

ABSTRACT
RNA has been classically known to play central roles in biology, including maintaining telomeres1, protein synthesis2, and in sex chromosome compensation in certain species3,4. At the center of these important biological systems are noncoding RNAs. While thousands of long noncoding RNAs (lncRNAs) have been identified in mammalian genomes5–8, attributing RNA-based roles to lncRNA loci requires an assessment of whether the observed effect could be due to DNA regulatory elements, the act of transcription, or the lncRNA transcript. Here, we use the syntenically conserved lncRNA locus, Functional intergenic repeating RNA element (Firre), that is located on the X chromosome as a model to discriminate between DNA- and RNA-mediated effects in vivo. To this end, we generated genetically defined loss-of-function, gain-of-function, and rescue mouse models for Firre and provide genetic evidence that the Firre locus produces a trans-acting RNA. We report that: (i) Firre mutant mice have cell-specific defects during hematopoiesis and changes in gene expression that can be rescued by induction of Firre RNA from a transgene in the Firre knockout background, (ii) mice overexpressing Firre from a transgene exhibit increased levels of pro-inflammatory cytokines and impaired survival upon exposure to lipopolysaccharide, and (iii) deletion of the Firre locus did not result in changes in local gene expression on the X chromosome in 9 different biological contexts, suggesting that Firre does not function by cis-acting RNA or DNA elements. Together, our results provide genetic evidence that the Firre locus produces a trans-acting lncRNA that has physiological roles in hematopoiesis and immune function.
INTRODUCTION
Transcription occurs at thousands of sites throughout the mammalian genome. Many of these sites are devoid of protein-coding genes and instead contain long noncoding RNAs (lncRNAs). While lncRNA loci have been implicated in a variety of biological functions, comparatively few lncRNA loci have been genetically defined to have RNA-based roles. Indeed, deletions of entire lncRNA loci have uncovered a number of in vivo phenotypes9–13; however, this approach alone is confounded because in addition to the lncRNA transcript, lncRNA loci can also exert function through DNA regulatory elements14–16, the promoter region17, as well as by the act of transcription18,19. Thus, attributing RNA-based role(s) to lncRNA loci requires testing whether other regulatory modes potentially present at the locus have molecular activity that could contribute to phenotypic effects11,20,21.
In this study, we use the Firre locus as a model to discriminate between DNA- and RNA-mediated effects in vivo. We selected this locus for our study because it is syntenically conserved in a number of mammals including human22–25, and because studies have reported diverse biological and molecular roles. Early characterization of the FIRRE locus in human cell lines identified it as a region that interacts with the X-linked macrosatellite region, DXZ4, in a CTCF-dependent manner26–29. Further analyses of the Firre locus demonstrated that it produces a lncRNA that escapes X-inactivation23,30–32, although it is predominately expressed from the active X chromosome29. Studies using cell culture models suggest that the Firre locus has biological roles in multiple processes, including adipogenesis33, nuclear architecture23,27,29, and regulation of gene expression programs23,34. Additionally, there is some evidence for roles in human development and disease35–38. Collectively, these studies demonstrate the diverse cellular and biological functions for the Firre locus. However, the biological roles of Firre as well as disentangling DNA- and RNA-mediated function(s) for the Firre locus have not been explored in vivo.
Using multiple genetic approaches, we describe an in vivo role for the Firre locus during hematopoiesis. We report that Firre mutant mice have cell-specific defects in hematopoietic populations. Deletion of Firre is accompanied by significant changes in gene expression in a hematopoietic progenitor cell type, which can be rescued by induction of Firre RNA from a transgene within the Firre knockout background. Mice overexpressing Firre have increased levels of pro-inflammatory cytokines and impaired survival upon exposure to lipopolysaccharide (LPS). Finally, the Firre locus does not contain cis-acting RNA or DNA elements (including the promoter) that regulate neighboring gene expression on the X chromosome (9 biological contexts examined), suggesting that Firre does not function in cis. Collectively, our study provides evidence for a trans-acting RNA-based role for the Firre locus that has physiological importance for hematopoiesis and immune function.
RESULTS
The Firre locus produces an abundant lncRNA
We first sought to investigate the gene expression properties for Firre RNA in vivo. To determine potential spatial and temporal aspects of Firre RNA expression during development, we performed in situ hybridization in wildtype (WT) mouse embryos (E8.0 – E12.5). Notably, we detected Firre RNA in many embryonic tissues, including the forebrain, midbrain, pre-somitic mesoderm, lung, forelimb, hindlimb, liver, and heart (Fig. 1A). Since noncoding RNAs have been described to be generally expressed at lower levels compared to protein-coding genes39–42, we determined the relative abundance of Firre RNA in vivo. We performed RNA-seq on eight different WT embryonic tissues and plotted the expression of noncoding and coding transcripts. Consistent with previous reports39–42, we observed that noncoding transcripts were generally less abundant than protein-coding transcripts (Fig. 1B). Despite most lncRNAs being expressed at low levels, we found that Firre, like Malat143–45, is an abundant transcript (Fig. 1B). Next, since Firre is located on the X chromosome and escapes X-inactivation23,30–32, we investigated whether Firre has different expression levels in male and female WT tissues. While levels of Firre RNA varied across embryonic tissue types, within individual tissues, male and female samples exhibited similar expression levels of Firre, despite escaping X inactivation (Fig. 1C).

(A) Whole-mount in situ hybridization for Firre RNA in WT mouse embryos at E8.0 (n=4), E9.5 (n=4), E10.5 (n=5), E11.5 (n=6), and E12.5 (n=4) and ΔFirre E11.5 embryos (n=3). Scale bar is equal to 1 mm. (B) Abundance for protein coding transcripts (light gray) and noncoding transcripts (dark gray) in WT E11.5 heart tissue (representative tissue shown from 7 additional tissues). Vertical lines indicate Firre (red) and Malat1 (blue). (C) Expression of Firre in E11.5 WT male (n=3) and female (n=3) tissues shown as fragments per kilobase of transcript per million mapped reads (FPKM) from RNA-seq. Data shown as mean ± standard error of the mean (SEM). Tissue abbreviations: forebrain (FB), midbrain (MB), pre-somitic mesoderm (PSM), lung (LU), forelimb (FL), hindlimb (HL), liver (LIV), and heart (HRT). (D) Firre knockout mouse (red). Schematic of mouse X chromosome ideogram showing the Firre locus relative to Xist. UCSC genome browser diagram of the Firre locus (shown in opposite orientation). Dashed lines indicate the genomic region that is deleted in ΔFirre mice; single loxP scar upon deletion (gray triangle). Histone modifications and transcription factor binding sites in mouse embryonic stem cells (mESC-Bruce4, ENCODE/LICR, mm9). RNA-seq tracks for the Firre locus in WT and ΔFirre E11.5 forelimbs. (E) Schematic of doxycycline(dox)-inducible Firre overexpression mouse (FirreOE, blue). Tet-responsive element (TRE), minimal CMV promoter (mCMV), reverse-tetracycline transcriptional activator (rtTA), beta-globin polyA terminator (pA). (F) in situ hybridization for Firre at E11.5 in control (WT or tg(Firre) +dox) (n=4) and FirreOE +dox (n=3) embryos. (G) qRT-PCR for Firre expression shown as fold-change (FC) in dox-treated E11.5 control and FirreOE hrt, fb, and fl. Expression normalized to beta-actin in the control sample and data plotted as mean ± confidence interval (CI) at 98%. (H) RNA-FISH for Firre in male WT, ΔFirre, and Firrerescue MEFs. DAPI (blue) marks the nucleus and Firre RNA is shown in green. (I) qRT-PCR for Firre expression shown as FC in male WT, ΔFirre, Firrerescue +dox, and Firrerescue no dox MEFs. Expression normalized to beta-actin in the WT sample and data plotted as mean ± CI at 98%.
