

Abstract
Coordination of polarity between cells in tissues is key to multicellular organism development. In animals, coordination of this tissue cell polarity often requires direct cell-cell interactions and cell movements, which are precluded in plants by a wall that separates cells and holds them in place; yet plants coordinate the polarity of hundreds of cells during the formation of the veins in their leaves. Overwhelming experimental evidence suggests that the plant signaling molecule auxin coordinates tissue cell polarity to induce vein formation, but how auxin does so is unclear. The prevailing hypothesis proposes that GNOM, a regulator of vesicle formation during protein trafficking, positions auxin transporters of the PIN-FORMED family to the correct side of the plasma membrane. The resulting cell-to-cell, polar transport of auxin would coordinate tissue cell polarity and would induce vein formation. Here we tested this hypothesis by means of a combination of cellular imaging, molecular genetic analysis, and chemical induction and inhibition. Contrary to predictions of the hypothesis, we find that auxin-induced vein formation occurs in the absence of PIN-FORMED proteins or any known intercellular auxin transporter, that the residual auxin-transport-independent vein-patterning activity relies on auxin signaling, and that a GNOM-dependent signal that coordinates tissue cell polarity to induce vein formation acts upstream of both auxin transport and signaling. Our results reveal synergism between auxin transport and signaling, and their unsuspected control by GNOM, in the coordination of tissue cell polarity during vein patterning, one of the most spectacular and informative expressions of tissue cell polarization in plants.
Introduction
How the polarity of the cells in a tissue is coordinated is a central question in biology. In animals, the coordination of this tissue cell polarity requires direct cell-cell communication and often cell movements (Goodrich and Strutt, 2011), both of which are precluded in plants by a wall that holds cells in place; therefore, tissue cell polarity is coordinated by a different mechanism in plants.
Plant veins are an expression of coordinated tissue cell polarity (Sachs, 1991b; Sachs, 2000; Boutte et al., 2007; Nakamura et al., 2012). This is reflected in the relation between the parts of the vein, and between the veins and the parts of the plant: vascular elements are elongated along the length of the vein and are connected to one another at their ends (Esau, 1942), and veins primarily connect shoot organs with roots (Dengler, 2006); therefore, veins and their elements are unequal at their ends — one end connects to shoot tissues, the other to root tissues — and are thus polar (Sachs, 1975). Not all the veins in closed networks such as those of Arabidopsis leaves have unambiguous shoot-to-root polarity, but the vein networks themselves are polar (Sachs, 1975).
Just as veins are an expression of coordinated tissue cell polarity, their formation is an expression of coordination of tissue cell polarity; this is most evident in developing leaves. Consider, for example, the formation of the midvein at the center of the cylindrical leaf primordium. Initially, the plasma-membrane (PM)-localized PIN-FORMED1 (PIN1) protein of Arabidopsis (Galweiler et al., 1998), which catalyzes cellular efflux of the plant signal auxin (Petrasek et al., 2006), is expressed in all the inner cells of the leaf primordium (Benkova et al., 2003; Reinhardt et al., 2003; Heisler et al., 2005; Scarpella et al., 2006; Wenzel et al., 2007; Bayer et al., 2009; Verna et al., 2015); over time, however, PIN1 expression becomes gradually restricted to the file of cells that will form the midvein. PIN1 localization at the PM of the inner cells is initially isotropic, or nearly so, but as PIN1 expression becomes restricted to the site of midvein formation, PIN1 localization becomes polarized: in the cells surrounding the developing midvein, PIN1 localization gradually changes from isotropic to medial, i.e. toward the developing midvein, to mediobasal; and in the cells of the developing midvein, PIN1 becomes uniformly localized toward the base of the leaf primordium, where the midvein will connect to the pre-existing vasculature. Both the restriction of PIN1 expression and the polarization of PIN1 localization initiate and proceed away from pre-existing vasculature and are thus polar.
The correlation between (1) coordination of tissue cell polarity, as expressed by the coordination of PIN1 polar localization between cells, (2) polar auxin transport, as expressed by the auxin-transport-polarity-defining localization of PIN1 (Wisniewska et al., 2006), and (3) the polar formation of veins, themselves polar, does not seem to be coincidental. Auxin application to developing leaves induces the formation of broad expression domains of isotropically localized PIN1; such domains become restricted to the sites of auxin-induced vein formation, and PIN1 localization becomes polarized toward the pre-existing vasculature (Scarpella et al., 2006). Both the restriction of PIN1 expression domains and the polarization of PIN1 localization are delayed by chemical inhibition of auxin transport (Scarpella et al., 2006; Wenzel et al., 2007), which induces vein pattern defects similar to, though stronger than, those of pin1 mutants (Mattsson et al., 1999; Sieburth, 1999; Sawchuk et al., 2013).
Therefore, available evidence suggests that auxin coordinates tissue cell polarizaty to induce polar-vein-formation, and it seems that such coordinative and inductive property of auxin strictly depends on the function of PIN1 and possibly other PIN genes. How auxin precisely coordinates tissue cell polarity to induce polar-vein-formation is unclear, but the current hypothesis is that the GNOM (GN) guanine-nucleotide exchange factor for ADP-rybosilation-factor GTPases, which regulates vesicle formation in membrane trafficking, controls the cellular localization of PIN1 and other PIN proteins; the resulting cell-to-cell, polar transport of auxin would coordinate tissue cell polarity and control polar developmental processes such as vein formation (reviewed in, e.g., (Berleth et al., 2000; Richter et al., 2010; Nakamura et al., 2012; Linh et al., 2018)).
Here we tested this hypothesis by a combination of cellular imaging, molecular genetic analysis and chemical induction and inhibition. Contrary to predictions of the hypothesis, we found that auxin-induced polar-vein-formation occurs in the absence of PIN proteins or any known intercellular auxin transporter, that the residual auxin-transport-independent veinpatterning activity relies on auxin signaling, and that a GN-dependent tissue-cell-polarizing signal acts upstream of both auxin transport and signaling.
Results
Contribution of the GNOM Gene to Coordination of Tissue Cell Polarity During Arabidopsis Vein Formation
The current hypothesis of how auxin coordinates tissue cell polarity to induce polar-vein-formation proposes that the GNOM (GN) guanine-nucleotide exchange factor for ADP-ribosylation-factor GTPases, which regulates vesicle formation in membrane trafficking, controls the cellular localization of PIN1; the resulting cell-to-cell, polar transport of auxin would coordinate cell polarity between cells, and control polar developmental processes such as vein formation (reviewed in, e.g., (Berleth et al., 2000; Richter et al., 2010; Nakamura et al., 2012; Linh et al., 2018)). As such, the hypothesis predicts that the restriction of PIN1 expression domains and coordination of PIN1 polar localization that normally occur during vein formation (Benkova et al., 2003; Reinhardt et al., 2003; Heisler et al., 2005; Scarpella et al., 2006; Wenzel et al., 2007; Bayer et al., 2009; Sawchuk et al., 2013; Marcos and Berleth, 2014; Verna et al., 2015) would occur abnormally, or fail to occur altogether, during gn-mutant leaf development.
We first tested this prediction by imaging expression domains of PIN1::PIN1:YFP (PIN1:YFP fusion protein expressed by the PIN1 promoter (Xu et al., 2006)) in WT and in the new strong allele gn-13 (Table S1) during first-leaf development.
Consistent with previous reports (Benkova et al., 2003; Reinhardt et al., 2003; Heisler et al., 2005; Scarpella et al., 2006; Wenzel et al., 2007; Bayer et al., 2009; Sawchuk et al., 2013; Marcos and Berleth, 2014; Verna et al., 2015), in WT leaves PIN1::PIN1:YFP was expressed in all the cells at early stages of tissue development; over time, epidermal expression became restricted to the basal-most cells and inner tissue expression became restricted to files of vascular cells (Fig. 1A-J).

(A–Q,T,U) Top right: leaf age in days after germination (DAG). (A–E) Veins form sequentially during Arabidopsis leaf development: the formation of the midvein (mv) is followed by the formation of the first loops of veins (“first loops”; 11), which in turn is followed by the formation of second loops (12) and minor veins (hv) (Mattsson et al., 1999; Sieburth, 1999; Kang and Dengler, 2004; Scarpella et al., 2004). Loops and minor veins differentiate in a tip-to-base sequence during leaf development. Increasingly darker grays depict progressively later stages of vein development. Boxes in C and D illustrate positions of closeups in P and T. 13: third loop. (F–W) Confocal laser scanning microscopy. First leaves. For simplicity, only half-leaves are shown in H–J and L–O. Dashed white line in F–R, T, U and V delineates leaf outline. (F–Q,T,U) Top right: genotype. (F–P,R–T,V,W) Bottom left: reproducibility index. (F–O) PIN1::PIN1:YFP expression; look-up table (ramp in F) visualizes expression levels. (P,R–T,V,W) PIN1::PIN1:GFP expression; look-up table (ramp in P) visualizes expression levels. Red: autofluorescence. Stars in P label cells of the developing second loop. (Q,U) PIN1::PIN1:YFP expression. Boxes in Q and in U illustrate positions of closeups in R and S and in V and W, respectively. Bars: (F,P,R–T,V,W) 10 μm; (G,I,L,Q) 30 μm; (H,K) 20 μm; (J,M–O,U) 60 μm.
In gn leaves too, PIN1::PIN1:YFP was expressed in all the cells at early stages of tissue development and over time epidermal expression became restricted to the basal-most cells; however, inner tissue expression failed to become restricted to files of vascular cells and instead remained nearly ubiquitous even at very late stages of leaf development (Fig. 1K-O).
We next tested the prediction by imaging cellular localization of expression of PIN1::PIN1:GFP (Benkova et al.,2003) in WT and during first-leaf development. Hereafter, we use “basal” to describe localization of PIN::PIN1:GFP expression oriented toward pre-existing veins, irrespective of how these veins are positioned within a leaf.
