

Abstract
Background Among hematophagous arthropods, ticks transmit the greater variety of pathogens of public health and veterinary importance whose (re)-emergence is recognized worldwide. However, the epidemiological situation of the Caribbean area with regard to tick-borne diseases is poorly documented, mainly focusing on livestock pathogens such as Ehrlichia ruminantium, Babesia (bovis and bigemina) and Anaplasma marginale. These observations underline the need to undertake new large-scale epidemiological surveys to better assess the distribution of tick-borne pathogens and anticipate the risk of (re)-emergence of tick-borne diseases in these areas. To ease and reduce the cost of such large-scale surveys, the development of a fast and cheap high-throughput detection technics is desirable.
Methods In this study, we first implemented a high-throughput microfluidic real-time PCR (BioMark™ dynamic arrays, Fluidigm Corporation) adapted for the large-scale screening of tick-borne pathogens. The system developed here includes 57 designs allowing both the screening of bacteria and protozoans potentially circulating in the West Indies (5 bacterial genera, 30 bacterial species, 1 protozoan phylum, 2 protozoan genera and 17 protozoan species), and the molecular identification of three tick species mainly involved in tick-borne pathogens transmission in the Caribbean (Amblyomma variegatum, Rhipicephalus microplus and Rhipicephalus sanguineus sensu lato). Then, using the new high-throughput microfluidic real-time PCR system, we performed an exploratory epidemiological study on 132 specimens of Amblyomma variegatum and 446 Rhipicephalus microplus collected in Guadeloupe and Martinique.
Results We successfully detected tick-borne pathogens expected to be present in the area – Ehrlichia ruminantium, Rickettsia africae, Anaplasma marginale, Babesia bigemina, Babesia bovis, Theileria velifera and Theileria mutans – as well as unsuspected pathogens and microorganisms belonging to the genera Anaplasma, Ehrlichia, Borrelia, and Leishmania.
Conclusions We demonstrated the ability of the Biomark system to give a rapid overview of the pathogens/microorganisms diversity present in ticks, thus opening new research perspectives on tick-borne pathogens epidemiology in the Caribbean. Our study demonstrated how high-throughput microfluidic real-time PCR technology is a major improvement in large-scale epidemiological studies, giving a rapid overview of the tick-borne pathogens and microorganisms present in ticks in a given area.
Background
Among hematophagous arthropods, ticks transmit the greatest variety of pathogens of public health and veterinary importance which incidence is growing worldwide (Dantas-Torres et al., 2012). The French West Indies, Guadeloupe and Martinique islands, are located in the heart of the Caribbean Neotropical Zone, a cosmopolite area characterized by a tropical climate, intercontinental exchanges, and animal movements (legal or illegal trade and bird migration) that are favorable for ticks and tick-borne pathogens (TBPs) introduction and dispersal (George et al., 2002). The epidemiological situation of the Caribbean area with regard to the diversity of tick species and tick-borne diseases (TBD) is poorly documented (Gondard et al., 2017). Amblyomma variegatum also known as Tropical Bont Tick (TBT) in the Caribbean and Rhipicephalus microplus (“Cattle tick”) are the two main tropical livestock pests since their introduction in the Caribbean through the importation of infested animals from Africa and Asia during the 18th-19th century (Barré and Uilenberg, 2010). Both tick species are present in the French Antilles where they are involved in the transmission of TBPs of medical and veterinary importance (Morel, 1967; Uilenberg et al., 1983; Camus and Barré 1995; Alonso et al., 1992; Parola et al., 1999). R. microplus, a one-host tick highly specific of cattle, is responsible for the transmission of Anaplasma marginale, Babesia bovis and Babesia bigemina, causing bovine anaplasmosis and babesiosis, respectively. These pathogens are considered as endemic in the Caribbean (Camus and Montenegro-James 1994; Camus and Barré 1995). A. variegatum, is a three-host tick species, with immature stages that can parasitize a large host range, including rodent, mongoose, bird, and adult stage that is more specific to cattle (Barré, et al., 1988). This tick species can transmit Ehrlichia ruminantium the causative agent of heartwater, a fatal ruminant ehrlichiosis. Although A. variegatum is present in both Martinique (mainly in the South) and Guadeloupe (widespread), E. ruminantium is only reported in Guadeloupe (Vachiéry et al., 2008). This tick species is also vector of Rickettsia africae, common in the Caribbean, which can induce human rickettsiosis called African Tick Bite Fever (Parola et al., 1999, 2003; Kelly et al., 2010). Despite high level of tick infection and seroprevalence in human and cattle sera, to date only two human cases of African Tick Bite Fever related to travelers returning from Guadeloupe have been reported (Parola et al., 1999; Raoult et al., 2001). Finally, A. variegatum is also involved in Theileria mutans and Theileria velifera epidemiology, two cattle parasites of low and no virulence, respectively (Uilenberg 1983; Alonso et al., 1992). The majority of the epidemiological information available on TBPs in the Caribbean is based on serological studies in animals or humans, or molecular biology testing (PCR, nested PCR). These studies did not survey nor discriminate the whole repertoire of infectious agents since they were often limited to the detection of some well-known pathogens (Estrada-Peña et al., 2013; Cabezas-Cruz et al., 2018). Thus, new insights into tick and TBPs epidemiology are needed to address TBD prevalence and (re-) emergence in the Caribbean.
The recent development and use of a microfluidic real-time PCR system for rapid and concomitant detection of a large panel of TBPs in European ticks has paved the way for a promising wider surveillance capacity (Michelet et al., 2014; Dupraz et al., 2017; Gioia et al., 2018; Gondard et al., 2018; Malmsten et al., 2018). In this study, we adapted and designed a new microfluidic real-time PCR system adapted to the simultaneous screening of the main bacteria and protozoans potentially transmitted by ticks in the Caribbean area in 95 DNA tick samples in a single experiment. Not only does the system allowed the direct detection of 49 bacterial species but it also allowed in a single experiment a broader surveillance ability of microorganisms, potentially pathogenic, by targeting the main bacterial and protozoan genera involved in human or animal vector-borne diseases (one protozoan phylum, eight bacterial and protozoan genera). In addition, the system allowed the molecular identification of the three well known tick species involved in TBDs in the Caribbean. Finally, we used the new high-throughput detection tool, to conduce a large scale screening of TBPs in 132 A. variegatum and 446 R. microplus collected in Guadeloupe and Martinique. We demonstrated the ability of the system to detect well-known TBPs occurring in the French Antilles, as well as unsuspected TBPs and potential new microorganisms. This new method can improve considerably the monitoring capacity of emergent and non-emergent TBPs through large-scale surveys in the Caribbean area.
Methods
Tick collected in Guadeloupe and Martinique
The ticks used in this study were collected within the framework of two distinct epidemiological surveys conducted in Guadeloupe (between February 2014 and January 2015) and Martinique (between February and March 2015), respectively. In Guadeloupe, adult ticks (any species, any sex) were collected on 40 bovines originating from 22 different herds that were sampled in nine localities situated in six different biotopes (urban area, dry coastal regions, valleys and hills, evergreen seasonal forest, sub-mountainous rain forest, swamp forest). In Martinique, engorged females of R. microplus only were collected from cattle in 29 farms participating in a study on acaricide resistance in ticks. All ticks were collected from cattle with the permission from farmers and cattle owners. Ticks were morphologically identified at the species level (Walker et al., 2003). A total of 578 adult ticks were included in the study: 132 A. variegatum and 165 R. microplus from Guadeloupe and 281 R. microplus from Martinique (see maps, Fig 2). GPS coordinates of tick collection sites are available in Additional file 1. All the ticks were partially engorged, and then conserved at -80°C.
