

Abstract
Microbes often live in multispecies communities where interactions among community members impact both the individual constituents and the surrounding environment. Here, we developed a system to visualize interspecies behaviors at initial encounters. By imaging two prevalent pathogens known to be coisolated from chronic illnesses, Pseudomonas aeruginosa and Staphylococcus aureus, we observed P. aeruginosa can modify surface motility in response to secreted factors from S. aureus. Upon sensing S. aureus, P. aeruginosa transitioned from collective to single-cell motility with an associated increase in speed and directedness – a behavior we refer to as ‘exploratory motility’. Through modulation of cAMP, explorer cells moved preferentially towards S. aureus and invaded S. aureus colonies through the action of the type IV pili. These studies reveal previously undescribed motility behaviors and lend insight into how P. aeruginosa senses and responds to other species. Identifying strategies to harness these interactions may open avenues for new antimicrobial strategies.
Introduction
It has been clear since the 1600’s that many microbial infections do not occur with a single species, but we have only recently begun to understand the profound impacts microbial species have on each other and patients (Nguyen and Oglesby-Sherrouse 2016)}. Studies of dental biofilms, intestinal communities, chronic wounds, and respiratory infections in patients with cystic fibrosis (CF) demonstrate that community interactions influence microbial survival and disease progression (Limoli and Hoffman 2019; Gabrilska and Rumbaugh 2015). For example, we and others find an association between coisolation of Pseudomonas aeruginosa and Staphylococcus aureus from the CF airway or chronic wounds and poor patient outcomes, including decreased lung function and shortened life-spans (Limoli et al. 2016; Maliniak, Stecenko, and McCarty 2016; Hubert et al. 2013). Laboratory studies also reveal interactions between these two pathogens can alter virulence factor production by one or both species, potentially influencing pathogenesis, persistence, and/or antibiotic susceptibility (Hotterbeekx et al. 2017). For example, in a model of coinfection on CF-derived bronchial epithelial cells, we observed P. aeruginosa and S. aureus form mixed microcolonies, which promotes the survival of S. aureus in the presence of vancomycin (Orazi and O’Toole 2017). One strategy to improve outcomes for these coinfected patients is to block harmful interspecies interactions before they begin.
Here, we designed a system to visualize early interactions between P. aeruginosa and S. aureus, and follow single-cell behaviors over time with live imaging. We show that P. aeruginosa can sense S. aureus secreted products from a distance and in turn, dramatically alter the motility behaviors of this Gram-negative bacterium. In response to S. aureus, individual P. aeruginosa cells transition from collective to single cell movement, allowing exploration of the surrounding environment and directional movement towards S. aureus. We find that such ‘exploratory motility’ is driven primarily by the P. aeruginosa type IV pili and modulation of the intracellular second messenger, cAMP. Thus, we provide a new means to study polymicrobial interactions at the single-cell level and reveal that P. aeruginosa can sense the presence of other microbial species and dramatically, yet specifically, modify its behavior in response to such interspecies signals.
Results
S. aureus promotes exploratory motility in P. aeruginosa
To understand early microbial interactions, we established an in vitro coculture system to monitor P. aeruginosa and S. aureus at first encounters. Bacteria were inoculated at low cell densities between a coverslip and an agarose pad in minimal medium, supplemented with glucose and tryptone, and imaged with phase contrast time-lapse microscopy every 15 minutes for 8 hours. Alone, P. aeruginosa cells replicate and expand outward as raft-like groups, as previously described for P. aeruginosa surface-based motility (Anyan et al. 2014; Burrows 2012) (Movie 1; Figure 1, still montage, top row). In comparison, coincubation with S. aureus resulted in dramatically altered behavior (Movie 2, Figure 1, still montage, bottom row). After two to three rounds of cell division, instead of remaining as a group, individual P. aeruginosa cells began to move as single cells, suggesting that P. aeruginosa responds to the presence of S. aureus by altering motility behaviors. P. aeruginosa significantly inhibited S. aureus growth, as previously reported (see Movie 3 for representative time-lapse movie of S. aureus alone).

Live imaging of polymicrobial interactions. P. aeruginosa (rod-shaped) was inoculated between a coverslip and an agarose pad, either in monoculture (top) or in coculture with equal numbers of S. aureus (cocci-shaped, bottom). Images were acquired every 15m for 9h. Representative snap-shots of Movies 1 (top) and 2 (bottom) are shown. Founding cells identified in the first frame are indicated with green rods (P. aeruginosa) or yellow circles (S. aureus). The location of the founding cell is indicated in each subsequent frame for positional reference.
To visualize P. aeruginosa motility in the presence of S. aureus in more detail, the inoculating cell density was reduced (2-3 cells of each species per field of view), and images were taken at 5s intervals for 8h. Movie 4 shows images taken during hours 4 – 6 of coculture, when P. aeruginosa initiates single-cell movement under these conditions (Figure 2A, still montage). We observed a number of surprising behaviors by P. aeruginosa in the presence of S. aureus. P. aeruginosa cells initially replicated and remained in a raft (t=4h:28m), as we and others have observed for P. aeruginosa in monoculture, but as the community approached S. aureus, individual cells: (1) exit the raft (t=4h:34m), (2) move with increased speed, and (3) move towards and surround S. aureus, “exploring” the surface of the colony until (Figure 2A-B), (4) P. aeruginosa cells enter the S. aureus colony (Figure 2C, t=6h,), and finally, (5) by 8h, P. aeruginosa completely dismantles the S. aureus community (Figure 2D, t=8h). P. aeruginosa was also observed to adopt a swift motion, beginning between 4.5 - 6h, moving in and out of the plane of focus during imaging (Movie 5 and Figure 2D, red arrows).

Single-cell live-imaging of P. aeruginosa with WT S. aureus (Movie 4). A. Montage of representative snap-shots are shown beginning at 4h:28m. Founding cells identified in the first frame are indicated with green rods (P. aeruginosa) or yellow circles (S. aureus). The location of the founding cell is indicated in each subsequent frame for positional reference. B. Snap-shot at 5h, zoomed in to visualize single cells. C. Snap-shot at 6h of P. aeruginosa (PA1/04/03 -mKO) and S. aureus (sarAP1-sGFP) illustrating P. aeruginosa invasion into S. aureus colonies. D. Snap-shot of coculture at 8h, showing disruption of S. aureus colonies (green arrows) and swift-moving P. aeruginosa cells out of the plane of focus (red arrows). Single P. aeruginosa cells and the leading edge of rafts were tracked in the presence of S. aureus and the speed (µm/s), acceleration (µm/s2), and mean squared displacement (µm2) for four independent movies are indicated, respectively, in E – G.
To quantitate the movement of P. aeruginosa single-cell motility in comparison to collective motility in rafts, the movement of individual cells and the leading edge of the rafts was tracked over time. In comparison to cells moving in rafts, individual cells moved with increased speed (µm/s), acceleration (µm/s2), and mean squared displacement (MSD, µm2) (Figure 2 E, F, and G, respectively). MSD represents a combined measure of both the speed and directional persistence of the cell, thus an increased MSD in single cells over a change in time suggests single cells exhibit directed motion, followed by a decreased MSD when P. aeruginosa cells reach the S. aureus colony.
