

Abstract
The world is astoundingly variable, and individuals to whole communities must respond to variability to survive. One potent example of nature’s variability is the massive fluctuations in spruce budworm (Choristoneura fumiferana Clemens) populations that occur over 35 years. We examined how the parasitoid community altered its parasitism of budworm and other caterpillars in response to these fluctuations. Budworm and other caterpillar species were sampled from balsam fir in three plots for 13 years in Atlantic Canada. These caterpillars were reared to identify any emerging parasitoids. We found that the parasitoid community showed a simple linear, and indiscriminate, response (i.e., no preference and so densities purely dictated parasitism rates) to changes in budworm densities relative to other caterpillar species on balsam fir. Furthermore, we observed strong changes in topology and distributions of interaction strengths. These observations suggest parasitoid movement between hardwood stands and balsam fir stands is integral to the population dynamics of budworm. Furthermore, our study remarkably shows that species communities coherently alter species interactions in response to variable resources, fundamentally shifting food web pathways in a manner similar to generalist apex predators.
Introduction
Ecologists have long used equilibrium assumptions to study the complex suite of interactions that make up food webs [1, 2]. Although a reasonable first approach, in fact, both abiotic and biotic conditions are notoriously variable [3, 4]. Despite this, our understanding of how organisms respond to variation remains surprisingly limited. In light of human-driven impacts including climate change that promise to significantly alter this variation [5, 6], it behooves ecologists to embrace variation and to ask how individuals to whole communities respond to both natural variation and changes in this natural variation caused by human modifications.
Community-level responses to variability are a function of individual-level responses and recent work has found compelling evidence that individuals and species behave to take advantage of strongly changing conditions. An example of individuals taking advantage of changing conditions is grizzly bears in Alaska following the ephemeral pulses of salmon densities across the river and stream landscape [7]. The bears track the phenological variation of salmon spawning across space and time, so maximising the bear’s energy intake. Another example is individuals switching their diet depending on the abundances of different resource items, where an individual preferentially consumes whichever resource is at the highest abundance [8, 9]. Again, diet switching can maximize the individual’s energy intake. These individual responses can add together to produce a community-level response. One known example of a community-level response to variability is the impressive convergence of parasitoids on the periodic spruce budworm (Choristoneura fumiferana Clemens) outbreaks on balsam fir (Abies balsamea Miller) [10]. Similar to how many species of birds converge on a full birdfeeder, parasitoid diversity sampled from balsam fir increases when spruce budworm densities increase (the birdfeeder effect). The individual parasitoids likely all respond to the changing densities of budworm in order to maximize their fitness [11] and collectively they converge on high densities of budworm during outbreaks. This convergence leads to a diversity cascade across trophic levels suggesting that such strong changes as budworm outbreaks may produce seemingly coherent community responses as a function of all individual responses.
Because of the possibility of coherent behavior within the parasitoid community, the budworm – parasitoid food web provides an excellent system to examine community-level responses to variability. Budworm have massive and relatively predictable outbreaks every thirty five years, followed by periods of budworm rarity [12]. This cycle is considered to be a predator – prey cycle, where the predator is a complex of natural enemies including insects that parasitize and then kill a caterpillar host (parasitoids) [13]. Although budworm are consumed by many invertebrate predators and birds, about 50% of budworm caterpillars are killed by 50 species of parasitoids [13]. Furthermore, this parasitoid community likely has the strongest response to changing budworm densities because invertebrate predators and birds are more generalist than parasitoids, and birds are also limited in numbers due to territory competition [13, 14]. Therefore, we expect to find a community-level response in this parasitoid community. From Eveleigh et al. [10], which showed a birdfeeder effect, we know the parasitoid community response when budworm are plentiful, but we do not know the parasitoid community response when budworm are rare. In effect, we have not observed what happens when the “bird feed” has gone.
For the birdfeeder effect to occur, the parasitoids either remain dormant when budworm are rare, or move between patches of outbreaking budworm, or find alternative sources of caterpillar hosts when budworm are rare. We can immediately discount parasitoids remaining dormant as a hypothesis for maintaining the birdfeeder effect because no parasitoid of budworm is known to enter dormancy longer than a year [15, 16]. We can also discount parasitoids moving between patches of budworm when budworm are rare because budworm are uniformly rare over their whole range for these longer periods. When budworm are rare, parasitoid populations decrease, but a reserve population of parasitoids could be maintained by attacking other caterpillar species. Parasitoids attacking other caterpillar species is a possibility because we know that of the whole parasitoid community that attacks budworm, some parasitoids are generalists that attack both budworm and other caterpillar species [10, 17, 18]. Therefore, the parasitoid community could converge on high budworm densities and then leave to attack other caterpillars when budworm densities decline. The full birdfeeder effect, including other caterpillar species, has as yet not been examined. Specifically, we do not know the relative attack rates of the parasitoid community on budworm and other caterpillar species as budworm densities change. There is the potential for the whole parasitoid community to flexibly and coherently respond to fluctuating budworm densities by collectively altering the relative parasitism rates of budworm and other caterpillar species. These are fundamental questions because scaling this community-level response to the landscape scale has the potential to moderate the amplitude and severity of budworm outbreaks.
Whereas Eveleigh et al. [10] provided a qualitative examination of the entire budworm food web on balsam fir, and Royama et al. [13] examined the impact of parasitoids on budworm only, in this exploratory study, we aimed to quantify the changing trophic interactions of parasitoids with both budworm and other caterpillars. Here, we take the first step towards this goal by examining the interactions on balsam fir solely. First, we extended the birdfeeder effect pattern observed by Eveleigh et al. [10] to a longer dataset by evaluating whether a reversal of the cascading increases in parasitoid species diversity occurred or not when budworm densities decrease. Second, we examined whether the whole parasitoid community exhibited host preference by caterpillar frequency or type. Third, we evaluated whether the parasitoid community response was an aggregate response and not a single species response by dropping the most abundant species and examining species turnover. Fourth, because total diversity changes, parasitism rates, and species turnover all impact the structure and dynamics of food webs, we examined how the topology and interaction strengths of the budworm food web on balsam fir changed. We analyzed rearing data of budworm and other caterpillar species collected from balsam fir branches sampled from 1982 to 1995. During this time period, balsam fir branches were collected from three plots and a representative sample of budworm and all other caterpillar species were placed into feeding vials to identify any parasitoids that emerged. Importantly, we found that the parasitoid community coherently and indiscriminately tracked changes in relative densities of budworm and other caterpillars. Furthermore, the observed changes in total diversity, parasitism rates, and species turnover produced a rewiring of the budworm food web on balsam fir.