Consistent with previous reports (Benkova et al., 2003; Reinhardt et al., 2003; Heisler et al., 2005; Scarpella et al., 2006; Wenzel et al., 2007; Bayer et al., 2009; Sawchuk et al., 2013; Marcos and Berleth, 2014; Verna et al., 2015), in the cells of the second pair of vein loops (“second loop” hereafter) at early stages of its development in WT leaves, PIN1::PIN1:GFP expression was mainly localized to the basal side of the plasma membrane (PM), toward the midvein; in the inner cells flanking the developing loop, PIN1::PIN1:GFP expression was mainly localized to the side of the PM facing the developing loop; and in the inner cells further away from the developing loop, PIN1::PIN1:GFP expression was localized isotropically, or nearly so, at the PM (Fig. 1C,P). At later stages of second-loop development, by which time PIN1::PIN1:GFP expression had become restricted to the sole, elongated cells of the developing loop, PIN1::PIN1:GFP expression was localized to the basal side of the PM, toward the midvein (Fig. 1D,T).
At early stages of development of the tissue that in gn leaves corresponds to that from which the second loop forms in WT leaves, PIN1::PIN1:GFP was expressed uniformly in the outermost inner tissue and expression was localized isotropically, or nearly so, at the PM (Fig. 1Q,R). PIN1::PIN1:GFP was expressed more heterogeneously in the innermost inner tissue, but expression remained localized isotropically, or nearly so, at the PM, except in cells near the edge of higher-expression domains (Fig. 1Q,S); in those cells, localization of PIN1::PIN1:GFP expression at the PM was weakly polar, but such weak cell polarities pointed in seemingly random directions (Fig. 1Q,S).
At late stages of gn leaf development, heterogeneity of PIN1::PIN1:GFP expression had spread to the outermost inner tissue, but expression remained localized isotropically, or nearly so, at the PM, except in cells near the edge of higher-expression domains (Fig. 1U,V); in those cells, localization of PIN1::PIN1:GFP expression at the PM was weakly polar, but such weak cell polarities pointed in seemingly random directions (Fig. 1U,V). Heterogeneity of PIN1::PIN1:GFP expression in the innermost inner tissue had become more pronounced at late stages of gn leaf development, and the weakly polar localization of PIN1:: PIN1:GFP expression at the PM had spread to the center of the higher-expression domains (Fig. 1U,W); nevertheless, such weak cell polarities still pointed in seemingly random directions (Fig. 1U,W). Finally, none of the cells had acquired the elongated shape characteristic of vascular cells in WT (Fig. 1U-W).
In conclusion, consistent with previous observations (Steinmann et al., 1999; Kleine-Vehn et al., 2008), both restriction of PIN1 expression domains and coordination of PIN1 polar localization occur only to a very limited extent or fail to occur altogether during gn leaf development, which is consistent with the current hypothesis of how auxin coordinates tissue cell polarity to induce polar-vein-formation.
Contribution of GN to Vein Patterning
We asked whether the very limited or altogether absent restriction of PIN1 expression domains and coordination of PIN1 polar localization occurring during gn leaf development (Figure 1) were associated with vein pattern defects in mature gn leaves.
WT Arabidopsis grown under normal conditions forms separate leaves whose vein networks are defined by at least four reproducible features (Telfer and Poethig, 1994; Nelson and Dengler, 1997; Kinsman and Pyke, 1998; Candela et al., 1999; Mattsson et al., 1999; Sieburth, 1999; Steynen and Schultz, 2003; Sawchuk et al., 2013; Verna et al., 2015) (Fig. 2A,B): (1) a narrow I-shaped midvein that runs the length of the leaf; (2) lateral veins that branch from the midvein and join distal veins to form closed loops; (3) minor veins that branch from midvein and loops, and either end freely or join other veins; (4) minor veins and loops that curve near the leaf margin, lending a scalloped outline to the vein network.

(A,B) Vein pattern of WT mature first leaf. In A: red, midvein; orange, loops; gray, minor veins. (B–J) Dark-field illumination of mature first leaves illustrating phenotype classes (top right): class 0, narrow I-shaped midvein and scalloped vein-network outline (B); class a1, dense vein network and apically thickened vein-network outline (not shown); class a2, open vein-network outline (C); class a3, fragmented vein network (D); class a4, open veinnetwork outline and fragmented vein network (E); class a5, open vein-network outline, fragmented vein network and apically thickened vein-network outline (F); class a6, wide midvein, dense network of thick veins and jagged vein-network outline (G); class a7, dense network of thick veins that fail to join the midvein in the bottom half of the leaf and pronouncedly jagged vein-network outline (H); class a8, wide midvein and shapeless vascular cluster (I); class a9, fused leaves with wide midvein and shapeless vascular cluster (not shown); class a10, shapeless vascular cluster (J). (K–M) Details of vascular clusters illustrating vascular elements uniformly oriented perpendicular to the leaf margin (K) (class a6), vascular elements oriented seemingly randomly at the distal side of the cluster and parallel to the leaf axis at the proximal side of the cluster (L) (classes a8 and a9), and seemingly random orientation of vascular elements (M) (classed a8–a10). (N) Percentages of leaves in phenotype classes. Difference between gn-18 and WT, between gnfwr and WT, between gnB/E and WT, between gnR5 and WT, between gnvan7 and WT, between gnvan7+fwr;gn-13 and WT, between gnSALK_103014 and WT, between gn-13 and WT, and between emb30-8 and WT was significant at P<0.001 (***) by Kruskal-Wallis and Mann-Whitney test with Bonferroni correction. Sample population sizes: WT, 58; gn-18, 172; gnfwr, 43; gnB/E, 80; gnR5, 93; gnvan7, 109; gnvan7+fwr;gn-13 no. 5, 97; gnvan7+fwr;gn-13 no. 7, 93; gnSALK_103014, 32; gn-13, 56; gnemb30-8, 45. Bars: (B–F) 1 mm; (G) 0.75 mm; (H,I) 0.5 mm; (J) 0.25 mm; (K–M) 50 μm.
In ~25% of the leaves of the new weak allele gn-18 (Table S1) (Figure S1) closed loops were often replaced by open loops, i.e. loops that contact the midvein or other loops at only one of their two ends (Fig. 2C,N). Moreover, in ~50% of gn-18 leaves veins were often replaced by “vein fragments”, i.e. stretches of vascular elements that fail to contact other stretches of vascular elements at either one of their two ends (Fig. 2D,E,N). Loops were open and veins were fragmented also in the leaves of both gnfwr (Okumura et al., 2013) and gnB/E (Geldner et al., 2004) (Fig. 2N). In addition, the vein network of gnB/E leaves was denser and its outline was thicker near the leaf tip (Fig. 2F,N).
The vein network was denser also in all the leaves of gnR5 (Geldner et al., 2004), in nearly 70% of those of gnvan7 (Koizumi et al., 2000) and in ~40% of those of gnvan7+fwr;gn-13 — in which we had combined the van7 and fwr mutations (Table S1) (Fig. 2G,N). However, in the leaves of these backgrounds — unlike in those of gnB/E — all the veins were thicker; lateral veins failed to join the midvein but ran parallel to it to form a “wide midvein”; and the vein network outline was jagged because of narrow clusters of vascular elements that were oriented perpendicular to the leaf margin and that were laterally connected by veins (Fig. 2G,K,N). These features were enhanced in ~20% of the leaves of gnvan7, in ~55% of those of gnvan7+fwr;gn-13 and in ~5% of those of gnSALK_103014 (Okumura et al., 2013): the vein network was denser, veins failed to join the midvein in the bottom half of the leaf, and the vein network outline was pronouncedly jagged (Fig. 2H,N).
Consistent with previous observations (Shevell et al., 2000), in the few remaining leaves of gnvan7 and gnvan7+fwr; gn-13, and in most of those of gnSALK_103014, gn-13 and gnemb30-8 (Franzmann et al., 1989; Moriwaki et al., 2014), a central, shapeless vascular cluster was connected with the basal part of the leaf by a wide midvein, and vascular elements were oriented seemingly randomly at the distal side of the cluster and progressively more parallel to the leaf axis toward the proximal side of the cluster (Fig. 2I,L–N).
Finally, in the remaining leaves of gnSALK_103014, gn-13 and gnemb30-8, vascular differentiation was limited to a central, shapeless cluster of seemingly randomly oriented vascular elements (Fig. 2J,M,N).
We conclude that defects in coordination of PIN1 polar localization and possible derived defects in polar auxin transport during gn leaf development are associated with vein pattern defects in mature gn leaves.
Contribution of Plasma-Membrane-Localized PIN Proteins to Vein Patterning
Were the vein pattern defects of gn the sole result of loss of PIN1-mediated polar auxin-transport induced by defects in coordination of PIN1 polar localization, the vein pattern defects of gn would be phenocopied by simultaneous mutation in all the PIN genes with function in PIN1-dependent vein patterning; we asked whether that were so.
In Arabidopsis, the PIN family of auxin transporters is composed of eight members (Paponov et al., 2005; Krecek et al., 2009; Viaene et al., 2012): PIN5, PIN6 and PIN8, which are primarily localized to the endoplasmic reticulum (ER) (Mravec et al., 2009; Bosco et al., 2012; Ding et al., 2012; Sawchuk et al., 2013), and PIN1, PIN2, PIN3, PIN4 and PIN7 are primarily localized to the plasma membrane (PM) and catalyze cellular auxin efflux (Chen et al., 1998; Galweiler et al., 1998; Luschnig et al., 1998; Muller et al., 1998; Friml et al.,2002a; Friml et al., 2002b; Friml et al., 2003; Petrasek et al., 2006; Yang and Murphy, 2009; Zourelidou et al., 2014). Sequence analysis divides the PM-localized subfamily of PIN (PM-PIN) proteins into three groups: the PIN1 group, the PIN2 group and the PIN3 group, which also contains PIN4 and PIN7 (Krecek et al., 2009; Viaene et al., 2012).
Mutants of PIN1 are the only pin single mutants with vein pattern defects, and the vein pattern defects of double mutants between pin1 and mutants of PIN2, PIN3, PIN4 or PIN7 are no different from those of pin1 single mutants (Sawchuk et al., 2013), suggesting that either PIN2, PIN3, PIN4 and PIN7 have no function in PIN1-dependent vein patterning or their function in this process is redundant. To discriminate between these possibilities, we first assessed the collective contribution to PIN1-dependent vein patterning of the PM-PIN genes of the PIN3 group (PIN3, PIN4 and PIN7), whose translational fusions to GFP (Zadnikova et al., 2010; Bennett et al., 2016; Belteton et al., 2018) (Table S1) are all expressed — as are translational fusions of PIN1 to GFP (Benkova et al., 2003; Heisler et al., 2005; Scarpella et al., 2006; Wenzel et al., 2007; Bayer et al., 2009; Marcos and Berleth, 2014) — in both epidermal and inner cells of the developing leaf (Fig. 3A,C-E).