DNA extraction
For 20 mg of ticks, 1 mL of recently prepared PBS 1X was added to the sample, then ticks were washed by gentle shaking over 2-3 min at 7 Hz/s in a Tissue Lyzer (Qiagen, Germany). After discarding the supernatant, ticks were frozen at -80°C during 15-20 min. Then a steel ball was added and samples were crushed twice during 2 min at 30 Hz/s with the Tissue Lyzer (Qiagen, Germany). 450 µL of fresh PBS 1X were added to the samples. Samples were vortexed during 10 s, and then centrifuged during 2-3 min at 8000 g. Finally, 20 µL of Proteinase K were added to 180 µL of crushed tick sample and DNA was extracted using the NucleoSpin® 96 virus Core kit (Macherey-Nagel, Germany) and the automatic platform Biomek4000 (Beckman Coulter). This protocol allows the simultaneous extraction of both DNA and RNA. Total nucleic acid per sample was eluted in 160 µl of rehydration solution and stored at −80°C until further use. A. variegatum ticks were individually extracted. R. microplus ticks were extracted both individually and in pools of two to four adult specimens when too small to be treated individually.
Assay design
The list of pathogens to be monitored, the sets of primers and probes required for their detection, as well as the targeted genes are shown on Table 1. Some of the oligonucleotides were specifically designed for the purpose of this study; the others came from Michelet et al., 2014. The newly developed oligonucleotides were validated on dilution range of positive controls including cultures, plasmids or DNA samples (Table 1) by real-time TaqMan PCRs assays on a LightCycler® 480 (LC480) (Roche Applied Science, Germany). Real-time PCR assays were performed with LightCycler® 480 Probe Master Mix 1× (Roche Applied Science, Germany), using 200 nM of primers and probes in a final volume of 12 µl and 2 µl of control DNA were added. The thermal cycling program was as follows: 95°C for 5 min, 45 cycles at 95°C for 10 s and 60°C for 15 s and one final cooling cycle at 40°C for 10 s.
Pre-amplification of DNA samples
All the DNA samples were subject to preamplification in order to enrich the pathogen DNA content comparatively to tick DNA. The Perfecta® PreAmp SuperMix (Quanta Biosciences, Beverly, USA) was used for DNA pre-amplification, following the manufacturer’s instructions. All the primers were pooled (except the ones targeting the tick species) with a final and equal concentration of 45 nM each. Pre-amplification reaction was performed in a final volume of 5 µL containing 1 µL of PerfecTa PreAmp SuperMix (5X), 1.25 µL of pooled primers mix, 1.25 µL of DNA and 1.5 µL of MilliQ water with one cycle at 95°C for 2 min, 14 cycles at 95°C for 10 s and 60°C for 3 min. At the end of the cycling program the reactions were diluted 1:10. Pre-amplified DNAs were stored at −20°C until use.
High-throughput microfluidic real-time PCR
High-throughput microfluidic real-time PCR amplifications were performed using the BioMark™ real-time PCR system (Fluidigm, USA) and 96.96 dynamic arrays (Fluidigm, USA), that allows to perform up to 9,216 individual reactions in one run (Michelet et al., 2014). Real-time PCRs were performed using 6-carboxyfluorescein (FAM) and black hole quencher (BHQ1)-labeled TaqMan probes with TaqMan Gene Expression Master Mix (Applied Biosystems, USA) following the manufacturer’s instructions. Cycling conditions were as follows: 2 min at 50°C, 10 min at 95°C, followed by 40 cycles of 2-step amplification of 15 s at 95°C, and 1 min at 60°C. The BioMark™ Real-Time PCR system was used for data acquisition and the Fluidigm Real-time PCR Analysis software for Ct values determination. Three kind of control were used per chip for experiment validation: a negative water control to exclude contaminations; a DNA extraction control, corresponding to primers and probe targeting a portion of the 16S rRNA gene of ticks; and an internal control to check the presence of PCR inhibitors made of a DNA of Escherichia coli strain EDL933 added to each sample with specific primers and probe targeting the eae gene (Nielsen and Andersen, 2003).
Infection rates of ticks from the French Antilles
According to the tick species and the island of origin, for each detected pathogen, infection rates (the proportion of infected ticks divided by the total number of ticks analyzed) were calculated. The majority of the samples were single specimens of ticks. When ticks were too small to be treated individually, they were pooled by two to four specimens. Thus, among the 523 samples analyzed, 47 consisted of a pool of two to four tick specimens. The final estimation of the infection rate also includes the pools and is therefore expressed as the minimal (assuming at least one positive ticks in the pools) and maximal (assuming all ticks positive in the pools) proportions of infected ticks out of the total number of ticks analyzed.
PCRs and sequencing for results confirmation
Conventional PCR/Nested PCR using primers targeting different genes or regions than those of the BioMark™ system were used to confirm the presence of pathogenic DNA in some field samples and positive controls (Table 2). Amplicons were sequenced by Eurofins MWG Operon (BIOMNIS-EUROFINS GENOMICS, France), and then assembled using BioEdit software (Ibis Biosciences, Carlsbad, CA, USA). An online BLAST (National Center for Biotechnology information) was used to compare results with published sequences listed in GenBank sequence databases.
Phylogenetic sequence analysis
Alignments were performed using ClustalW (Thompson et al., 1994). Maximum Likelihood trees were generated by 1,000 bootstrap repetitions under the Tamura-Nei model (Tamura and Nei, 1993) by MEGA7 (Kumar et al., 2016). Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value. The tree was drawn to scale, with branch lengths measured in the number of substitutions per site. Codon positions included were 1st+2nd+3rd+Noncoding. All positions containing gaps and missing data were eliminated. Further information is provided in the figure legends.
Results
Implementation of the high-throughput microfluidic real-time PCR system for tick-borne pathogens survey
The high-throughput microfluidic real-time PCR system developed for the screening of known and potential TBPs included 61 sets of primers and probes. Among them, 49 designs were developed for the detection of bacterial (n=32) and protozoan (n=17) species, bacterial (n=5) and protozoan genera/phylum (n=3) (Table 1). Three sets of primers and probes were developed for the molecular identification of the three tick species found in the Caribbean, including A. variegatum, R. microplus and R. sanguineus sensu lato(Table 1). Finally, we developed a design targeting a conserved region of the 16S rRNA genes within ticks, called Tick spp., used as a control of DNA/RNA extraction (Table 1).