Materials and methods
Study sites
Three plots in balsam fir forests in New Brunswick, Canada were established. Plot 1 was in the Acadia Research Forest near Fredericton (46°00’N, 66°25’W). Balsam fir branches were sampled in this plot from 1982 to 1989. Because budworm caused 60% tree mortality in Plot 1 by the mid-1980s, Plot 2 was added, which was also in the Acadia Research Forest. Balsam fir branches were sampled in this plot from 1986 to 1995. In the late 1980s, the budworm populations in Plot 1 and 2 were so low that Plot 3 was added, approximately 170km farther north near Saint-Quentin (47°29’N, 67°15’W). Balsam fir branches were sampled in Plot 3 from 1988 until 1994 when budworm populations also declined to a low level. All plots had mostly balsam fir but also contained spruces and a variety of hardwood trees [10]. Both the Acadia Research Forest and the Restigouche River watershed (where Plot 3 is located) contained balsam fir dominated, mixed, and hardwood dominated stands [19, 20]. All plots were outside areas of biopesticide application. Full details of the three plots and all sampling and rearing procedures can be found in Lucarotti et al. [21], Eveleigh et al. [10] (SI Materials and Methods) and Royama et al. [13]. Here, we present only a brief synopsis.
Sampling
At the beginning of each season, a group of codominant balsam fir trees were selected in 20 random locations within each plot. Every year and for each plot, before larval emergence from winter diapause, one balsam fir branch from each of the 20 locations was collected. As soon as second instar larvae in the field began emerging from diapause, balsam fir branches were sampled approximately every day until the end of budworm adult eclosion. On each sampling day during the earlier years when budworm populations were high, one foliated mid-crown balsam fir branch from one of the trees in each of the 20 locations was collected. During the later years when budworm populations were low, two or more branches were collected from each location to increase the number of collected budworm larvae at each sample date and location
Laboratory work
All budworm and other caterpillars were collected for rearing from all 20 branches sampled before budworm emergence from winter diapause. For branches sampled after budworm emergence from winter diapause, one of the 20 sampled branches was selected and all budworm and other caterpillars on that branch were reared. If a minimum of 100 budworm were obtained for rearing from this branch, no more branches were selected for collection of caterpillars for rearing. If less than 100 budworm were obtained from the first branch selected, then another branch was selected and all budworm and all other caterpillars from that branch were collected and reared, even if the final total number of budworm exceeded 100. When budworm populations were low, obtaining more than 100 budworm individuals became difficult. As a result, all budworm and all other caterpillars that were found on the sampled branches were collected for rearing. Overall, for every sampling day, all budworm and all other caterpillars were reared from the subset of branches of the 20 collected each sampling day. All collected caterpillars (budworm and other caterpillar species) were individually reared on artificial diet [22] and inspected every weekday for mortality. There was high rearing success of both budworm and other caterpillars because all of these hosts feed on balsam fir and therefore readily feed on the artificial diet. On average, 317 other caterpillars were collected each year. All parasitoids that emerged from any reared caterpillars were morphologically identified to genus and where possible to species. Any parasitoids unidentifiable to genus or species were excluded from our analysis (1.6% of the total number of emergences from spruce budworm or other caterpillars).
Statistical Analyses
Because we were interested in quantifying the trophic interactions of parasitoids that attack both budworm and other caterpillars, we excluded all parasitoid taxa that attacked only budworm or only other caterpillars. The 31 parasitoid taxa (listed in Fig. 4) found to attack both budworm and other caterpillars formed 89% of all trophic interactions with budworm and 80% of all trophic interactions with all other caterpillar species. Using Chao2 (function specpool, R package vegan, version 2.5.2, [23]) to estimate the total potential number of interactions between parasitoids and budworm or other caterpillars, this subsetted dataset captures 50% of the potential interactions between parasitoids and budworm and 63% of the potential interactions between parasitoids and other caterpillars.
