

Summary
Programmed cell death (PCD) is widespread during nervous system development, eliminating up to half of the neurons in certain regions of mammalian brains1,2. PCD can serve to counterbalance the surpluses of neural production, and contribute to the formation of correct connectivity3-6. Here we show that cells normally fated to die also represent a reservoir of potential neurons that could contribute to neural circuit evolution. We used as a model the Drosophila peripheral olfactory system, whose lineages exhibit extensive, stereotyped patterns of PCD7-10. Inhibition of developmental PCD is sufficient to generate many new cells in the antenna that express neural markers. Electrophysiological recordings from these “undead” neurons’ sensory dendrites reveal that they exhibit basal and odour-evoked activity similar to wild-type neurons. Transcriptomic and in situ analyses demonstrate that undead antennal neurons express a subset of olfactory receptor genes, including those expressed naturally in other olfactory organs in adults and larvae. Intriguingly, this subset is enriched for relatively young gene duplicates that are normally co-expressed in wild-type neurons. Undead neurons therefore provide a potential cellular substrate to allow the switch of receptors between sensory organs or life stages, as well as accommodate recently-generated receptor genes. Finally, we show that undead neurons can extend axons to novel regions in the primary olfactory centre in the brain, where they may form synaptic connections with second order projection neurons. These data indicate that undead neurons retain a molecular programme that enables their functional integration into the extant olfactory system, raising the possibility that alterations in PCD patterning during evolution is a simple way to generate new sensory pathways. Consistently, comparative analysis of homologous olfactory lineages across the drosophilid phylogeny revealed multiple independent examples where the presence of an additional neuron is consistent with evolutionary fate changes from PCD to a functional olfactory sensory neuron.
A fundamental way in which nervous systems evolve is through increases in the numbers of neurons11-13. Additional sensory neurons can enable higher sensitivity to environmental signals or lead to functional diversification to support acquisition of novel detection abilities. Increases in central neuron number might underlie diverse enhancements in cognitive abilities14, such as parallel processing and memory storage. The generation of more neurons could be achieved through greater production during development – by increasing the number and/or proliferation of neural precursor cells – a process that appears to underlie neocortical expansion during primate evolution15. Alternatively (or additionally), given the widespread occurrence of programmed cell death (PCD) during neural development1,2, prevention of this process can potentially yield a pool of additional neurons. Consistent with this idea, genetic blockage of PCD in mice or D. melanogaster results in the development of enlarged, albeit malformed, nervous systems16,17. Moreover, in C. elegans, the function of an experimentally-ablated pharyngeal neuron can be partially compensated by a sister cell rescued from PCD by a caspase mutation18.
Here we examined the potential of PCD blockage in the formation of novel neural pathways in the D. melanogaster olfactory system. This model is of interest because its development involves prevalent PCD and its molecular neuroanatomy is well-described. The principal olfactory organ, the third antennal segment, is covered with ∼400 porous sensory hairs (sensilla) of morphologically-diverse classes (Fig. 1a)19. An individual sensillum derives from a single sensory organ precursor (SOP) cell that is specified in the antennal imaginal disc20,21. Each SOP gives rise to a short, fixed lineage of asymmetric cell divisions that produces eight terminal cells with distinct identities8,21 (Fig. 1b). Four adopt non-neural (“support cell”) fates, and are involved in the construction of the hair, amongst other roles. The other four cells can potentially differentiate as olfactory sensory neurons (OSNs), which express a single (or rarely two) sensory receptor genes, develop ciliated dendrites that innervate the lumen of the sensillum hair, and project axons towards a specific glomerulus in the primary olfactory centre in the brain (antennal lobe).

a, Schematic of the Drosophila third antennal segment showing the different sensory structures.
b, Schematic of the lineage of an antennal disc sensory organ precursor (SOP) giving rise to a sensillum containing two neurons (illustrated on the right). The expression of a subset of molecular markers is shown; Elav is expressed in only three of four neural precursors; one of these (Naa) as well as the Elav-negative cell (Nbb) are eliminated by PCD. The lineage is based upon data from8,9,21.
c, Simplified schematic of the PCD pathway in Drosophila, highlighting the elements relevant for this study. Several intermediate steps between the pro-apoptotic proteins (Rpr, Grim, Hid and Skl) and the executioner caspases are not shown.
d, Elav expression in whole-mount antennae from control (Df(3L)H99/+; the wild-type chromosome here and in other genotypes was derived from a w1118 parent) and PCD-deficient (Df(3L)H99/Df(3L)XR38) animals. Scale bar = 10 μm. Right: quantifications of antennal neuron numbers of the indicated genotypes, including an additional control genotype (Df(3L)XR38)/+). *** indicates p = 0.0007216 for the comparison to Df(3L)H99/+ and p = 0.0013224 for the comparison to Df(3L)XR38/+ (Wilcoxon-sum rank test, corrected for multiple comparisons using a Bonferroni correction). In this and subsequent panels, individual data points are shown, overlaid with boxes indicating the median and first and third quartile of the data; whiskers showing the limits of the distribution.
e, Elav expression in whole-mount antennae from control (peb-Gal4/+) and PCD-blocked (peb-Gal4/+;UAS-miR(grim,rpr,hid)) animals. Scale bar = 10 μm. Right: quantifications of neuron numbers of these genotypes. *** indicates p = 0.0024×10-4 (t-test).
f, Elav expression in whole-mount antennae from control (peb-Gal4/+) and PCD-blocked (peb-Gal4/+;UAS-p35/+) animals. Scale bar = 10 μm. Right: quantifications of neuron numbers of these genotypes. * indicates p = 0.024 (t-test).