The detection ability of each designs and the effect of pre-amplification on detection signals were first checked by Taqman real-time PCRs on a LightCycler 480 apparatus using dilution range of positives controls (Table 1). Three kind of positive controls were used, including bacterial or protozoan cultures when available, DNA from infected ticks or blood samples, and plasmidic constructions as a last resort (Table 1). Except the design targeting Borrelia burgdorferi sensu stricto which never succeeded in detecting the positive controls even after a preamplification step, the remaining 60 designs targeting TBPs and ticks species were able to detect their target with Ct values between 6 to 38 (data not showed). Pre-amplification improved the quality of the detection and was therefore validated as part of the screening protocol (see Additional file 2).
Then, the specificity of the designs was evaluated using the Biomark system and a total of 62 positive controls (Table 1, Fig 1).

Each square corresponds to a single real-time PCR reaction, where rows indicate the pathogen in the sample and columns represent the target of the primers/probe set. Ct values for each reaction are represented by a color gradient; the color scale is shown on the right y-axis. The darkest shades of blue and black squares are considered as negative reactions with Ct > 30.
42 primers/probe sets were able to detect and amplify specifically their target using a Ct cut-off value of 30 and were then directly validated (Fig 1). The design Tick spp., used as tick acid nucleic extraction control, was able to detect A. variegatum and R. sanguineus s.l. samples, as well as the DNA of R. sanguineus s.l. tick present in the Rickettsia conorii positive control as expected (Fig 1). However the DNA of ticks from the R. microplus control sample and others positive controls including tick DNA (such as the controls Borrelia lonestari, Anaplasma phagocytophilum, etc.) were not detected (Fig 1). The detection ability of this designed has been corrected by the addition of the Tick spp. primers during the preamplification step, which were initially excluded since the objective was to enrich pathogen DNA content comparatively to tick DNA (data not shown).
The remaining designs gave false-positive results in outgroup controls in addition to their target detection. Interestingly, two kind of unsuspected signals were observed, some related to cross-reactions with closely related species and some related to potential co-infections in controls corresponding to field samples (Fig 1). Eight designs - Rickettsia massiliae, Rickettsia conorii, Bartonella henselae, Bartonella bacilliformis, Babesia canis vogeli, Babesia microti, Theileria parva, Hepatozoon americanum - gave positive results in outgroup controls, revealing cross-reactions with one to two closely related species (Fig 1). Babesia ovis and Rickettsia rickettsii designs gave multiple cross-reactions with closely related species or distant outgroups and thus were considered as non-specific and removed from the rest of the study (Fig 1). Finally, the seven remaining designs - Rickettsia spp., Rickettsia felis, Rickettsia africae, Apicomplexa, Babesia bigemina, Hepatozoon spp., Hepatozoon canis - gave positive results in outgroup controls linked to potential co-infection in controls corresponding to DNA from ticks collected in the field or DNA from infected blood (Fig 1). As co-infections may occur in natural tick or blood samples, these unexpected detections in biological samples could be due to natural (co-)occurrence of microorganisms rather than cross-reactions (see details of co-infection analysis in Additional file 3).
To conclude, with the exception of the sets of primers and probes targeting Borrelia burgdorferi sensu stricto, Babesia ovis and Rickettsia rickettsii that were finally removed from the study, the 58 remaining designs were validated for the high-throughput screening of pathogens in Caribbean ticks, taking into account the notified cross-reactions.
Large scale TBPs detection survey in ticks from Guadeloupe and Martinique
A total of 578 adults ticks were collected from cattle in Guadeloupe and Martinique. A total of 523 were tested using the BioMarkTM system developed in this study. The number of positive ticks and the corresponding infection rates for each pathogen detected were calculated for 132 A. variegatum, 165 and 281 R. microplus from Guadeloupe and Martinique, respectively (Fig 2). As some of the R. microplus samples corresponded to pool of two to four adult specimens, we reported the minimum and maximum infection rates (see Material and methods).
Conventional PCR/Nested PCR followed by amplicon sequencing were performed on several tick samples to confirm some of the results of the newly designed BioMarkTM system (Table 2). Percentage of identity of the sequences obtained with reference sequences available in NCBI are presented in Table 3.

Number of positive ticks Amblyomma variegatum (out of the 132), Rhipicephalus microplus from Guadeloupe (out of 165) and Martinique (out of 281). Dots on the map indicate the tick collection sites in Guadeloupe and Martinique. Yellow: collection site of Rhipicephalus microplus ticks; Red: collection site of Amblyomma variegatum; Blue: collection site of both tick species, sometimes on the same animal. IR: Infection rate. As some samples of Rhipicephalus (B.) microplus were pooled, we present minimum and maximum infection rates of infected ticks.
Detection of known TBPs in Caribbean ticks
Seven TBPs known to circulate in the Caribbean were detected in ticks from Guadeloupe and Martinique, including R. africae, E. ruminantium, An. marginale, B. bigemina, B. ovis, T. mutans and T. velifera (Fig 2).
Rickettsia spp. were only detected in ticks collected in Guadeloupe (Fig 2). R. africae was identified in 95.6% of the A. variegatum samples (Fig 2). In contrast, Rickettsia spp. detected in 15.7-23.5% of the R. microplus samples from Guadeloupe were not directly identified as R. africae with the BiomarkTM system (Fig 2). Thus, fourteen samples of A. variegatum (6/14) and R. microplus (8/14) positive for Rickettsia spp. were tested by nested PCR with primers targeting the ompB gene, followed by sequencing (Table 2). All the sequences recovered were identical, and displayed 100% of identity with R. africae (Table 3). The consensus sequence was deposited under the name Rickettsia africae Tick208 (accession number MK049851). E. ruminantium was identified in 5.1% of the A. variegatum ticks from Guadeloupe (Fig 2). We confirmed the presence of E. ruminantium nucleic acids by testing one sample of A. variegatum by conventional PCR targeting the 16S rRNA genes followed by amplicon sequencing (Table 2). The sequence obtained displayed 100% of sequence identity with E. ruminantium and were deposited under the name Ehrlichia ruminantium Tick116 (accession number MK049848) (Table 3, Fig 3).

Phylogenetic analysis of 16S rRNA sequences of Ehrlichia spp. by maximum likelihood method based on the Tamura-Nei model. In the phylogenetic tree, GenBank, species designation and strain name are given. The sequences investigated in the present study are marked with a black circle (Ehrlichia sp. Tick428, accession number MK049849) and a black diamond (Ehrlichia ruminantium Tick116, accession number MK049848). The tree with the highest log likelihood (-413.76) is shown. The percentage of trees in which the associated taxa clustered together is shown above the branches (bootstrap values). The analysis involved 25 nucleotide sequences. There were a total of 206 positions in the final dataset.
An. marginale was identified in R. microplus ticks from the both islands, with infection rates reaching 3.6-4.8% and 39.5-41.3% of specimens from Guadeloupe and Martinique, respectively (Fig 2). We confirmed the detection of An. marginale by testing two samples of R. microplus by conventional PCR targeting the 16S rRNA genes followed by amplicon sequencing (Table 2). We obtained two identical sequences, deposited under the name Anaplasma sp. Tick283 (accession number MK049844) that displayed 100% of sequence identity with Anaplasma spp. including An. marginale (Table 3, Fig 4).