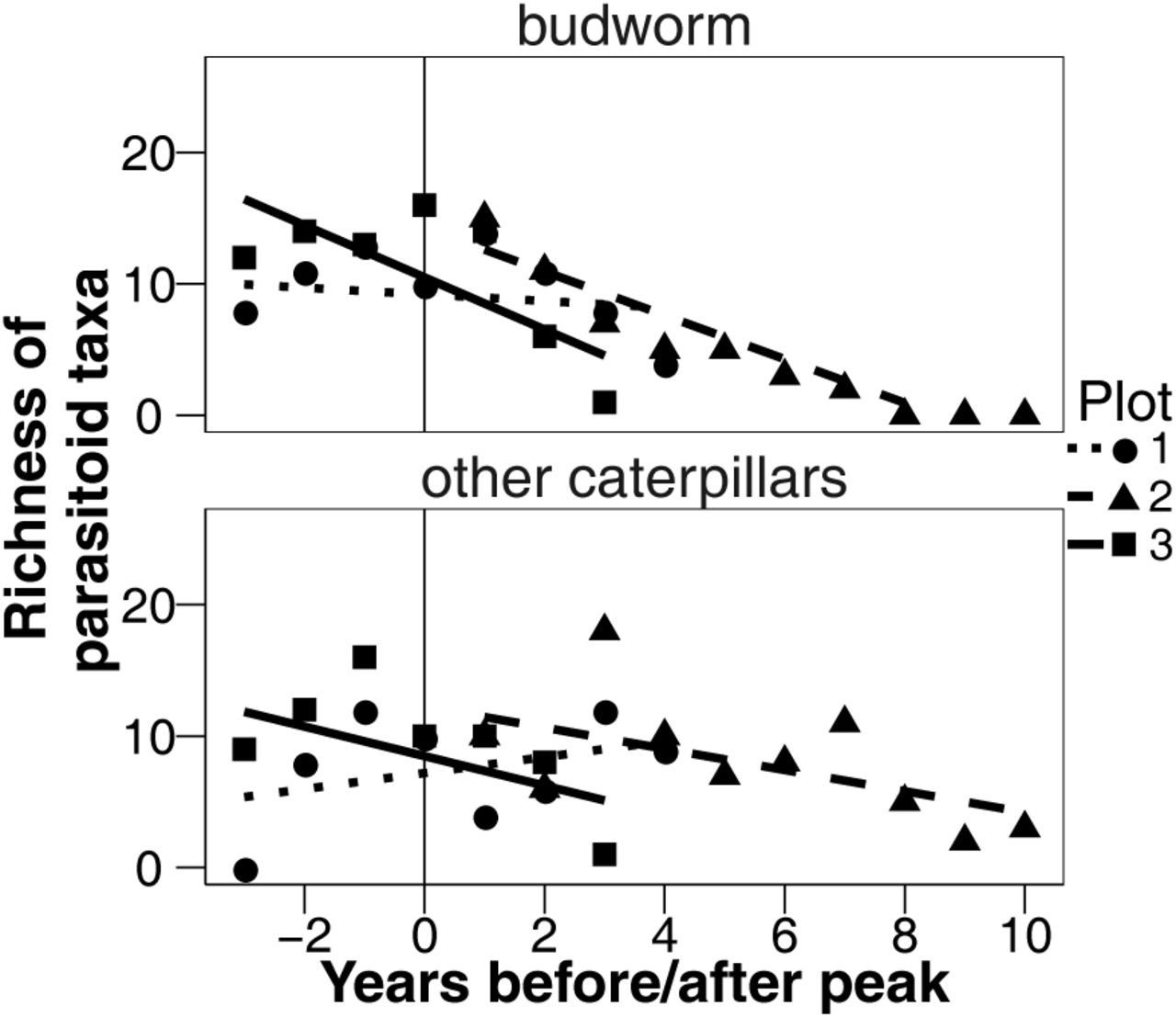
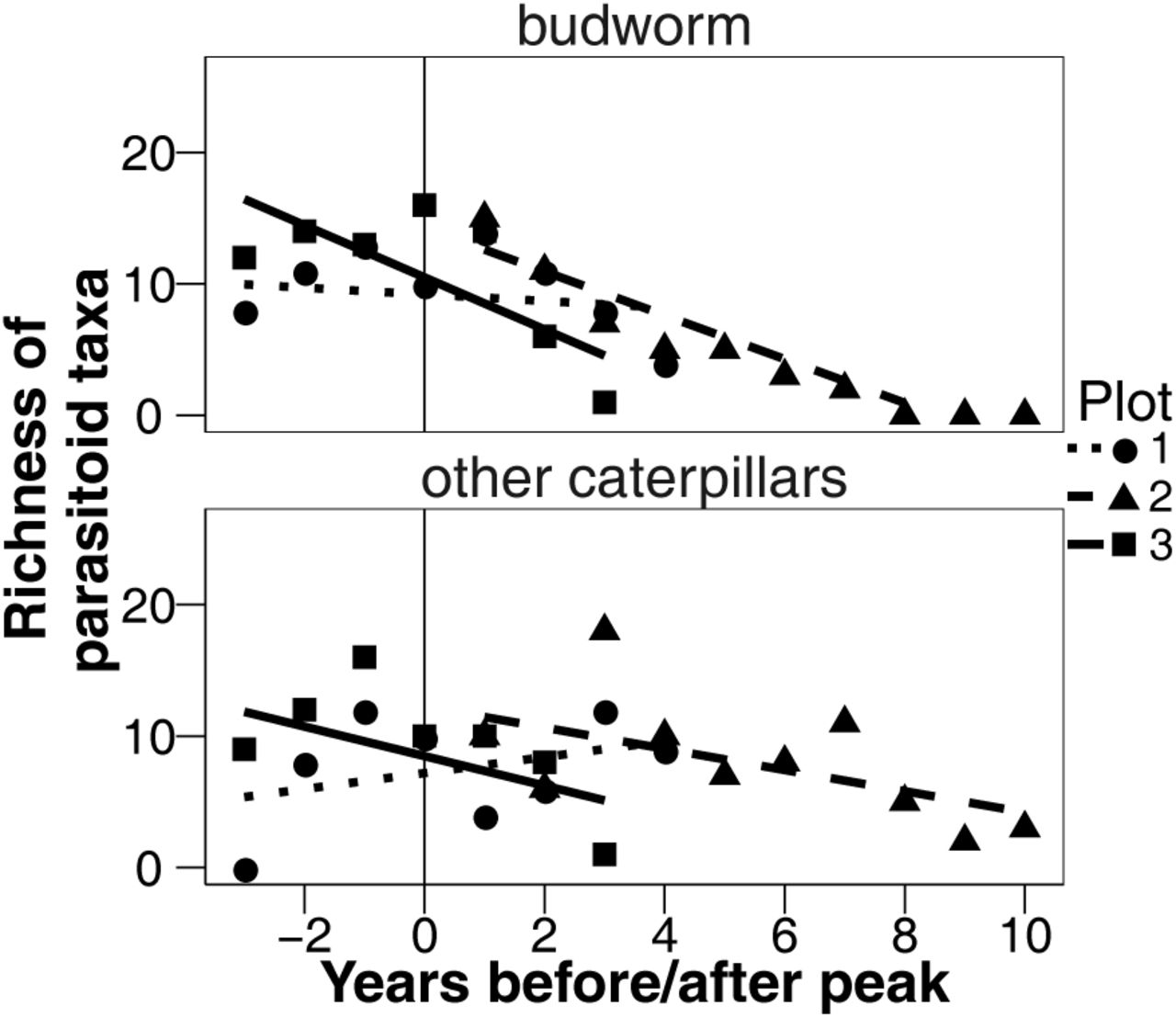
Number of parasitoid taxa that emerged from budworm and other caterpillars over time. Each point is for a single relative year and a single plot. Year zero is the peak of the budworm populations in each plot.