There are ∼20 sensillum classes, housing stereotyped combinations of OSNs (Extended Data Table 1)22-24. Of these, only one class (antennal basiconic 1 [ab1]) contains four neurons, with the others containing fewer – mostly two or three – OSNs. The “missing” neurons are removed by PCD ∼22-32 hours after puparium formation (APF)7-10, when OSN terminal fate is established25.
To block PCD during OSN development, we first used animals bearing deletions in the tandem cluster of pro-apoptotic genes (head involution defective (hid), grim, reaper (rpr) and sickle (skl)), which encode transcription factors critical for promoting developmentally-regulated PCD in diverse tissues (Fig. 1c)4,26. Homozygous chromosomal deficiencies that span the entire cluster cause embryonic lethality. However, a trans-heterozygous combination (Df(3L)H99/Df(3L)XR38), which removes both copies of rpr, and one copy each of hid, grim and skl, allowed recovery of a few viable adults. Immunofluorescence on whole-mount antennae with an antibody against the neural nuclear marker, Elav, revealed a clear increase in the number of labelled cells in mutant animals compared to controls (Fig. 1d), indicating that new neurons form when cell death is prevented.
PCD might be impaired in these mutants at any stage of olfactory system development, including during SOP specification. To selectively block the terminal PCD of OSN lineages (Fig. 1b), we down-regulated expression of hid, grim and rpr by transgenic RNAi from ∼18 h APF using the pebbled-Gal4 (peb-Gal4) driver, which is broadly-expressed in post-mitotic terminal OSN precursors27. Blockage of OSN-specific PCD in this manner also led to a significant increase in Elav-positive cells (Fig. 1e). The number of extra Elav-positive cells observed in these experiments (∼200-300, recognising the limits of automated neuron counting in nuclei-dense tissue (Extended Data Fig. 1)) is in line with estimates of the total number of potential undead neurons (∼300-400) (Extended Data Table 1). We further confirmed the role of the PCD pathway in the antenna through expression of the baculoviral caspase inhibitor p3528 with the same driver. peb-Gal4>UAS-p35 (hereafter “PCD-blocked”) animals displayed higher numbers of Elav-positive cells compared to a peb-Gal4 (control) (Fig. 1f), consistent with a caspase-dependent PCD pathway in this sensory organ.
To determine whether these additional Elav-positive cells are functional neurons, we performed single-sensillum electrophysiological recordings. We focused on one class of trichoid sensilla, at1, which houses a single OSN in wildtype animals, due to PCD of the other three potential neurons in the lineage9. This OSN expresses OR67d, a receptor for the pheromone 11-cis-vaccenyl acetate (cVA)29. at1 sensilla are easily recognised by their sparse basal (spontaneous) pattern of spikes of a single amplitude, and the robust train of spikes that occur only upon cVA presentation (Fig. 2a,d). In PCD-blocked animals, these sensilla often contain additional spikes of smaller amplitude (Fig. 2b-c), suggesting the presence of one or more extra, active OSNs (spike amplitude is defined by the OSN not the receptor gene30). Moreover, exposure to a blend of food-derived odours (which activate many different ORs31) led to responses of the undead neurons in about one-third of the tested sensilla (Fig. 2d-e); the non-responding undead neurons may express receptors activated by other stimuli. These observations indicate that blocking PCD can lead to the development of functional OSNs. The variable odour-evoked responses of these undead neurons to food-derived odours (Fig. 2e) suggests that these cells do not have a fixed identity in at1 sensilla but rather express one of several different types of receptors.

a, Representative extracellular electrophysiology traces of spontaneous activity from neurons in an at1 sensillum of control (peb-Gal4/+) and PCD-blocked (peb-Gal4/+;UAS-p35/+) animals. Automatically-detected spikes (see Methods) from the neuron expressing OR67d are shown in blue, and those of the additional, undead neuron(s) in black, as schematised in the cartoon on the left (cells fated to die are shown with dashed outlines).
b, Quantifications of the proportion of sensilla containing either one neuron (grey) or two (or more) neurons (red) in control (peb-Gal4/+) and PCD-blocked (peb-Gal4/+;UAS-p35/+) animals.
c, Quantifications of the spontaneous activity of the indicated neurons for the control and PCD-blocked genotypes.
d, Representative electrophysiology traces from at1 sensillum recordings in control (peb-Gal4/+) and PCD-blocked (peb-Gal4/+;UAS-p35/+) animals upon stimulation with a 0.5 s pulse (black horizontal bar) of the pheromone 11-cis-vaccenyl acetate (cVA) (10-2 dilution (v/v) in paraffin oil) or a mix of fruit odours (butyl acetate, ethyl butyrate, 2-heptanone, hexanol, isoamyl acetate, pentyl acetate; each odour at 10-2 dilution (v/v) in paraffin oil). Automatically-detected spikes from the neuron expressing OR67d are shown in blue, and those of the undead neuron(s) in black.
e, Quantifications of odour-evoked responses to fruit odours (see Methods) in control (peb-Gal4/+) and PCD-blocked (peb-Gal4/+;UAS-p35/+) animals.