Firre knockout and overexpression mice are viable and fertile
To investigate the in vivo role of Firre and assess DNA- and RNA-mediated effects, we generated both Firre loss-of-function and Firre overexpression mice. To delete the Firre locus in vivo, we generated a mouse line containing a floxed allele (Firrefloxed) from a previously targeted mouse embryonic stem cell line23 and matted to a CMV-Cre deleter mouse46. This produced a genomic deletion (81.8 kb) that removed the entire Firre gene body and promoter (henceforth called ΔFirre) (Fig. 1D). We confirmed the deletion of the Firre locus by genotyping (Extended Data Fig. 1) and examined Firre RNA expression. As expected, we did not detect Firre RNA in ΔFirre embryos by whole-mount in situ hybridization or by RNA-seq (Fig. 1A, D).
Since Firre is found on the X chromosome, we first sought to determine if deletion of the locus had an effect on the expected sex ratio of the progeny. Matings between ΔFirre mice produced viable progeny with a normal frequency of male and female pups that did not exhibit overt morphological, skeletal, or weight defects (Extended Data Table 1 and Extended Data Fig. 2). Moreover, deletion of Firre did not impact expression levels of Xist RNA in embryonic tissues or perturb Xist RNA localization during random X chromosome inactivation (XCI) in mouse embryonic fibroblasts (MEFs) (Extended Data Fig. 3A-C).
Genotyping from progeny at P7 from intercrosses between male WT and female WT; male ΔFirre and female ΔFirre; male rtTA and female FirreOE no dox; and male rtTA and female FirreOE dox-diet mice. Litter size shown as mean with standard deviation (s.d.), not determined (n.d.), Chi-square statistic reported (p-value).
Because the ΔFirre allele removes the entire gene body, this model does not allow us to distinguish between DNA- and RNA-mediated effects. Therefore, in order to be able to investigate the role of Firre RNA, we generated a doxycycline (dox)-inducible Firre overexpression mouse. This mouse model was engineered to contain a Firre cDNA downstream of a tet-responsive element (henceforth called tg(Firre)) and was mated to mice that constitutively express the reverse tetracycline transcriptional activator (rtTA) gene (combined alleles henceforth called FirreOE) (Fig. 1E). This approach enabled systemic induction of Firre RNA in a temporally controllable manner by the administration of dox. Moreover, by combining the FirreOE and ΔFirre alleles (henceforth called Firrerescue) we could test whether Firre RNA expression alone is sufficient to rescue phenotypes arising in the ΔFirre mice, thereby distinguishing DNA- and RNA-based effects.
To confirm expression of transgenic Firre RNA, tg(Firre) females were mated with rtTA males and placed on a dox diet the day a copulatory plug was detected, and embryos were collected at E11.5 for analyses. Compared to sibling control embryos, we detected increased Firre RNA in FirreOE embryos by whole-mount in situ hybridization (Fig. 1F) and by quantitative reverse transcription-PCR (qRT-PCR) (heart, 16 fold; forebrain, 26.6 fold; and forelimb, 11.5 fold) (Fig. 1G). Moreover, mice overexpressing Firre are viable and detected at expected male and female frequencies (Extended Data Table 1).
Firre RNA has been reported to be largely enriched in the nucleus of mouse embryonic stem cells (mESCs)23,47, neuronal precursor cells39, and HEK293 cells17, but also has been reported in the cytoplasm of a human colon cell line34. Thus, we investigated the subcellular localization of Firre in the genetic models using RNA fluorescent in situ hybridization (RNA FISH). In contrast to ΔFirre MEFs, we detected pronounced localization of Firre RNA in the nucleus of WT MEFs (Fig. 1H). In dox-treated Firrerescue MEFs, which only produce Firre RNA from the transgene, we detected Firre RNA in both the nucleus and cytoplasm (Fig. 1H), which corresponded to approximately a 2.7-fold increase in Firre RNA relative to WT (Fig. 1I). Notably, the Firrerescue transgenic model showed both nuclear and cytoplasmic localization of Firre, suggesting a threshold level control for nuclear localized Firre.
RNA-seq of ΔFirre embryonic tissues identifies tissue-specific gene dysregulation
Given the broad expression profile of Firre RNA (Fig. 1A), we took an initial unbiased approach to explore the potential biological roles for the Firre locus, and performed poly(A)+ RNA-seq on eight E11.5 tissues from WT and ΔFirre embryos (forebrain, midbrain, heart, lung, liver, forelimb, hindlimb, and pre-somitic mesoderm). As expected, Firre expression was not detected in any of the ΔFirre tissues (Extended Data Tables 2-9). Deletion of Firre was accompanied by significant changes in gene expression in all tissues examined (>1FPKM, FDR<0.05) (Fig. 2A,B and Extended Data Tables 2-9).

(A) Schematized E11.5 tissues used for RNA-seq. WT (n=6) shown in black and ΔFirre (n=6) shown in red. Number of differentially expressed genes shown below each tissue. (B) Heatmap of replicate embryonic tissues. (C) GO analysis for genes found dysregulated in four or more tissues. (D) Firre expression across multiple mouse blood cell lineages (RNA-seq data from bloodspot.eu, GSE60101). (E) Experimental approach for cytokine and survival experiments. (F) Cytokine measurements in serum at 5 hours post intraperitoneal (i.p.) injection of 5 mg/kg LPS (broad-acting) in WT (n=5), ΔFirre (n=5), FirreOE control diet (n=3), and FirreOE dox diet (n=2 to 3). Data are shown as mean ± SEM and significance determined by an unpaired two-tail t-test. (G) Cytokine measurements in serum at 5 hours post i.p. injection of 5mg/kg LPS (specific-acting) in WT (n=6), FirreOE control diet (n=5), FirreOE dox diet (n=4). Data are shown as mean ± SEM and significance determined by an unpaired two-tail t-test. (H) 6-day survival plot of mice injected with 5 mg/kg LPS (specific-acting) or saline over two independent experiments in WT (n=30), ΔFirre (n=18), FirreOE control diet (n=13), and FirreOE dox diet (n=17). Saline control group (n=10) consisting of WT, ΔFirre, and FirreOE mice. Significance determined by Mantel-Cox test.
Across these eight tissues, we identified a total of 3,910 significantly differentially expressed genes, of which 271 genes were differentially expressed in two or more tissues (Extended Data Tables 2-9). Interestingly, gene ontology (GO) analysis of the commonly dysregulated genes showed that deletion of the Firre locus affected genes involved in hemoglobin regulation and general blood developmental processes (Fig. 2C). We therefore analyzed publicly available mouse RNA-seq datasets and found that Firre is expressed across many blood cell types and note that expression is found highest in hematopoietic stem cells (HSCs)48 and then decreases in conjunction with hematopoietic differentiation49 (Fig. 2D). Based on this information we narrowed our investigation to evaluate potential roles for Firre in the blood system, and leveraged the genetic mouse models to test DNA- and RNA-mediated effects.
LPS exposure to mice overexpressing Firre RNA impacts the innate immune response
Firre is expressed in many innate immune cell types (Fig. 2D) and has been shown to regulate the levels of inflammatory genes in human intestinal epithelial and mouse macrophage cell lines34. Thus, we hypothesized that dysregulation of Firre might alter the inflammatory response in vivo. To test this, we employed a commonly used endotoxic shock model by administering lipopolysaccharide (LPS) intraperitoneally to cohorts of WT, ΔFirre, FirreOE no dox, and dox-fed FirreOE mice in order to stimulate signaling pathways that regulate inflammatory mediators50 (Fig. 3E).

(A) Schematic of hematopoiesis. (B) Frequencies of CD4, CD8, and NK cells from the peripheral blood from WT (black circle) and ΔFirre (red square) mice. Three representative experiments combined (seven independent experiments). Frequencies of common myeloid progenitors (CMP) and common lymphoid progenitors (CLP) in the bone marrow shown from WT and ΔFirre mice. Two representative experiments combined (three independent experiments). (C) Frequencies of CMPs and CLPs from the bone marrow from control (tg(Firre) or WT or rtTA with dox) (black circle) and dox-treated FirreOE (blue square) mice. One representative experiment shown (two independent experiments). Frequencies of CD4, CD8, and NK cells from the peripheral blood from control (WT or tg(Firre) or rtTA with dox) (black circle) and dox-treated FirreOE (blue square) mice. One representative experiment shown (three independent experiments). All cell frequencies determined by flow cytometry analysis. All data are plotted as percent (%) of live cells showing the mean ± SEM and statistical significance determined by a two-tailed Mann-Whitney U test.