(A–K) Top right: expression-reported gene, phenotype class or genotype. (B–E) Bottom left: reproducibility index. (A–E) Confocal laser scanning microscopy with (A) or without (B–E) transmitted light; 4-day-old first leaves. Dashed magenta line delineates leaf outline. (A) PIN1::PIN1:GFP expression. (B) PIN2::PIN2:GFP expression. (C) PIN3::PIN3:GFP expression. (D) PIN4::PIN4:GFP expression. (E) PIN7::PIN7:GFP expression. (F–I) Dark-field illumination images of mature first leaves illustrating phenotype classes: class b1, Y-shaped midvein and scalloped vein-network outline (F); class b2, fused leaves with scalloped vein-network outline (G); class b3, thick veins and scalloped vein-network outline (H); class b4, fused leaves with thick veins and scalloped vein-network outline (I). (J,K) Differential interference images of details of WT (J) or pin1-1,3;4;7 (K) illustrating normal (classes 0, b1 and b2) or thick (classes b3 and b4) veins, respectively. (L) Percentages of leaves in phenotype classes. Difference between pin1-1 and WT, between pin1-134 and WT, between pin1-1,3;4;7 and pin1-1, and between pin1-134,3;4;7 and pin1-134-wassignificantat P<0.001 (***) by Kruskal-Wallis and Mann-Whitney test with Bonferroni correction. Sample population sizes: WT, 58; pin2;5;4;7, 49; pin3,4;7, 102; pin1-1, 81; pin1-134, 48; pin1-1,3;4;7,7ĩ>; pin1-134,3;4;7, 45, pin1-1,3;2;4;7, 99. Bars: (A–E) 0.1 mm; (F–H) 1 mm; (I) 5 mm; (J,K) 50 μm.
Consistent with previous reports (Sawchuk et al., 2013; Verna et al., 2015), the vein patterns of most of the pin1 leaves were abnormal (Fig. 3F,G,L). pin3;pin4;pin7 (pin3;4;7 hereafter) embryos were viable and developed into seedlings (Table S2) whose vein patterns were no different from those of WT (Fig. 3L). pin1,3;4;7 embryes were viable (Table S3) and developed into seedlings (Table S4) that were smaller than pin1 seedlings (Fig. S2A,B). The cotyledon pattern defects of pin1,3;40;7 were more severe than those of pin1 (Fig. S3A–H), and the vein pattern defects of pin1,3;4;7 were more severe than those of pin1: no pin1,3;4;7 leaf had a WT vein pattern; pin1,3;4;7 veins were thicker; and ~15% of pin1,3;4;7 leaves were fused (Fig. 3H–L). However, as in WT, in pin1,3;4;7 vascular elements were elongated and aligned along the length of the vein (Fig. 3J,K).
Next, we asked whether mutation of PIN2 — whose translational fusion to GFP (Xu and Scheres, 2005) is only expressed in epidermal cells in the developing leaf (Fig. 3B) — changed the spectrum of vein pattern defects of pin1,3;4;7.
pin2;3;4;7 embryos were viable and developed into seedlings (Table S2) whose vein patterns were no different from those of WT (Fig. 3L). pin1,3;2;4;7 embryos were viable (Table S3) and developed into seedlings (Table S4) whose vein pattern defects were no different from those of pin1,3;4;7 (Fig. 3L). The cotyledon pattern defects of pin1,3;2;4;7 were more severe than those of pin1,3;4;7 (Fig. S3A–H), but the size of pin1,3;2;4;7 seedlings was similar to that of pin1,3;4;7 seedlings (Fig. S2A–C).
In conclusion, the PIN3 group of PM-PIN genes (PIN3, PIN4 and PIN7) provides no nonredundant function in vein patterning, but it contributes to PIN1-dependent vein patterning; PIN1 and the PIN3 group of PM-PIN genes redundantly restrict vascular differentiation to narrow zones; and PIN2 seems to have no function in any of these processes. Most important, loss of PM-PIN function fails to phenocopy the vein pattern defects of gn.
Contribution of PIN Genes to Vein Patterning
Expression and genetic analyses suggest that PIN1, PIN3, PIN4 and PIN7 redundantly define a single auxin-transport pathway with vein patterning functions whose loss fails to phenocopy the vein pattern defects of gn (Figure 2; Figure 3). The ER-localized PIN (ER-PIN) proteins PIN6 and PIN8, but not the ER-PIN protein PIN5, define a distinct auxin-transport pathway with vein patterning functions that overlap with those of PIN1 (Sawchuk et al., 2013; Verna et al., 2015). We asked what the collective contribution of these two auxin-transport pathways were to vein patterning and whether simultaneous mutation in all the PIN genes with vein patterning function phenocopied the vein pattern defects of gn.
As previously reported (Sawchuk et al., 2013), the vein pattern of pin6;8 was no different from that of WT (Fig. 4C). pin1,3,6;4;7;8 embryos were viable (Table S3) and developed into seedlings (Table S4) whose vein patterns differed from those of pin1,3,4;7 in four respects: (1) the vein network comprised more lateral veins; (2) lateral veins failed to join the midvein but ran parallel to it to form a wide midvein; (3) lateral veins ended in a marginal vein that closely paralleled the leaf margin, lending a smooth outline to the vein network; (4) veins were thicker (Figure 3; Fig. 4A–C). Simultaneous mutation of PIN6 and PIN8 in the pin1,3;4;7 background shifted the distribution of pin1,3;4;7 cotyledon pattern phenotypes toward stronger classes (Fig. S3A–H), but the size of pin1,3,6;4;7;8 seedlings was similar to that of pin1,3;4;7 seedlings (Fig. S2A,B,D).

(A,B) Dark-field illumination of mature first leaves illustrating phenotype classes (top right): class b7, wide midvein, more lateral-veins and conspicuous marginal vein (A); class b8, fused leaves with wide midvein, more lateral-veins and conspicuous marginal vein (B). (C) Percentages of leaves in phenotype classes (Classes 0, b3 and b4 defined in Figures 2 and 3). Difference between pin1-1,3;4;7;8 and pin1;4;7 was significant at P<0.001 (***) by Kruskal-Wallis and Mann-Whitney test with Bonferroni correction. Sample population sizes: WT, 51; pin6;8, 47; pin1-1,3;4;7, 49; pin1-1,3,6;4;7;8, 73. Bars: (A,B) 0.5 mm.
Because pin6;8 synthetically enhanced vein pattern defects of pin1,3;4;7, we conclude that the auxin-transport pathway mediated by PIN1, PIN3, PIN4 and PIN7 and that mediated by PIN6 and PIN8 provide overlapping functions in vein patterning. Nevertheless, loss of PIN-dependent vein patterning function fails to phenocopy the vein pattern defects of gn.
Genetic Versus Chemical Inhibition of Auxin Transport During Vein Patterning
Loss of PIN-dependent vein patterning function fails to phenocopy the vein pattern defects of gn (Figure 2; Figure 4), suggesting that these latter are not the sole result of loss of PIN-dependent polar auxin-transport induced by defects in coordination of PIN polar localization. However, it is possible that the vein pattern defects of gn result from additional or exclusive defects in PIN-independent polar auxin-transport pathways; we asked whether that were so.
Cellular auxin efflux is inhibited by a class of structurally related compounds referred to as phytotropins, exemplified by N-1-naphthylphthalamic acid (NPA) (Cande and Ray, 1976; Katekar and Geissler, 1980; Sussman and Goldsmith, 1981). Because PM-PIN proteins catalyze cellular auxin efflux (Chen et al., 1998; Petrasek et al., 2006; Yang and Murphy, 2009; Zourelidou et al., 2014), we first asked whether defects resulting from simultaneous mutation of all the PM-PIN genes with vein patterning function were phenocopied by growth of WT in the presence of NPA. To address this question, we compared defects of pin1,3;4;7 to those induced in WT by growth in the presence of 100 μM NPA, which is the highest concentration of NPA without toxic, auxin-efflux-unrelated effects (Petrasek et al., 2003; Dhonukshe et al., 2008). Because leaves develop more slowly at this concentration of NPA (Mattsson et al., 1999; Sieburth, 1999), to ensure maximal vascular differentiation we allowed them to grow for four weeks before analysis.
Consistent with previous reports (Mattsson et al., 1999; Sieburth, 1999), NPA only rarely induced leaf fusion in WT (see Fig. 6I for one such rare occurrence) but reproducibly induced characteristic vein-pattern defects: (1) the vein network comprised more lateral veins; (2) lateral veins failed to join the midvein but ran parallel to it to form a wide midvein; (3) lateral veins ended in a marginal vein that closely paralleled the leaf margin, lending a smooth outline to the vein network; (4) veins were thicker, though vascular elements were elongated and aligned along the length of the vein (Fig. 5A,D,E,H).

(A–G) Top right: genotype and treatment. (A–G) Dark-field illumination (A–D,F,G) or confocal laser scanning microscopy (E) of mature first leaves. (A) WT. (B) pin1-1,3;4;7. (C) pin1-1,3,6;4;7;8. (D) NPA-grown WT. (E) Detail illustrating thick veins in NPA-grown WT (compare with Fig. 3J). (F) NPA-grown pin1-1,3;4;7. (G) NPA-grown pin1-1,3,6;4;7;8. (G) Percentages of leaves in phenotype classes (defined in Figures 2-4). Sample population sizes: WT, 38; pin1-1,3;4;7, 30; pin1-1,3,6;4;7;8, 73; NPA-grown WT, 41; NPA-grown pin1-1,3;4;7, 58; NPA-grown pin1-1,3,6;4;7;8, 48. Bars: (A-D,F,G) 0.5 mm, (E) 25 μm.
By contrast, 20% of pin1,3;4;7 leaves were fused, and though pin1,3;4;7 veins were thick, pin1,3;4;7 vein patterns lacked all the other characteristic defects induced in WT by NPA (Fig. 5B,H). However, such defects were induced in pin1,3;4;7 by NPA (Fig. 5F,H), suggesting that this background has residual NPA-sensitive vein-patterning activity. The vein pattern defects induced in WT or pin1,3;4;7 by NPA were no different from those of pin1,3,6;4;7;8 (Fig. 5C,D–F,H). Because no additional defects were induced in pin1,3,6;4;7;8 by NPA (Fig. 5G,H), the residual NPA-sensitive vein-patterning activity of pin1,3;4;7 is likely provided by PIN6 and PIN8.