To identify the receptor genes expressed by undead OSNs, we performed comparative transcriptomics of whole antennae of control and PCD-blocked animals by RNA-sequencing. As a positive control, we first examined the changes in transcript levels of grim, rpr, hid and skl, reasoning that inhibition of PCD downstream in the pathway should lead to the presence of undead cells expressing mRNAs for these pro-apoptotic genes (Fig. 1c). Indeed, three of these genes showed higher expression levels in PCD-blocked antennae (Fig. 3a).

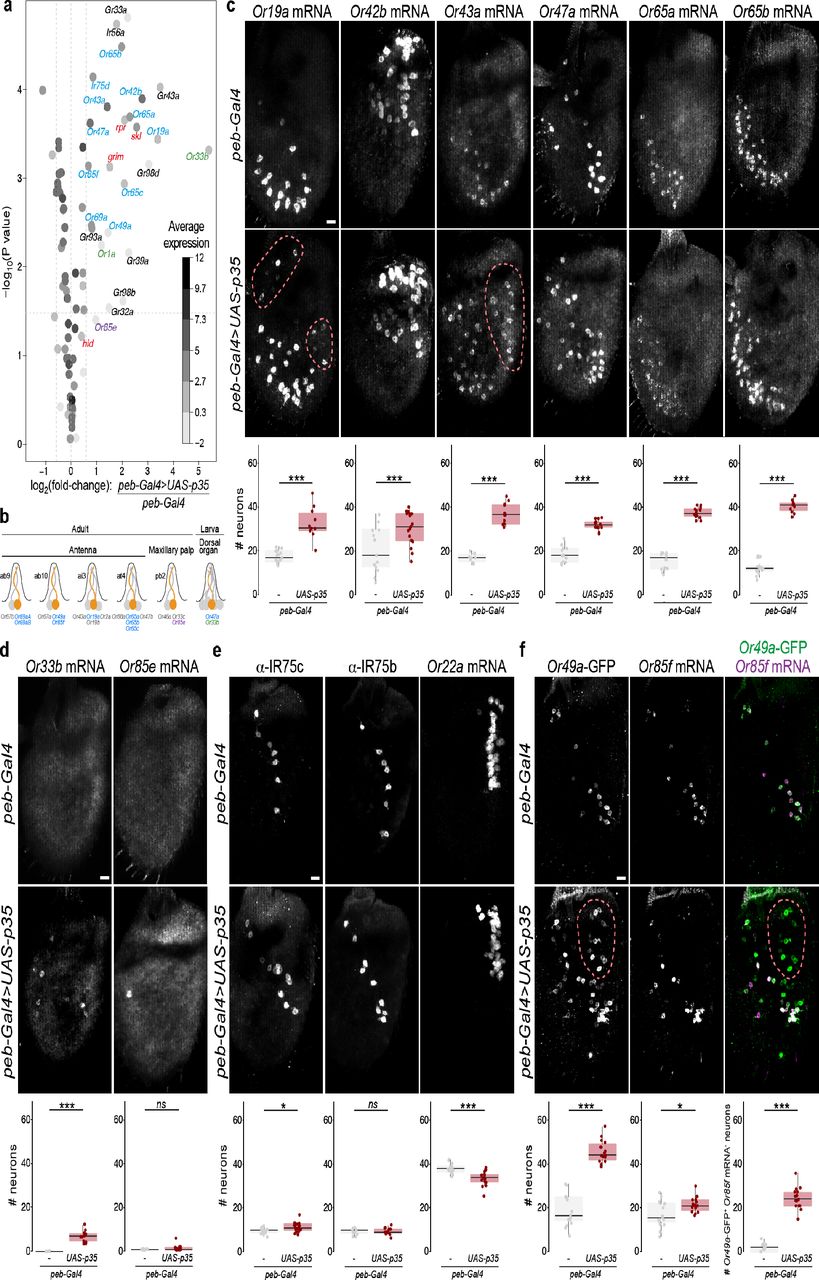
a, Gene expression differences between control and PCD-blocked antennae. The volcano plot shows the differential expression (on the x-axis) of D. melanogaster Or, Ir and Gr gene transcripts (each gene represented by a dot), as well as the four pro-apoptotic genes (grim, rpr, hid and skl; red labels), plotted against the statistical significance (on the y-axis). The mean expression level of individual genes across all samples is shown by the shading of the dot, as indicated by the grey scale on the right (units: log2(counts per million)). Only chemosensory genes showing a >1.5-fold increase in PCD-blocked antennae are labelled: blue labels indicate genes whose expression in the antenna has previously been demonstrated by RNA in situ hybridisation; magenta and green labels indicate receptors normally only expressed in the adult maxillary palps and larval dorsal organ, respectively; black labels indicate receptors that are expressed in gustatory organs. The horizontal dashed line indicates a false discovery rate threshold of 5%. Data for all Or, Ir and Gr genes are provided in Extended Data Table 2.