We administered two different LPS preparations, one which broadly stimulates the pattern recognition receptors toll-like receptors (TLR) 2, 4 and nitric oxide synthase, and an ultrapure LPS preparation that specifically stimulates TLR451–53. At 5 hours post LPS injection we measured serum cytokine levels. Notably, we observed that FirreOE dox-fed mice administered broad-acting LPS had significantly higher levels of inflammatory cytokines, including TNFα, IL12-p40, and MIP-2 compared to WT (Fig. 2F). In contrast, we did not observe a significant difference for these cytokines in LPS-treated ΔFirre mice (Fig. 2F). Consistent with the increased cytokine response using broad-acting LPS, dox-fed FirreOE mice administered TLR4 specific-acting LPS also had significantly higher levels of TNFα, IL12-p40, and MIP-2 compared to WT (Fig. 2G), albeit at lower serum concentrations compared to the broad-acting LPS (Fig. 2F,G). In addition, we confirmed that overexpressing Firre RNA alone (without LPS) does not result in increased serum levels of TNFα, IL12-p40, and MIP-2 (Extended Data Fig. 4).
Because increased levels of TNFα is a hallmark of endotoxic shock54–56, we next tested whether the levels of Firre RNA had an impact on survival following LPS treatment. We administered 5mg/kg of TLR4 specific-acting LPS to WT (n=30), ΔFirre (n=18), FirreOE no dox (n=13), and FirreOE dox-fed (n=17) mice, as well as a saline control group and monitored for 6 days. At this dose, across two independent cohorts, dox-treated FirreOE mice showed a significantly higher susceptibility to LPS compared to WT mice (P<0.0001, Mantel-Cox) and uninduced FirreOE animals (P=0.0063, Mantel-Cox) (Fig. 2H). Whereas ΔFirre mice did not show a significant difference in the level of susceptibility to LPS (P=0.1967, Mantel-Cox test) (Fig. 2H). Collectively, these results indicate that increased levels of Firre RNA can modulate the inflammatory response in vivo independent of genomic context, suggesting an RNA-based role for Firre.
Modulating the Firre locus and RNA results in cell-specific defects during hematopoiesis
Having observed an effect of Firre in regulating gene expression and accentuating the inflammatory response, we further investigated the role of Firre in hematopoiesis (Fig. 3A). We first examined cell populations in the peripheral blood in ΔFirre mice and observed a modest but significant reduction in the frequencies of CD4 and CD8 T cells (P=0.0002 and P=0.0081, respectively), whereas the frequencies of B and NK cells were unaffected compared to WT (Fig. 3B and Extended Data Fig. 5A). To investigate the cause of this reduction we examined the thymus (to assess for a defect in T cell development) and the bone marrow (to assess for a defect in hematopoietic progenitor cells). There was no block in thymic development in ΔFirre mice, as normal frequencies of cells were observed at each developmental stage (Extended Data Fig. 5B, upper panels). However, we noticed that the absolute number of cells was generally lower in ΔFirre mice at every developmental stage, suggestive of a pre-thymic defect in progenitor development (Extended Data Fig. 5B, lower panels). Consistent with this, in the bone marrow compartment, we observed a significant reduction in both the frequency and number of the common lymphoid progenitors (CLPs) (lineage(lin)-Sca-1lo-c-KitloIL7Rα+), a hematopoietic progenitor cell type, in ΔFirre mice (P=0.0069 and P<0.0001, respectively) (Fig. 3B and Extended Data Fig. 5C).
To assess whether the observed defect in hematopoiesis could be due to a progenitor-intrinsic effect of Firre deficiency, we performed competitive chimera assays. Briefly, we isolated an HSC-enriched population (lin-Sca-1+c-Kit+CD34+/-CD135-) from WT (CD45.2) and ΔFirre (CD45.2), mixed the cells separately at an equal ratio with congenic WT (CD45.1) HSCs, and transplanted this mixture into lethally irradiated CD45.1 recipient mice (Extended Data Fig. 6A,B). We assessed the long-term reconstitution ability of WT and ΔFirre HSCs to repopulate blood cell lineages in vivo. We observed that ΔFirre/CD45.2-donors had reduced frequencies of CD4 and CD8 T-cells (n=10, P=0.0028 and P=0.0051), B-cell (n=10, P=0.0114), and NK cell (n=10, 0.0068) populations in the peripheral blood of recipient mice compared to WT/CD45.2-donors, suggesting that ΔFirre-donors were markedly outcompeted at repopulating the blood (Extended Data Fig. 6C). These data are consistent with a progenitor-intrinsic role for Firre in hematopoiesis.
In contrast to the ΔFirre model, mice overexpressing Firre RNA in the WT background (FirreOE), had normal frequencies of CD4, CD8 and B cells, but had a significant reduction in the frequency of NK cells (P=0.0043) in the peripheral blood compared to control mice (Fig. 3C, Extended Data Fig. 5D). A decrease in the frequency of NK cells in dox-fed Firrerescue mice, where only Firre RNA from the transgene is expressed, was also observed (Extended Data Fig. 7). In the bone marrow of dox-treated FirreOE mice, we did not observe significant changes in the frequencies of HSC, multipotent progenitor (MPP), common myeloid progenitor (CMP), or CLPs compared to control samples (Fig. 3C and Extended Data Fig. 5E). Taken together, these results identify cell type-specific defects during hematopoiesis, whereby alterations of Firre impact the ratios and numbers of particular blood cells produced during hematopoiesis.
Firre lncRNA has a trans-acting role in vivo
Next, we wanted to further investigate the DNA- and RNA-mediated effects of the Firre locus using a cell type that was dysregulated in the ΔFirre immunophenotyping analysis. We selected to use the CLP as a model because this was the earliest hematopoietic defect identified and because Firre is highly expressed in this progenitor cell type. Therefore, further investigation could provide insight into the physiological effects of modulating Firre in a progenitor cell population. Because the ΔFirre mouse contains a deletion that removes all potential DNA-regulatory elements, the lncRNA, and the promoter (thus removing the act of transcription), this mouse model does not allow us to distinguish between DNA- and RNA-mediated effects.