Phylogenetic analysis of 16S rRNA sequences of Anaplasma spp. by maximum likelihood method based on the Tamura-Nei model. In the phylogenetic tree, GenBank, species designation and strain name are given. The sequences investigated in the present study are marked with a black triangle (Anaplasma sp. Tick283, accession number MK049844) and a black square (Anaplasma sp. Tick314, accession number MK049845). The tree with the highest log likelihood (-473.44) is shown. The percentage of trees in which the associated taxa clustered together is shown above the branches (bootstrap values). The analysis involved 12 nucleotide sequences. There were a total of 243 positions in the final dataset.
B. bigemina was detected in 0.6-1.2% and 12.5-12.8% of the R. microplus ticks from Guadeloupe and Martinique respectively (Fig 2). B. bovis was only detected in tick from Martinique, with an infection rate of 0.7% in R. microplus samples (Fig 2). As conventional and nested PCR did not succeed in detecting those parasites, we directly sequenced amplicons obtained with the B. bigemina and B. bovis designs developed here and corresponding sequences were identified (accession numbers MK071738 and MK071739 respectively) (Table 3).
T. velifera and T. mutans were detected in both tick species and in the two islands. T. velifera was identified in 42.3% of the A. variegatum samples and in 24.1-31.9% and 25.6-26% of the R. microplus samples from Guadeloupe and Martinique, respectively (Fig 2). As well, T. mutans was detected in 1.5% of the A. variegatum samples, and in 1.8-2.4% and 1.4% of the R. microplus samples from Guadeloupe and Martinique, respectively (Fig 2). Unfortunately, neither conventional PCR nor Biomark amplicon sequencing succeeded in confirming the Biomark results.
Detection of unexpected microorganisms in Caribbean ticks
Unexpected signals were obtained during the screening of microorganisms in ticks from Guadeloupe and Martinique, including the first detection of untargeted species belonging to the genera Anaplasma, Ehrlichia, Borrelia and Leishmania (Fig 2).
Ehrlichia spp. were detected in R. microplus ticks from both islands, with infections rates reaching 4.2-6.6% and 47.7-49.1% in Guadeloupe and Martinique, respectively (Fig 2). We tested two of the Ehrlichia spp. positive R. microplus samples by conventional PCR targeting the 16S rRNA genes (Table 2). We obtained two identical sequences, deposited under the name Ehrlichia sp. Tick428 (accession number MK049849) (Table 3). Phylogenetic and genetic distance analyses were performed using fragment of the 16S rRNA genes of several Ehrlichia species (Fig 3). The Ehrlichia sp. Tick428 sequence was found within a cluster including various uncharacterized Ehrlichia species detected in ticks from Asia and Africa.
In addition, in around 50% and 18% of the R. microplus specimens positive for Anaplasma spp., none of the Anaplasma species targeted by the BiomarkTM system gave signals, suggesting the presence of an unexpected or a new Anaplasma spp. (Fig 2). We tested two of the Anaplasma spp. positive R. microplus samples by conventional PCR targeting the 16S rRNA genes (Table 2). We obtained two identical sequences, deposited under the name Anaplasma sp. Tick314 (accession number MK049845) (Table 3). This sequence displayed 100% of sequence identity with Candidatus Anaplasma boleense. Phylogenetic and genetic distance analyses were performed using fragment of the 16S rRNA genes of several Anaplasma species (Fig 4). Anaplasma sp. Tick314 sequence was found in a cluster including Candidatus Anaplasma boleense, Anaplasma platys and Anaplasma phagocytophilum.
Borrelia spp. was detected in both tick species from both islands (Fig 2). Infection rates reached 5.1% in A. variegatum, and 0.6% and 4.3% in R. microplus from Guadeloupe and Martinique, respectively (Fig 2). None of the specific Borrelia species targeted causing Lyme disease (Borrelia burgdorferi sensu lato) or Borrelia Relapsing Fever group, gave any positive results suggesting the occurrence of a new or unexpected Borrelia spp. in our samples (Fig 2). We tested 30 of the Borrelia spp. positive ticks by nested PCR targeting the flaB genes (Table 2). Interestingly, we obtained two sequences according to the tick species analyzed. Borrelia sp. Tick7 (accession number MK049846) sequence was recovered from one A. variegatum samples from Guadeloupe and Borrelia sp. Tick457 sequence (accession number MK049847) was recovered from four R. microplus samples from Martinique (Table 3). Phylogenetic and genetic distance analyses were performed using fragment of the flaB gene of several Borrelia species (Fig 5). Surprisingly, the Borrelia sp. Tick7 sequence recovered from the A. variegatum sample, and found closely related to Bo. anserina, displayed an intermediate position, sharing homology with both the relapsing fever and Lyme disease groups (Fig 5). Finally, the Borrelia sp. Tick457 sequence recovered from the R. microplus samples confirmed the previous observations, forming a cluster with various Relapsing Fever Borrelia species encountered in hard ticks, including Bo. lonestari and Bo. theileri (Fig 5).

Phylogenetic analysis of flaB sequences of Borrelia spp. by maximum likelihood method based on the Tamura-Nei model. In the phylogenetic tree, GenBank, species designation and strain name are given. The sequences investigated in the present study are marked with a black circle (Borrelia sp. Tick457, accession number MK049847) and a black triangle (Borrelia sp. Tick7, accession number MK049846). The Lyme disease and relapsing fever clades of Borrelia are marked. The tree with the highest log likelihood (-963.24) is shown. The percentage of trees in which the associated taxa clustered together is shown above the branches (bootstrap values). The analysis involved 16 nucleotide sequences. There were a total of 245 positions in the final dataset.
Finally, 0.7% of R. microplus ticks from Martinique were positive for Leishmania spp. (Fig 2). We tested two of the Leishmania spp. positive ticks by nested PCR targeting the small sub-unit rRNA gene (Table 2). We obtained one sequence from one sample, deposited under the name Leishmania martiniquensis Tick389 (accession number MK049850) (Table 3). This sequence displayed 100% of identity with both Leishmania martiniquensis and Leishmania siamensis sequences (Table 3).
Co-infections in ticks of Guadeloupe and Martinique
In Guadeloupe, almost all A. variegatum (99.3%) were infected by at least one pathogen whereas only 39.8 – 56.6% of R. microplus were infected. In contrast, 79.4 – 81.5% of R. microplus from Martinique were infected by at least one pathogen. High and similar percentages of the two tick species were infected by either one or two pathogens. The percentages drastically felt down from co-infection by three pathogens with less than 10% of the ticks infected. Respectively one and nine A. variegatum and R. microplus, from Guadeloupe and Martinique were co-infected by four pathogens and one R. microplus from Martinique was found infected by five pathogens. In the rest of the analysis on co-infections, only pathogens present in a sufficient number of ticks (more than 25) were further considered for possible infectious interactions in the ticks. These pathogens are R. africae and T. velifera in A. variegatum of Guadeloupe and T. velifera, An. marginale and Ehrlichia spp. in R. microplus of Martinique. In A. variegatum, massively infected by R. africae, there are up to 40.1% of the ticks infected by T. velifera, suggesting that this pathogen is not excluded by R. africae. There is no difference in the infection rates of T. velifera in R. microplus of Guadeloupe and Martinique (24-26%). However, the infection rates of An. marginale and Ehrlichia spp. in R. microplus of Guadeloupe were much lower than the ones observed in Martinique: 4% for both pathogens in Guadeloupe versus 40% and 48% for A. marginale and Ehrlichia spp., respectively in Martinique. The detailed analyses of the presence of T. velifera, An. marginale and Ehrlichia spp. in the Martinique ticks did not show any significant biases in the infection rates between single versus dual-infection, suggesting that none of these pathogens can interfere with the infection by the two others. Details of Co-infections in ticks of Guadeloupe and Martinique are available in Additional file 4.