P. aeruginosa type IV pili drive exploratory motility
How is P. aeruginosa motility generated in response to S. aureus? When grown in monoculture, P. aeruginosa performs cellular movement through the action of a single polar flagellum and the type IV pili (TFP) (Conrad et al. 2011; Gibiansky et al. 2010; Merritt et al. 2010). To determine how the observed P. aeruginosa “exploratory motility” is generated, P. aeruginosa strains deficient in the production of either TFP (ΔpilA; encoding the pilin monomers), flagella (ΔflgK; encoding the flagellar hook protein), or both (ΔpilA ΔflgK) were examined. Time-lapse images were taken as described for Figure 2, except they were acquired at 50 ms intervals for visualization of specific motility patterns. Representative movies and snap-shots (Figure 3A) were chosen at the time-point where P. aeruginosa was found to exhibit both slow and swift single-cell movements. The pilA mutant was unable move away from the group as single cells (Movie 6), as seen for the parental P. aeruginosa (Movie 5), suggesting the TFP are required for P. aeruginosa exploratory motility. However, the swift movement observed in the WT (Figure 3A, see boxed inset with red arrows), was maintained in ΔpilA, demonstrating that TFP are not required for this behavior.

Live, single cell imaging of P. aeruginosa with WT S. aureus. A. Representative snap-shots of coculture with WT S. aureus and P. aeruginosa (WT, ΔpilA, ΔflgK, and ΔpilA ΔflgK, left to right, Movies 5-8, respectively) are shown at t=4.5h. Boxed insets show swift-moving P. aeruginosa cells out of the plane of focus (red arrows). B. The area of S. aureus in monoculture or in the presence of the indicated P. aeruginosa strain was calculated at t=5h by dividing the total area in a single frame by the number of S. aureus colonies. A minimum of four movies were analyzed per condition. The mean and standard deviation are indicated. Statistical significance was determined by one-way ANOVA followed by Tukey’s Multiple Comparisons Test - a indicates a statistically significant difference (P ≤ 0.05) between S. aureus in monoculture and in the presence of either WT P. aeruginosa or ΔflgK; b indicates a statistically significant difference between ΔpilA and ΔpilA ΔflgK. C. Representative snap-shots of Movie 5, WT P. aeruginosa and S. aureus beginning at 4h with 50 ms intervals, showing back-and-forth motion. Red arrows indicate when a P. aeruginosa cell is moving in towards the S. aureus colony and green arrows indicate when a cell is moving away.
We next examined the ΔflgK mutant (Movie 7), which was seen to adopt the single-cell behaviors of the WT, except the swift movements were not observed, supporting the hypothesis that this movement is generated by the flagellum. These data suggest that while S. aureus modulates both TFP and flagella-mediated motility, the early events necessary for exploration (initiation of single cell movement and directional movement towards S. aureus) requires the TFP.
We also examined the response of a double ΔpilA ΔflgK mutant in the presence of S. aureus (Movie 8). Surprisingly, this mutant exhibited a phenotype distinct from either the WT, or the individual ΔpilA and ΔflgK mutants. P. aeruginosa cells deficient in both TFP and flagella were not only unable to produce the respective movements characteristic of these motility motors, but also were unable to remain within a raft-like group (Figure 3A). This behavior was not dependent upon the presence of S. aureus, as a similar pattern was observed for ΔpilA ΔflgK when visualized in the absence of S. aureus (Figure S1).

Live imaging of P. aeruginosa ΔflgK ΔpilA in monoculture. Representative snapshot at t=4h.
The influence of P. aeruginosa exploratory motility on S. aureus growth was also examined by measuring the area of the S. aureus colonies at 4.5 hours post coculture. S. aureus colonies were significantly smaller in the presence of WT P. aeruginosa or the ΔflgK mutant in comparison to S. aureus grown in monoculture (Figure 3B). However, in the presence the ΔpilA mutant, which was deficient in exploratory motility, colonies were significantly larger in comparison to WT and the ΔflgK mutant. Moreover, when grown in the presence of the double mutants, the area of the S. aureus colonies was not significantly different from S. aureus monoculture. These data suggest that ability of P. aeruginosa to perform exploratory motility influences P. aeruginosa inhibition of S. aureus growth.
While visualizing P. aeruginosa-S. aureus interactions at 50 ms intervals, we observed an additional P. aeruginosa behavior. When P. aeruginosa first encounters the S. aureus colony, the cell body orients perpendicular to the surface of the colony and moves back-and-forth (Movie 5 and Figure 3C) and appears to drive P. aeruginosa into the S. aureus colony. This behavior was only observed in the WT and ΔflgK mutant, suggesting that it is driven by the TFP.
Agr-regulated secreted S. aureus factors promote P. aeruginosa motility
Live-imaging suggests P. aeruginosa is capable of sensing S. aureus and initiating exploratory motility from a distance. Thus, we hypothesized that P. aeruginosa responds to S. aureus secreted factors. Since we observed that TFP were required for exploratory motility, we sought to develop a macroscopic assay where TFP motility could be monitored in the presence of S. aureus secreted factors only. Macroscopically, population-scale TFP-mediated twitching motility can be visualized by inoculating P. aeruginosa cells onto the bottom of a plate though 1.5% agar (sub-surface); cells move on the surface of the plate under the agar and can be stained with crystal violet for visualization (Turnbull and Whitchurch 2014). To test the hypothesis that P. aeruginosa responds to S. aureus secreted products, cell-free supernatant derived from overnight cultures of WT S. aureus, (normalized to OD600 = 5.0) was spread on the plate, prior to pouring molten agar. S. aureus supernatant significantly increased the motility diameter of P. aeruginosa in dose-dependent manner (Figure 4A), supporting the hypothesis that S. aureus secreted factors increase P. aeruginosa twitching motility.

A – D. Motility of WT P. aeruginosa was monitored by macroscopic sub-surface inoculation assays in the presence of medium alone or cell-free supernatant from the indicated S. aureus strains. A illustrates representative motility zones stained with crystal violet for visualization. The dilution factor of the supernatant is indicated in A and B. Undilute supernatant was used in C. In D and E, the motility of the indicated P. aeruginosa mutants was analyzed in the presence of medium alone or supernatant derived from WT S. aureus (0.5 dilution) under 1.5% agar or within 0.3% agar (E). The mean and standard deviation are indicated for at least three biological replicates. Statistical significance was determined by one-way ANOVA followed by Tukey’s Multiple Comparisons Test - a indicates a statistically significant difference (P ≤ 0.05) between the motility observed in the presence of S. aureus supernatant compared to medium alone, and b indicates a statistically significant difference (P ≤ 0.05) between the motility observed in the mutant strain (S. aureus mutants in B and C, P. aeruginosa mutants in D and E) compared to the parental.