In conclusion, our results suggest that growth in the presence of NPA phenocopies defects of loss of PIN-dependent vein patterning function, that in the absence of this function any residual NPA-sensitive vein-patterning activity — if existing — becomes inconsequential, and that loss of neither PIN-dependent vein-patterning function nor NPA-sensitive vein-patterning activity phenocopies the vein pattern defects of gn.
Contribution of ABCB Genes to Vein Patterning
Loss of PIN-dependent vein-patterning function or of NPA-sensitive vein-patterning activity fails to phenocopy the vein pattern defects of gn (Figure 2; Figure 5), suggesting that these latter are not the sole result of loss of PIN-dependent or NPA-sensitive polar auxin-transport induced by defects in coordination of PIN polar localization. However, it is possible that the vein pattern defects of gn result from additional or exclusive defects in another polar auxin-transport pathway; we asked whether that were so.
Cellular auxin efflux is catalyzed not only by PM-PIN proteins but by the PM-localized ATP-BINDING CASSETTE B1 (ABCB1) and ABCB19 proteins (Geisler et al., 2005; Bouchard et al., 2006; Petrasek et al., 2006; Blakeslee et al., 2007; Yang and Murphy, 2009), whose fusions to GFP (Dhonukshe et al., 2008; Mravec et al., 2008) are expressed at early stages of leaf development (Fig. 6A,B). We asked whether ABCB1/19-mediated auxin efflux were required for vein patterning.
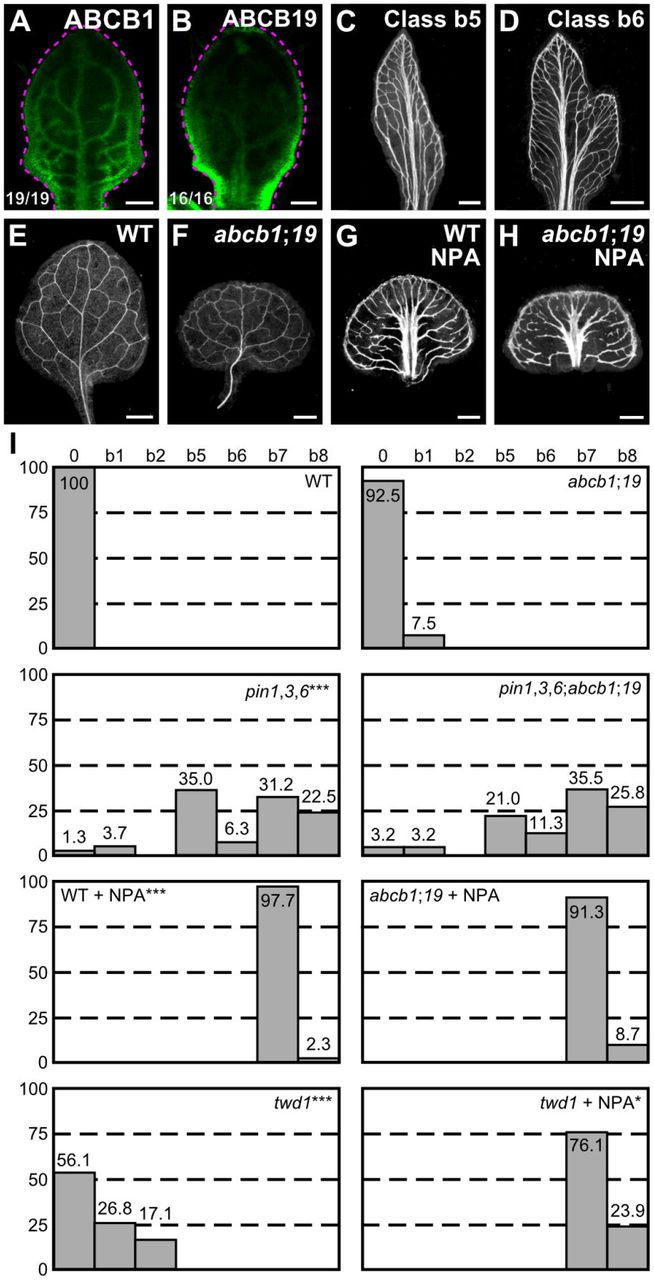
(A,B,E–H) Top right: expression-reported gene, genotype and treatment. (A–B) Bottom left: reproducibility index. (A–B) Confocal laser scanning microscopy; 5-day-old first leaves. Dashed magenta line delineates leaf outline. (A) ABCB1::ABCB1:GFP expression. (B) ABCBI9::ABCBI9:GFP expression. (C–H) Dark-field illumination of mature first leaves. (C,D) Phenotype classes: class b5, thick veins and conspicuous marginal vein (C); class b6, fused leaves with thick veins and conspicuous marginal vein (D). (I) Percentages of leaves in phenotype classes (Classes 0, b1, b2, b7 and b8 defined in Figures 2-4). Difference between pin1-1,3,6 and WT, between twd1 and WT, and between NPA-grown WT and WT was significant at P<0.001 (***), and between NPA-grown twd1 and NPA-grown WT was significant at P<0.05 (*) by Kruskal-Wallis and Mann-Whitney test with Bonferroni correction. Sample population sizes: WT, 41; abcb1;19, 40; pin1-1,3,6, 80; pin1-1,3,6;abcb1;19, 62; NPA-grown WT, 43; NPA-grown abcb1;19, 46; twd1, 41; NPA-grown twd1, 46. Bars: (A–B) 0.1 mm; (C–H) 0.5 mm.
The embryos of abcb1 and abcb19 were viable, but ~15% of abcb1;19 embryos died during embryogenesis (Table S5); nevertheless, the vein patterns of abcb1, abcb19 and abcb1;19 were no different from the vein pattern of WT (Fig. 6E,F,I), suggesting that ABCB1/19-mediated auxin efflux is dispensable for vein patterning.
Developmental functions of ABCB1/19-mediated auxin transport overlap with those of PIN-mediated auxin transport (Blakeslee et al., 2007; Mravec et al., 2008). We therefore asked whether vein pattern defects resulting from simultaneous mutation of PIN1, PIN3 and PIN6, or induced in WT by 100 μM NPA — which phenocopies loss of PIN-dependent vein-patterning function (Figure 5) — were enhanced by simultaneous mutation of ABCB1 and ABCB19.
pin1,3,6 embryos were viable (Table S6) and developed into seedlings (Table S7). The proportion of embryos derived from the self-fertilization of PIN1,PIN3,PIN6/pin1,pin3,pin6;abcb1/abcb1;abcb19/abcb19 that died during embryogenesis was no different from the proportion of embryos derived from the self-fertilization of abcb1/abcb1;abcb19/abcb19 that died during embryogenesis (Table S6), suggesting no nonredundant functions of PIN1, PIN3 and PIN6 in ABCB1/ABCB19-dependent embryo viability.
Consistent with previous reports (Blakeslee et al., 2007; Mravec et al., 2008), simultaneous mutation of ABCB1 and ABCB19 in the pin1,3,6 background shifted the distribution of pin1,3,6 cotyledon pattern phenotypes toward stronger classes (Figure S4). However, the spectrum of vein pattern phenotypes of pin1,3,6;abcb1;19 was no different from that of pin1,3,6, and the vein pattern defects induced in abcb1;19 by NPA were no different from those induced in WT by NPA (Fig. 6C,D,G–I), suggesting no vein-patterning function of ABCB1 and ABCB19 in the absence of function of PIN1, PIN3 and PIN6 or of NPA-sensitive, PIN-dependent vein-patterning function.
ABCB1 and ABCB19 are members of a large family (Geisler and Murphy, 2006); therefore, vein patterning functions of ABCB1/19-mediated auxin efflux might be masked by redundant functions provided by other ABCB transporters. The TWISTED DWARF1/ULTRACURVATA2 (TWD1/UCU2; TWD1 hereafter) protein (Kamphausen et al., 2002; Perez-Perez et al., 2004) is a positive regulator of ABCB-mediated auxin transport (Geisler et al., 2003; Bouchard et al., 2006; Bailly et al., 2008; Wu et al., 2010; Wang et al., 2013). Consistent with this observation, defects of twd1 are more severe than, though similar to, those of abcb1;19 (Geisler et al., 2003; Bouchard et al., 2006; Bailly et al., 2008; Wu et al., 2010; Wang et al., 2013). We therefore reasoned that analysis of twd1 vein patterns might uncover vein patterning functions of ABCB-mediated auxin transport that could not be inferred from the analysis of abcb1;19.
Approximately 25% of twd1 leaves had Y-shaped midveins and ~15% of twd1 leaves were fused (Fig. 6I), suggesting possible vein-patterning functions of TWD1-dependent ABCB-mediated auxin transport. However, vein pattern defects induced in twd1 by 100 μM NPA were no different from those induced in WT or abcb1;19 by NPA (Fig. 6I), suggesting that vein patterning functions of TWD1-dependent ABCB-mediated auxin transport — if existing — become inconsequential in the absence of NPA-sensitive, PIN-dependent vein-patterning function. By contrast, NPA enhanced leaf separation defects of twd1 (Fig. 6I), suggesting overlapping functions of TWD1-dependent ABCB-mediated auxin transport and NPA-sensitive, PIN-dependent auxin transport in leaf separation.
In conclusion, the residual vein patterning activity in pin mutants or in their NPA-induced phenocopy is not provided by ABCB1, ABCB19 or TWD1-dependent ABCB-mediated auxin transport, and loss of PIN- and ABCB-mediated auxin transport fails to phenocopy vein pattern defects of gn.
Contribution of AUX1/LAX Genes to Vein Patterning
Loss of PIN- and ABCB-mediated auxin transport fails to phenocopy vein pattern defects of gn (Figure 2; Figure 6), suggesting that these latter are not the sole result of loss of PIN-dependent, NPA-sensitive or ABCB-dependent polar auxin-transport. However, it is possible that the vein pattern defects of gn result from additional or exclusive defects in yet another auxin-transport pathway; we asked whether that were so.