Discussion
In this study, a high-throughput microfluidic real-time PCR system based on the use of multiple primers/probes was developed for large scale surveys of bacteria and protozoans potentially transmitted by ticks from the Caribbean area.
The association of genus and species primer/probe designs targeting TBPs improved the screening capacity of the technology, allowing not only the identification of infectious agents known to circulate in the studied area, but also the detection of unsuspected TBPs or new microorganisms belonging to the main bacterial and protozoan genera/phylum involved in TBD worldwide. Nevertheless, as some endosymbiotic microorganisms could belong to known TBPs genera, such as Rickettsia and Coxiella genus, confirmatory testings are required before suggesting the presence of a pathogenic microorganism (Ahantarig et al., 2013; Duron et al., 2015; Bonnet et al., 2017).
While analyzing the specificity of the microfluidic real-time PCR system, cross-reactions have been observed for some designs targeting closely related species that must be taken into account when interpreting results. Due to high design constraints and a lack of available sequences in public databases, the improvement of such cross-reacting oligonucleotides remain challenging. Here, the concomitant use of the bacterial and protozoan genera can assist non-specific signals identification.
In addition to microorganism’s detection, we developed sets of primers and probes allowing the molecular identification of the three main tick species involved in TBDs in the Caribbean, including A. variegatum, R. microplus and R. sanguineus s.l.. As Morphological identification of ticks collected on the field remains challenging, molecular identification can assist the correct identification of the tick species analyzed.
We used the newly developed high-throughput microfluidic real-time PCR system to perform an exploratory epidemiological study of TBPs and microorganisms potentially circulating in Caribbean ticks. The analysis gave an overview of the diversity of microorganisms belonging to the main bacteria and protozoan genera potentially transmitted by ticks. It allowed both the detection of known TBPs of public and animal health importance in the area which require surveillance, and unexpected microorganisms within Caribbean ticks.
The four main pathogens responsible for ruminant diseases in the Caribbean – currently classified as notifiable diseases by the World Organisation of Animal Health (OIE) –have been detected by the microfluidic real-time PCR system, including E. ruminantium in A. variegatum specimens and An. marginale, B. bigemina and B. bovis in R. microplus.
Interestingly, E. ruminantium infection rate in A. variegatum reported in our study was much lower compared to previous studies conducted between 2003 and 2005 in Guadeloupe (5.1% versus 36.7%) (Vachiéry et al., 2008). Although study designs were distinct (different sampling strategy, study periods, or detection methods, etc.) and can explain this difference, it is worth investigating further whether the tick infection rate for E. ruminantium decreased in Guadeloupe and possibly assess the epidemiological impact in term of incidence and prevalence of heartwater in the ruminant population. Those results are all the more surprising that systematic TBT surveillance and control programs have stopped after the end of the eradication programs POSEIDOM in 2006.
In this study, we document for the first time infection rates for B. bigemina, B. bovis and An. marginale in the vector tick R. microplus in the French Antilles. Indeed, records of such pathogens are mostly based on seroprevalence studies in cattle (Camus and Montenegro-James, 1994; Camus and Barre, 1995; Alonso et al., 1992).
R. microplus ticks are both vector and reservoir of B. bigemina and B. bovis, transmitting transovarially and trans-stadially the parasites (Howell, et al., 2007; Chauvin et al., 2009). As R. microplus ticks and bovine are both reservoir of the infection, the infection rates reported here seemed quite low (Howell, et al., 2007; Chauvin et al., 2009). The life cycle of Babesia spp. require complex interactions with its two host, the tick vector and the vertebrate host. The efficiency of tick acquisition, transovarial and trans-stadial transmissions of B. bovis and B. bigemina by R. microplus, involved in long-term persistence of Babesia spp. in nature, are still poorly understood and deserved further investigations (Chauvin et al 2009, Howell et al., 2009).
Interestingly, An. marginale was detected in R. microplus from the two islands but the infection rate reported in ticks from Guadeloupe seemed lower compared to Martinique. The same tendency has been reported during previous seroprevalence studies (Camus and Montenegro-James, 1994; Camus and Barre, 1995; Alonso et al., 1992). Anaplasmosis can be transmitted by other vectors than ticks and some cattle breeds are known to be more susceptible than others to Anaplasma infection (Camus and Montenegro, 1994). The difference of Anaplasma prevalence in ticks between the two islands could be explained by the difference in cattle populations. Indeed, there are mainly local Creole or mixed European-Creole breeds in Guadeloupe – known to be more resistant to anaplasmosis than Brahman and European breeds – which are mainly bred in Martinique (Camus and Montenegro, 1994). In addition, other factors, like differences in the population dynamics of alternate vectors such as flies could also contribute to this difference.
Among the other known TBPs detected, we also found pathogens with low impact on health in the Caribbean – almost considered as endosymbionts – such as R. africae, T. velifera and T. mutans in their vector A. variegatum and surprisingly in R. microplus ticks.
With almost all A. variegatum found infected, the R. africae infection rate is the higher ever reported in the Caribbean (Parola et al., 1999; 2003; Robinson et al., 2009; Kelly et al., 2010). As A. variegatum is both the vector and the reservoir of the pathogen, with transovarial and trans-stadial transmission rates reaching 100%, this high level of R. africae infection rate is not surprising per se (Socolovshi et al., 2009; Kelly et al., 2010). Interestingly, the high R. africae infection rates in vector ticks, associated to a very low number of African tick bite Fever cases in the Caribbean, highlights the difficulty in some cases to clearly distinguish between endosymbiosis and pathogenicity (Parola et al., 1999; Raoult et al., 2001). The biological relationship between R. africae and A. variegatum as well as the strains variety and virulence of R. africae in the Caribbean should be investigated in order to better assess the risks and guide prevention measures, especially for the travelers (Ahantarig et al., 2013; Maina et al., 2014; Duron et al., 2017). The absence of direct identification of R. africae in R. microplus ticks was probably due to a lower sensitivity of the specific target design compared to the genus target design. Indeed, Rickettsia spp. positive R. microplus samples displayed quite high Ct values, suggesting a low infection level that might be below the detection limit for R. africae. The unusual presence of R. africae in R. microplus tick could be explained by the co-occurrence of the two tick species, R. microplus and A. variegatum, on cattle. As the ticks here were collected partially engorged, the presence of R. africae in R. microplus may be due to bacteria circulating in cattle blood which would have been picked up by engorging ticks, or to a cross-contamination during R. microplus ticks co-feeding next to infected A. variegatum (Yssouf et al., 2014; Matsimbe et al., 2017).