b, Schematic summarising the normal olfactory organ/sensillum expression pattern of the subset of up-regulated Or genes that display co-expression in wild-type neurons (colour-coded as in a; genes showing no changes in transcript levels are labelled in grey).
c, Representative images of RNA FISH for the indicated Or genes in whole mount antennae of control (peb-Gal4/+) and PCD-blocked (peb-Gal4/+;UAS-p35/+) animals. Scale bar = 10 μm. Quantifications of neuron numbers are shown at the bottom. *** indicates Or19a p = 6.526 x 10-05 (t-test), Or42b p = 0.008486 (t-test), Or65a p = 4.701 x 10-06 (Wilcoxon-sum rank test), Or43a p = 5.888×10-07 (t-test) (see also Extended Data Fig. 2a). The pink dashed line encircles those cells in the PCD-blocked antenna that express the corresponding Ors outside their usual spatial domain (see also Extended Data Fig. 2b).
d, Representative images of RNA FISH for the indicated Or genes in whole mount antennae of control (peb-Gal4/+) and PCD-blocked (peb-Gal4/+;UAS-p35/+) animals. Scale bar = 10 μm. Quantifications of neuron numbers are shown at the bottom.. *** indicates Or33a p = 1.812×10-07 (t-test), Or85e p = 0.053 (Wilcoxon-sum rank test).
e, Representative images of immunohistochemistry or RNA FISH for the indicated olfactory receptors in whole mount antennae of control (peb-Gal4/+) and PCD-blocked (peb-Gal4/+;UAS-p35/+) animals. Scale bar = 10 μm. Quantifications of neuron numbers are shown at the bottom. *, ns, and *** indicate, respectively, IR75c p = 0.01 (t-test), IR75b p = 0.9246 (t-test), Or22a p = 0.0002472 (t-test).
f, Representative images of anti-GFP and RNA FISH for Or85f in whole mount antennae of control (peb-Gal4/+;Or49a-GFP/+) and PCD-blocked (peb-Gal4/+;Or49a-GFP/UAS-p35) animals. Scale bar = 10 μm. Quantifications of neuron numbers are shown at the bottom. *** indicates Or49a-GFP p = 1.444×10-12 (t-test), Or85f p = 0.01375 (t-test), merged panel Or49a-GFP+/Or85f mRNA- population p = 5.48×10-12 (t-test). The pink dashed line encircles those cells in the PCD-blocked antenna that express Or49a-GFP outside its usual spatial domain. We used an Or49a-GFP reporter, due to our inability to reliably detect Or49a transcripts in situ; the higher number of Or49a-CD8:GFP-positive Or85f RNA-positive cells is not an artefact of the analysis method, as an Or85f-CD8:GFP reporter revealed a similarly limited increase in neuron number (Extended Data Fig. 2e).
We next queried the transcript levels for all chemosensory receptors, comprising Odorant receptor (Or), Ionotropic receptor (Ir) and Gustatory receptor (Gr) gene families (Extended Data Tables 1-2). Of the receptors previously detected in antennal neurons in situ22,23,32, we found that 10/36 Ors, 1/17 Irs and 0/3 Grs displayed a >1.5-fold increase in expression, suggesting that only subsets of these receptors are expressed in the undead neurons (Fig. 3a-b and Extended Data Tables 2-3).
To validate these transcriptomic data, we visualised the neuronal expression of several of the Ors in situ. Transcripts for all of those tested by RNA fluorescent in situ hybridisation (FISH) were detected in more neurons in PCD-blocked antennae compared to controls (Fig. 3c and Extended Data Fig. 2a). In some cases, these neurons were found only within the same region of the antenna as the endogenous OSNs (e.g., Or42b, Or65a) while in others (e.g., Or19a, Or43a) undead neurons were observed in novel locations (Fig. 3c and Extended Data Fig. 2b).
Notably, many of the other receptors displaying increases in transcript levels normally act in other chemosensory organs, including one Or expressed in the maxillary palp (Or85e), two larval Ors, and seven Gr genes, which function in various gustatory organs (Fig. 2a and Extended Data Tables 2-3). In situ analysis revealed the presence of transcripts for Or85e and the larval-specific Or33b in populations of undead neurons in PCD-blocked antennae (Fig. 3d).
