

Abstract
The size of the pupil depends on light level. Watson & Yellott (2012) developed a unified formula to predict pupil size from luminance, field diameter, age, and number of eyes. Luminance reflects input from the L and M cones in the retina but ignores the contribution of intrinsically photosensitive retinal ganglion cells (ipRGCs) expressing the photopigment melanopsin, which are known to control the size of the pupil. We discuss the role of melanopsin in controlling pupil size by reanalysing an extant data set. We confirm that melanopsin-weighted quantities, in conjunction with Watson & Yellott’s formula, adequately model intensity-dependent pupil size. We discuss the contributions of other photoreceptors into pupil control.
In a paper adequately described as a tour de force, Watson and Yellott [1] developed a unified formula to predict pupil size from luminance, field diameter, age, and number of eyes. This letter concerns the parametrisation of the retinal intensity, which in Watson and Yellott’s model is given in terms of luminance, i.e. the radiance of the stimulus weighted by the photopic luminosity curve V(λ). V(λ) corresponds to a mixture of the L and M cones in the retina, thereby largely ignoring the potential role of S cones, rods, and the intrinsically photosensitive retinal ganglion cells (ipRGCs) expressing the photopigment melanopsin [2-4].
The observation that V(λ)-weighted quantities do not predict pupil size is not new [5]. In 1962, Bouma [6] noted that the spectral sensitivity of pupil control is neither V(λ) nor the rod-based V’(λ), interjecting that the outcome of his experiments “may turn out to be related to other adaptive processes in the human eye”. Bouma himself modelled the spectral sensitivity as a combination of S cones and rods. We know now that steady-state pupil size is largely controlled by melanopsin.
To test if Bouma’s data is consistent with melanopsin-based pupil control, we reanalysed the intensity-response curves from Bouma [6] as follows. We first extracted the data from Bouma’s Figure 1 (Figure 1A, B). For monochromatic lights, which we assumed Bouma used, it is simple to convert the reported V(λ)-weighted luminous flux into a melanopsin-weighted radiant flux [7]. As radiant flux describes the total amount of energy emitted by a source, it is not an appropriate measure to describe corneal or retinal intensity, so the absolute quantities are not informative unless a geometry is specified. Allowing for an arbitrary horizontal shift, Watson and Yellott’s model accounts well for the shape of the pupil response as a function of melanopic radiant flux, except for long-wavelength lights (Fig. 1C).
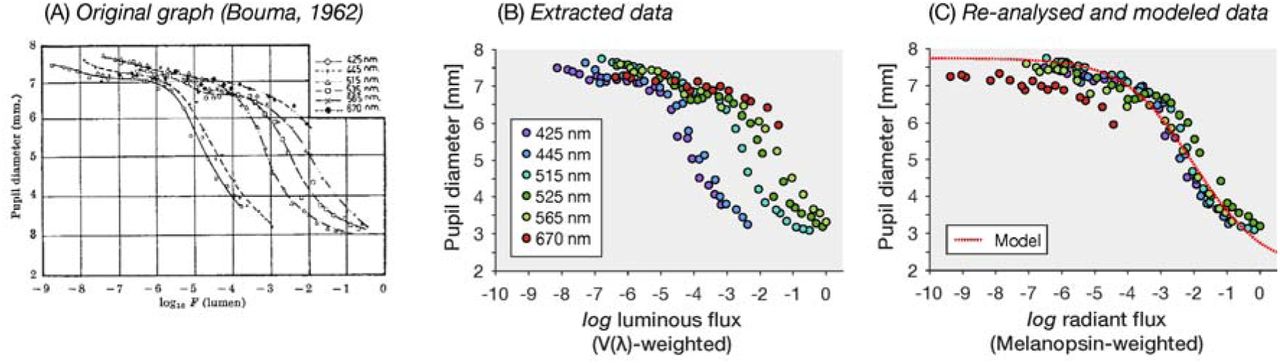
A Original graph from Bouma [6] relating luminous flux to pupil diameter in milimeters. B Replotted extracted pupil size data (using WebPlotDigitizer, https://automeris.io/WebPlotDigitizer/). C Re-analysed (in terms of normalised melanopic radiant flux) and modelled pupil size data.
There is now a good body of evidence that all photoreceptors can control the diameter of the pupil. The best evidence comes from studies examining pupil size using the method of silent substitution, in which pairs of lights are alternated such that only one photoreceptor class is stimulated [8, 9]. Studies examining pupil control using this method are given in Table 1.
Evidence of photoreceptor contributions to pupil control
A key realisation is that while all photoreceptors may contribute to controlling the pupil size, the when and how is important. For example, due to rod saturation [10], rods are not expected to contribute to pupil control at photopic light levels. The temporal regimes in which the photoreceptors contribute are also different. Notably, L+M stimulation is band-pass, while S cones and melanopsin are tuned to low frequencies in driving the pupil [11]. McDougal and Gamlin [12] found that cones and rods account for pupil constriction between 1 and 10 seconds from the onset of the light exposure, at 100 seconds, pupil size is largely controlled by melanopsin with some contribution from the rods.
To what extend does Watson and Yellott’s use of luminance as an input parameter call into question the generalizability of their model? From first principles, differences between V(λ)-weighted and melanopic quantities are largest with monochromatic lights. But we typically do not live under monochromatic illumination. We explored this question by examining the range of melanopic irradiances at a fixed illuminance. In other words, how wrong would we be if we continued using V(λ)-weighted quantities to predict pupil size? Using a database of 401 polychromatic (“white”) illuminant spectra [13], we calculated the range of melanopic irradiance while keeping the photopic illuminance fixed at 100 lux (Figure 2). Across all 401 spectra, a 100 lux light source has a melanopic irradiance of 75.5±23.4 mW/m2. The range of melanopic irradiances is between 20.4 and 164 melanopic mW/m2, i.e. in the worst case a factor of ∼8. Whether or not this worst-case misprediction by using a V(λ)- weighted quantity has tangible consequences depends on the application. Predicting pupil size in a psychophysical experiment at mesopic light levels requires less stringent estimation of retinal intensity than safety-critical calculations.

Variability of the melanopic irradiance of 401 polychromatic “white” light sources [13] at 100 lux.
A recent study reported attempted to derive a formula for predicting pupil size also from melanopsin activation but only focused on a rather narrow luminance range (50-300 cd/m2) [14]. While this is a good start, it might be a useful empirical exercise to collect natural pupil sizes under a large range of illumination conditions (indoors, outdoors) under natural behaviour with cojoint spectral measurements.





{kind=link}
{kind=link}