The study provides the first update of the detection of T. mutans and T. velifera in Caribbean ticks. Indeed, references to these parasites in the Caribbean are quite old, and no prevalence studies neither in ticks nor in cattle, have been conducted since then (Morel et al., 1967; Uilenberg et al., 1983; Rodriguez et al., 1989). The low pathogenicity of these piroplasms may explain the lack of diagnostic and the scarce information available on their distribution and prevalence in the Caribbean. However, these parasites species may play an important role in theileriosis management and protection as chronically infected cattle can develop immunity and heterologous protection against other pathogenic Theileria species, such as Theileria parva (Woolhouse et al., 2015). Unfortunately, we did not succeed in confirming these results by conventional or nested PCR, which can suggest either a level of infection under the detection threshold, or simply false signals.
Finally, the high-throughput microfluidic real-time PCR system allowed the detection of unexpected and/or potentially new microorganisms, leading to the recovery of nucleotide sequences of Anaplasma spp., Ehrlichia spp., Borrelia spp. and Leishmania spp. in ticks collected in Guadeloupe and Martinique.
The Ehrlichia sp. Tick428 sequence detected here formed a cluster with other uncharacterized Ehrlichia species detected in ticks from Asia and Africa (Parola et al., 2001, 2003; Inokuma et al., 2004; Loftis et al., 2006; Kang et al., 2014; Lu et al., 2017). However, regarding the highly conserved nature of the 16S rRNA genes, we could not define more accurately the phylogenetical relationships within the Ehrlichia species group. The Anaplasma sp. Tick314 sequence was identified as Candidatus Anaplasma boleense, a bacteria described in ticks and mosquitoes in China (Kang et al., 2014; Guo et al., 2016). No further information are available regarding the epidemiology of Candidatus Anaplasma boleense. These observations highlight the need to set up characterization studies. Indeed, high-throughput detection technologies can highlight the presence of DNA from potentially new microorganisms, but it will be still necessary to isolate and characterize them in order first to confirm their existence and second to determine if their presence in ticks represents a risk for public or animal health.
We provided here the first report of Borrelia spp. in ticks from Guadeloupe and Martinique. Two different sequences have been recovered, according to the tick species analyzed. In A. variagetum, a sequence named Borrelia sp. Tick7 have been detected, closely related to Bo. anserina, the agent of avian spirochetosis. Both of them seemed to define an intermediate position between the relapsing fever and the Lyme disease group. In contrast, the Borrelia sp. Tick457 sequence found R. microplus, clustered with uncharacterized Borrelia spp., Borrelia sp. strain Mo063b and Borrelia sp. BR, detected in R. microplus from Madagascar and Brazil and Relapsing Fever Borrelia species encountered in hard ticks, including Borrelia lonestari and Bo. theileri (Yparraguirre et al., 2007; Hagen et al., 2017). Interestingly, the same observations were made recently about Borrelia spp. found in A. variegatum and R. microplus ticks from Ethiopia and Côte d’Ivoire (Kumsa et al., 2015; Ehounoud et al., 2016). As A. variegatum and R. microplus were imported in the Caribbean from Africa during the time of the Atlantic triangular trade, we may have detected bacteria probably characterized by an old introduction through infected ticks and subsequent local evolution within their vector over a long period (Barré and Uilenberg, 2012; Estrada-Peña et al., 2018). Borrelia spp. and borreliosis cases reports in the Caribbean are scarce and still in debate. In Cuba, one study suggested the presence of antibodies to Borrelia burgdorferi sensu stricto in human serums associated with clinical cases of Lyme disease-like syndrome (Rodríguez et al., 2004, 2012). However, the real specificity of these serum antibodies was questioned (Dessau, 2012). In the US Virgin Islands, seropositivity to Borrelia hermsii and closely related species have been reported in association with a human case of relapsing fever (Flanigan et al., 1991). Finally, Erythema Migrans– like skin lesions and illness have been reported in four Caribbean nationals (Sharma et al., 2010). Regarding the importance of Borrelia spp. in human and animal health, the characterization of these potential new Borrelia species which seemed associated to tropical tick species require further investigations.
Lastly, Leishmania spp. were detected in R. microplus specimens from Martinique, and a sequence was identified as Leishmania martiniquensis Tick389 (accession number MK049850). Studies on Leishmania nomenclature highlighted the fact that isolates of “L. siamensis” have never been officially characterized and therefore, this name should not be used (Pothirat et al., 2014; Espinosa et al., 2018; Leelayoova et al., 2017; Steverding, 2017). Thus, as all the sequence reported as “L. siamensis” – except one – in databases should be considered as a synonym of L. martiniquensis, we assumed here the occurrence of L. martiniquensis. Parasites of the genus Leishmania are usually transmitted by female phlebotomine sand flies, (Diptera: Pshychodidae: Phlebotominae), and involve generally a wide variety of animal species, mostly dogs and canids in the epidemiological cycle. They are responsible for Leishmaniasis, a zoonosis widespread in tropical and sub-tropical areas (Steverding, 2017). L. martiniquensis belongs to the L. enriettii complex and has been described in Martinique and in Thailand, where it was responsible for both cutaneous and visceral leishmaniosis (Desbois et al., 2014; Pothirat et al., 2014; Liautaud et al., 2015; Steverding, 2017). L. martiniquensis is suspected to be endemic in Martinique (Desbois et al., 2014). Although phlebotomines and rodents are present in Martinique, neither a vector nor a reservoir of this parasite have yet been described (Desbois et al., 2014). Our study represents the first report of L. martiniquensis in R. microplus ticks from the French Antilles. Althought Leishmania spp. have been reported in ticks, such as L. infantum in R. sanguineus, or L. guyanensis in R. microplus ticks in Peru, the role of ticks in Leishmania transmission is still a debate and no evidence of vector capacity have been reported yet (Dantas-Torres, 2011; Rakhshanpour et al., 2017; Rojas-Jaimes et al., 2017). In addition, the finding of Leishmania spp. in a tick species that feed mainly on cattle also raises question on the potential role of cattle in the epidemiology of leishmaniasis (Alam et al., 2018, Paixão-Marques et al., 2019). The participation of ticks in Leishmania epidemiology deserve further investigation, especially since R. microplus ticks could parasitizing human (Lamattina and Nava, 2016).
Surprisingly, co-infections with two or more TBPs were found in more than 50% of the infected ticks, both for A. variegatum and R. microplus and in the two islands. In addition, we could not identify any association or exclusion of the infection by one pathogen to another one. These observations illustrate the efficiency of ticks as reservoir of multiple pathogens with apparently no significant effects on their life traits.
To conclude, although screening tools are useful for pathogen discovery in ticks, the epidemiological meaning of such results warrants further analyzes. Detecting microorganisms DNA in ticks, especially in partially engorged ticks, does not necessarily mean that the tick is involved in the microorganism’s life cycle, however they provide useful and complementary information to vector competence studies (Estrada-Peña et al., 2013). Nevertheless, the detection of potentially new microorganisms in ticks from the French Antilles open new research perspectives on TBPs epidemiology in the Caribbean for the future. A region-wide epidemiological survey on TBPs in ticks collected in different countries and territories of the Caribbean area to strengthen our results and organized in collaboration with the Caribbean Animal Health Network, CaribVET and the ticks and tick-borne diseases working group may be an interesting way to complete and strengthen some of the findings of this paper.