To directly test whether the hematopoietic CLP defect in the ΔFirre mice is mediated by an RNA-based mechanism, we reasoned that overexpressing Firre RNA in the ΔFirre background would enable us to identify RNA-mediated effects. To this end, we generated multiple cohorts of compound mice (Firrerescue) that contained the FirreOE alleles in the ΔFirre background and induced transgenic Firre expression by placing mice on a dox-diet. From multiple cohorts of WT, ΔFirre, and dox-fed Firrerescue mice, we assessed CLP frequency by flow cytometry in total bone marrow and lineage depleted bone marrow (to enrich for hematopoietic non-lineage committed cells) (Fig. 4A).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
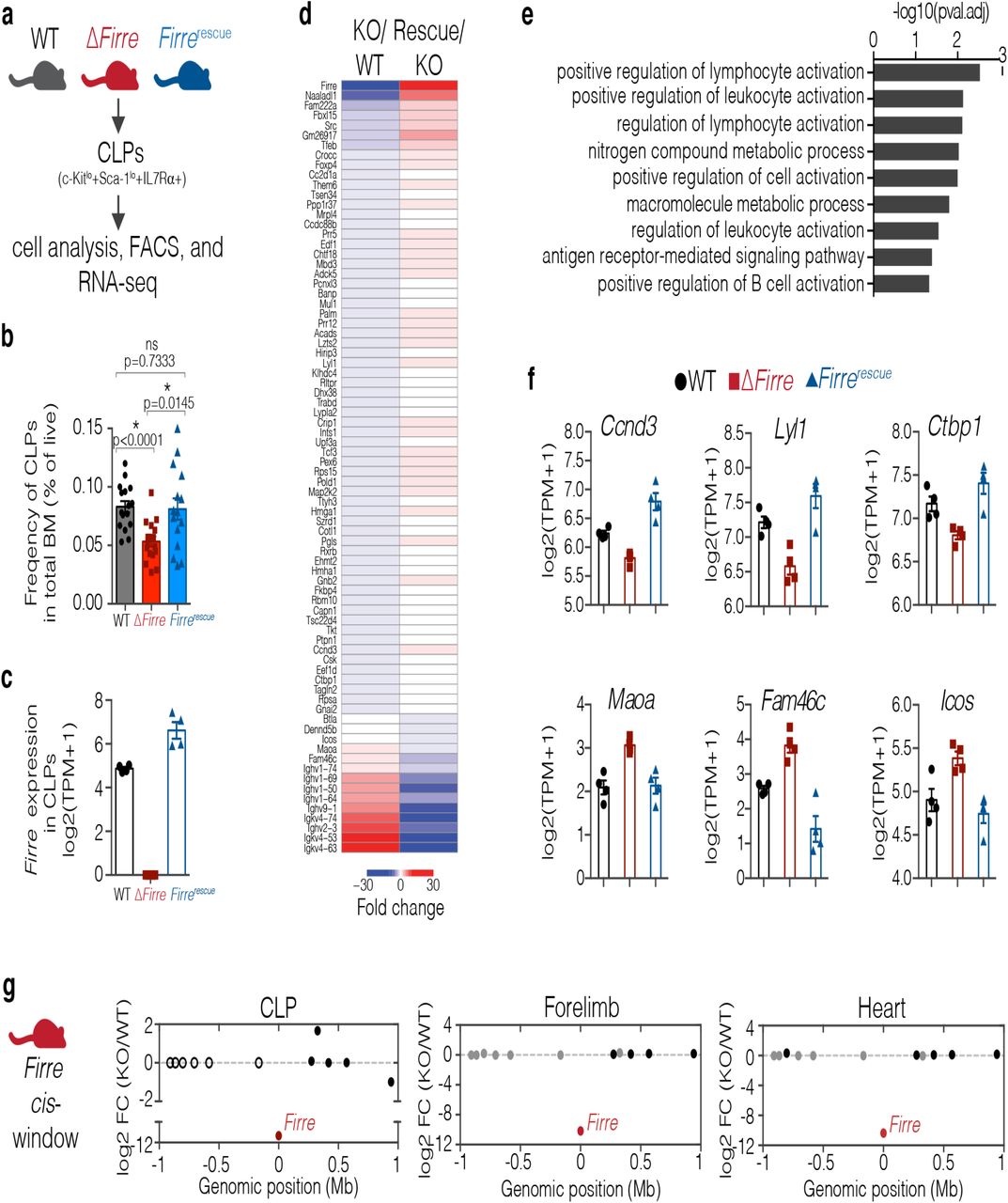
(A) Schematic of experimental approach. (B) Bar graph indicating the frequency of CLPs shown as percent of live in total bone marrow from 3-7 month old WT (n=16, mean age=26 weeks), ΔFirre (n=17, mean age=23 weeks), and Firrerescue dox diet (n=15, mean age=23 weeks) mice over three independent experiments. Data are shown as mean ± SEM and statistical significance determined by a two-tailed Mann-Whitney U test. (C) Firre RNA expression in CLPs from WT (n=4), ΔFirre (n=4), and dox-treated Firrerescue (n=4) determined by RNA-seq. Data plotted as transcripts per million (TPM +1) showing the mean ± SEM. (D) Heatmap showing significantly differentially expressed genes in CLPs in ΔFirre / WT comparison and dox-treated Firrerescue / ΔFirre and comparison. (E) GO analysis for significantly dysregulated genes in ΔFirre CLPs. (F) Examples of genes that show significant reciprocal regulation in WT, ΔFirre, and dox-treated Firrerescue CLPs. (G) Firre locus region (2 Mb) showing gene expression differences in log2 FC between ΔFirre and WT CLPs, mouse embryonic forelimb, and heart. Firre is shown in red, significantly dysregulated genes are shown in red, genes that are not significantly changed are shown in black, and genes that were not detected shown in white.
Consistent with our previous data (Fig. 2B), we observed a significant decrease in the frequency of CLPs in total bone marrow from ΔFirre mice (n=17, mean CLP frequency = 0.0532) compared to WT mice (n=16, mean CLP frequency = 0.0829) (P<0.0001) (Fig. 4B). Further, in separate experimental cohorts, the frequency of CLPs in lineage-depleted bone marrow from ΔFirre mice was significantly decreased (n=9, mean CLP frequency = 0.3011, P=0.0071) compared to WT (n=9, mean CLP frequency = 0.2167) (Extended Data Fig. 8). Notably, the frequency of CLPs was significantly increased in dox-fed Firrerescue mice compared to ΔFirre mice and restored to approximately that of WT in both total bone marrow (n=15, mean CLP frequency = 0.0810, P=0.0145) (Fig. 4B) and lineage depleted bone marrow (n=11, mean CLP frequency = 0.2809, P=0.0234, respectively) (Extended Data Fig. 8). Thus, induction of transgenic Firre RNA alone is sufficient to rescue the reduction in frequency of CLPs observed in ΔFirre bone marrow. These data suggest that Firre RNA, rather than the DNA, exerts a biological function in the CLPs during hematopoiesis.
Expression of transgenic Firre restores gene expression programs in vivo
To gain further insight into the molecular roles of Firre in the CLPs, we took a gene expression approach because alterations of the Firre locus and RNA have previously been shown to impact gene expression23,29,57. Moreover, we reasoned that we could test if changes in gene expression in the loss-of-function model could be rescued by expressing only Firre RNA. To this end, we isolated CLPs by fluorescence activated cell sorting (FACS) from the bone marrow of age- and sex-matched WT, ΔFirre, and dox-fed Firrerescue mice and performed poly(A)+ RNA-seq. As expected, Firre RNA was not detected in the ΔFirre samples, and Firre RNA levels were restored to levels above WT in the Firrerescue samples (Fig. 4C). Differential gene expression analysis between WT and ΔFirre CLPs identified 89 significantly differentially expressed genes (FDR<0.1) (Fig. 4D and Extended Data Table 10). GO analysis of the differentially expressed genes showed that deletion of Firre in CLPs affected genes involved in lymphocyte activation, cell adhesion, and B cell activation (Fig. 4E).
Next, we determined if induction of Firre RNA in the Firrerescue model could rescue expression of the 89 significantly dysregulated genes found in ΔFirre CLPs. We compared the CLP RNA-seq from ΔFirre and Firrerescue mice and identified 4,656 genes with significant changes in gene expression (FDR<0.1) (Extended Data Table 11). Notably, 78 of the 89 genes that were significantly differentially expressed in ΔFirre CLPs, were found to be significantly and reciprocally regulated in Firrerescue CLPs (P=2.2e-16, Fisher exact test) (Fig. 4D). For example, Ccnd3, Lyl1, and Ctbp1 are significantly downregulated in ΔFirre CLPs, but are found significantly upregulated in Firrerescue CLPs to (Fig. 4F). Further, genes such as Maoa, Fam46c, and Icos were found significantly upregulated in ΔFirre CLPs, but their expression was significantly reduced in Firrerescue CLPs (Fig. 4F). We also noted that several immunoglobin heavy and light chain variable region genes were reciprocally regulated in our analyses (Fig. 4D). Taken together, these data suggest that ectopic expression of Firre is sufficient to restore a gene expression program in an RNA-based manner in vivo.
Firre does not function in cis
Many lncRNA loci exert function to control the expression of neighboring genes, a biological function called cis regulation58. This occurs through a variety of mechanisms including, cis-acting DNA regulatory elements, the promoter region, the act of transcription, and the lncRNA (a biological function called cis regulation)18,59–62. The ΔFirre mouse model enables to test for a potential cis regulatory roles for Firre on the X chromosome because the knockout removes the entire Firre locus (Fig. 1D). To investigate local (cis) effects on gene expression, we generated a 2 Mb windows centered on the Firre locus and examined whether the neighboring genes were significantly dysregulated across 9 biological contexts.