Beyond these cases, the RNA levels of the vast majority of receptor genes were either unchanged or slightly down-regulated in PCD-blocked antennae (Fig. 3a and Extended Data Table 2). Consistently, in situ analysis of a sample of antennal genes revealed only a very small increase (e.g., Ir75c), no change (e.g., Or13a, Or67d, Ir75b), or a decrease (e.g., Or35a, Or22a) in the size of the corresponding neuron populations (Fig. 3e, and Extended Data Fig. 2c). The latter, unexpected phenotype raises the possibility that undead neurons impact (directly or indirectly) the specification and/or survival of certain populations of neurons.
What properties characterise the small subset of receptors that are expressed in undead neurons? They are normally expressed in neurons housed in diverse sensillum types: basiconic (e.g., Or42b), trichoid (e.g., Or65a), intermediate (e.g., Or19a), and coeloconic (e.g., Ir75d) (Fig. 3b). By contrast, most of these receptors (including 9/10 Ors) are activated in OSNs derived from the Nba precursor cell; the remaining Or (Or43a) and the sole Ir (Ir75d) are expressed in Naa-derived OSNs (Fig. 1b and Extended Data Table 3). This pattern suggests that undead neurons (which are largely Naa-derived (Fig. 1b) preserve gene-regulatory networks that are more similar to Nba/Naa cells than Nab cells, perhaps reflecting the shared Notch activity in Naa and Nba precursors (Fig. 1b)21. Finally, of the 13 Ors detected in undead neurons (including those from other olfactory organs), ten are normally co-expressed with other Or genes, a striking enrichment given the rarity of receptor co-expression within this repertoire22,32. While some co-expressed receptors remain co-expressed in undead OSNs (e.g., Or65a and Or65b (Extended Data Fig. 2d)), this is not always the case. For example, Or19a, but not the co-expressed Or19b, displays up-regulation by RNA-seq (Extended Data Table 2). In addition, in situ analysis of Or49a (using an Or49a-CD8:GFP (Or49a-GFP) reporter22) and Or85f, reveals that while these are always co-expressed in control antennal OSNs, in PCD-blocked antennae there is a novel population of undead neurons that expresses Or49a-GFP, but not Or85f (Fig. 3f).
We next investigated whether undead OSNs project their axons to the antennal lobe. To label these neurons, we used an EGFP gene trap allele of grim (grimMI03811(EGFP)), in which the fluorophore should report on the expression pattern of this pro-apoptotic gene. In controls, grimMI03811(EGFP) expression is detected only at background levels across the antenna; this is expected, as cells that induce grim (and so EGFP) expression are fated to die (Fig. 4a). By contrast, in PCD-blocked antennae, EGFP was detected in many soma (Fig. 4a), which presumably represent the undead neurons previously observed with Elav antibodies (Fig. 1f). In the brains of these animals, we observed that the EGFP-labelled neurons innervate multiple glomeruli of the antennal lobe, indicating that undead neurons can form axonal projections to the primary olfactory centre. Antennal deafferentation experiments confirmed that the specific glomerular signals in PCD-blocked animals were entirely due to the contribution of OSNs (Fig. 4b).

a, Representative images of anti-GFP immunofluorescence in whole mount antennae of control (peb-Gal4/+;+;grimMI03811(EGFP)/+) and PCD-blocked (peb-Gal4/+; UAS-p35/+;grimMI03811(EGFP)/+) animals. Blind scoring by two independent observers of antennae as belonging to control or PCD-blocked group was 100% accurate (see methods). Scale bar = 10 μm.
b, Representative images of combined anti-GFP and nc82 immunofluorescence in whole mount brains of control (peb-Gal4/+;;grimMI03811(EGFP)/+) and PCD-blocked (peb-Gal4/+;UAS-p35/+;grimMI03811(EGFP)/+) animals with intact (left) or excised antennae (right). Blind categorisation of brains (n = 9-12 brains per genotype) as belonging to the control or PCD-blocked set was 95% accurate (2 independent observers). Scale bar = 10 μm.
c, Representative images of combined anti-GFP and nc82 immunofluorescence in whole mount brains of control (peb-Gal4/+;Or49a-GFP/Or49a-GFP;GH146-QF,QUAS-Tomato/+) and PCD-blocked (peb-Gal4/+;Or49a-GFP/Or49a-GFP,UAS-p35;GH146-QF,QUAS-Tomato/+) animals. Blind categorisation of brains (n = 9 and 7 brains, respectively, per genotype) as belonging to the control or PCD-blocked group was 100% accurate (1 observer). Scale bar = 10 μm.
d, Representative images of combined anti-GFP, anti-RFP and nc82 immunofluorescence in whole mount brains of control (peb-Gal4/+;Or49a-GFP/Or49a-GFP;GH146-QF,QUAS-Tomato/+) and PCD-blocked (peb-Gal4/+;Or49a-GFP/Or49a-GFP, UAS-p35;GH146-QF,QUAS-Tomato/+) animals. Scale bar = 10 μm.
e, Representative images of PN soma (bounded by the dashed lines) labelled by GH146>Tomato in whole mount brains of control (peb-Gal4/+;Or49a-GFP/ Or49a-GFP;GH146-QF,QUAS-Tomato/+) and PCD-blocked (peb-Gal4/+;Or49a-GFP/Or49a-GFP,UAS-p35;GH146-QF,QUAS-Tomato/+) animals. Scale bar = 10 μm. Quantifications of neuron numbers are shown to the right. ns indicates p = 0.819 (t-test).