Auxin is predicted to enter the cell by diffusion and through an auxin influx carrier (Rubery and Sheldrake, 1974; Raven, 1975). In Arabidopsis, auxin influx activity is encoded by the AUX1, LAX1, LAX2 and LAX3 (AUX1/LAX) genes (Parry et al., 2001; Yang et al., 2006; Swarup et al., 2008; Peret et al., 2012). We asked whether AUX1/LAX-mediated auxin influx were required for vein patterning.
aux1;lax1;2;3 embryos were viable (Table S8). Because the vein patterns of aux1;lax1;2;3 were no different from those of WT (Fig. 7A,C,D), we conclude that AUX1/LAX function is dispensable for vein patterning.

(A–D) Dark-field illumination of mature first leaves. Top right: genotype and treatment. (E) Percentages of leaves in phenotype classes (defined in Figures 2-4 and 6). Difference between pin1-1,3,6 and WT, between NPA-grown WT and WT, and between NPA-grown aux1-21;lax1;2;3 and NPA-grown WT was significant at P<0.001 (***) by Kruskal-Wallis and Mann-Whitney test with Bonferroni correction. Sample population sizes: WT, 53; aux1-21;lax1;2;3, 60; aux1-355;lax1-064, 77; pin1-1,3;6, 75; pin1-1,3,6;aux1-355;lax1-064, 58; NPA-grown WT, 46; NPA-grown aux1-21;lax1;2;3, 40. Bars: (A–D) 1 mm.
We next asked whether contribution of AUX1/LAX genes to vein patterning only became apparent in conditions of extremely reduced PIN-mediated auxin transport. To address this question, we tested whether vein pattern defects resulting from simultaneous loss of function of PIN1, PIN3 and PIN6 or induced in WT by 100 μM NPA, which phenocopies simultaneous mutation of all the PIN genes with vein patterning function (Figure 4), were enhanced by simultaneous mutation of AUX1 and LAX1 — the two AUX1/LAX genes that most contribute to shoot organ patterning (Bainbridge et al., 2008) — or of all AUX1/LAX genes, respectively.
The embryos derived from the self-fertilization of PIN1,pin3,PIN6/pin1,pin3,pin6;aux1/aux1;lax1/lax1 were viable (Table S9) and developed into seedlings (Table S10). The spectrum of vein pattern phenotypes of pin1,3,6;aux1;lax1 was no different from that of pin1,3,6 and the vein pattern defects induced in aux1;lax1;2;3 by NPA were no different from those induced in WT by NPA (Fig. 7B,D,E), suggesting no vein-patterning function of AUX1/LAX genes in conditions of extremely reduced auxin transport. On the other hand, simultaneous mutation of AUX1 and LAX1 in the pin1,3,6 background shifted the distribution of pin1,3,6 cotyledon pattern phenotypes toward stronger classes (Figure S4), and NPA induced leaf fusion in aux1;lax1;2;3 but not in WT (Fig. 7E), suggesting that AUX1/LAX-mediated auxin influx and NPA-sensitive, PIN-dependent, auxin transport have overlapping functions in cotyledon and leaf separation and that — consistent with previous observations (Reinhardt et al., 2003; Bainbridge et al., 2008; Kierzkowski et al., 2013) — AUX1/LAX-mediated auxin influx contributes to maintaining cotyledon and leaves separate in conditions of reduced auxin transport. Nevertheless, loss of PIN- and AUX1/LAX-mediated auxin transport fails to phenocopy the vein pattern defects of gn.
Genetic Interaction Between GN and PIN Genes
The vein pattern defects of gn are not the sole result of loss of PIN-dependent auxin transport (Figure 2; Figure 4; Figure 5); however, they could be the result of abnormal polarity of PIN-mediated auxin transport induced by defects in coordination of PIN polar localization. Were that so, the vein pattern defects of gn would depend on PIN genes, and therefore the vein pattern defects of gn;pin mutants would resemble those of pin mutants; we tested whether that were so.
We first asked what the phenotype were of the quintuple mutant between the strong allele gn-13 (Figure 2) and mutation in PIN1, PIN3, PIN4 and PIN7 — i.e. the PM-PIN genes with vein patterning function (Figure 3).
Consistent with previous observations (Mayer et al., 1993; Shevell et al., 1994), in gn seedlings hypocotyl and root were replaced by a basal peg, and the cotyledons were most frequently fused (Fig. S5A,C; Fig S6B; Fig. S7A,B). As shown above (Fig. S2A,B; Fig. S3A,H), pin1,3;4;7 seedlings had hypocotyl, short root, and a single cotyledon or two — either separate or fused — cotyledons (Fig. S5A,B; Fig S6C,D; Fig. S7B).
gn;pin1,3;4;7 embryos were viable (Table S11). A novel phenotype segregated in approximately one-sixteenth of the progeny of plants homozygous for pin3, pin4 and pin7 and heterozygous for pin1 and gn — no different from the one-sixteenth frequency expected for the gn;pin1,3;4;7 homozygous mutants by Pearson’s chi-squared (χ2) goodness-of-fit test (Table S12). We genotyped 10 of the seedlings with the novel mutant phenotype and found they were gn;pin1,3;4;7 homozygous mutants. gn;pin1,3;4;7 seedlings had hypocotyl, no root and the cotyledons were fused (Fig. S5A,D; Fig S6E; Fig. S7B).
WT cotyledons have a I-shaped midvein and three or four loops (Fig. S8A,B,K). All the veins of pin1,3;4;7 cotyledons were thick, and all pin1,3;4;7 cotyledons had three or four loops (Fig. S8C,D,K). In pin1,3;4;7 cotyledons, the proximal end of the first loops joined the midvein more basally than in WT, and minor veins branched from midvein and loops (Fig. S8C,D,K). Approximately 60% of pin1,3;4;7 cotyledons had an I-shaped midvein, while the remaining ~40% of them had a Y-shaped midvein (Fig. S8C,D,K).
Consistent with previous observations (Mayer et al., 1993; Shevell et al., 1994), in ~70% of gn cotyledons short stretches of vascular elements connected the proximal side of a central, shapeless cluster of seemingly randomly oriented vascular elements with the basal part of the cotyledon, while vascular differentiation was limited to a central, shapeless vascular cluster in the remaining ~30% of gn cotyledons (Fig. S8F,G,K). The vein pattern defects of gn;pin1,3;4;7 cotyledons were no different from those of gn; cotyledons (Fig. S8H,K), suggesting that the vein pattern phenotype of gn cotyledons is epistatic to that of pin1,3;4;7 cotyledons. Likewise, the vein pattern defects of gn;pin1,3;4;7 leaves were no different from those of gn leaves (Fig. 8A,B,E), suggesting that the vein pattern phenotype of gn leaves is epistatic to that of pin1,3;4;7 leaves.

(A–D) Dark-field illumination of mature first leaves. Top right: genotype and treatment. (E) Percentages of leaves in phenotype classes (defined in Figures 2-4). Difference between pin1-1,3;4;7 and WT, between pin1-1,3,6;4;7;8 and WT, between gn and WT, between gn-13;pin1-1,3;4;7 and pin1-1,3;4;7, between gn-13;pin1-1,3;6;4;7;8 and pin1-1,3,6;4;7;8 and between NPA-grown gn-13 and pin1-1,3,6;4;7;8, was significant at P<0.001 (***) by Kruskal-Wallis and Mann-Whitney test with Bonferroni correction. Sample population sizes: WT, 63; pin1-1,3;4;7, 53; pin1-1,3,6,4;7;8, 52; gn-13, 69; gn-13;pin1-1,3;4;7, 21; gn-13;pin1-1,3,6;4;7;8, 15; NPA-grown gn-13, 60. Bars: (A–D) 0.5 mm.
We next asked what the phenotype were of the septuple mutant between the strong allele gn-13 (Figure 2) and mutation in all the PIN genes with vein patterning function (Figure 4).
As shown above (Fig. S2A,D; Fig. S3A,H), pin1,3,6;4;7;8 seedlings had hypocotyl, short root and a single cotyledon or two fused cotyledons (Fig. S5A,E; Fig S6G,H; Fig. S7B).
gn;pin1,3,6;4;7;8 embryos were viable (Table S11). A phenotype similar to that of gn;pin1,3;4;7 segregated in approximately one-sixteenth of the progeny of plants homozygous for pin3, pin4, pin6, pin7 and pin8 and heterozygous for pin1 and gn — no different from the one-sixteenth frequency expected for the gn;pin1,3,6;4;7;8 homozygous mutants by Pearson’s χ2 goodness-of-fit test (Table S12). We genotyped 10 of the seedlings with the novel mutant phenotype and found they were gn;pin1,3,6;4;7;8 homozygous mutants. Like gn;pin1,3;4;7 seedlings, gn;pin1,3,6;4;7;8 seedlings had hypocotyl and no root, but unlike gn;pin1,3;4;7 seedlings ~90% of gn;pin1,3,6;4;7;8 seedlings had completely fused cup-shaped cotyledons (Fig. S5A,F; Fig S6I; Fig. S7B).
The vein pattern defects of pin1,3,6;4; 7;8 cotyledons were similar to those of pin1,3;4;7 cotyledons, but in ~85% of pin1,3,6;4;7;8 cotyledons the loops joined the midvein at the base of the cotyledon and the top half of the vein network outline was thick (Fig. S8C-E,K). The vein pattern defects of gn;pin1,3,6;4;7;8 cotyledons were no different from those of gn cotyledons (Fig. S8I-K), suggesting that the vein pattern phenotype of gn cotyledons is epistatic to that of pin1,3,6;4;7;8 cotyledons. Likewise, the vein pattern defects of gn;pin1,3,6;4;7;8 leaves were no different from those of gn leaves (Fig. 8C,E), suggesting that the vein pattern phenotype of gn leaves is epistatic to that of pin1,3,6;4;7;8 leaves. Finally, 100 μM NPA, which phenocopies loss of PIN-dependent vein-patterning function (Figure 5), failed to induce additional vein pattern defects in gn leaves (Fig. 8D,E).
In conclusion, our results suggest that the vein pattern defects of gn are not the result of either the sole loss of PIN-mediated auxin transport or the sole abnormal polarity of PIN-mediated auxin transport induced by defects in coordination of PIN polar localization.