Differential gene expression analysis for WT and ΔFirre CLPs showed that of the 12 genes within a 2 Mb window (excluding Firre), none were differentially expressed (Fig. 4G). Consistent with this finding, we did not observe significant changes in gene expression (2 Mb windows centered on the Firre locus) in seven of the eight embryonic tissues (Fig. 4G and Extended Data Fig. 9A-F). Indeed, we observed one instance of differential expression in one embryonic tissue (Hs6st2 was slightly but significantly downregulated in the embryonic forebrain, −0.38 log2 fold change, FDR<0.05) (Extended Data Fig. 9A-F). These data demonstrate that the Firre locus does not exert a local effect on gene expression in vivo, and suggest that the Firre lncRNA regulates gene expression in a trans-based manner. Collectively, our study investigates the roles of DNA and RNA at the Firre locus in vivo and genetically defines that the Firre locus produces as a trans-acting lncRNA molecule in a hematopoetic context.
DISCUSSION
Classic models used to study noncoding RNAs – ribosomal RNAs, small nucleolar RNAs, tRNAs, and telomerase RNA component (TERC) – have demonstrated that these RNAs species serve important cellular functions. This core of possible RNA biology has been greatly expanded by studies that have identified tens-of-thousands of lncRNAs39,40,63. Indeed, subsequent molecular and genetic interrogation of lncRNA loci have identified diverse molecular roles and biological phenotypes. However, lncRNA loci potentially contain multiple modes that can exert function, including DNA regulatory elements (including the promoter), the act of transcription, and the lncRNA. Therefore, attributing an RNA-based role to a lncRNA locus requires the development of multiple genetic models to determine the activities and contributions of potential regulatory modalities20,21,59.
In this study, we developed three genetic models in mice for the syntenically conserved lncRNA Firre: loss-of-function, overexpression, and rescue. Notably, we report that deletion of the Firre locus does not impact survival in mice, or despite escaping XCI, skew the sex ratio of progeny. We leveraged the genetic models to discriminate between DNA- and RNA-mediated effects in vivo. We determined that modulating Firre directs cell-specific defects during hematopoiesis, potentiates the innate immune response upon exposure to LPS, and can restore gene expression programs – all of which have an RNA-based functional modality. We also conclude that the Firre locus does not have a local cis-regulatory effect on gene-expression across numerous tissues. Together, by using multiple genetic and molecular approaches we identified that Firre produces a trans-acting lncRNA in a hematopoietic context. There are several important implications for these results.
First, our study indicates that Firre has trans RNA-based activity in vivo, and thus extends previous reports that have suggested RNA-based roles for Firre in cell culture models34,47. By using compound genetic approaches, we found that overexpression of Firre from a transgene in the Firre-deficient background was sufficient to rescue physiological and molecular phenotypes in ΔFirre CLPs in vivo. We speculate that early hematopoietic progenitor cells may represent a unique context to study the role(s) of FIRRE/Firre. In humans, FIRRE is expressed as both circular (circ-FIRRE) and linear forms in hematopoietic cells, and circ-FIRRE is abundant in all progenitor cell types except for the CLPs64. More studies will be needed to determine the functional differences between linear and circular isoforms of Firre in vivo.
Second, we observed that overexpression of Firre RNA in an endotoxic shock model potentiated the innate immune response in vivo. Our findings suggest that ectopic or high levels Firre lncRNA could be important for regulating the innate immune response. Consistent with our findings, modulating the levels of human and mouse FIRRE/Firre RNA in human intestinal epithelial and mouse macrophage cell lines perturbed the mRNA levels of inflammatory genes, including IL12-p4034. Interestingly, we identified increased levels of IL12-p40 protein in the serum from FirreOE mice upon exposure to LPS. We speculate that Firre could be important in setting functional thresholds for cells. For example, FIRRE is not only significantly increased in certain cancers38, but high levels of FIRRE expression have been significantly associated with more aggressive disease and poor survival in patients with large B-cell lymphoma36.
Finally, our study suggests that Firre does not have a cis-regulatory role on gene expression in vivo. Upon deletion of the Firre locus and its promoter region we observed global changes in gene expression. Yet, we did not find changes in local gene expression (2 Mb window) at the Firre locus in eight embryonic tissues and in CLPs. Thus, potential DNA-regulatory elements, the lncRNA, the promoter, and the act of transcription appear to not have regulatory roles on neighboring gene expression in the nine biological contexts (8 embryonic and 1 cell type) analyzed in this study. Moreover, we observed that deletion of Firre in vivo does not perturb Xist RNA expression in eight embryonic tissues and does not affect random XCI in MEFs, consistent with a previous study using cell culture models29. Together, these data are notable because cis-acting mechanisms are speculated to be common feature at lncRNA loci65. While we did not find any evidence for cis-activity at the Firre locus in vivo, a previous study from our group found active DNA elements within the Firre locus using a cell-based enhancer reporter assay in 3T3 cells22. We speculate that these candidate DNA regulatory elements are likely to regulate the Firre locus rather than neighboring genes, as we did not find evidence of dysregulation in gene expression for neighboring genes when the locus was deleted in vivo.
In summary, we have examined the role of Firre in the context of hematopoiesis in order to test DNA- and RNA-mediated effects. This study does not exclude that Firre could be functioning elsewhere, and even by other molecular modalities. Indeed, we identified that Firre is abundantly expressed in a number of tissues, therefore going forward it will be important to investigate the potential role(s) of Firre in other biological contexts as well as in pathological disease. Our findings provide evidence that the X chromosome lncRNA locus Firre has a role in hematopoiesis that is mediated by a trans-acting RNA, and further highlights the biological importance of lncRNA-based machines in vivo.
METHODS
Mouse care and ethics statement
Mice used in these studies were maintained in a pathogen-specific free facility under the care and supervision of Harvard University’s Institutional Animal Care Committee.
Mouse strains and genotyping
Firrefloxed mice were generated from 129/C57 F1 hybrid mouse embryonic cells as previously described23. Briefly, sequential targeting was used to insert a floxed-neomycin-floxed cassette in the 5’ end of the Firre locus between nucleotides 4790843-4790844 (mm9) and a floxed-hygromycin-floxed cassette was inserted into the 3’ end of the Firre locus between nucleotides 47990293-47990294 (mm9) (Extended Data Fig. 1A). 129/C57 F1 hybrid cells containing the Firrefloxed allele were injected into 129/C57 blastocysts (Harvard Genome Modification Facility). Transgenic mice were screened for the Firrefloxed allele by PCR genotyping. To generate a deletion of the Firre locus, female Firrefloxed mice were matted to a male B6.C-Tg(CMV-Cre)1Cgn/J mouse46 (Jackson Lab, 006054). Tail biopsies were collected from the progeny and were genotyped for WT, knockout, neomycin, hygromycin, and cre alleles (Extended Data Fig. 1B). Female mice heterozygous for the Firre deletion were subsequently mated to C57BL/6J mice. Firre WT and ΔFirre mice used in this study are from the F3 generation.
To generate an inducible Firre-overexpressing allele in mice (tg(Firre)), we cloned a Firre cDNA into a Tet-On vector (pTRE2) where the beta globin intron sequence was removed. We next used both EcoRI and NheI restriction enzymes to digest the cassette containing the tet-responsive element, CMV minimal promoter, Firre cDNA, and beta globin poly(A) terminator. This cassette was injected into the pronucleus of C57BL/6J zygotes (Harvard Genome Modification Facility). Male founder mice containing the tg(Firre) cassette were identified and individually mated to female C57BL/6J mice (Jackson Laboratory, 000664). To overexpress tg(Firre) F2 and F3 generation females were mated to male B6N.FVB(Cg)-Tg(CAG-rtTA3)4288Slowe/J (rtTA) mice (Jackson Laboratory, 016532) and at the plug date females were either put on a normal diet or 625 mg/kg doxycycline-containing food (Envigo, TD.01306) until experimental end points. A colony of male rtTA mice were maintained by breeding to C57BL6/J females for up to 4 generations.