Two primary regulators of secreted factors in S. aureus are the alternative stress sigma factor, sigma B (SigB) (Bæk et al. 2013) and the accessory gene regulator (Agr) quorum sensing system (Nair et al. 2011). S. aureus strains with transposon insertions in either sigB or agrB (Fey et al. 2013) were examined for their ability to induce P. aeruginosa twitching motility (Figure 4B). While the sigB::Tn strain phenocopied the WT, the agrB::Tn mutant lost all ability to induce P. aeruginosa twitching motility, suggesting that Agr regulates the production of the factors promoting motility in P. aeruginosa. The agr operon is organized around two divergent promoters, P2 and P3, and generates two primary transcripts, RNAII and RNAIII, respectively. RNAII encodes AgrB, AgrD, AgrC, and AgrA (Le and Otto 2015). To confirm a role for the Agr quorum sensing system, an unmarked deletion of agrBDCA was generated and complemented with WT agrBDCA. P. aeruginosa motility was examined in the presence of supernatant derived from these strains. As predicted, the ΔagrBDCA mutant was unable to enhance P. aeruginosa motility, and complementation restored activity to WT levels (Figure 4C).
To confirm that the TFP are necessary for increased P. aeruginosa motility in this assay, we examined the response of pilA deficient P. aeruginosa to S. aureus supernatant. In the absence of S. aureus supernatant, the pilA mutant was required for twitching motility, as previously reported (Darzins 1994). However, while motility was reduced in the presence of S. aureus supernatant, the ΔpilA mutant retained some ability to respond to S. aureus secreted factors (Figure 4D). Since we observed during live imaging that S. aureus can increase P. aeruginosa flagella-mediated motility, in addition to TFP-mediated motility, we hypothesized that the response retained in the ΔpilA mutant was due to increased flagellar-mediated motility. To test this hypothesis, we examined the response of ΔflgK and a double ΔpilA ΔflgK mutant to S. aureus supernatant. The single ΔflgK mutant phenotype trended lower, but was not significantly different from WT, while the motility of the ΔpilA ΔflgK mutant was reduced to levels not significantly different from ΔpilA or ΔpilA ΔflgK in the absence of supernatant (Figure 4D). These data support our observation from live-imaging that, while TFP were the primary contributors to increased motility, S. aureus secreted factors could increase both TFP- and flagellar-mediated motility.
To formally examine the response of P. aeruginosa flagella-mediated motility to S. aureus supernatant, traditional macroscopic low-percentage agar assays that measure the contribution of flagellar motility and chemotaxis were performed. Cell-free S. aureus supernatant was mixed into 0.3% agar prior to inoculating P. aeruginosa cells into the agar. The diameter of the P. aeruginosa motility zone showed a modest, but not significant increase in the presence of S. aureus supernatant, compared to medium alone (Figure 4E). To examine the role for flagella in response to S. aureus under swim assay conditions, the flgK mutant was tested. As expected, flagellar-mediated motility, both with and without S. aureus supernatant, was significantly reduced. To determine if TFP contribute under these assay conditions, the ΔpilA and ΔpilA ΔflgK mutants were also examined. The motility diameter of ΔpilA was not significantly different from WT, and the double mutant phenocopied the single flgK mutant, suggesting that flagella are primarily responsible for the motility observed here.
P. aeruginosa biases the directionality of movement up a concentration gradient of S. aureus-secreted factors
Plate-based macroscopic motility assays measure both an absolute increase in motility and directional movement up a self-generated gradient (chemotaxis) as bacterial populations metabolize the available substrates and expand outward radially (Shapiro 1984). While flagella-based movement though liquid has been extensively described in P. aeruginosa for a variety of chemoattractants, directional movement on a surface is poorly understood. Kearns and Shimkets previously reported P. aeruginosa pili-mediated biased movement up a gradient of phosphatidylethanolamine (PE) on the surface of an agar plate (Kearns, Robinson, and Shimkets 2001). To determine if P. aeruginosa is capable of moving up a previously established concentration gradient of S. aureus supernatant, supernatant derived from either WT or the ΔagrBDCA mutant of S. aureus was spotted onto the surface of 1.5% agar (containing medium buffered for pH only, no carbon source). The secreted factors were allowed to diffuse and establish a gradient for approximately 24 hours, prior to inoculating P. aeruginosa onto the surface, as previously described (Kearns, Robinson, and Shimkets 2001). Preferential movement of P. aeruginosa towards supernatant derived from WT S. aureus was observed, but not for medium alone or supernatant derived from the ΔagrBDCA mutant (Figure 5A). The response for the ΔpilA mutant was also examined and all motility was abrogated in this mutant, demonstrating motility observed in this assay is entirely pili-mediated. These data support the hypothesis that P. aeruginosa biases the directionality of TFP-mediated motility up a concentration gradient of S. aureus secreted factors.
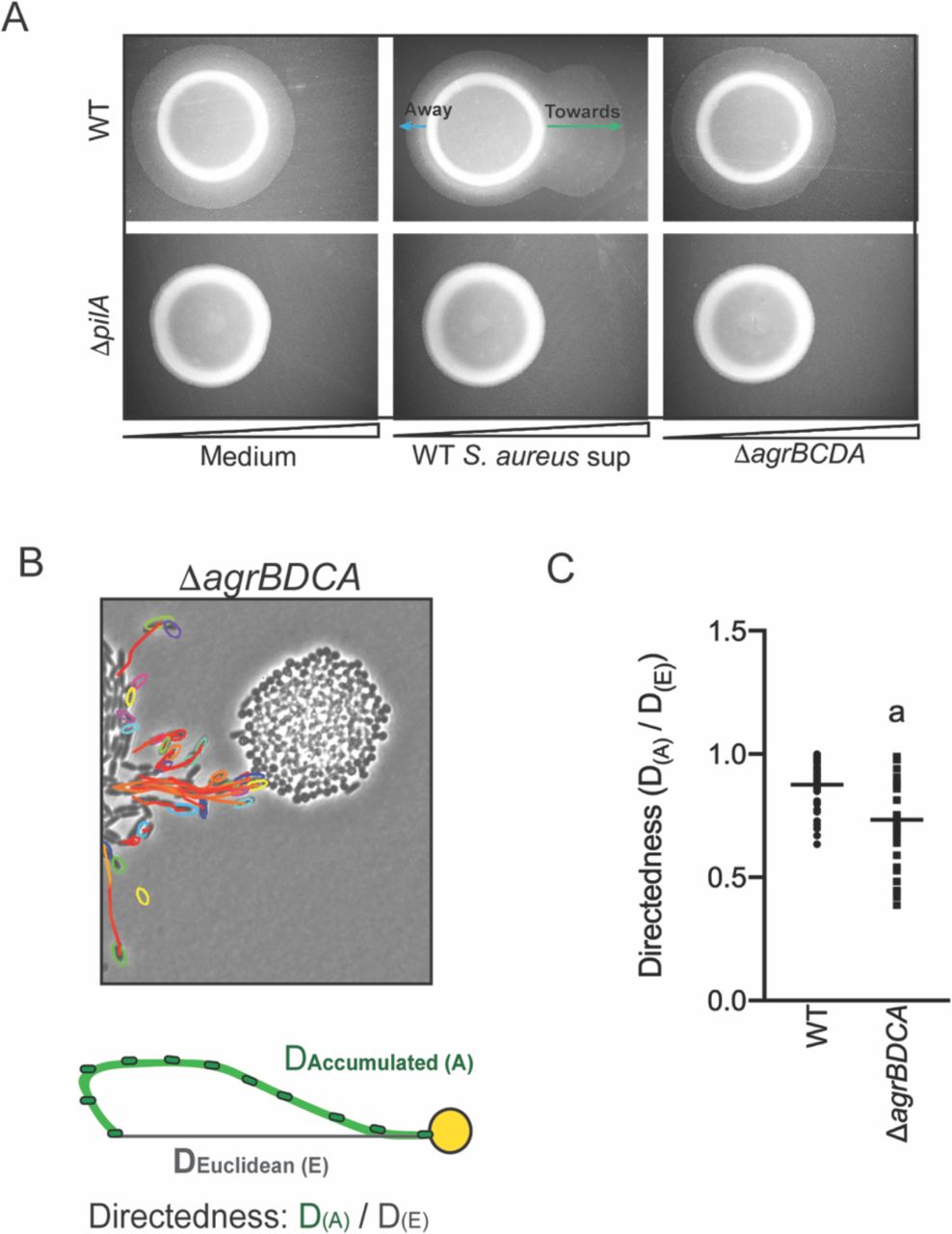
A. A concentration gradient of either S. aureus growth medium (TSB), S. aureus supernatant derived from WT or ΔagrBCDA was established by spotting onto the surface of the agar and allowing a concentration gradient to establish by diffusion for approximately 24h, prior to spotting P. aeruginosa onto the agar (6 mm to the left) and surface-based motility imaged after 24h. Representative images of at least three independent experiments are shown. B. Example of live imaging of WT P. aeruginosa with S. aureus ΔagrBCDA with tracks of single cells shown and schematic illustrating the methods for calculating the directedness. Single P. aeruginosa cells were tracked from first frame a cell exited the raft to the frame where it first encounters S. aureus. The accumulated track distance, D(A), was measured for at least 30 cells in four independent movies and compared to the Euclidean distance, D(E), between the position of the cell in the first and last frame tracked. The ratio of D(A) / D(E) (Directedness) is shown in C for P. aeruginosa towards WT S. aureus compared to ΔagrBDCA with the mean indicated. Statistical significance was determined by an unpaired Student’s t-test (a = P ≤ 0.05).