Response of pin Leaves to Auxin Application
The uniform vein-pattern phenotype of pin1,3,6;4;7;8 was phenocopied by growth of WT in the presence of high concentration of NPA (Figure 5). Moreover, the vein-pattern phenotype of pin1,3,6;4;7;8 was unchanged by NPA treatment, and the NPA-induced vein-pattern phenocopy of pin1,3,6;4;7;8 was unchanged by mutation in any other known intercellular auxin-transporter (Figure 6; Figure 7). These observations suggest that the function of known intercellular auxin-transporters in vein patterning is dispensable in the absence of the auxin transport activity of PIN1, PIN3, PIN4, PIN6, PIN7 and PIN8. Because auxin transport is thought to be essential for auxin-induced vascular-strand formation (reviewed in (Sachs, 1981; Berleth et al., 2000; Aloni, 2010; Sawchuk and Scarpella, 2013)), we asked whether auxin-induced vein formation in pin1,3,6;4;7;8 and consequently whether veins were formed by an auxin-dependent mechanism in pin1,3,6;4;7;8. To address this question, we applied lanolin paste containing 1% of the natural auxin indole-3-acetic acid (IAA) to one side of developing leaves of WT and pin1,3,6;4;7;8, and recorded tissue response in mature leaves.
Consistent with previous reports (Scarpella et al., 2006; Sawchuk et al., 2007), IAA induced formation of extra veins in ~70% of WT leaves (27/38) (Fig. 9A,B), while ~30% of WT leaves (9/38) failed to respond to IAA application.

(A–F) Top right: genotype and treatment. Dark-field illumination of mature first leaves of WT (A,B) or pin1-1,3,6;4;7;8 (C–F) at side of application of lanolin paste (A,C) or lanolin paste containing 1% IAA (B,D–F). Bars: (A) 0.5 mm; (B–E) 0.25 mm; (F) 0.1 mm.
The effects of IAA on pin1,3,6;4;7;8 leaves were variable. In 40% of the leaves (28/70), IAA induced formation of extra veins (Fig. 9C,D). In ~60% of the leaves in which IAA induced formation of extra veins (17/28), IAA also induced tissue outgrowth of varied shape (Fig. 9E,F). In 30% of pin1,3,6;4;7;8 leaves (21/70), IAA induced tissue outgrowth but failed to induce formation of extra veins in the leaf; however, in nearly 80% of the pin1,3,6;4;7;8 leaves in which IAA induced tissue outgrowth [30/(17+21)=30/38], IAA also induced formation of vascular strands in the outgrowth (Fig. 9E,F). Finally, as in WT, 30% of pin1,3,6;4;7;8 leaves (21/70) failed to respond to IAA application in any noticeable way.
We conclude that pin1,3,6;4;7;8 leaves respond to vein-formation-inducing auxin signals and consequently that veins are formed by an auxin-dependent mechanism in the absence of PIN-mediated auxin transport.
Contribution of Auxin Signaling to Vein Patterning
Leaves of pin1,3,6;4;7;8 respond to vein-formation-inducing auxin signals (Figure 9), suggesting that the residual vein-patterning activity in those leaves may be provided by an auxin-dependent mechanism. We therefore asked what the contribution of auxin signaling to vein patterning were in the absence of PIN-dependent vein patterning activity.
To address this question, we used mutants in AUXIN-RESISTANT1 (AXR1), which lack a required post-translational modification of the auxin receptor complex (reviewed in (Calderon-Villalobos et al., 2010; Schwechheimer, 2018)); double mutants in TRANSPORT INHIBITOR RESPONSE1 (TIR1) and AUXIN SIGNALING F-BOX2 (AFB2), which lack the two auxin receptors that most contribute to auxin signaling (Dharmasiri et al., 2005); and phenylboronic acid (PBA), which inhibits auxin signaling (Matthes and Torres-Ruiz, 2016).
The embryos of axr1 and tir1;afb2 were viable (Table S13). In ~40–65% of the leaves of axr1, of tir1;afb2 and of WT grown in the presence of 10 μM PBA — as in leaves of weak gn alleles (Figure 2) — loops were open (Fig. 10A,B,H). Furthermore, in ~20–50% of the leaves of axr1, of tir1;afb2 and of WT grown in the presence of 10 μM PBA — again as in leaves of weak gn alleles (Figure 2) — veins were fragmented (Fig. 10A,B,H).

(A–G) Dark-field illumination of mature leaves illustrating phenotype classes (A–F, top right; G, bottom left): class a2 (axr1-3; A); class a4 (tir1;afb2; B); class b7 (NPA-grown WT; C); class b7/a6, wide midvein, more lateral-veins, dense network of thick veins and conspicuous marginal vein (NPA-grown axr1-12; D); class b8/a6, fused leaves with wide midvein, more lateral-veins, dense network of thick veins and conspicuous marginal vein (not shown); class a6 (E: PBA- and NPA-grown WT; F: NPA-grown tir1;afb2; G: tir1;afb2;pin1-1,3,6;4;7;8); inset in (G) illustrates cluster of seemingly randomly oriented vascular elements. (H) Percentages of leaves in phenotype classes (Classes 0, a2, a3, a4, a6, b7 and b8 defined in Figures 2 and 4). Difference between axr1-3 and WT, between axr1-12 and WT, between tir1;afb2 and WT, between PBA-grown WT and WT, between pin1-1,3,6;4;7;8 and WT, between NPA-grown WT and WT, between NPA-grown axr1-3 and NPA-grown WT, between NPA-grown axr1-12 and NPA-grown WT, between NPA-grown tir1;afb2 and NPA-grown WT, between PBA- and NPA-grown WT and NPA-grown WT, between axr1-3;pin1-1,3,6;4;7;8, and pin1-1,3,6;4;7;8 and between tir1;afb2;pin1-1,3,6;4;7;8 and pin1-1,3,6;4;7;8 was significant at P<0.001 (***) by Kruskal-Wallis and Mann-Whitney test with Bonferroni correction. Sample population sizes: WT, 47; axr1-3, 41; axr1-12, 41; tir1;afb2, 42; PBA-grown WT, 58; pin1-1,3,6;4;7;8, 63; NPA-grown WT, 48 (25 μM) or 146 (100 μM); NPA-grown axr1-3, 101; NPA-grown axr1-12, 103; NPA-grown tir1;afb2, 65; PBA- and NPA-grown WT, 105; axr1-3;pin1-1,3,6;4;7;8, 62; tir1;afb2;pin1-1,3,6;4;7;8, 75. Bars: (A,B) 1 mm; (C–E) 0.75 mm (F,G) 0.5 mmm.
We next asked whether PBA, mutation of AXR1 or simultaneous mutation of TIR1 and AFB2 enhanced the vein pattern defects induced by NPA, which phenocopies loss of PIN-dependent vein-patterning activity (Figure 5).
Approximately 3-25% of the leaves of NPA-grown axr1, NPA-grown tir1;afb2 and NPA- and PBA-grown WT resembled those of NPA-grown WT or of pin1,3,6;4;7;8 (Fig. lOC,H). However, ~25-50% of the leaves of NPA-grown axr1, NPA-grown tir1;afb2 and NPA- and PBA-grown WT resembled those of intermediate gn alleles: veins were thicker; the vein network was denser; and its outline was jagged because of narrow clusters of vascular elements that were oriented perpendicular to the leaf margin and that were laterally connected by veins or that, in the most severe cases, were aligned in seemingly random orientations (Figure 2; Fig. 10E,F,H; Fig. 10G, inset). Finally, ~20-60% of the leaves of NPA-grown axr1, NPA-grown tir1;afb2 and NPA- and PBA-grown WT had features intermediate between those of NPA-grown WT or of pin1,3,6;4;7;8 and those of intermediate gn alleles (Fig. 10D,H).
We next asked whether the spectrum of vein pattern defects of NPA-grown axr1 and tir1;afb2 were recapitulated by axr1;pin1,3,6;4;7;8 and tir1;afb2;pin1,3,6;4;7;8.
axr1;pin1,3,6;4;7;8 embryos were viable (Table S14) and developed into seedlings (Table S15) that resembled pin1,3,6;4;7;8 seedlings (Figure S9; Figure S10). Also tir1;afb2;pin1,3,6;4;7;8 embryos were viable (Table S14), but they developed into seedlings (Table S15) whose cotyledon pattern defects were more severe than those of pin1,3,6;4;7;8 seedlings (Figure S10; Figure S11) and whose root was replaced by a basal peg (Fig. S11C), as in strong gn alleles (Mayer et al., 1993) (Fig. S6B). Nevertheless, the spectrum of vein pattern defects of axr1;pin1,3,6;4;7;8 and tir1;afb2;pin1,3,6;4;7;8 was no different from that of NPA-grown axr1 and NPA-grown tir1;afb2 (Fig. 10C–H).
These observations suggest that the residual vein-patterning activity in pin1,3,6;4;7;8 is provided, at least in part, by AXR1- and TIR1/AFB2-mediated auxin signaling. Because reduction of AXR1- and TIR1/AFB2-mediated auxin signaling synthetically enhanced vein pattern defects resulting from loss of PIN-dependent vein-patterning function, we conclude that PIN-mediated auxin transport and AXR1- and TIR1/AFB2-mediated auxin signaling provide overlapping functions in vein patterning. Finally, the similarity between the vein pattern defects of NPA-grown axr1 and tir1;afb2, of NPA- and PBA-grown WT, and of axr1;pin1,3,6;4;7;8 and tir1;afb2;pin1,3,6;4;7;8, on the one hand, and those of intermediate gn alleles, on the other, suggests that the vein pattern defects of gn are caused by simultaneous defects in auxin transport and signaling.
Contribution of GN to Auxin Signaling
Were the vein pattern defects of gn not only the result of abnormal polarity or loss of PIN-mediated auxin transport but that of defects in auxin signaling, the vein pattern defects of gn might be associated with reduced auxin response, and the reduced auxin response of gn would be recapitulated by NPA-grown axr1; we asked whether that were so.
To address this question, we imaged expression of the auxin response reporter DR5rev::nYFP (Heisler et al., 2005; Sawchuk et al., 2013) in developing first-leaves of WT, pin1,3,6;4;7;8, NPA-grown WT, axr1, gn and NPA-grown axr1.