Genotyping for mice was performed on tissue collected at P7. Primers used for genotyping: Firre wild-type allele, F-GGAGGAGTGCTGCTTACTGG, R-TCTGTGAGCCACCTGAAATG; ΔFirre allele, F-TCACAATGGGCTGGGTATTCTC, R- CCTGGGTCCTCTATAAAAGCAACAG; neomycin, F- GACCACCAAGCGAAACATC, R- CTCGTCAAGAAGGCGATAGAA; hygromycin, F-CGGAAGTGCTTGACATTGGG, R- CGTCCATCACAGTTTGCCAGTG; Cre, F- TAATCCATATTGGCAGAACG, R- ATCAATCGATGAGTTGCTTC; Sry, F- TTGTCTAGAGAGCATGGAGGGCCATGTCAA, R- CCACTCCTCTGTGACACTTTAGCCCTCCGA; tg(Firre) allele, F: TACCACTCCCTATCAGTGA, R: CGGCTTCATCTTCAGTCCTC; and the rtTA allele, F: AGTCACTTGTCACACAACG, R: CTCTTATGGAGATCCCTCGAC. Additional genotyping was performed by Transnetyx using real-time PCR.
Cytokine analysis and in vivo endotoxin challenge
To investigate the cytokine response in vivo, we used two different preparations of LPS from Escherichia coli (E. coli) O111:B4. (Sigma, L2630) and Ultrapure LPS, E. coli O111:B4 (InvivoGen, tlrl-3pelps) and dissolved in 0.9% saline solution (Teknova, S5825). We administered either 0.9% saline or 5 mg/kg broad-acting LPS (Sigma, L2630) by i.p. injection using a 30G needle (BD insulin syringes, 328411) to mice cohorts 8 to 10 weeks old (WT, ΔFirre, FirreOE no dox, and dox fed FirreOE). We also administered either a 0.9% saline or a 5 mg/kg dose of LPS that is TLR4-specific (InvivoGen, tlrl-3pelps) by i.p. injection in mice 5 to 10 weeks old (WT, FirreOE no dox, and dox fed FirreOE). At 5 hours post i.p. injection, mice were euthanized and peripheral blood was collected by cardiac puncture and allowed to clot for 30 minutes at room temperature with gentle rotation. After clotting, samples were centrifuged at 1000 x g for 10 minutes at 4°C and serum was collected. Cytokine analysis was performed on serum diluted 2-fold in PBS pH7.4 (Eve Technologies, Chemokine Array 31-Plex). Measurements within the linear range of the assay are reported.
Endotoxin survival experiments were performed over two independent experiments using mice 9 to 16 weeks old: WT (mean age = 12.9 weeks), ΔFirre (mean age = 16 weeks), FirreOE no dox (mean age = 13.6 weeks), and dox fed FirreOE (mean age = 13.7 weeks). Saline control group consisting of WT, ΔFirre, and FirreOE mice. 0.9% saline or 5 mg/kg LPS (InvivoGen, tlrl-3pelps) was prepared as described above and administered by i.p. injection and mice were monitored for moribund survival over 6 days. Mice were housed at a density of 3 to 5 mice per cage containing: Anderson’s Bed (The Andersons, Inc), Enviro-Dri (Shepherd Specialty Papers), compressed 2” x 2” cotton nestlet (Ancare), and a mouse hut (BioServ). The following supportive care was provided during the duration of the experiments: hydrogel, a small cup containing powdered diet mixed with water, and a heating pad (5” × 8.6” × 6”) was placed externally on the bottom of the cage (Kobayashi).
Whole-mount in situ hybridization
We generated an antisense digoxigenin-labeled antisense riboprobe against Firre from a 428bp sequence (Extended Data Fig. 1C) corresponding to the 5’ end of the Firre transcript. In situ hybridization was performed on a minimum of three embryos per stage and/or genotype. For whole-mount staining, we fixed embryos in 4% paraformaldehyde for 18 hours at 4°C, followed by 3 washes in 1× PBS for 10 minutes at room temperature. We then dehydrated the embryos them for 5 min at room temperature in a series of graded methanol solutions (25%, 50%, 75%, methanol containing 0.85% NaCl, and 100% methanol). Embryos were then stored in 100% methanol at −20°C. We then rehydrated embryos through a graded series of 75%, 50%, 25%, methanol/ 0.85% NaCl 5 min incubations at room temperature and then washed in twice in 1× PBS with 0.1% Tween-20 (PBST). Embryos were treated with 10mg/mL proteinase K in 1× PBST for 10 minutes (E8.0, E9.5) or 30 minutes (E10.5, E11.5 and E12.5). Samples were fixed again in 4% paraformaldehyde/0.2% glutaraldehyde in PBST for 20 minutes at room temperature and washed in twice in 1× PBST. We then incubated samples in pre-hybridization solution for 1 hour at 68°C and then incubated samples in 500 ng/mL of Firre antisense riboprobe at 68°C for 16 hours. Post hybridization, samples were washed in stringency washes and incubated in 100 μg/mL RNaseA at 37°C for 566 1 hour. Samples were washed in 1X maleic acid buffer with 0.1% Tween-20 (MBST) and then incubated in Roche Blocking Reagent (Roche, 1096176) with 10% heat inactivated sheep serum (Sigma, S2263) for 4 hours at room temperature. We used an anti-digoxigenin antibody (Roche, 11093274910) at 1:5000 and incubated the samples for 18 hours at 4°C. Samples were washed 8 times with MBST for 15 min, 5 times in MBST for 1 hour, and then once in MBST for 16 hours at 4°C. To develop, samples were washed 3× for 5 min at room temperature with NTMT solution (100 mM NaCl, 100 mM Tris-HCl (pH 9.5), 50 mM MgCl2, 0.1% Tween-20, 2 mM levamisole). The in situ hybridization signal was developed by adding BM Purple (Roche, 11442074001) for 4, 6, 8, and 12 hours. After the colorimetric development, samples were fixed in 4% paraformaldehyde and cleared through a graded series of glycerol/1× PBS and stored in 80% glycerol. Embryos were imaged on a Leica M216FA stereomicroscope (Leica Microsystems) equipped with a DFC300 FX digital imaging camera.
RNA-seq in embryonic tissues preparation and analysis
For WT and ΔFirre RNA-seq in embryonic tissues we dissected tissues (forebrain, midbrain, heart, lung, liver, forelimb, hindlimb, and presomitic mesoderm) from E11.5 embryos (44-48 somites) that were collected from matings between either male WT and female Firre+/- or male ΔFirre and female Firre+/- mice. Tissues were immediately homogenized in Trizol (Invitrogen) and total RNA was isolated using RNeasy mini columns (Qiagen) on a QIAcube (Qiagen). Samples were genotyped for the WT, ΔFirre, and sex alleles (Extended Data Fig. 1A,B). For each tissue, we generated the following libraries: WT male (n=3), WT female (n=3), ΔFirre male (n=3), and ΔFirre female (n=3). Poly(A)+ RNA-seq libraries were constructed using TruSeq RNA Sample Preparation Kit v2 (Illumina). The libraries were prepared using 500ng of total RNA as input, with the exception of the lung (200ng) and the presomitic mesoderm (80ng), and with a 10-cycle PCR enrichment to minimize PCR artifacts. The indexed libraries were pooled in groups of six, with each pool containing a mix of WT and ΔFirre samples. Pooled libraries were sequenced on an Illumina HiSeq 2500 in rapid-run mode with paired-end reads.
Reads were mapped to the mm10 mouse reference genome using TopHat v2.1.1 with the flags: “--no-coverage-search --GTF gencode.vM9.annotation.gtf” where this GTF is the Gencode vM9 reference gene annotation available at gencodegenes.org. Cufflinks v2.2.1 was used to quantify gene expression and assess the statistical significance of differences between conditions. Cuffdiff was used to independently compare the WT and ΔFirre samples and from each tissue and sex, and genes with FDR<0.05 were deemed significant (Extended Data Tables 2-9).