We next asked if P. aeruginosa would also fail to migrate towards S. aureus deficient in Agr activity at the single cell level. WT P. aeruginosa was visualized in coculture with ΔagrBDCA, as previously performed in the presence of WT S. aureus – with images acquired every 5s for 8 hours (Movie 4 for WT: Figure 2A and Movie 9 for the ΔagrBDCA mutant: Figure 5B). In comparison to WT S. aureus, P. aeruginosa behavior was significantly altered in the presence of ΔagrBDCA. P. aeruginosa remained capable of initiating single cell movement; however, once cells initiated single-cell movement, the path of their movement did not appear as directed towards the S. aureus colonies. In fact, some cells seemed to actively avoid the S. aureus colony all together.
To quantify the directedness of P. aeruginosa movement in the presence of WT and the ΔagrBDCA, single P. aeruginosa cells were tracked from the first frame a cell exited the raft to the frame where it first encounters S. aureus (Figure 5B). The accumulated track distance, D(A), was measured and compared to the Euclidean distance, D(E), between the position of the cell in the first and last frame tracked. The directness of P. aeruginosa towards S. aureus was calculated as a ratio of D(A) / D(E). P. aeruginosa exhibited a higher directedness ratio towards WT S. aureus compared to the ΔagrBDCA mutant (Figure 5C). These data support the hypothesis that Agr regulates the production of S. aureus secreted factors driving the directionality of P. aeruginosa motility.
P. aeruginosa responds to S. aureus with a decrease in cAMP levels
How does P. aeruginosa sense the presence of S. aureus secreted products and initiate TFP-driven exploratory motility? P. aeruginosa encodes three known chemotaxis pathways: two flagella-mediated (che and che2) and a putative TFP-mediated system (pil-chp) (Darzins 1994; Whitchurch et al. 2004). While several proteins encoded in the Pil-Chp pathway comprise a signal transduction pathway very similar (by gene homology) to the flagella-mediated chemotaxis pathway, their role in directional twitching motility remains unclear. A significant challenge lies in that many of these proteins are required for pilus assembly, thus teasing apart their requirement for motility per se verses regulation of a chemotactic response is challenging. Nonetheless, pilJ is predicted to encode the TFP methyl-accepting chemoreceptor protein (MCP) and by homology to the flagella MCPs, PilJ is expected to detect changes in the concentration of an attractant or repellent and initiate a signaling cascade to control twitching motility.
To determine if PilJ is necessary for P. aeruginosa to sense S. aureus, we examined the response of a ΔpilJ mutant to S. aureus (WT) by single-cell imaging. In the presence of S. aureus, the ΔpilJ mutant phenocopied the behavior of WT P. aeruginosa; moving as single cells and with preferred directionality towards S. aureus (Figure 6A, left), suggesting that PilJ is not necessary for exploratory motility. Surprisingly however, when ΔpilJ was imaged in the absence of S. aureus, this strain was capable of adopting many of the behaviors we observed in the presence of S. aureus (i.e. increased single-cell movement, Figure 6A center, Movie 10).

A. Representative snap-shots at t=4h of live imaging of P. aeruginosa ΔpilJ with WT S. aureus (left), ΔpilJ alone (Movie 10, center), and ΔpilJ ΔflgK alone. Red arrows indicate swimming cells. B. Kinetic analysis of cAMP levels in P. aeruginosa cells grown on an agar surface in monoculture and with S. aureus. The cellular cAMP level is expressed as lacP1-lacZ activity divided by the vector control. The mean +/- standard deviation of three biological replicates is indicated. C. Macroscopic twitch assay performed as previously described for the indicated mutants. The mean and standard deviation are indicated for at least three biological replicates. Statistical significance was determined by one-way ANOVA followed by Tukey’s Multiple Comparisons Test - a indicates a statistically significant difference (P ≤ 0.05) between the motility observed in the mutant strain compared to the parental, b indicates a statistically significant difference (P ≤ 0.05) in motility observed in the presence of S. aureus supernatant compared to medium alone, and c indicates a statistically significant difference (P ≤ 0.05) between ΔflgK and ΔflgK ΔpilA in the presence of supernatant.
How might sensing S. aureus and eliminating PilJ have similar phenotypic outcomes for P. aeruginosa? We previously reported that PilJ participates in a hierarchical regulatory cascade of the second messengers, cyclic AMP (cAMP) and cyclic di-GMP (c-di-GMP), which regulate transitions between motility and surface attachment in P. aeruginosa. In the absence of PilJ, cells are predicted to have low intracellular levels of cAMP and c-di-GMP, with a concomitant decrease in surface attachment and increase in flagella-mediated motility (Luo et al. 2015). Indeed, some of the movement in the pilJ mutant had visual appearance attributed in flagella-mediated motility in Figure 2 (red arrows in Figure 6A). To determine if increased flagella mediated-motility is responsible for the phenotypes observed during live imaging of the ΔpilJ mutant, we constructed a ΔpilJ ΔflgK double mutant. While the double mutant had significantly reducing swimming motility in the macroscopic motility assay (Figure S2), its phenotype in the live, single-cell imaging remained similar to both WT P. aeruginosa with S. aureus and ΔpilJ alone, suggesting that the ΔpilJ mutant is not only able to modulate TFP-mediated motility at the single cell level, but also adopts characteristics of exploratory motility.