(A) Log10 ratio of parasitoid emergences from budworm to other caterpillars for all parasitoid taxa used in our analysis as a function of the log10 ratio of all sampled budworm and other caterpillars. Each point is a single relative year and a single plot. The thin dashed line is the y = x line. (B) Per capita emergences of parasitoids from budworm and other caterpillars over time. Each point is a single relative year and a single plot. Year zero is the peak of the budworm population in each plot.
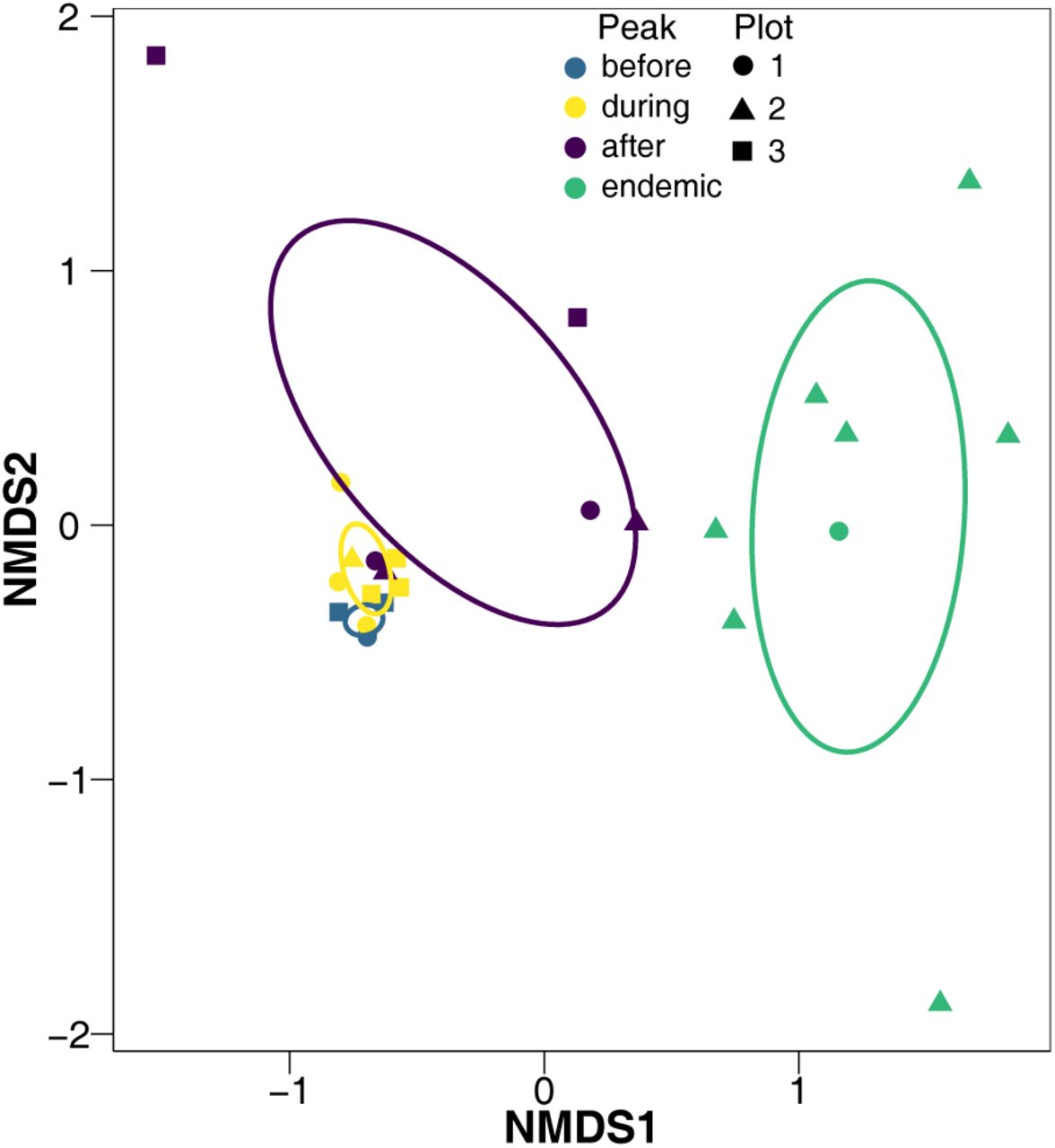
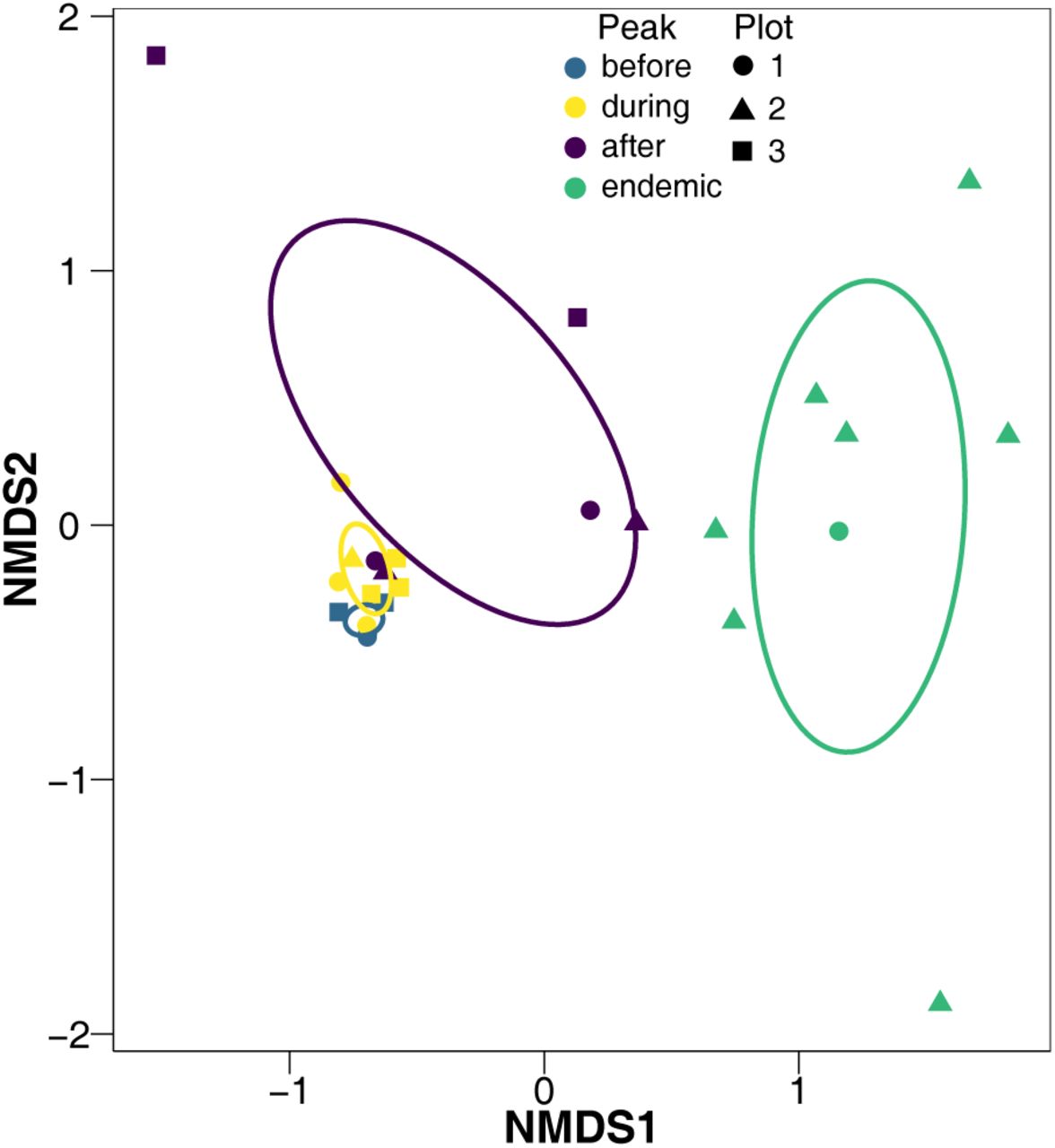
nMDS of parasitoid community over time. The colour of each point and ellipse corresponds to the four temporal groups: three and two relative years before the peak (before); one relative year before and after the peak, and the peak (during); two and three relative years after the peak (after); and four to ten relative years after the peak (endemic). Each point is a single relative year and a single plot. Each ellipse is a covariance ellipse. 26 iterations. Final stress of 0.0798. Instability for preceding 10 iterations was 0.0196.

Graphical representations of the number of emergences of each parasitoid taxon (top boxes) from budworm and other caterpillars (bottom boxes) over time. The width of links is proportional to the fraction of emergences of each parasitoid taxon from either budworm or other caterpillars. The width of the bottom boxes are proportional to the number of emergences from budworm versus other caterpillars, and the percentages show this quantitatively. Four different relative years are shown, where all plots were combined within a relative year: (A) three relative years before the peak, (B) peak relative year, (C) three relative years after the peak, and (D) ten relative years after the peak. All other relative years can be found in Figs S4 & S5. A star denotes a taxon that requires an alternate caterpillar host to overwinter in. To find the corresponding taxon in Eveleigh et al. [10], see Table S1.
Because budworm populations peaked in different years in the three different plots, we created a new time variable called years before/after peak. In this variable, zero was set as the relative year at which budworm populations peaked in each plot. For all analyses, plots were compared using this relative variable. Hereafter, the phrase relative year refers to this created variable “years before/after peak variable”. Plot 1 peaked in 1985 and Plot 3 peaked in 1991. We do not know exactly when budworm populations peaked in plot 2 but because population trends in plots 1 and 2 were nearly identical due to their close proximity, we assumed budworm populations peaked in 1985.