RNA-seq in CLPs preparation and analysis
We isolated CLPs (Lin-Sca-1locKitloIL7Rα+) by fluorescence activated cell sorting (FACS) from mice 27 to 32 weeks old: WT (n=4, mean age = 31 weeks), ΔFirre (n=4, mean age = 30.4 weeks), and dox fed Firrerescue (n=4, mean age = 29.3 weeks. CLPs were directly sorted into TRIzol. RNA was isolated using RNeasy micro columns (Qiagen) on a QIAcube (Qiagen) and we quantification the concentration and determined the RNA integrity using a BioAnalyzer (Agilent). Poly(A)+ RNA-seq libraries were constructed using CATS RNA-seq kit v2 (Diagenode, C05010041). Pooled libraries were sequenced on an Illumina HiSeq 2500 in rapid-run mode with paired-end reads.
The adapter-trimmed reads were mapped to the mm10 mouse reference genome using TopHat v2.1.1 with the flags: “--no-coverage-search --GTF gencode.vM9.annotation.gtf” (Gencode vM9 reference gene annotation) FeatureCounts and R-package, DESeq2, were used to quantify gene expression and assess the statistical significance of differences between conditions66,67 and the p-value of comparisons were empirically calculated by using fdrtools68. Genes with an FDR<0.1 were deemed significant in a comparison between wildtype and ΔFirre (Extended Data Table 10) and genes with an FDR<0.1 in the ΔFirre and Firrerescue comparison were deemed significant (Extended Data Table 11).
qRT-PCR
Embryonic tissues were homogenized in Trizol (Invitrogen) and total RNA was isolated using RNeasy mini columns (Qiagen) on a QIAcube (Qiagen). 300ng of total RNA was used as input to synthesize cDNA (SuperScript IV VILO Master Mix, Invitrogen, 11756050). Primers used qRT-PCR experiments: F_b-act: GCTGTATTCCCCTCCATCGTG, R_b-act: CACGGTTGGCCTTAGGGTTCAG; F_Firre: AAATCCGAGGACAGTCGAGC, R_Firre: CCGTGGCTGGTGACTTTTTG. Experiments were performed on a Viia7 (Applied Biosciences). qRT-PCR data was analyzed by the ΔΔCt method.
Distribution of Firre expression across wild-type tissues
For each of the eight WT embryonic tissues, FPKM estimates of all protein coding or noncoding genes were aggregated and filtered for expression > 1FPKM. Density plots were generated using ggplot2 (geom_density()). The distributions of wildtype FPKM estimates for these transcriptional types are indicated by black line for protein-coding and gray line for non-coding expression.
Flow Cytometry analysis
Age and sex-matched adult mice were used in all flow cytometry experiments. Zombie Aqua Fixable Viability Kit (Biolegend, 423101) was used as a live-dead stain. CountBright Absolute Counting Beads (Invitrogen, C36950) were added to bone marrow and thymi samples in order to enumerate cell populations.
For cell analysis, peripheral blood was collected by cardiac puncture. The following antibodies were added (1:100) to each sample and incubated for 30 minutes at room temperature Alexa Fluor 700 anti-mouse CD8a (Biolegend, 100730), PE/Dazzle-594 anti-mouse CD4 (Biolegend, 100456), APC anti-mouse CD19 (Biolegend, 115512), Alexa Fluor 488 anti-mouse NK-1.1 (Biolegend, 108718), PE anti-mouse CD3 (Biolegend, 100205), PE/Cy7 anti-mouse/human CD44 (Biolegend, 103030), eFluor 450 anti-Mouse CD62L (L-Selectin, eBiosciences, 48-0621-82), and TruStain FcX (anti-mouse CD16/32) antibody (1:50) (Biolegend, 101319). Red blood cells were then lysed for 15 minutes at room temperature using BD FACS Lysing Solution (BD, 349202). Cells were washed twice in 1× PBS with 1% BSA and then resuspended in 1% paraformaldehyde or 1× PBS with 0.2% BSA.
Thymi were collected and homogenized in ice cold PBS over a 40 micron filter. The cells were incubated with the following antibodies (1:100) for 30 minutes at room temperature: Alexa Fluor 488 anti-mouse CD25 (Biolegend, 102017), PE/Cy7 anti-mouse/human CD44 (Biolegend, 103030), PE anti-mouse TCR β chain (Biolegend, 109208), APC anti-mouse/human CD45R/B220 (Biolegend, 103212), eFluor 450 anti-Mouse CD69 (eBiosciences, 48-069182), Alexa Fluor 700 anti-mouse CD8a (Biolegend, 100730), and PE/Dazzle-594 anti-mouse CD4 (Biolegend, 100456). Cells were washed twice in 1x PBS with 1% BSA and then resuspended in 1% paraformaldehyde.
Bone marrow was collected from both femurs and tibias (four bones total per mouse) by removing the end caps and flushing with DMEM (Gibco, 11995-073) containing 5% FBS (Gibco, 26140079) and 10mM EDTA. Cells were then pelleted, re-suspended and passed through a 70 micron filter. The resulting single cell suspension was then incubated with the following antibodies (1:100) for 60 minutes on ice: Alexa Fluor 700 anti-Mouse CD16/CD32 (eBiosciences, 65016182), PE/Cy7 anti-mouse CD127 (IL-7Rα) (Biolegend, 135014), Alexa Fluor 488 anti-mouse CD117 (c-Kit) (Biolegend, 105816), PE/Dazzle-594 anti-mouse Ly-6A/E (Sca-1) (Biolegend, 108138), Pacific Blue anti-mouse Lineage Cocktail (Biolegend, 135306), PE anti-mouse CD135 (Biolegend, 135306), and APC anti-mouse CD34 (Biolegend, 128612). Red blood cells were then lysed for 15 minutes at room temperature using BD FACS Lysing Solution (BD, 349202) or BD Pharm Lyse (BD, 555899). Cells were washed twice in 1× PBS with 1% BSA and then resuspended in 1% paraformaldehyde or 1× PBS with 0.2% BSA.
Flow cytometry was performed on a LSR-II (BD) and the gating was performed using FlowJo software (Treestar) using the following criteria (applied to live singlets): CD4 T cells (CD3+CD4+CD8-CD19-; CD8 T cells (CD3+CD8+CD4-CD19-); NK cells (NK1.1+B220-CD3-); B cells (CD19+CD3-); double negative (DN) (B220-CD4-CD8-CD25varCD44var); double positive (DP) (CD4+CD8+B220-); single positive (SP) (CD8, CD8+CD4-B220-); single positive (SP) (CD4, CD4+CD8-B220-); hematopoietic stem cells (HSC) (LSK [Lin-, Sca-1+, c-Kit+)-CD34+-CD135-); multipotent progenitors (MPP) (LSK-CD34+CD135+); common lymphoid progenitors (CLP) (Lin-Sca-1loc-KitloIL7Rα+); and common myeloid progenitors (CMP) (CD34+CD16/32-). Negative gates were set using fluorescence-minus-one controls (FMO).
Competitive HSC transplant assay
Bone marrow from age- and sex-matched mice was collected and pooled with like genotypes (as described in flow cytometry analysis section) from mice that were 8 to 9 weeks in age: PepBoy/CD45.1 (n=3 females per experiment; mean age = 9 weeks) (Jackson Laboratory, 002014), Firre WT/CD45.2 (n=3 females per experiment mean age = 8.9 weeks), and ΔFirre/CD45.2 (n=3 females per experiment; mean age = 8.6 weeks). Bone marrow was lineage depleted according to the manufacture protocol (MiltinyiBiotec, 130-042-401), and cell marker surface staining was performed (as described for bone marrow in flow cytometry analysis section). Red blood cells were then lysed for 15 minutes at room temperature using BD Pharm Lyse (BD, 555899). Cells were washed twice in 1× HBSS with 5% FBS and 2 mM EDTA. We then double sorted lineage depleted cells for an HSC-enriched population (Lin- Sca-1+c-Kit+CD34+-CD135-) into 2% FBS in HBSS using fluorescent activated cell sorting (FACS) (BD Aria). Recipient mice, PepBoy/CD45.1 (n=10 males per experiment; mean age = 8.6 weeks), were lethally irradiated using a split 9.5γ split dose (3 hours apart). Firre WT and ΔFirre HSCs were mixed at a 1:1 ratio to PepBoy/CD45.1 HSCs and 100 ul containing 4,000 cells were transplanted by retro-orbital injection using 30-gauge insulin syringes (BD, 328411) into the lethally irradiated recipients. 48 hours post-transplant, 100,000 helper marrow cells from male PepBoy/CD45.1 were transplanted by retro-orbital injection into each experimental PepBoy/CD45.1 male recipient mouse.