The indicated P. aeruginosa strains were inoculated into soft agar (0.3%) and incubated for 24h. Representative images are shown of three independent experiments.
Since PilJ is necessary for P. aeruginosa to increase cAMP in response to contact with a surface, we hypothesized that S. aureus might influence P. aeruginosa cAMP levels. To test this model, we utilized a cAMP-dependent promoter fusion, lacP1-lacZ, whose activity has been shown to reflect cellular cAMP levels (Fulcher et al. 2010). We deduced cellular cAMP levels based on the fold change of lacP1-lacZ activity over a vector control, as reported (Fulcher et al. 2010; Luo et al. 2015), and measured these levels over time in the presence of S. aureus, while grown on the surface of an agar plate. For the first eight hours of coculture, P. aeruginosa cAMP levels in the presence of S. aureus were indistinguishable from P. aeruginosa grown in monoculture (Figure 6B). However, after eight hours, cAMP levels continue to rise when P. aeruginosa is grown in monoculture, as previously reported (Luo et al. 2015), but when cocultured with S. aureus, a similar increase in cAMP was not observed. Although the conditions and kinetics of this bulk cell assay are delayed from the single-cell live imaging, these data support the hypothesis that P. aeruginosa responds to S. aureus by modulating cAMP levels.
To further examine a role for P. aeruginosa cAMP modulation in response to S. aureus, we examined the ability of the ΔpilJ mutant to respond to S. aureus supernatant in the macroscopic twitching motility assay. As previously reported (Fulcher et al. 2010), a ΔpilJ mutant is deficient in macroscopic twitching motility; however, consistent with the live imaging, the ΔpilJ mutant remained capable of responding to S. aureus supernatant (Figure 6C). We also tested the double ΔpilJ ΔflgK mutant, which demonstrated a reduced response, in comparison to the single ΔpilJ mutant. However, it maintained the ability to respond to S. aureus, in comparison to the double ΔpilA ΔflgK mutant. These data support a model whereby the ΔpilJ mutant maintains the capacity to respond to S. aureus and to undergo TFP-mediated motility. To formally examine the role for cAMP, we tested a mutant in the cyaB gene, encoding the primary adenylate cyclase required for the generation of cAMP and a mutant in the cpdA gene, encoding the phosphodiesterase required to convert cAMP into ATP (Fulcher et al. 2010). Consistent with our model, ΔcyaB (low cAMP), demonstrated an enhanced response to S. aureus (Figure 6C). However, the phosphodiesterase mutant, predicted to have high cAMP (ΔcpdA), phenocopied the WT. These data support a model whereby S. aureus reduces cellular cAMP at the level adenylate cyclase and not though activation of CpdA (see Model, Figure 8 and discussion).

A. The motility of laboratory strain P. aeruginosa stain PA14 was monitored in the presence of undilute cell-free supernatant from clinical S. aureus CF isolates, CFBRSA47, 48, and 50, in comparison to the positive control S. aureus strain USA300 LAC. B. The motility of clinical P. aeruginosa isolates (CFBRPA38, 43, 44, and 37), in the presence of cell-free supernatant derived from S. aureus USA300 LAC (0.5 dilution) is indicated, in comparison to P. aeruginosa PA14. The mean and standard deviation are indicated for at least three biological replicates. Statistical significance was determined by one-way ANOVA followed by Tukey’s Multiple Comparisons Test - a indicates a statistically significant difference (P ≤ 0.05) between the motility observed in the presence of S. aureus supernatant compared to medium alone, and b indicates a statistically significant difference (P ≤ 0.05) between the motility observed in the CF isolates, in comparison laboratory strains (S. aureus USA300 LAC in A and P. aeruginosa laboratory strain in B).
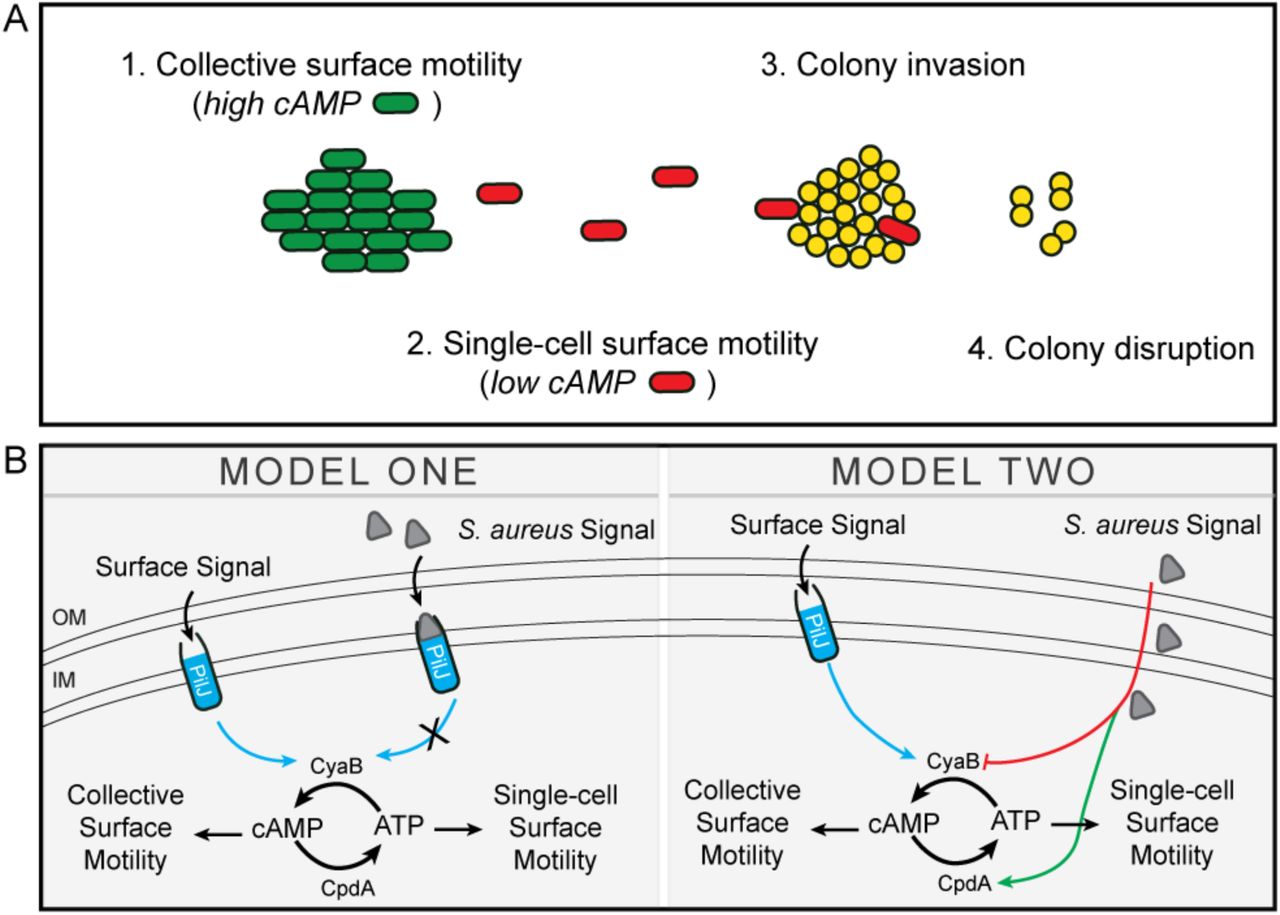
A. In monoculture P. aeruginosa minimally traverses surfaces via collective TFP-mediated motility in large rafts, modulated in part, through increased levels of the second messenger, cAMP. When P. aeruginosa senses the presence of S. aureus, single cells exit the group (with an associated decrease in cAMP), move towards S. aureus, invade, and disrupt the colony. B. Two proposed models for how S. aureus signals modulate cAMP. In Model One, S. aureus signals bind to the chemoreceptor, PilJ and inhibits its ability to activate the adenylate cyclase, CyaB, resulting in decreased cAMP levels. In Model Two, S. aureus signals lower cAMP levels through an unknown, PilJ-independent mechanism by either decreasing activity of the adenylate cyclase, CyaB or increasing the activity of the phosphodiesterase, CpdA (or both).