Using the same data, Eveleigh et al. [10] established through rarefaction that changes in diversity of parasitoid species were not due to sampling artefacts. Consequently, we are confident that any patterns found by the analyses below are not due to changes in sampling intensity but due to underlying ecological mechanisms.
All of the following analyses were done using R version 3.5.2 [24]. The data used here can be found on the Dryad Digital Repository (presently, privately archived on Dryad) and the R script can be found on GitHub [25].
Parasitoid taxa diversity
To establish whether the richness of parasitoid taxa emerging from budworm and other caterpillars declined with decreasing budworm densities, we calculated the number of parasitoid taxa that emerged from budworm and other caterpillars for every combination of relative year and plot. We ran a generalized least squares (GLS) regression to test the effects of relative year, caterpillar type (budworm or other caterpillars), plot, and their interactions on the numbers of parasitoid taxa emerging from budworm and other caterpillars (function gls, R package nlme, version 3.1-137, [26]). We added a corAR1 autocorrelation structure to account for temporal autocorrelation. We fitted the full model using maximum likelihood estimation (ML) and then used backwards selection with log likelihood ratio tests (LLRT) to select the final fixed effects. We refitted the final model using restricted maximum likelihood estimation (REML) to give unbiased ML predictors [27].
Parasitoid community host preference
To examine whether the parasitoid community exhibited preference for budworm or other caterpillars on balsam fir, we calculated two values for every combination of relative year and plot: the ratio of parasitoid emergence from budworm to other caterpillars for all parasitoid taxa combined, and the ratio of abundances of budworm to other caterpillars. We ran a GLS regression with the log10 of the ratio of emergence as the response variable and the log10 of the ratio of the abundances of budworm to other caterpillars, plot, and their interaction as the explanatory variables. We fitted the full model using ML, used backwards selection with LLRT to select the final fixed effects, and refitted the final model using REML. We tested whether the average slope for all plots was significantly different from one and whether the average intercept for all plots was different from zero using one sample t-tests. As per the methods in Greenwood & Elton [28], a slope different from one indicates frequency dependent host preference and a intercept different from zero indicates preference for a specific host type (budworm or other caterpillars).