As previously shown (Sawchuk et al., 2013; Verna et al., 2015), strong DR5rev::nYFP expression was mainly associated with developing veins in WT (Fig. 11A). In pin1,3,6;4;7;8 and NPA-grown WT, DR5rev::nYFP expression was weaker and mainly confined to areas near the margin of the leaf (Fig. 11B–E). DR5rev::nYFP expression was weaker also in axr1 but was still associated with developing veins (Fig. 11F,G). Finally, in both gn and NPA-grown axr1, DR5rev::nYFP expression was much weaker and scattered across large areas of the leaf (Fig. 11H-K), suggesting that the vein pattern defects of gn are associated with reduced auxin response and that the reduced auxin response of gn is recapitulated by NPA-grown axr1.

(A–K) Confocal laser scanning microscopy; first leaves 4 (A,D,E), 5 (B,C,F–I) or 6 (J,K) days after germination. DR5rev::nYFP expression; look-up table (ramp in F) visualizes expression levels. Top right: genotype and treatment. Bottom left: reproducibility index. Dashed white line delineates leaf outline. Images in A,B,D,F,H,J were taken at identical settings. Images in A,C,E,G,I,K were taken by matching signal intensity to detector’s input range (~5% saturated pixels). Bars: (A–K) 100 μm.
Were the vein pattern defects of gn caused by simultaneous defects in auxin transport and signaling and did GN control auxin signaling as it controls auxin transport, the vein pattern defects of gn;axr1 should resemble those of gn, just as the vein pattern defects of gn;pin1,3;4;7 and gn;pin1,3,6;4;7;8 resemble those of gn; we tested whether that were so.
gn;axr1 embryos were viable (Table S16) and developed into seedlings (Table S17) that resembled gn seedlings (Figure S12; Figure S13), and the vein pattern defects of gn;axr1 were no different from those of gn (Fig. 12A–C), suggesting that the phenotype of gn is epistatic to that of axr1.

(A,B) Dark-field illumination of mature first leaves. Top right: genotype. (C) Percentages of leaves in phenotype classes (defined in Figures 2). Difference between axr1-3 and WT, between axr1-12 and WT, and between gn-13 and WT was significant at P<0.001 (***) by Kruskal-Wallis and Mann-Whitney test with Bonferroni correction. Sample population sizes: WT, 49; axr1-3, 42; axr1-12, 49; gn-13, 47; gn-13;axr1-3, 45; gn-13;axr1-12, 45. Bars: (A,B) 0.75 mm.
We conclude that the vein pattern defects of gn are caused by simultaneous defects in auxin transport and signaling and that GN controls both auxin signaling and auxin transport.
Contribution of Auxin Transport and Signaling to Coordination of Tissue Cell Polarity During Vein Formation
The vein pattern defects of gn are caused by simultaneous defects in auxin transport and signaling. We finally asked whether simultaneous defects in auxin transport and signaling recapitulated gn defects in coordination of tissue cell polarity.
To address this question, we imaged cellular localization of PIN1::PIN1:GFP expression during first-leaf development in WT, tir1;afb2, NPA-grown WT, NPA-grown tir1;afb2, and gnvan7.
Consistent with previous reports (Benkova et al., 2003; Reinhardt et al., 2003; Heisler et al., 2005; Scarpella et al., 2006; Wenzel et al., 2007; Bayer et al., 2009; Sawchuk et al., 2013; Marcos and Berleth, 2014; Verna et al., 2015), and as shown above (Fig. 1P,T), in the cells of the second loop at early stages of its development in WT leaves, PIN1::PIN1:GFP expression was mainly localized to the basal side of the PM, toward the midvein; in the inner cells flanking the developing loop, PIN1::PIN1:GFP expression was mainly localized to the side of the PM facing the developing loop; and in the inner cells further away from the developing loop, PIN1::PIN1:GFP expression was localized isotropically, or nearly so, at the PM (Fig. 13B). At later stages of second-loop development, by which time PIN1::PIN1:GFP expression had become restricted to the sole, elongated cells of the developing loop, PIN1::PIN1:GFP expression was localized to the basal side of the PM, toward the midvein (Fig. 13H). We observed a similar pattern of localization of PIN1::PIN1:GFP expression in tir1;afb2, but in this background stages of second-loop development comparable to those in WT appeared at later stages of leaf development, and nearly 70% (24/35) of second loops failed to connect to the first loop (Fig. 13C,I).
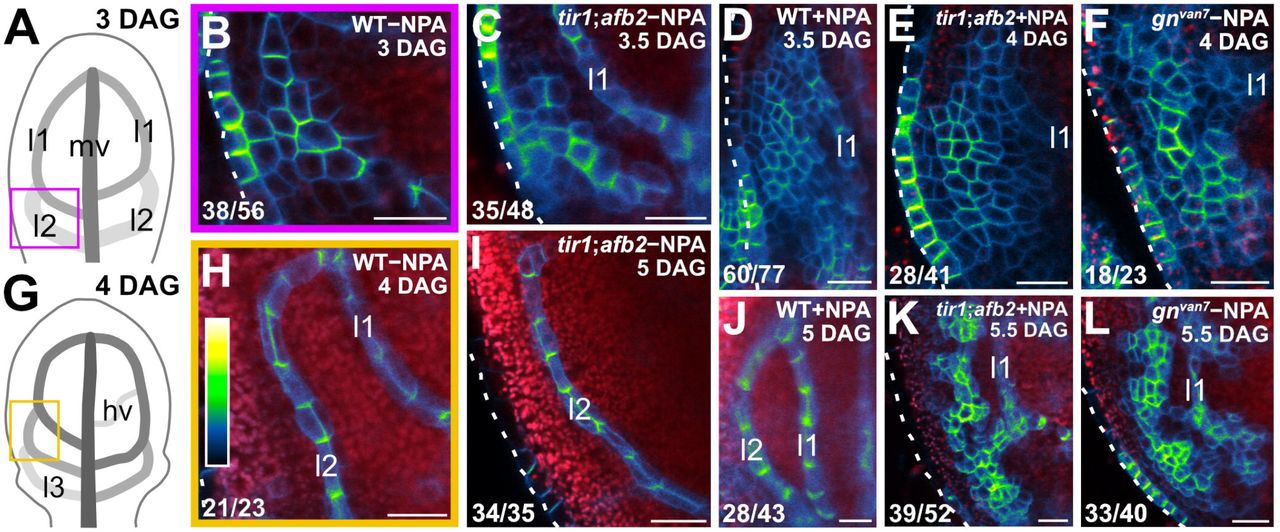
(A,G) Increasingly darker grays depict progressively later stages of vein development Boxes illustrate positions of closeups in B and H, respectively, hv: minor vein; 11, 12 and 13: first, second and third loops; mv: midvein. (B–F,H–L) Confocal laser scanning microscopy. First leaves. Top right: genotype, treatment and leaf age in days after germination (DAG). Dashed white line delineates leaf outline. Bottom left: reproducibility index. PIN1::PIN1:GFP expression; look-up table (ramp in H) visualizes expression levels. Red: autofluorescence. Bars: (B–F,H–L) 20 μm.
Consistent with previous reports (Scarpella et al., 2006; Wenzel et al., 2007), PIN1::PIN1:GFP expression domains were broader at early stages of development of the tissue that in NPA-grown WT corresponds to that from which the second loop forms in WT; PIN1::PIN1:GFP expression was localized isotropically, or nearly so, at the PM in the outermost inner cells but was mainly localized to the basal side of the PM in the innermost inner cells (Fig. 13D). At later stages of second-loop development in NPA-grown WT, by which time PIN1::PIN1:GFP expression had become restricted to the sole, elongated cells of the developing loop, PIN1::PIN1:GFP expression was localized to the basal side of the PM (Fig. 13J).
As in NPA-grown WT, in both gnvan7 and NPA-grown tir1;afb2 PIN1::PIN1:GFP expression domains were broader at early stages of development of the tissue that corresponds to that from which the second loop forms in WT, but PIN1::PIN1:GFP was expressed more heterogeneously in gnvan7 and NPA-grown tir1;afb2 than in NPA-grown WT (Fig. 13E,F). Nevertheless, as in NPA-grown WT, in both gnvan7 and NPA-grown tir1;afb2 PIN1::PIN1:GFP expression remained localized isotropically, or nearly so, at the PM, except in cells near the edge of higher-expression domains; in those cells, localization of PIN1::PIN1:GFP expression atthe PM was weakly polar, but such weak cell polarities pointed in seemingly random directions (Fig. 13E,F). At later stages of second-loop development of both gnvan7 and NPA-grown tir1;afb2, heterogeneity of PIN1::PIN1:GFP expression had become more pronounced, and PIN1::PIN1:GFP expression had become restricted to narrow clusters of cells; in those cells, localization of PIN1::PIN1:GFP expression at the PM was weakly polar, but such weak cell polarities still pointed in seemingly random directions (Fig. 13K,L).
In conclusion, simultaneous defects in auxin transport and signaling recapitulate gn defects in coordination of PIN1 polar localization, suggesting not only that the vein pattern defects of gn are caused by simultaneous defects in auxin transport and signaling, but that simultaneous defects in auxin transport and signaling recapitulate gn defects in coordination of tissue cell polarity during vein formation.
Discussion
The current hypothesis of how auxin coordinates tissue cell polarity to induce polar-veinformation proposes that GN controls the cellular localization of PIN1 and other PIN proteins; the resulting cell-to-cell, polar transport of auxin would coordinate tissue cell polarity and control polar developmental processes such as vein formation (reviewed in, e.g., (Berleth et al., 2000; Richter et al., 2010; Nakamura et al., 2012; Linh et al., 2018)).
Contrary to predictions of the hypothesis, we find that auxin-induced polar-veinformation occurs in the absence of PIN proteins or any known intercellular auxin transporter, that the residual auxin-transport-independent vein-patterning activity relies on auxin signaling, and that a GN-dependent signal that coordinates tissue cell polarity to induce polar-vein-formation acts upstream of both auxin transport and signaling (Fig. S14).