Although the global architecture of the antennal lobe, as visualised by the synaptic marker nc82 (Bruchpilot), is similar in control and PCD-blocked animals, we did detect minor, and somewhat variable, morphological differences, including less distinct boundaries between certain glomeruli and apparently novel regions of neuropil. We wondered whether these differences reflect the innervation patterns of populations of undead OSNs. To test this possibility, we examined the projections of the neurons expressing the Or49a-GFP reporter, which labels many more neurons in PCD-blocked antennae; some of these are different from control neurons as they do not co-express Or85f (Fig. 3f). In control animals, these neurons project to a single glomerulus, DL4, as previously described22. In PCD-blocked animals, labelled axons projected to DL4, as well as to a second, more anterior, glomerulus-like structure (Fig. 4c), which presumably correspond to the wild-type Or49a neuron population and the undead neurons that express this reporter, respectively (Fig. 3f). This observation suggests that undead neurons can acquire distinct fates from control neurons, both by expressing distinct receptor combinations and by forming different glomerular targets in the brain.
We next asked whether such novel OSN axons can potentially synapse with second-order projection neurons (PN). We combined the Or49a-GFP reporter with a genetic driver for the majority of PNs (GH146-QF>QUAS-Tomato) in control and antennal PCD-blocked flies. GH146-labelled processes were detected in the novel Or49a-GFP-labelled glomerulus (Fig. 4d). Moreover, nc82 immunoreactivity, which reflects the presence of the active zone scaffolding protein Bruchpilot33 was also detected in this region (Fig. 4c), implying the formation of synapses between OSNs and these second order neurons. The novel connectivity does not result from the production of additional PNs (Fig. 4e), suggesting that there is no mechanism to match OSN and PN numbers.
Our demonstration that inhibition of PCD is sufficient to allow the development of new functional OSN populations that integrate into the olfactory circuitry is consistent with the hypothesis that modulation of cell death patterns during evolution can be a mechanism to create (or, conversely, remove) olfactory channels. While the variation in OSN number per sensilla within D. melanogaster implies that different SOP lineages have distinct regulation of PCD, we wondered whether we could identify examples of divergent PCD patterning across shorter evolutionary timescales by comparing homologous sensilla in different drosophilids. Previous cross-species analyses suggested this is likely to be relatively rare, as no differences in neuron numbers or pairing were reported in at least a subset of basiconic and coeloconic sensilla in a limited range of drosophilids (although receptor tuning properties do vary) (e.g.,34-36). We therefore performed a broader electrophysiological screening of at1 sensilla in 24 drosophilid species. While the at1 sensilla of most species house a single cVA-responsive neuron (Fig. 5a), similar to D. melanogaster, we identified several species in which this sensillum houses two neurons of distinct spike amplitudes (Fig. 5b), only one of which is cVA-responsive. The lack of genomic of these species currently precludes further molecular analysis, although we assume that cVA-responsive neurons express an OR67d orthologue and the partner neurons a distinct OR of still-unknown sensory specificity. The at1 phenotype in these species is reminiscent of that observed in D. melanogaster when PCD was inhibited (Fig. 2) and provides natural examples of potential changes in PCD patterns leading to novel neuronal circuit elements. Mapping the species whose at1 sensilla house >1 OSN onto a phylogenetic tree, reveals that the acquisition of an additional neuron has occurred independently multiple times during the diversification of the drosophilid clade (Fig. 5c).

a, Representative traces of extracellular recordings of neuronal responses to a 0.5 s pulse (black horizontal bar) of solvent (dichloromethane) or cVA in D. melanogaster and D. subobscura (n = 5). A single cVA-responsive neuron (known or assumed to express OR67d orthologues) is detected (blue spikes), as schematised in the cartoon on the left.
b, Representative traces of extracellular recordings of neuronal responses to a 0.5 s pulse (black horizontal bar) of solvent (dichloromethane) or cVA in D. nasuta, D. pallidipennis and D. testacea (n = 3-5). Two spike amplitudes are detected: a cVA-responsive neuron (assumed to express OR67d orthologues) (blue spikes) and second neuron with a larger spike amplitude, which does not respond to cVA (black spikes), as schematised in the cartoon on the left.
c, Phylogeny of 24 drosophilid species, representing the majority of the Drosophila genus subgroups, based on the protein sequences of housekeeping loci (see Methods). Species names are coloured to reflect the presence of one or two neurons in at1 sensilla. Numbers next to the tree nodes indicate the support values. The scale bar for branch length represents the number of substitutions per site.
A future challenge will be to understand how PCD is determined in OSN lineages and how these mechanisms relate to those defining the fate of the OSNs that survive and express specific receptor genes. Our RNA-seq dataset provides a molecular entry-point to answer these questions by identifying candidate genes expressed highly in cells normally fated to die, similar to the pro-apoptotic factors. Such knowledge is an essential pre-requisite to address how PCD pathways are modified during evolution to selectively eliminate or create OSN populations.