To unpack any parasitoid commmunity host preference response found in the analysis above, we examined how the per capita emergences of parasitoids from budworm and other caterpillars changed over time. For every combination of relative year and plot, we calculated the average per capita parasitoid emergences from budworm and other caterpillars for all parasitoid taxa (per capita emergences being the number of emergences from budworm or other caterpillars divided by the total number of budworm or other caterpillars reared). We ran a GLS regression to test the effects of relative year, caterpillar type (budworm or other caterpillars), plot, and their interactions on per capita parasitoid emergence. We added a corAR1 autocorrelation structure to account for temporal autocorrelation. We fitted the full model using ML, used backwards selection with LLRT to select the final fixed effects, and refitted the final model using REML.
Aggregate response
To identify whether the response to the changing relative abundances of budworm to other caterpillar species was driven by a single parasitoid taxon or by the whole community, we found the three most frequently emerging parasitoid taxa. We then removed in turn the top parasitoid taxon, the top two parasitoid taxa, and the top three parasitoid taxa from the data and ran GLS regressions with the same final model as for the first analysis in the parasitoid community host preference analysis. Using one-sample t-tests, we compared the average slopes and intercepts for all plots of each of these models with the average slope and intercepts for all plots produced in the model with all parasitoid taxa included. To examine turnover in parasitoid taxa over time, we ran an nMDS analysis using the Bray-Curtis dissimilarity measure where the abundances of individual taxa were divided by the total number of parasitoid emergences (all taxa) for each relative year and plot (function metaMDS, R package vegan, version 2.5.2, [23]). We ran a perMANOVA between four groups (function adonis, R package vegan): three and two relative years before the peak; one relative year before and after the peak, and the peak; two and three relative years after the peak; and four to ten relative years after the peak. In this perMANOVA, we used the Bray-Curtis dissimilarity measure, constrained permutations within each plot, and maintained the temporal order of permutations.
Food web topology and interaction strengths
Given the potential for changes in total diversity, parasitism rates, and species turnover to change food web structure, we examined how topology and interaction strengths changed in the budworm food web on balsam fir. We calculated the number of emergences of each parasitoid taxon from either budworm or other caterpillars for every relative year. To examine changes in topology, we produced visual bipartite food webs from these numbers of emergences (R package bipartite, version 2.11, [29]). To examine changes in interactions strengths, we calculated the ratio of the median to maximum interaction strengths for every relative year, where the number of emergences was used for interaction strengths. Note, using the number of emergences or the per capita emergences for calculating the ratio of median to maximum interaction strengths yields the same answer. Calculating the ratio from per capita emergences cancels out the division by the number of sampled caterpillars which is the same for the median and maximum, returning the ratio of median to maximum number of emergences. We used the change in ratio of median to maximum interaction strengths to qualitatively assess how the distributions of weak to strong interactions strengths changed over time.
Results
Parasitoid taxa diversity
The final model explaining the number of parasitoid taxa emerging from budworm and other caterpillars included the explanatory variables of relative year, caterpillar type, plot, the interactions of relative year and caterpillar type, and the interaction relative year and plot (Fig. 1). Graphically, we see that the number of parasitoid taxa that emerged from budworm decreased at a faster rate than the number of parasitoid taxa that emerged from other caterpillars (relative year:caterpillar type interaction, L = 7.988, P = 0.0047, df = 1, log likelihood ratio test, Fig. 1).
Parasitoid community host preference
The final model explaining the log10 ratio of parasitoid emergence from budworm to other caterpillars included the explanatory variables of the log10 ratio of abundances of budworm to other caterpillars, plot, and their interaction (Fig. 2 A). The average slope for all plots, signifying whether the parasitoid community had a statistical frequency preference for either budworm or other caterpillars, was not significantly different from 1 (0.939±0.282, df=15, P = 0.675, one-sample t-test). The average intercept for all plots, indicating whether the parasitoid community had density independent preference for budworm or other caterpillars, was not significantly different from 0 (0.136±0.399, df=15, P = 0.514, one-sample t-test).
The final model explaining the per capita parasitoid emergences from budworm and other caterpillars included the variables of relative year, caterpillar type, plot, and all their interactions (relative year:caterpillar:plot interaction, L = 13.494, P = 0.0012, df = 1, log likelihood ratio test, Fig. 2 B).
Aggregate response
Neither dropping the most abundant parasitoid taxon with the most emergences from all caterpillars (Apanteles fumiferanae), nor dropping the two most abundant parasitoid taxa (A. fumiferanae and Glypta fumiferanae), nor dropping the three most abundant taxa (A. fumiferanae, G. fumiferanae, and Meteorus trachynotus) caused the average slopes and intercepts for all plots to be significantly different from when all parasitoid taxa were included (original average slope was 0.939 and original average intercept was 0.136, Table 1). However, as each most abundant parasitoid taxon was dropped, there was a trend for decreasing slopes and intercepts. The community of parasitoid taxa did not differ between before and during the peak, but the parasitoid community in these two periods did differ from after the peak and during the endemic periods (F = 6.198, P = 0.003, 999 permutations, perMANOVA, Fig. 3).
Slopes and intercepts with corresponding standard errors, t statistics, p values, and degrees of freedom when the three most abundant parasitoid taxa were dropped consecutively. The explanatory variables in this model were Log10 ratio of abundance of budworm to other caterpillars, plot, and their interaction. The response variable was Log10 ratio of emergences from budworm to other caterpillars. Taxon 1 is Apanteles fumiferanae. Taxon 7 is Glypta fumiferanae. Taxon 6 is Meteorus trachynotus.
Food web topology and interaction strengths
There were some parasitoid taxa (e.g. Diadegma pulicalvariae) that were ephemerally found in the food web (Figs 4, S4, & S5). Those parasitoid taxa (e.g. Apanteles fumiferanae) that were found in the food web consistently through time, often were found to change as to whether they emerged from both budworm and other caterpillars or just one caterpillar type. The distribution of interactions strengths for budworm and other caterpillars changed from a skewed distribution dominated by weak interactions towards a uniform distribution, though the variation in the median:maximum interaction strengths between sequential years is greater for other caterpillars than for budworm (Fig. 5).

Median:maximum interaction strength over time, for each plot and for each caterpillar type, where the number of emergences was used for interaction strengths. Figure inspired by the median:maximum interaction strength figure in Ushio et al. [30].