Clinical isolates of P. aeruginosa and S. aureus perform exploratory motility
Our data thus far demonstrate the P. aeruginosa can sense S. aureus from a distance and modulate both flagella and TFP-mediated motility. To begin to examine if these behaviors might be relevant during airway infection in CF patients, we sought to determine if S. aureus isolates from CF patients are capable of inducing P. aeruginosa motility in the macroscopic twitching assay. Supernatant derived from two out of three clinical CF S. aureus isolates (each from different patients) was capable of inducing motility of the WT P. aeruginosa laboratory strain, to an extent not significantly different from that previously observed with WT S. aureus (Figure 7A). Since we previously observed Agr was necessary for S. aureus to promote motility, we hypothesized that CFBRSA48 was unable to induce P. aeruginosa due to reduced activity of the Agr quorum sensing system – an adaptation previously reported for S. aureus CF isolates (Nair et al. 2011; Goerke and Wolz 2010). Since Agr also positively regulates the production of S. aureus α-hemolysin, hemolysis on sheep blood agar plates is often used as an indicator of Agr activity (Peng et al. 1988). However, all three clinical isolates maintained WT levels of hemolysis, suggesting Agr is active in these strains and an unknown mechanism accounts for reduced activity towards S. aureus in CFBRSA isolate 48 (Figure S3).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Laboratory S. aureus isolates USA300 LAC (WT) and ΔagrBCDA and clinical CF S. aureus isolates (CFBRSA47, 48, and 50) were grown on sheep’s blood agar and examined for hemolysis of red blood cells surrounding the colonies.
Next, we examined the capacity of a panel of P. aeruginosa isolates from CF patients to respond to S. aureus. Phenotypic loss of twitching motility in monoculture is often observed in chronic P. aeruginosa isolates from CF patients (Mayer-Hamblett et al. 2014), thus it was not surprising that two of the four isolates examined exhibited no detectable twitching motility in the absence of S. aureus. Nonetheless, each P. aeruginosa isolate was able to respond to S. aureus supernatant, although to varying degrees (Figure 7B). Importantly, even mucoid P. aeruginosa isolates (38, 43, 37), exhibited at least a two-fold increase in motility in the presence of S. aureus supernatant, above medium alone.
Discussion
Current massive sequencing efforts yield unprecedented information regarding microbial community composition during infection, but fail to provide the necessary information required to functionally understand microbial behaviors in mixed species communities. Moreover, while the bulk assays most frequently utilized to study microbial interactions have provided insight into how bacterial species can influence community survival, metabolism, or virulence factor production (Hotterbeekx et al. 2017), we lack detailed information regarding how a single cell responds to the presence of another species. Here, we developed quantitative live-imaging methods to visualize the behaviors of two clinically important organisms and track their behavior overtime. Through these studies, we uncovered microbial behaviors that could not have been predicted from bulk assays alone. We observed that P. aeruginosa is able to sense S. aureus from a distance by responding to secreted factors. P. aeruginosa is then able to tune its motility patterns, adopting a behavior we refer to as ‘exploratory motility’. The studies presented here open many questions regarding how P. aeruginosa is able to sense and respond to other microbial species. Our current working model is illustrated in Figure 8 and some of the many questions produced from these studies are discussed in detail below.
What interspecies signals does P. aeruginosa sense and respond to? Our data thus far reveal that P. aeruginosa can sense secreted products from S. aureus. We also show that these products are regulated by the Agr quorum sensing system in S. aureus. Identification of the nature of these interspecies signals, as well as investigation into how conserved these processes are among other microbial species is underway. Our current data suggest that P. aeruginosa can sense multiple secreted factors from S. aureus. This hypothesis is supported by the observation that an Agr-deficient strain is unable to induce motility in the macroscopic motility assays, but remains capable of promoting single cell motility during live imaging. Thus, it is formally possible that separate signals are required for initiating single-cell motility and directional motion. Live imaging also suggests that P. aeruginosa cells may actively avoid Agr-deficient S. aureus, raising the possibility that the Agr system negatively regulates a secreted factor that is unfavorable for P. aeruginosa.
When P. aeruginosa senses S. aureus signals, we observed a transition from collective TFP-mediated movement to single-cell motility. TFP-mediated motility is most often described as a behavior where cells are primarily motile only in large groups. The behaviors for WT P. aeruginosa in monoculture observed here are consistent with previous descriptions of twitching motility, including the movement of cells in rafts, preferentially aligned along their long axis, and group tendril formation at the edges of the expanding community (Burrows 2012). In contrast, in the presence of S. aureus, P. aeruginosa cells were seen to exit an expanding raft and transition to single-cell motility. In the process of interrogating P. aeruginosa genes necessary for response to S. aureus, we also uncovered pathways which were necessary for P. aeruginosa transition between collective and individual movements, in the absence of S. aureus. First, while a P. aeruginosa mutant deficient in TFP was unable to exhibit characteristics of twitching motility, the cells also exclusively remained tethered to the group while dividing and expanding outward. However, when flagella were also deleted from this strain, the community appeared to lose its integrity and fall apart. We had initially predicted that maintenance of cells within a group under these conditions (cells sandwiched between a 2% agarose pad and a coverslip) resulted from immobilization of the cells under the pad, preventing P. aeruginosa from generating sufficient force to become motile by either the TFP or flagellum to move away from the group. However, the observation that the ΔpilA ΔflgK mutant was unable to retain group structure suggests the flagella may be important to actively maintain cell-cell contact during collective movement on a surface.