Conclusion
Our study demonstrated the high ability of microfluidic real time PCR technology to give a rapid overview of the diversity of TBPs of veterinary and medical importance present in ticks from the Caribbean. This innovative high-throughput tool is promising and could improve significantly TBPs surveillance and study, allowing a rapid screening of multiple especially in regions where few epidemiological data are available and TBD are numerous.
Declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Availability of data and material
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Competing interests
The authors declare that they have no competing interests
Funding
This publication was funded by grants from The French Agency for Food, Environmental and Occupational Health & Safety CoVetLab grant (ANSES), the French Agricultural Research and International Cooperation Organization (CIRAD) and the French National Institute for Agricultural Research (INRA, PATHO-ID Metaprogram). This study was partly supported by the project MALIN “Surveillance, diagnosis, control and impact of infectious diseases of humans, animals and plants in tropical islands” supported by the European Union in the framework of the European Regional Development Fund (ERDF) and the Regional Council of Guadeloupe. The study used the ticks collected in Guadeloupe and Martinique within the framework of previous projects: PathoID “Rodent and tick Pathobiome” funded by INRA and Resist project “Assessment of Ticks Resistance to Acaricides in the Caribbean - Development of strategies to improve surveillance and control of tick-borne diseases in Ruminants” funded by the Fond de Cooperation Régionale (FCR) of Guadeloupe and Martinique. This work was performed under the framework of the EurNegVec COST Action TD1303.
Authors’ contributions
MG wrote the paper. MVT, EA, JP and SM reviewed the manuscript. EA, JP and SM designed the experiment and supervised the manuscript. MG performed the experiment. ED, VP, RA, CG, MVT contributed to sampling and sample preparation. SD contributed to technical support.
Additional Files
Additional file 1.pdf: GPS coordinates of ticks collection sites
A total of 578 adult ticks collected on bovine from Guadeloupe and Martinique were used for the screening of tick-borne pathogens with the newly implemented BioMark™ Real-Time PCR system. 132 Amblyomma variegatum and 165 Rhipicephalus microplus adult specimens were collected on cattle in Guadeloupe in this study. Ticks were sampled between February 2014 and January 2015, on 40 bovines originating from 22 different herds including : four herds from Les Abymes (N 16° 16’; O 61° 31’), three from Sainte-Anne (N 16° 14’; O 61° 23’), Petit Bourg (N 16° 10’; O 61° 36’), Capesterre (N 16° 2’; O 61° 34’), Le Gosier (N 16° 13’; O 61° 28’), Morne-À-l’Eau (N 16° 19’; O 61° 28’), two from Pointe-Noire (N 16° 13’; O 61° 45’), and one from Bouillante (N 16° 7’; O 61° 46’).
In Martinique, only Rhipicephalus microplus specimens were collected on bovine. Indeed, ticks were provided in the frame of the ResisT project, funded by the Regional Cooperation Fund of Guadeloupe and Martinique which aimed to analyze Rhipicephalus microplus acaricides resistance in the Caribbean. Thus, 281 Rhipicephalus microplus adult specimens were collected on cattle in Martinique between February and March 2015, on 29 bovines originating from 29 herds: eight from Le Vauclin (N 14° 32’; O 60° 50’), four from Sainte-Anne (N 14° 26’; O 60° 50’), Le Robert (N 14° 40’; O 60° 56’), two from Le François (N 14° 36’; O 60° 53’), Saint-Joseph (N 14° 40’; O 61° 2’), one from Rivière-Pilote (N 14° 28’, O 60° 54’), Le Diamant (N 14° 29’; O 61° 1’), Le Lamentin (N 14° 37’; O 60° 59’), Le Lorrain (N 14° 49’; O 61° 2’), Le Morne-Vert (N 14° 42’; O 61° 8’), Rivière-Salée (N 14° 31’; O 60° 57’), Saint-Esprit (N 14° 33’; O 60° 55’), Sainte-Luce (N 14° 29’; O 60° 56’), and Sainte-Marie (N 14° 46’; O 60° 59’). All the ticks were collected on cattle, partially engorged, and then conserved at - 80°C. Ticks were morphologically identified at the species level (Walker et al., 2003).
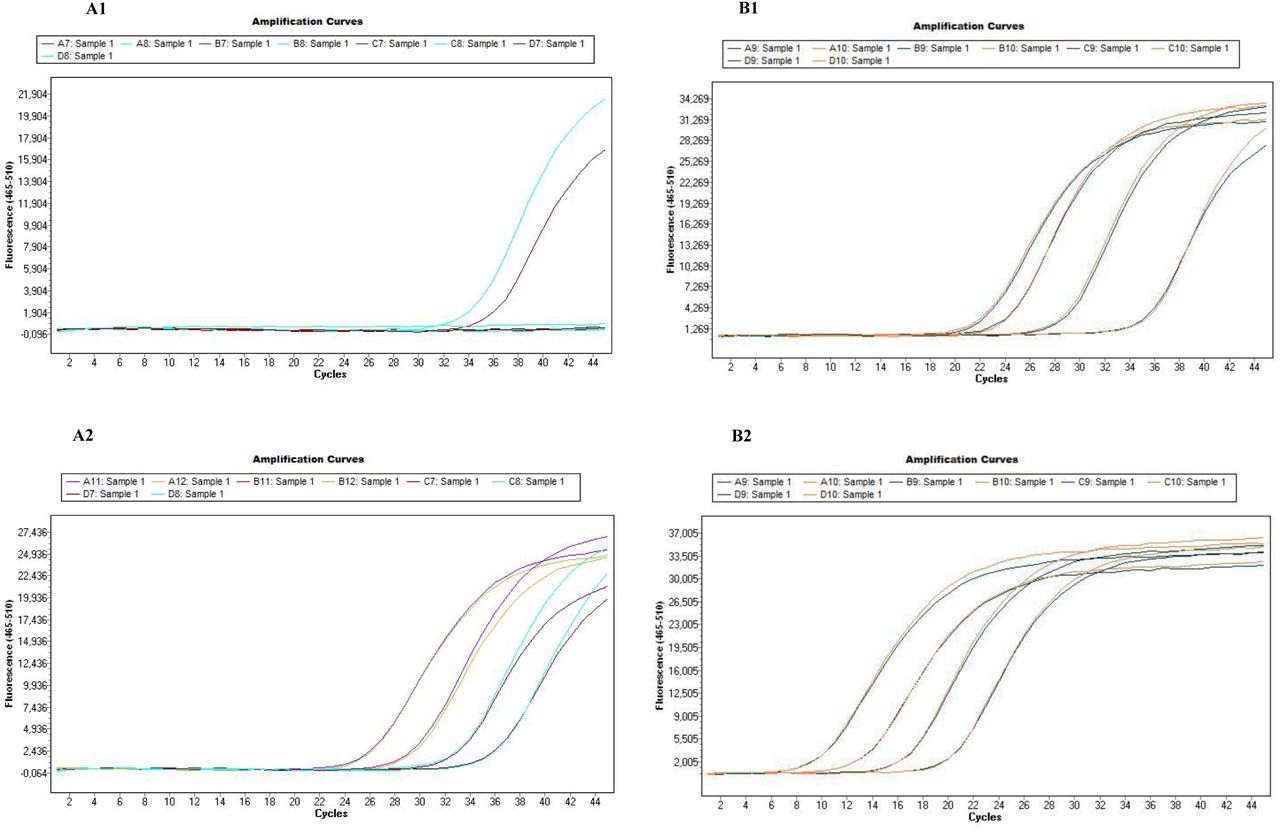
Additional file 2.pdf: Improvement of detection signals by pre-amplification
Test of primers/probe set sensitivity on a dilution range of positive controls by Taqman real-time PCRs using LightCycler 480, before and after pre-amplification. A1/A2: Test of sensitivity of the design Leishmania infantum using Leishmania infantum culture, before (A1) and after pre-amplification (A2); B1/B2: Test of sensitivity of the design Rickettsia spp. using Rickettsia conorii positive controls (extract from infected Rhipicephalus sanguineus tick), before (B1) and after pre-amplification (B2).