One intriguing observation is that undead neuron populations do not necessarily exhibit functional or anatomical properties that match those of existing OSNs, for example, by expressing receptor genes not normally activated in antennal neurons, or just one of two normally co-expressed receptors. These traits presumably reflect properties of undead OSNs’ “latent” gene regulatory networks. Our work, together with a related study37, reveals the outstanding potential for modulation of cell death patterns to generate new neurons with unique functions and wiring patterns.
Author contributions
L.L.P.-G. and R.B. conceived the project. All authors contributed to experimental design, analysis and interpretation of results. Experimental contributions were as follows: L.L.P.-G. (Fig. 2; Fig. 3e; Fig. 4a; ED Fig. 1); A.F.S. (Fig. 1d-f; Fig. 3e,f; Fig. 4b-e; ED Fig. 1, ED Fig. 2c,e), M.A.K. (Fig. 5 and ED Fig. 3), S.C. (Fig. 3a,c,d and ED Fig. 2a-d). K.B. and S.P. performed RNA-seq data analysis. R.B., L.L.P.-G. and A.F.S. wrote the paper with input from all other authors.
Methods
Drosophila culture
Flies were maintained at 25°C in 12 h light:12 h dark conditions, except where noted. D. melanogaster strains were cultured on a standard cornmeal diet; other drosophilid species were grown on food sources as indicated in Extended Data Table 4 (for recipes: http://blogs.cornell.edu/drosophila/recipes). Published mutant and transgenic D. melanogaster are described in Extended Data Table 4. Df(3L)H99/Df(3L)XR38 (and their controls) were cultured at 22°C to increase the number obtained of adult offspring of the desired genotype. For most histological experiments, only female flies were analysed, to avoid confounding variation due to known sexual dimorphisms24. Mixed genders were used for Df(3L)H99/Df(3L)XR38 flies in Fig. 1d due to the limitation in the recovery of this genotype, as well as for anti-IR75b and anti-IR75c immunofluorescence in Fig. 3e (there is no sexual dimorphism in the numbers of these OSNs). For histological experiments, flies were 1-12 days old. Animals subjected to antennal deafferentation (and control intact flies) were left for 10 days post-surgery to permit degeneration of OSN axons. For the experiments in Fig. 5 and Extended Data Fig. 3, all experiments were carried out with 8-15 day old, mated female flies.
Histology and image analysis
Whole mount antennal immunofluorescence and RNA fluorescent in situ hybridisation were performed essentially as described38. Whole mount brain immunofluorescence was performed essentially as described39. Primary and secondary antibodies are listed in Extended Data Table 5. Sources and/or construction details of templates for RNA probes are provided in Extended Data Table 6. Imaging was performed on a Zeiss confocal microscope LSM710 or LSM880 using a 40x oil immersion objective.
For automated counting of Elav-positive cell bodies, confocal stacks were imported into Fiji40 and passed through a median 3D filter of radius 1 in all dimensions. Images were subsequently thresholded using the 3D iterative thresholding plug-in41, and cells automatically counted using the 3D object counter.
Analyses of OSN numbers expressing specific olfactory receptor genes, and morphological differences of the antennal lobes of control and PCD-blocked animals were performed by experimenters blind to the genotype, using RandomNames.bat(https://github.com/DavidOVM/File-Name-Randomizer/blob/master/RandomNames.bat) to encode image names.
Electrophysiology
Single-sensillum recordings were performed and analysed essentially as described42,43. at1 sensilla were identified based upon their morphology and characteristic distal distribution on the antenna; they could also be clearly distinguished from the only other trichoid sensillum class, at4, which houses three OSNs (Extended Data Fig. 3 and data not shown). Chemical stimuli and solvents are described in Extended Data Table 7. For the experiments in Fig. 2, neuron activity was recorded for 10 s, starting 3 s before a stimulation period of 0.5 s. For the experiments in Fig. 5 and Extended Data Fig. 3, neuron activity was recorded for 6 s, starting 2 s before a stimulation period of 0.5 s. Traces were analysed by sorting spike amplitudes in AutoSpike; representative traces presented in the figures were further processed in Adobe Illustrator CS (Adobe systems, San Jose, CA). Spontaneous neuron activity was quantified by counting spontaneous spikes in a 10 s recording window. Stimulus-evoked activity was quantified by counting spikes in a 0.5 s window during odour stimulation, and then subtracting this count from a 0.5 s recording window just prior to stimulation. For the solvent-corrected quantifications in Fig. 2d-e, the responses to solvent (paraffin oil) were subtracted from the responses to the odour. In Fig. 2b, sensilla were classified as having two neurons if two different spike amplitudes were automatically detected and/or corrected responses to the fruit odour mix were above 20 Hz.