Additional insight into P. aeruginosa regulation of collective verses single cell behaviors was revealed through investigation of a P. aeruginosa strain deficient in PilJ. PilJ is a predicted chemoreceptor, by gene homology to the flagella chemotaxis system (Darzins 1994); however, its functional role in chemotaxis remains unclear. Instead, PilJ participates in sensing surfaces and relaying this information to the cell via modulation of cAMP (Luo et al. 2015; Persat et al. 2015). PilJ is required for twitching motility in macroscopic twitching motility assays (Fulcher et al. 2010; Kearns, Robinson, and Shimkets 2001); a phenotype we confirmed here in monoculture. However, in both our macro- and microscopic assays, the ΔpilJ mutant maintained the ability to undergo TFP-mediated motility in the presence of S. aureus. In fact, the ΔpilJ mutant was visualized to retain motility patterns characteristic of TFP-mediated motility in monoculture during live-imaging. Together these data suggest that the ΔpilJ mutant is indeed capable of performing TFP-mediated motility, but fails to expand outward sufficient to generate a macroscopic motility zone under standard macroscopic conditions – a phenotype characteristic of classic chemotaxis systems.
We also observed that similar to the ΔpilA ΔflgK mutant, the ΔpilJ mutant failed to remain as a group during live imaging, but instead initiated single cell movement. However, once the pilJ mutant (and ΔpilJ ΔflgK), exited the raft, the cells traveled with increased speed and directional persistence, in comparison to the ΔpilA ΔflgK mutant whose single cells were unable to remain tethered, but did not significantly move away from their initial location. The ΔpilJ mutant produces reduced levels of the second messengers cAMP and c-di-GMP, resulting in increased swarming motility (Luo et al. 2015), raising the possibility that modulation of the flagellum may influence initiation of single-cell motility in this strain as well, although further studies are necessary to examine this hypothesis.
The observation that S. aureus reduces P. aeruginosa cAMP levels further supports the hypothesis that modulation of cAMP is necessary for exploratory motility. However, whether maintenance of low cAMP levels in the presence of S. aureus drives the initiation of single-cell motility on a surface or is concurrent with, is unknown. The observation that the PilJ-deficient P. aeruginosa, which also exhibits low cAMP levels, responds similarly to S. aureus as WT and in monoculture also travels as single cells, leads us to posit two models. First, it is possible that S. aureus secreted factors bind to PilJ and inhibit its ability to increase cAMP levels. Second, S. aureus factors inhibit cAMP production independent of PilJ. In this model, S. aureus could lower cAMP levels by inhibiting CyaA/B or by activating the phosphodiesterase, CpdA. Studies to differentiate these hypotheses are underway (Figure 8).
Myxococcus xanthus is one of the most notorious predatory bacteria – elaborating an array of social behaviors reminiscent of what we observe here for P. aeruginosa. M. xanthus coordinates a cooperative, density-dependent feeding behavior, resulting in propulsion of the cells rapidly through the colony of prey, leading to prey lysis, and nutrient acquisition. When M. xanthus contacts prey cells, pili-dependent reversals are stimulated, which keeps the cells “trapped” near the prey colony, promoting contact-dependent prey killing and increased local concentration of antimicrobials (Muñoz-Dorado et al. 2016). Similarly, we observed that when P. aeruginosa encounters S. aureus, the P. aeruginosa cells appear to mount a coordinated response whereby P. aeruginosa surrounds the S. aureus colony and eventually invades the colony – a behavior referred to as the “wolf pack” strategy. P. aeruginosa produces an arsenal of secreted antimicrobials shown to inhibit the growth of S. aureus, yet these secreted factors alone are insufficient for cellular lysis (Limoli and Hoffman 2019). Thus, it is interesting to speculate that these behaviors function to synergistically increase cellular contacts necessary to kill S. aureus by a contact-dependent mechanism and to locally increase the concentration of secreted products.
P. aeruginosa and S. aureus can be coisolated from the lungs of approximately 30% of patients with CF, which is associated with poor clinical outcomes (Limoli et al. 2016). Whether interactions between these pathogens during infection directly influence pulmonary decline is unknown. However, in addition to regulation of surface behaviors, cAMP is a primary regulator of P. aeruginosa virulence factors (Wolfgang et al. 2003), which may influence pathogenic behaviors for these bacteria during polymicrobial infection. Moreover, during chronic infection we find P. aeruginosa evolves phenotypes more permissive to growth with S. aureus (Limoli et al. 2017), suggesting a potential advantage for P. aeruginosa to interact with S. aureus during infection.
By acquiring a fundamental understanding of how bacteria sense and respond to life with each other, we move closer to learning how to rationally manipulate interspecies behaviors during infection and in the environment. While for CF patients, this may mean preventing P. aeruginosa and S. aureus physical interactions, in other instances, we might bring species together who synergize to produce a beneficial compound.
Materials and Methods
Bacterial strains and culture conditions
P. aeruginosa and E. coli were routinely cultured in lysogeny broth (LB; 1% tryptone, 0.5% yeast extract, 1% sodium chloride) and S. aureus in tryptic soy broth (TSB, Becton Dickenson) at 37°C, with aeration. For coculture assays, both species were grown in TSB or M8 minimal medium (48 mM sodium phosphate dibasic, 22 mM potassium phosphate monobasic, 8.6 mM sodium chloride, 2.0 mM magnesium sulfate, 0.1 mM calcium chloride) supplemented with 1% glucose and tryptone. When necessary for strain construction, medium was supplemented with the following antibiotics: gentamicin (10 µg/ml E. coli; 30 µg/ml P. aeruginosa), ampicillin (100 µg/ml E. coli), carbenicillin (200 µg/ml P. aeruginosa), or chloramphenicol (10 µg/ml S. aureus).
S. aureus genetic manipulation. In-frame deletion of the agrBDCA operon in a S. aureus USA300 LAC strain (JE2) was generated using pMAD-mediated allelic replacement (Arnaud et al. 2004). Briefly, a pMAD deletion vector was created by Gibson assembly (Gibson et al. 2009) using EcoRI/BamHI-linearized pMAD and PCR amplicons of 1kb regions up- and downstream of agrBDCA (primer sets agr-a/agr-b and agr-c/agr-d, respectively). The vector to chromosomally complement this deletion strain was similarly produced using a Gibson assembly on the EcoRI/BamHI-lineararized pMAD vector and the PCR product of the agr-a/agr-d primers. Following the standard protocol of heat shift on selective media, strains with successful deletions and restorations of agrBDCA were verified by PCR analysis and chromosomal DNA sequencing.
P. aeruginosa genetic manipulation. In-frame deletion mutants in P. aeruginosa were constructed via allelic exchange as previously described (Shanks et al. 2006). DNA fragments were amplified from P. aeruginosa PA14 genomic DNA by PCR to generate upstream and downstream fragments of the flgK gene with nucleotide tails complementary to plasmid pMQ30 using the following primer pairs: flgK KO P1/flgK KO P2 and flgK KO P3/flgK KO. PCR products were cloned into pMQ30 by in vivo homologous recombination in Saccharomyces cerevisiae INVSc1 (Invitrogen) as previously described (Shanks et al. 2006). The pMQ30-flgK deletion construct was transformed by electroporation into E. coli strain S17 and introduced into P. aeruginosa ΔpilJ (SMC2992) mutants by conjugation. Integrants were isolated on Vogel-Bonner minimal medium (VBMM; 10 mM sodium citrate tribasic, 9.5 mM citric acid, 57 mM potassium phosphate dibasic, 17 mM sodium ammonium phosphate, 1 mM magnesium sulfate, 0.1 mM cal-cium chloride, pH 7.0) agar with gentamicin (30 µg/ml) followed by sucrose counterselection. Resolved integrants were confirmed by PCR and sequencing.
