameter that admits bifurcation of its solutions. Thus, this system can function in different dynamic modes which are defined by the behaviour of order parameters. Switch between the vibrational modes occurs at a change in the bifurcation parameter λs which in according to eq. (37) determines interaction of the macromolecule with its environment and phonon pumping into the molecule. The PG chain dynamics below and above of the switching threshold are significantly different. At λs > 0, the system is characterised by the absence of excited modes, and the PGs fluctuate that results in a zero-mean amplitude of the phonon modes. At λs < 0, behavithe of the system changes so that either one or several vibrational modes become unstable and their amplitudes grow up to macroscopic values. This transition to a state when the energy is condensated in a set of the low-frequency vibrational modes can lead to the excitation of coherent vibrations in proteins in terahethe frequency range. Thus, dynamithe c behaviour of the macromolecule is defined by both fluctuations governed by the dissipation in the system and switching regime as a result of an order parameter change controlled by environment.
As shown, the condition of the vibration mode excitation in the model is the energy pumping to the system that agrees with the condition of Fröhlich vibrational mode excitation which can be formed at a high rate of energy channelling into the lowest frequency vibration of protein macromolecules [12]. We suggest that the vibrational energy may be channelled to the protein through its interaction with the environment by mechanism considered in the model. A key role in this energy pumping can be played by the ordered water clusters bound to the hydration shell of proteins [9], [32] and the hydrogen bond network of water molecules which was suggested to possess the collective vibrational sub-picosecond dynamics and propagating phonon-like modes in terahertz and IR spectra [7], [29].
Another condition of the collective vibrations obtained in our model is a low rate of the relaxation of the PG vibrations (eq. (59)). The reasonable mechanisms of the reduced relaxation of protein collective motion, which were discussed in connection with the Fröhlich condensation in biological structures, can be linked again to the interaction of macromolecules with their environment e.g. with ordered water clusters possessing slow dynamics [33], [34].
According to our model, space-temporal structures of the type of solitary waves (Davydov regime) can emerge in the PG chain. The following features of the Davydov regime in the PG chain were defined in this work. First, soliton formation is governed by phonon-phonon interaction between PGs and radical chain in alpha-helical proteins, and second, Davydov regime realises at the values of bifurcation parameter when excitation of vibrational mode (Fröhlich-like regime) occurs in the system. It was suggested that cooperative behaviour of the PGs may lead to the formation of a giant oscillating dipole [12] due to a large dipole moment of the PGs that is likely to be associated with some protein function such as selective forces, protein-ligand interaction, molecular recognition, and catalytic enzyme activity. On the other hand, soliton formation including acoustic solitons [16] can lead to energy and electron transport along alpha-helical protein structures due to either exciton or electron-phonon interaction [27], [35]. A similar mechanism of collective excitation was explored in the theoretical description of energy transport in microtubule [36], [37].
The obtained results on the dynamics of alpha-helical protein interacting with environment showed that vibrational mode instability induced by a change in the protein-environment interaction parameter can cause a formation of new macroscopic space-temporal structures in the system. Joint action of random and deterministic forces can lead to the switching of the system to a dynamic state characterised by cooperative behaviour of subsystems (subunits) making up protein macromolecule. One of the factors of this selforganisation was shown to be the nonlinear interaction of several subsystems in protein interacting with environment, i.e. the regular chains of hydrogen bonds, side chain radicals, and hydrogen bond network of water molecules.
Progress in terahertz spectroscopic techniques and their combination with other spectroscopic methods led to a revival of interest to experimental observation of Fröhlich coherent excitation in biological structures [26], [38]. For example, experimental confirmation of a long-range quantum coherent state in proteins comprised of alpha-helical structures were undertaken in experiments with interaction of biomolecular structures with microwave and terahertz radiation [25]. Authors used X-ray crystallographic methods to visualise low-frequency collective vibration modes in the lysozyme protein crystal irradiated by 0.4 THz radiation [25]. Estimated microsecond lifetime of these excited longitudinal compression modes in the alpha-helix turned out to be significantly longer then femto-and nanosecond time scale of the decay of intramolecular vibrations due to their interaction with environment (thermalization). Authors refer this underdamped vibration to Frohlich condensation mode excited by terahertz radiation. The existence of these persisting motions indicates that damping and intermode coupling are weaker than previously assumed [3].
Vibrational wave packets with a long lifetime over 500 picoseconds were observed in bacteriorhodopsin exposed by picosecond far IR sources [34]. Authors discussed a possible mechanism of slow relaxation due to quantum effects of restricted interaction of the low-frequency collective modes with solvent and suggested a link between collective undamped collective vibration and the conformational transitions in proteins enriched by alpha-helical structures.
Experimental investigation of the coherent vibrational dynamics in proteins was intensified by the observation of long-lived coherent excitonic states in light-harvesting proteins in photosynthetic bacteria [8]. The results of 2D IR coherent spectroscopy suggests that the coherent vibrations in photosynthetic pigment–protein complexes contribute to the effective electron and energy transport due to the electron-vibrational couplings [6], [39]–[41]. Note that a role of coherent longitudinal electric modes (polarization waves) of lowest frequency (0.01 THz – 1 THz) in a storage of light energy in photosynthesis was proposed by Fröhlich in 1968 [10].
As seen current experiments using new experimental techniques in terahertz range and 2D IR spectroscopy can provide new valuable data on the molecular mechanisms of the quantum coherent vibration excitations in protein one of which was suggested in the paper. Together theoretical and experimental investigation of the conditions for the triggering of long-live, low-frequency vibrations in alpha-helical protein interacting with environment may help to elucidate a role of one-dimensional dynamics in extraordinary effectiveness of protein biological functions such as energy storage, transport and transformation.
Conflict of interest
The authors report no conflict of interest.
Acknowledgements
This work was supported by personal support to AG from Scottish Informatics and Computer Science Alliance (SICSA).
References