Additional file 3.pdf: Analysis of the specificity of the 61 designs constituting the Biomark system developed for Caribbean tick-borne pathogen screening
Among the 61 design used in this study, 1 were removed because of absence of signals, 43 were specific for their target; 8 displayed cross-reactions with one to two closely related species, 2 were removed because of multiple cross-reactions and 7 displayed unexpected signals due to the potential presence of co-infections in complex control samples corresponding to DNA extracted from wild ticks or blood samples.
We explored the hypothesis of potential co-infection in controls corresponding to DNA from tick collected in the field or DNA from infected blood and we confirmed some results using conventional PCR and amplicon sequencing (Table S2). Both Rickettsia spp. and Rickettsia felis designs gave positive results when testing the Borrelia lonestari, Panola Mountain Ehrlichia, Ehrlichia ewingii, Ehrlichia chaffensis controls. All those samples corresponded to DNA extracted from infected Amblyomma americanum collected on the field. This result supports the fact that a Rickettsia species may be present in those samples, being Rickettsia felis or a closely related Rickettsia spp. infecting Amblyomma americanum ticks. When tested the Panola Mountain Ehrlichia sample for Rickettsia spp. by conventional PCR targeting the gltA gene (Regnery et al., 1991), we obtained a 382 bp sequence (accession number MK049843) sharing 99% of sequence identity with Rickettsia spp. (MF511253.1) (Table S2). This result, in addition to natural co-infections documented in Amblyomma americanum ticks, suggest that the detection of Rickettsia spp. - within the four outgroup positive controls corresponding to Amblyomma americanum DNA – in this assay were not cross-reactions (Mixson et al., 2006; Trout Fryxell and DeBruyn, 2016). Moreover, the Amblyomma variegatum sample, corresponding to ticks collected from the field, was found positive for both Rickettsia spp. and Rickettsia africae. As this sample originated from Guadeloupe, where Rickettsia africae circulates, this result could be in agreement with a natural infection (Parola et al., 1999). As well, the Rhipicephalus sanguineus sample was positive with four designs targeting parasites – Apicomplexa, Hepatozoon spp., Hepatozoon canis and Hepatozoon americanum. As Rhipicephalus sanguineus ticks are involved in tick-borne parasites epidemiology including Hepatozoon spp., those results suggest strongly the occurrence of such parasite in the biological sample (Aktas and Özübek, 2017). Thus, those observations suggest that our designs were actually capable of detecting pathogens present in naturally-infected ticks rather than giving an unsuspected cross-reaction. The Neoehrlichia mikurensis sample, corresponding to DNA extracted from rodent blood, was also found positive for both Apicomplexa and Hepatozoon spp.. Amplicon obtained with the Hepatozoon spp. design from this sample was sequenced. The 169 bp sequence obtained, (accession number MK071735) displayed 99% of sequence identity with Hepatozoon spp. sequences (AB771515.1) (Table S2). As rodents could be infected by Hepatozoon parasites, this result could reflect also a natural infection (Hamšíková et al., 2016). The Anaplasma marginale sample, corresponding to a blood sample of experimentally infected cow, was found also positive for both Apicomplexa and Babesia bigemina. Amplicon obtained with the Apicomplexa design from this sample was sequenced. The 104 bp sequence obtained (accession number MK071737) displayed 99% of sequence identity with Apicomplexan sequences, including Babesia bigemina (MG604302.1) (Table S2). As Babesia bigemina and Anaplasma marginale are two cattle pathogens which often evolved in the same region, and are transmitted by the same vector tick, co-infections by those two pathogens have already been reported (Hailemariam et al., 2017). Thus, this cross-reaction could be again the reflection of parasites co-infections. Finally, Apicomplexa gave also positive results in Borrelia lonestari and the Panola Mountain Ehrlichia controls. As highlighted previously, those two controls corresponded to DNA extracted from A. americanum ticks. Amplicons obtained with the Apicomplexa design from those two samples were sequenced. The two sequences of 102 bp obtained were identical, (accession number MK071736) and displayed 98% of sequence identity with sequences of Theileria cervi (MH085203.1) (Table S2). As Theileria cervi is a common deer pathogen found in Amblyomma americanum, the occurrence of this parasite in those two control samples could explain the unexpected signals (Laird et al., 1988; Mans et al., 2015).
CR: Cross-reactions with closely related species samples; CI: Potential co-infections in control samples; *: designs removed from the screening analysis.
AN: Accession number, % I: Percentage of identity, % C: Percentage of coverage.
Additional file 4.pdf: Co-infections found in ticks of Guadeloupe and Martinique
Focus on Rickettsia africae and Theileria velifera in A. variegatum in Guadeloupe: 132 ticks were analyzed, the percentages of non-, single- and dual-infected ticks are presented in the table. There is no bias in the prevalence of T. velifera in ticks infected by R. africae.
Focus on Anaplasma marginale and Theileria velifera in R. B. microplus in Martinique: 281 ticks were analyzed, the percentages of non-, single- and dual-infected ticks are presented in the table. There are no biases in the prevalence of the two pathogens in single versus dual-infected ticks.
Focus on Ehrlichia spp. and Theileria velifera in R. B. microplus in Martinique: 281 ticks were analyzed, the percentages of non-, single- and dual-infected ticks are presented in the table. There are no biases in the prevalence of the two pathogens in single versus dual-infected ticks.
Focus on Ehrlichia spp. and Anaplasma marginale in R. B. microplus in Martinique: 281 ticks were analyzed, the percentages of non-, single- and dual-infected ticks are presented in the table. There are no biases in the prevalence of the two pathogens in single versus dual-infected ticks.
Raw data
Acknowledgments
We thank the “Tiques et Maladies à Tiques (TMT)” group of the CNRS “Réseau Ecologie des Interactions Durables” for stimulating discussion and support. We thank M. Hamon, S. Depraz, L. Felixine, R. Aprelon and J.F. Cosson and D. Pleydell for their contribution during tick collection, and research partners that gave us control samples.
Footnotes
Please note that this paper has already been published online as a preprint on bioRxiv: https://doi.org/10.1101/532457
Section results and discussion revised





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}