Live imaging of interspecies interactions
Bacteria were inoculated between a coverslip and an agarose pad and imaged with time-lapse microscopy. Pads were made by adding 900 µl of M8 medium with 1% glucose, 1% tryptone, and 2% molten agarose to a 10 mm diameter silicone mold on a coverslip. A second coverslip was placed on top of the agarose while still molten. Pads were allowed to dry for 1h at room temperature, followed by 30m at 37°C. Meanwhile, bacteria were prepared by growing to mid-log phase in M8 medium with 1% glucose and 1% tryptone, diluting to OD600 = 0.15 in warm media, and mixing P. aeruginosa and S. aureus 1:1. 3 µl of inoculum was spotted evenly onto the center of a warm 35 mm glass bottom dish, #1.5 mm coverglass (Cellvis). The agarose pad was removed from the mold and placed on top of the bacterial inoculum. Bacteria were immediately imaged with an inverted Nikon TiE or Ti2 at 37°C for 8h. Images were acquired with a 100x oil objective (1.45NA) with phase contrast and an ORCA Flash4.0 Digital CMOS camera (Hammamatsu).
Movies were generated and cells tracked and analyzed in Nikon Elements with General Analysis 3. For single cells, binary images were generated and single P. aeruginosa cells were identified and tracking began when the first P. aeruginosa cell exited a raft. Rafts were tracked up until the time point where single cell tracking began. Rafts edges were manually identified. The speed (µm/s), acceleration (µm/s2), mean squared displacement (µm2), accumulated track distance, D(A), and the Euclidean distance, D(E), were measure for at least 30 tracks (for single cells) in four independent movies. The area of S. aureus was determined by dividing the total area occupied in a field of view divided by the number of colonies.
Macroscopic Coculture Assays
P. aeruginosa sub-surface twitch (Turnbull and Whitchurch 2014) and 0.3% soft agar motility assays (Ha, Kuchma, and O’Toole 2014) were performed as previously described with modifications for treatment with S. aureus supernatant. S. aureus was grown in TSB to OD600 = 5.0, centrifuged to pellet cells, and supernatant filtered through a 0.22 µm filter. For soft agar assays, cell-free supernatant was mixed at the indicated concentrations with cooled, but still molten agar prior to pouring. For twitch assays, 100 µl of supernatant was spread onto the bottom of a petri plate before pouring media (1.5% agar). P. aeruginosa was grown to OD600 = 5.0 in TSB and inoculated into motility plates by either stabbing with a toothpick halfway through the agar (0.3%), all the way though (sub-surface). Plates were incubated at 37°C for 24h, followed by 24h at room temperature. The diameter of the motility zones was measured in mm. For twitch plates, agar was dropped out and the P. aeruginosa biomass stained with 1% crystal violet for visualization.
Directional Twitching Motility Assay
Experiments were performed as previously described (R. M. Miller et al. 2008). In brief, buffered agar plates (10 mM Tris, pH 7.6; 8 mM MgSO4; 1 mM NaPO4, pH 7.6; and 1.5% agar) were poured and dried at room temperature for 24h. 4 µl of supernatant or medium (TSB) was spotted on the plates, and gradients were established by incubating the plates at 30°C for 24h. Supernatants were prepared as described above. P. aeruginosa strains were grown to early stationary phase, pelleted, and resuspended in 100 ml of MOPS buffer (10 mM MOPS, pH 7.6, and 8 mM MgSO4) at an OD600 of 1.2. 2 µl of this cell suspension was spotted approximately 6 mm from the center of the supernatant spot. Plates were incubated for 24h at 37°C followed by 24h at room temperature before images were taken of the twitching zones.
ß-Galactosidase activity assays
To measure ß-galactosidase activity of the cAMP responsive promoter P1, 200 µl of mid-log phase P. aeruginosa harboring either the plasmid containing lacP1-lacZ or the empty vector were spread onto the surface of M8 + 1.5% agar, either in monoculture or coculture with equal numbers mid-log phase S. aureus. Cell were incubated for the indicated time points, scraped from the plates and the ß-galactosidase activity was measured as previously described (J. H. Miller 1972) and results were presented as Miller units, except that the reactions were performed at room temperature and the absorbance was measured in 96-well flat-bottom plates using a SpectraMax M2 microplate reader (Molecular Devices). For coculture conditions, in order to account for the contribution of S. aureus cells to the OD600 in the calculation for Miller Units, the OD600 measured for P. aeruginosa in monoculture was utilized. P. aeruginosa growth was not affected by the presence of S. aureus, which was confirmed by selectively plating for CFU on Pseudomonas Isolation Agar.
Competing Interests
The authors declare no financial or non-financial competing interests.
Supplemental Figures
Movie Legends
Movie 1. WT P. aeruginosa in monoculture. Duration 8h. Acquisition interval 15m. Playback speed 3054x.
Movie 2. WT P. aeruginosa in coculture with WT S. aureus. Duration 8h. Acquisition interval 15m. Playback speed 3054x.
Movie 3. WT S. aureus in monoculture. Duration 8h. Acquisition interval 15m. Playback speed 3054x.
Movie 4. WT P. aeruginosa in coculture with WT S. aureus. Duration 10m. 4h post inoculation. Acquisition interval 5s. Playback speed 50x.
Movie 5. WT P. aeruginosa in coculture with WT S. aureus. Duration 10s. 4.5h post inoculation. Acquisition interval 50 ms. Playback speed 3x.
Movie 6. P. aeruginosa ΔpilA in coculture with WT S. aureus. Duration 10s. 4.5h post inoculation. Acquisition interval 50 ms. Playback speed 3x.
Movie 7. P. aeruginosa ΔflgK in coculture with WT S. aureus. Duration 10s. 4.5h post inoculation. Acquisition interval 50 ms. Playback speed 3x.
Movie 8. P. aeruginosa ΔpilA ΔflgK in coculture with WT S. aureus. Duration 10s. 4.5h post inoculation. Acquisition interval 50 ms. Playback speed 3x.
Movie 9. WT P. aeruginosa in coculture with S. aureus ΔagrBCDA. Duration 10m. 4h post inoculation. Acquisition interval 5s. Playback speed 50x.
Movie 10. P. aeruginosa ΔpilJ in monoculture. Duration 10m. 4h post inoculation. Acquisition interval 5s. Playback speed 50x.
Acknowledgements
This work was supported by funding from NIH Grant R37 AI83256 (GAO), CFF Postdoctoral Fellowship LIMOLI15F0, and CFF Postdoc-to-Faculty Transition Award LIMOLI18F5. We thank Dr. Carey Nadell for strain PA14 attB:: PA1/04/03 -mKO and consultation on imaging studies, Drs. Ethan Garner, Gerard Wong, and Jeffrey Meisner for consultation on imaging studies, and Dr. Timothy Yahr for careful reading of the manuscript.
References
Supplemental References