Discussion
In our study, we have shown that this boreal insect food web is highly responsive and flexible in time to changing budworm densities. We used long-term data of host/parasitoid abundance and diversity to assess how parasitism rates and trophic interactions changed over the course of a budworm cycle. We found a dramatic aggregated whole community correspondence of parasitism rates with caterpillar relative abundance (budworm:other caterpillar species abundance) and a sharp change in topology and interaction strength distributions on balsam fir as budworm densities fluctuated.
A strong increase in parasitoid diversity was found when budworm densities increased [10]. In this study, as budworm densities decreased, we found a precipitous decline in parasitoid diversity emerging from budworm in comparison to other caterpillars. This pattern is not due simply to a drop in effective sampling effort when budworm densities decline because Eveleigh et al. [10] mass implanted budworm on balsam fir at a time when budworm was rare and found similar low parasitoid diversity. Instead, declining parasitoid populations likely goes part way to explain this drop in diversity. A complementary explanation is implicitly suggested by the birdfeeder analogy. Birds leave a birdfeeder when the feed deplenishes. Similarly, parasitiods likely leave balsam firs when budworm densities decrease. Therefore, combined with Eveleigh et al.’s [10] study, the full extent of the birdfeeder effect can be seen: parasitoid diversity tracks budworm density. Implicit in this birdfeeder effect is a source of parasitoids when budworm densities increase and a destination for the parasitoids when budworm densities decrease. We suggest that hardwood stands are the source and destination, where balsam fir is a softwood tree, and white birch (Betula papyrifera Marshall) and red maple (Acer rubrum Linnaeus) are hardwood trees. Our suggestion stems from two observations. First, we already know that some parasitoid species including Meteorus trachynotus alternate between budworm and other caterpillars on hardwoods every year [31]. We suggest that hardwood stands play a major role not just for these parasitoid species that require alternate hosts but also for the larger parasitoid community. Second, there have been several observations that budworm densities in stands that contained a mixture of softwoods and hardwoods, otherwise known as mixed forest stands, were lower than budworm densities in balsam fir dominated stands during an outbreak, even after accounting for tree densities [10, 32]. Consequently, these researchers hypothesized that there must be greater diversity and abundances of parasitoids in mixed forest stands, maintained by the greater diversity and abundances of caterpillar hosts in mixed forest stands over the full duration of a budworm population cycle. We hypothesize that hardwood trees are an important part of the budworm system by providing alternative caterpillar hosts for the parasitoids that attack budworm.
Interestingly, we found that the parasitoid community overall did not show a preference for budworm or other caterpillars by either frequency or type. This pattern suggests that the parasitoid community indiscriminately attacks budworm and other caterpillars on balsam fir. One implication of this parasitoid community indiscriminate response is for modeling of the spruce budworm system, where we could model the parasitoids as a single entity with a simple linear response to densities of budworm and other caterpillars on balsam fir. Clearly, the addition of hardwood trees would be integral to any future modeling and further work is required to ascertain the parasitoid community’s response to other caterpillars on hardwoods as well. Comparing between plots, plot 3 had a shallower slope and a intercept greater than 0 indicating that the parasitoid community prefered the least abundant host but still had a slight frequency independent preference for budworm. Similarly, Eveleigh et al. [10] and Smith et al. [18] found different responses of parasitoids in plot 3 compared to plots 1 and 2. Eveleigh et al. [10], using morphological methods, and Smith et al. [18], using DNA barcoding methods, found greater parasitoid diversity in plot 3 compared to plots 1 and 2. Plot 3 had a lower dominance of balsam fir compared to plots 1 and 2, but plots 2 and 3 had equal proportions of hardwood trees (Eveleigh et al. 2007 SI Table 1). The similar observations in host preference and parasitoid diversity between plots combined with the observation of lower budworm density and balsam fir mortality in plots 2 and 3 from Eveleigh et al. [10] lead us to speculate that tree composition and likely hardwood trees impacts the parasitoid community. Further support from our data is the increasing and decreasing of per capita emergences of parasitoids on budworm (Fig. 2B). We again acknowledge that population sizes of parasitoids increase and decrease in response to budworm populations. However, we argue that changing parasitoid population densities could not solely produce the increase and decrease in per capita emergence from budworm. Instead, we suggest that immigration of parasitoids from other stands and changes in searching efficiency both could produce this pattern. In support of immigration, Murakami, Hirao & Kasei [33] found greater parasitism rates in a high-patch connectivity oak plot compared to a low-patch connectivity oak plot. Although this study examined dispersal between trees instead of between stands as in our case, the study still highlights the importance of dispersal in host-parasitoid systems. In support of searching effiency, increasing host density plus increasing concentration of released host tree volatiles due to herbivory likely reduce searching time for the parasitoids, increase oviposition rates and lead to higher per capita emergences of parasitoids [34–36]. Whether on hardwood trees or balsam fir trees, other caterpillars are important to the parasitoid community that attacks spruce budworm. Even our study undersamples the interactions between other caterpillars on balsam fir and parasitoids (63% of the potential interactions between parasitoids and other caterpillars were sampled in this study), and yet the interactions of other caterpillars on hardwoods with parasitoids are sampled even less. Consequently, to fully understand budworm dynamics, it is imperative to sample the interactions of parasitoids with other caterpillars on balsam fir and hardwoods.
The indiscriminate response by the whole parasitoid community could be caused either by a few dominant parasitoid taxa or be a summation of all parasitoid taxa responses. When we excluded the three most abundant parasitoid taxa from our dataset, the resultant parasitoid communities still largely exhibited no host preference by type or by frequency. When we examined when and what caterpillar type these three most abundant parasitoid taxa were attacking (see Figs S1, S2, & S3), we see that a trend for decreasing slopes and intercepts is unsurprising. These three parasitoid taxa either were found emerging from caterpillars when budworm were abundant or were found to emerge more from budworm than other caterpillars, thus dropping the slope and intercept. This indicates that parasitoid taxa have differing preferences for budworm and other caterpillars, but collectively, the community exhibits no preference. Corroborating this suggestion, we also found species turnover over time. Parasitoid species turnover on balsam fir is likely produced from the differing preferences of each parasitoid taxon. In support of differing preferences leading to species turnover, Royama et al. [13] also found that no single parasitoid functional group determined the yearly budworm cycle. Instead, as budworm densities changed, there was turnover in the parasitoid functional group that attacked budworm the most, which produced a relatively constant overall parasitism rate of budworm. As a possible mechanism, Royama et al. [13] posited that the profitability of budworm and other caterpillar species changes in time differently for each parasitoid species. Consequently, different parasitoid species would attack budworm at different time periods during the budworm cycle. Theoretical work supports this supposition where two consumers attack a common resource at different rates during the cycling of the resource [37, 38]. Overall, our results suggest that the parasitiod community act coherently in response to fluctuating budworm densities.