RNA-sequencing and analysis
Antennal RNA was extracted from three biological replicates of control (peb-Gal4/+;Or49a-GFP/+) and PCD-blocked (peb-Gal4/+;Or49a-GFP/UAS-p35) animals. (The increased numbers of neurons labelled by Or49a-GFP was noted in preliminary studies and we therefore incorporated this transgene into the genotypes used in these experiments as an internal control; see below). For each pair of biological replicates, ∼200 animals were grown under identical conditions and RNA was extracted in parallel using 2-5 day old flies, as described9. RNA quality was assessed on a Fragment Analyzer (Advanced Analytical Technologies, Inc.); all RNAs had an RQN of 9.8-10. From 100 ng total RNA, mRNA was isolated with the NEBNext Poly(A) mRNA Magnetic Isolation Module. RNA-seq libraries were prepared from the mRNA using the NEBNext Ultra II Directional RNA Library Prep Kit for Illumina (New England Biolabs). Cluster generation was performed with the resulting libraries using the Illumina TruSeq PE Cluster Kit v4 reagents and sequenced on the Illumina HiSeq 2500 using TruSeq SBS Kit v4 reagents (Illumina). Sequencing data were demultiplexed using the bcl2fastq Conversion Software (version 2.20, Illumina).
Purity-filtered reads were adapters-and quality-trimmed with Cutadapt (version 1.844). Reads matching to ribosomal RNA sequences were removed with fastq_screen (version 0.11.1). Remaining reads were further filtered for low complexity with reaper (version 15-065)45. Reads were aligned to the Drosophila melanogaster BDGP6.92 genome using STAR (version 2.5.3a46). The number of read counts per gene locus was summarised with htseq-count (v. 0.9.1)47 using Drosophila melanogaster.BDGP6.92 gene annotation. The quality of the RNA-seq data alignment was assessed using RSeQC (v. 2.3.7)48.
Statistical analysis was performed for genes in R (version 3.5.3). Genes with low counts were filtered out according to the rule of 1 count per million in at least 1 sample. Library sizes were scaled using TMM normalisation (EdgeR package version 3.24.3)49 and log-transformed with limma cpm function (Limma package version 3.38.3)50.
Differential expression was computed with limma for paired samples by fitting the 6 samples into a linear model and performing the comparison PCD-blocked antennae versus controls. Comparison of read number for GFP (encoded by the Or49a-GFP transgene) was performed by mapping reads to the GFP sequence with Bowtie251: control antennal RNA: 139±9.5 reads/sample (mean ± standard deviation); PCD-blocked antennal RNA: 227±8.7 reads/sample.
Moderated t-test was used for the comparison on a subset of 83 expressed D. melanogaster genes including: Or, lr and Gr genes as well as the four pro-apoptotic genes (grim, rpr, hid and skl). For multiple testing correction, the p-values were adjusted by the Benjamini-Hochberg method, which controls the false discovery rate52. The volcano plot was generated in R by plotting the log2(fold change PCD-blocked vs control) against the -log(p value). Data points were shaded according to mean expression value across all samples.
Phylogenetic analysis
Phylogenetic analysis of drosophilid species was conducted using 6 housekeeping proteins, encompassing two nuclear loci (Adh and Xdh) and four mitochondrial loci (COI, COII, COIII and ND2). Available amino acid sequences from Uniprot (https://www.uniprot.org, accession numbers are listed in Extended Data Table 8) of each species were concatenated in Geneious (v11.0.5). A multiple sequence alignment of 2939 positions was generated using the MAFFT (v7.309) tool with E-INS-I parameters and scoring matrix 200 PAM / K=253. The final tree was reconstructed using a maximum likelihood approach with the GTR+G+I model of nucleotide substitution and 1000 rate categories of sites in FastTree (v2.1.5). The tree was visualised and processed in Geneious (v11.0.5).
Statistics and reproducibility
Statistical analyses and plotting were made in RStudio (v1.1.463 R Foundation for Statistical Computing, Vienna, Austria, 2005; R-project-org), except for the RNA-seq analyses (described above). For statistical analyses, normality was first assessed on datasets using a Shapiro test. If both datasets were normally distributed, a two-sided t-test was performed; otherwise, a Wilcoxon-rank sum test was performed.
Data availability
All relevant data supporting the findings of this study are available from the corresponding author on request. RNA-seq data are available in GEO (Accession GSE128725).
Acknowledgements
We are grateful to I. Alali and M. Erdogmus for technical assistance, Darren Williams and Sînziana Pop for sharing flies and discussions, Raquel Alverez Ocana for sharing information on OSN numbers, Mattias Alenius, the Bloomington Drosophila Stock Center (NIH P40OD018537), and the Developmental Studies Hybridoma Bank (NICHD of the NIH, University of Iowa) for reagents. We thank Roman Arguello and members of the Benton laboratory for comments on the manuscript. L.L.P.-G. was supported by a FEBS Long-Term Fellowship. M.A.K., B.S.H., and M.K. are supported by the Max Planck Society. Research in R.B.’s laboratory is supported by the University of Lausanne, an ERC Consolidator Grant (615094) and the Swiss National Science Foundation.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}