MEF preparations and culture
We generated Firre WT, Firre knockout, and Firrerescue MEFs at E13.5 from intercrosses between male Firre-/y with female Firre+/- and male Firrerescue with female Firre-/-. Individual embryos were dissected into 1× phosphate-buffered saline (PBS) and were eviscerated, and the head, forelimbs, and hindlimbs were removed. Embryo carcasses were placed into individual 6 cm2 tissue culture plates containing 1 mL of pre-warmed 37°C TrypLE (Thermo Fisher, 12604013) and were incubated at 37°C for 20 min. Embryos were dissociated by gently pipetting using a P1000 tip and MEF media was added. Cells were cultured for 5 to 7 days and cryostocks of individual lines were generated. Subsequent experiments were performed from thaws from the cryostocks up to passage 3. MEFs were genotyped for Firre WT, knockout, rtTA, tg(Firre), and sry alleles. MEF culture media: 1× Dulbecco’s modified Eagle’s medium (DMEM) (Invitrogen 11965-118), 10% fetal bovine serum (Gibco, 10082139), L-glutamine (Thermo Fisher, 25030081), and penicillin/streptomycin (Thermo Fisher, 15140122).
Firre RNA FISH
Firre WT, ΔFirre, and Firrerescue MEFs were plated at a density of 50,000 cells per well onto round glass cover slips in a 24-well plate. Firrerescue MEFs were cultured with either 2 ug/mL dox (Sigma, D9891) or vehicle (ddH2O) for 24 hours. Replicate wells were processed for either RNA FISH or to isolate RNA for Firre induction analysis by qRT-PCR. RNA FISH using oligo probes was performed as previously described69. Briefly, Firre oligo probes were designed using Primer3 (http://frodo.wi.mit.edu/primer3/) and synthesized by Integrated DNA Technologies. After Amine-ddUTP (Kerafast) was added to 2 pmol of pooled oligos by terminal transferase (New England Biolabs), oligos were labeled withAlexa647 NHS-ester (Life Technologies) in 0.1 M sodium borate. Cells grown on glass coverslips were rinsed in PBS and fixed in 4% paraformaldehyde. After permeabilization in 0.5% Triton X-100 at room temperature, cells were washed in PBS and dehydrated in a series of increasing ethanol concentrations. 6 labeled oligo probes were added to hybridization buffer containing 25% formamide, 2× SSC, 10% dextran sulfate, and 1 mg/mL yeast tRNA. RNA FISH was performed in a humidified chamber at 42°C for 4 hours. After being washed three times in 2X SSC, cells were mounted for wide-field fluorescent imaging or dehydrated for STORM imaging. Nuclei were counter-stained with Hoechst 33342 (Life Technologies). The following pooled oligos against Firre were used: (1) AGCAGCAAATCCCAGGGGCC, (2) TTCCTCATTCCCCTTCTCCTGG, (3) CCCATCTGGGTCCAGCAGCA, (4) ATCAGCTGTGAGTGCCTTGC, (5) TCCAGTGCTTGCTCCTGATG, (6) GCCATGGTCAAGTCCTGCAT
Firre DNA/RNA and Xist RNA co-FISH
Primary MEF cells were trypsinzed and cytospun to glass slides. After brief air drying, cells were incubated in PBS for 1min, CSK/0.5% Triton X-100 for 2 min on ice, and CSK for 2 min on ice. Cells were fixed in 4% formaldehyde in PBS for 10 min at RT and washed twice in PBS. After dehydrated through series of EtOH, cells were subject to hybridization at 37°C O/N with denatured digoxigenin-labeled Xist probe (50% formamide, 2× SSC, 10% dextran sulfate, 0.1 mg/mL CoT1 DNA). Cells were washed in 50% formamide, 2X SSC at 37 ° C and in 2× SSC at RT, three times each. RNA FISH signal was detected by incubating FITC-labeled anti-digoxygenin antibody (Roche) in 4× SSC, 0.1% Tween-20 at 37 °C for 1 hour and followed by washing in 4× SSC, 0.1% Tween-20 at 37 °C three times. Cells were fixed again in 4% formaldehyde in PBS for 10 min and washed twice in PBS. Cellular RNAs were removed by RNase A (Life Technology) in PBS at 37°C. After dehydrated through series of EtOH, cells were sealed in hybridization buffer (50% formamide, 2X SSC, 10% dextran sulfate, 0.1 mg/mL CoT-1 DNA) containing Cy3-labeled Firre probe (Fosmid WI-755K22). Chromosomal DNA and probes were denaturated at 80°C for 15 min and allowed to renature by cooling down to 37° C O/N. Cells were washed in 50% formamide, 2X SSC at 37°C and in 2X SSC at RT, three times each. Nuclei were counter-stained with Hoechst 33342 (Life Technology). Imaging was performed on Nikon 90i microscope equipped with a 60X/1.4 N.A. VC objective lens, Orca ER camera (Hamamatsu) and Volocity software (Perkin Elmer). All probes were prepared by nick translation using DNA polymerase I (New England Biolab), DNase I (Promega), and Digoxigenin-dUTP (Roche), or Cy3-dUTP (Enzo Life Sciences)
Skeletal preparations
WT and ΔFirre E18.5 embryos were dissected and eviscerated. Samples were fixed in 100% ethanol for 24 hours at room temperature. Embryos were then placed in 100% acetone for 24 hours at room temperature and then incubated in staining solution (0.3% alcian blue 8GS (Sigma) and 0.1% Alizarin Red S (Sigma) in 70% ethanol containing 5% acetic acid) for three days at 37°C. Samples were then rinsed with distilled water and then placed in 1% potassium hydroxide at room temperature for 24 hours. Samples were then cleared in a series of 1% potassium hydroxide / 20%, 50%, and 80% glycerol.
AUTHOR CONTRIBUTIONS
Study conceptualization and design: J.P.L, J.C.L, and J.L.R; Firre ES cell targeting: A.W., J.H., R.A.F; Transgenic mice generation and mouse husbandry, N.C. and J.P.L; Immunophenotyping experiments: J.P.L, J.C.L, and J.M.G.; Competitive chimera design and analysis: J.M.G, J.P.L, A.J.W; Endotoxic shock experiments: J.P.L, N.C, and C.G; RNA-sequencing design and analysis: T.H, J.P.L, C.G, W.M, A.G; RNA FISH for Firre: H.S and J.T.L; Funding and supervision: A.J.W and J.L.R; Writing manuscript J.P.L, J.C.L, and J.L.R with input from all of the authors.
ACKNOWLEDGEMENTS
We thank Dr. Martin Sauvageau for providing a Firre clone to generate a riboprobe; Dr. Diana Sanchez for assistance in the mouse facility; Dr. Susan Carpenter, Dr. Kate Pritchett-Corning, and Elektra Robinson for discussions on the LPS study; Joyce LaVecchio and Silvia Ionescu in the HSCRB flow cytometry core for FACS assistance; the Harvard Bauer Core for sequencing; Dr. Marta Mele for initial optimization for RNA-seq analysis; and Dr. Laurie Chen and Dr. Lin Wu at the Harvard Genome Modification Facility. This research was supported by the National Institutes of Health (NIH) General Medical Sciences postdoctoral fellowship award 1F32GM122335-01A1 (to J.P.L) and support from NIH National Heart, Lung, and Blood Institute T32HL007893; NIH postdoctoral fellowship F32AG050395 (to J.M.G.); R.A.F is supported by the Howard Hughes Medical Institute; NIH RO1 AG048917 and the Dean’s Initiative Award Program for Innovation Grants in the Basic and Social Sciences (to A.J.W). the Institute of Mental Health grant R01MH102416-03 and the NIH Institute of General Medical Sciences grant P01GM099117 (to J.L.R).
REFERENCES