The observed large coherent changes in total diversity, parasitism rates, and species turnover should alter the topology and interactions strengths of the food web. Indeed from our study, we can see large changes in topology with many parasitoid taxa emerging from budworm and/or other caterpillars in some years and not others. We also found dramatic shifts in the distribution of interaction strengths over the budworm cycle. When budworm were at high densities, the distribution of interaction strengths showed a dichotomy of strong-weak interaction strengths but skewed with a preponderance of weak interactions. As budworm densities declined, the distribution of interaction strengths became uniform. We acknowledge that spatial sampling effort (in terms of number of plots) differs between years and because different plots were sampled at different times along the budworm cycle, plot identity may impact the interactions found. However, there were several relative years when multiple plots were sampled in the same relative year (−3 to 4). In these subset of relative years, we see still see a trend of increasing median:maximum interaction strength. Therefore, we predict that even if we had sampled balsam fir in all three plots from budworm population peak to trough, then we would still find a change from skewed interaction strengths distributions to uniform. Similarly, Ushio et al. [30] found that interaction strengths distributions in a marine fish community were dominated by weak interactions in the summer and were more uniform in the winter. A skewed distribution dominated by weak interaction strengths is thought to stabilize foods webs [30, 39, 40]. However, apart from Ushio et al.’s [30] study, the pattern of fluctuating interaction strength distributions has not been well explored. Ushio et al. [30] posited a few drivers for these fluctuations including higher productivity in the summer months leading to higher fish abundance [41] and behavioral or physiological responses that vary over time [42–44]. We agree with Ushio et al.’s [30] assessment. Greater budworm densities could be thought of as the same as high fish abundance in the summer. We suggest too that the behavioral responses by the individual parasitoids are integral to the fluctuations in interaction strength distributions.
Theory may help to further explain this coherent community response. One theoretical model proposes that higher trophic level generalist apex consumers react to variation in their resources by either starting consumer interactions with a species in one separated subgroup of an entire food web (coupling to a resource compartment) or stopping consumer interactions with a different species in another separate subgroup of the entire food web (decoupling from a resource compartment) [45, 46]. This coupling and decoupling of different resource compartments can mute large population variation in lower trophic level organisms and so can stabilize food webs. In the budworm – parasitoid food web, although individual parasitoid species may be specialists or generalists, the collective parasitoid community acts as a generalist consumer and can couple or decouple the resource compartment with balsam fir as the basal resource (hereafter referred to as balsam fir resource compartment). This theoretical model suggests multiple resource compartments, more than the single balsam fir resource compartment that we have examined in this study. We suggest that the separate resource compartment has hardwood trees as the basal resource. Another framework for understanding the dynamics of the parasitoid community is rewiring of the food web, where rewiring is defined as large changes in food web structure that alter nutrient and energy channels in an ecosystem [47]. The rewiring framework has been applied to changes in food web topology and interaction strengths spatially due to climate change. Here, there is likely a periodic rewiring of the interactions of parasitoids with budworm on balsam fir and other caterpillars on hardwoods, due to the cyclical fluctuations in budworm densities.
The parasitoid community response to changing budworm populations illustrates the fantastic flexibility of food webs. Previous research found that as budworm densities increase on balsam fir, the diversity of parasitoid species found on balsam fir increase at all trophic levels [10]. In times of budworm rarity, parasitoid species diversity on balsam fir drops and yet the parasitoid community must be maintained by some mechanism otherwise the swift parasitoid community response to increased budworm abundance could not occur. Our study revealed that the parasitoid community responded to changing densities of budworm by linearly and indiscriminately following the relative densities of budworm and other caterpillars on balsam fir. Large changes in topology and interaction strengths in the budworm food web on balsam fir resulted from the changes in total diversity, parasitism rates and species turnover. The other caterpillar species that these parasitoids attack are not solely found on balsam fir, and in fact, many researchers have suggested that caterpillars on hardwoods should be the dominant resource while budworm are rare. We have corroborated this suggestion by observing that there must be net immigration of parasitoids into balsam fir as budworm densities are increasing. Consequently, further research should include caterpillars on hardwoods and could identify whether the parasitism rates of budworm on balsam fir compared to the parasitism rates of caterpillars on hardwoods change as budworm densities peak and ebb away. Such a response, which appears to be created by the combined actions of all parasitoid species, is an excellent example of community ecology driving the population ecology of a dominant species. For budworm management, we have highlighted the importance of the potential use of other caterpillars by parasitoids to the population dynamics of budworm. Therefore, increasing the abundance and diversity of these other caterpillar species has the potential to mute the amplitude of budworm outbreaks, thus helping to reduce the defoliation and destruction of balsam fir forests in eastern North America.
Author’s Contributions
ESE designed the initial study. ESE and CJL performed the field and laboratory work. CJGG and JF did the statistical analysis with assistance from ESE, MAS, and KSM. CJGG wrote the first draft of the manuscript. All authors contributed to editing the manuscript.
Data accessibility
The data is currently privately archived in Dryad for journal editors and reviewers. If/when the manuscript is accepted, the data will be publically available.
Acknowledgements
We thank the members of the McCann and Eveleigh laboratories for their comments. We are grateful to the many experts for their aid in identifying the insect parasitoids. These experts were J. Barron, A. Bennett, H. Goulet, J. Huber, J. O’Hara, M. Wood, and M. Sharkey. None of this research would have been possible without the many technicians who painstakingly sorted the balsam fir branches searching for caterpillars.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}