Control of Vein Patterning by Carrier-Mediated Polar Auxin-Transport
Overwhelming experimental evidence places polar auxin transport at the core of the mechanism that defines sites of vein formation (reviewed in (Sachs, 1981; Sachs, 1991a; Berleth et al., 2000; Sachs, 2000; Sawchuk and Scarpella, 2013)). The polarity of auxin transport is determined by the asymmetric localization of efflux carriers of the PIN family at the PM of auxin-transporting cells (Wisniewska et al., 2006). Therefore, loss of function of all the PM-PIN proteins should lead to loss of reproducible vein-pattern features or even, in the most extreme case, to the inability to form veins. Neither prediction is, however, supported by evidence: mutants in all the PM-PIN genes with vein patterning function — PIN1, PIN3, PIN4 and PIN7 — or in all the PM-PIN genes — PIN1–PIN4 and PIN7 — form veins, and these veins are arranged in reproducible, albeit abnormal, patterns. The most parsimonious account for the discrepancy between the observed and expected mutant defects is that vein patterning is controlled by additional, PM-PIN-independent auxin-transport pathways.
The existence of PM-PIN-independent auxin-transport pathways with vein patterning function can also be inferred from the discrepancy between the vein pattern defects of pin1,3;4;7, or of pin1,3;2;4;7, and those induced by NPA, which is thought to be a specific inhibitor of carrier-mediated cellular auxin-efflux (Cande and Ray, 1976; Sussman and Goldsmith, 1981; Petrasek et al., 2003; Dhonukshe et al., 2008). The vein pattern defects of WT grown in the presence of NPA are more severe than those of pin1,3;4;7 or pin1,3;2;4;7, suggesting the existence of an NPA-sensitive auxin-transport pathway with vein patterning function in addition to that controlled by PM-PIN proteins, a suggestion that is supported by the observation that growth in the presence of NPA enhances the vein pattern defects of pin1,3;4;7 to match those induced in WT by NPA.
Such PM-PIN-independent NPA-sensitive auxin-transport pathway with vein patterning function depends on the activity of the ER-PIN proteins PIN6 and PIN8, as inferred from the identity of the vein pattern defects induced in WT by NPA and those of pin1,3,6;4;7;8, and from the inability of NPA to induce further defects in pin1,3,6;4;7;8. Moreover, that NPA-grown WT phenocopies pin1,3,6;4;7;8, that no further defects can be induced in pin1,3,6;4;7;8 by NPA, and that the vein patterns of pin1,3,6;4;7;8 and NPA-grown WT fall into the same single phenotype-class suggest the absence of NPA-sensitive vein-patterning activity beyond that provided by PIN1, PIN3, PIN4, PIN6, PIN7 and PIN8, and hence the existence of NPA-insensitive vein-patterning pathways. It is of course possible that PIN6 and PIN8 are partially localized to the PM, and PM-localization of PIN5 and PIN6 has indeed been reported (Ganguly et al., 2014; Bennett et al., 2016; Simon et al., 2016; Ditengou et al., 2018); most important, however, that observation would not argue against the existence of NPA-insensitive vein patterning pathways, which is a logical conclusion, not a hypothesis.
These NPA-insensitive vein-patterning pathways are unlikely to be mediated by known intercellular auxin transporters — the AUX1/LAX influx carriers (Yang et al., 2006; Swarup et al., 2008; Peret et al., 2012) and the ABCB efflux carriers (Geisler et al., 2005; Bouchard et al., 2006; Petrasek et al., 2006) — as their mutation fails to enhance the vein pattern defects of pin1,3,6 and of the NPA-induced phenocopy of pin1,3,6;4;7;8. Though it remains unexplored whether the NPA-insensitive vein-patterning pathways depend on the function of the PIN-LIKES intracellular auxin-transporters (Barbez et al., 2012), and though we cannot rule out the existence of unknown auxin transporters, it is unlikely that the NPA-insensitive vein-patterning pathways depend on NPA-insensitive carrier-mediated auxintransport because as little as 10 μM NPA (a fraction of the concentration we used) is sufficient to inhibit carrier-mediated polar auxin-transport completely in tissue segments (Okada et al., 1991; Kaneda et al., 2011). Whatever the molecular nature of the NPA-insensitive vein-patterning pathways, they do contribute to the polar propagation of the inductive auxin signal: application of auxin to pin1,3,6;4;7;8 leaves, just as to WT leaves, induces the formation of veins that connect the applied auxin to the pre-existing vasculature basal to the site of auxin application.
Control of Vein Patterning by Auxin Signaling
The residual NPA-insensitive auxin-dependent vein-patterning activity of pin1,3,6;4;7;8 relies, at least in part, on the signal transduction mediated by the TIR1/AFB auxin receptors and their post-translational regulator AXR1. Loss of AXR1 or of TIR1 and AFB2, the two auxin receptors that most contribute to auxin signaling (Dharmasiri et al., 2005), or growth in the presence of the auxin signaling inhibitor PBA (Matthes and Torres-Ruiz, 2016), induces entirely new vein-pattern defects in pin1,3,6;4;7;8 or in its NPA-induced phenocopy, defects never observed in pin1,3,6;4;7;8 or NPA-grown WT: in the more-severely affected leaves of axr1;pin1,3,6;4;7;8, tir1;afb2;pin1,3,6;4;7;8, NPA-grown axr1, NPA-grown tir1;afb2 and NPA- and PBA-grown WT, the end-to-end alignment of vascular elements oriented with their axis along the axis of the vein is often replaced by the clustered differentiation of abnormally oriented vascular elements. Not only are these defects never observed in pin1,3,6;4;7;8 or NPA-grown WT, but they are more severe than the predicted sum of the defects of pin1,3,6;4;7;8 or NPA-grown WT, on the one hand, and of axr1, tir1;afb2 or PBA-grown WT, on the other. These observations are particularly interesting because genetic analysis of auxin signaling components has so far implicated auxin signaling only in the differentiation of normally patterned veins (Przemeck et al., 1996; Hardtke and Berleth, 1998; Hardtke et al., 2004; Alonso-Peral et al., 2006; Candela et al., 2007; Esteve-Bruna et al., 2013). Instead, the mutual synthetic enhancement between the vein pattern defects caused by reduced auxin signaling and those caused by reduced auxin transport suggests non-homologous redundancy of auxin signaling and auxin transport in vein patterning, a conclusion which is consistent with observations in the shoot apical meristem (Schuetz et al., 2008). Unlike in the shoot apical meristem, however, in the leaf such redundancy is unequal: whereas auxin transport is required for vein patterning even in the presence of normal auxin signaling, the vein patterning activity of auxin signaling is only exposed in conditions of compromised auxin transport.
How auxin signaling, inherently non-directional (Leyser, 2018), could contribute to the polar propagation of the inductive auxin signal in the absence of carrier-mediated polar auxin-transport is unclear. One possibility is that auxin signaling promotes the passive diffusion of auxin through the tissue by controlling, for example, the proton gradient across the PM (Fendrych et al., 2016). However, it is difficult to conceive how auxin diffusion through a specific side of the PM would positively feed back on the ability of auxin to diffuse through that specific side of the PM — a positive feedback that would be required to drain neighboring cells from auxin and thus to form veins, i.e. channels of preferential auxin movement (Sachs, 1969).
One other possibility is that auxin signaling promotes the facilitated diffusion of auxin through the plasmodesmata intercellular channels, a possibility that had previously been suggested (Mitchison, 1980) and that has recently received some experimental support (Han et al., 2014). Here, how auxin movement through a specific side of the PM could positively feed back on the ability of the cell to move auxin through that specific side of the PM is conceivable (e.g., (Cieslak et al., 2015)), but no experimental evidence exists of such feedback or that auxin movement through plasmodesmata controls vein patterning.
Yet another possibility is that auxin signaling activates an unknown mobile signal. Such signal need not be chemical and alternatives, for example a mechanical signal, have been suggested (Couder et al., 2002; Laguna et al., 2008; Corson et al., 2009; Lee et al., 2014) and have been implicated in other auxin-driven processes (e.g., (Hamant et al., 2008; Heisler et al., 2010; Peaucelle et al., 2011; Nakayama et al., 2012; Braybrook and Peaucelle, 2013)). However, whether a mechanical signal controls vein patterning remains to be tested.
A Tissue-Cell-Polarizing Signal Upstream of Auxin Transport and Signaling
The vein pattern defects of leaves in which both transport and transduction of the auxin signal are compromised are never observed in leaves in which either process is; yet those defects are not unprecedented: they are observed — though in more extreme form — in leaves of gn mutants, suggesting that GN controls both transport and transduction of the auxin signal during vein patterning.
That GN controls PM-PIN-mediated auxin transport during vein patterning is also suggested by the very limited or altogether missing restriction of PIN1 expression domains and coordination of PIN1 polar localization during gn leaf development, which is consistent with observations in embryos and roots (Steinmann et al., 1999; Kleine-Vehn et al., 2008). However, if failure to coordinate the polarization of the localization of PIN1 — and possibly other PM-PIN proteins — were the sole cause of the vein pattern defects of gn, these defects would be dependent on PM-PINfunction and would therefore be masked by those of pin1,3;4;7 in the gn;pin1,3;4;7 mutant. The epistasis of the vein pattern defects of gn to those of pin1,3;4;7 instead suggests that the vein pattern defects of gn are independent of PM-PIN function, and therefore that they are not the sole result of loss or abnormal polarity of PM-PIN-mediated auxin transport and that GN-acts upstream of PM-PIN genes in vein patterning. Moreover, the epistasis of the vein pattern defects of gn to those of pin1,3,6;4;7;8 and the inability of NPA, which phenocopies the vein pattern defects of pin1,3,6;4;7;8, to induce additional defects in gn suggest that the vein pattern defects of gn are independent of all the PIN genes with vein patterning function, and therefore that those defects are not the sole result of loss or abnormal polarity of PIN-mediated auxin transport, and that GN acts upstream of all the PIN genes in vein patterning. Whereas mechanisms by which GN may control PM-PIN-mediated auxin transport have been suggested (e.g., (Richter et al., 2010; Luschnig and Vert, 2014; Naramoto et al., 2014)), it is unclear how GN could control auxin transport mediated by the ER-localized PIN6 and PIN8; it is possible, however, that such control is mediated by GN function in ER-Golgi trafficking (Richter et al., 2007; Teh and Moore, 2007; Nakano et al., 2009).
These observations suggest that the function of GN in coordination of tissue cell polarity and vein patterning entails more than the regulation of PIN-mediated auxin transport, a conclusion which is consistent with functions of GN that do not seem to be related to auxin transport or mediated by PIN genes (Shevell et al., 2000; Fischer et al., 2006; Irani et al., 2012; Nielsen et al., 2012; Moriwaki et al., 2014).
The auxin-transport-, PIN-independent functions of GN in coordination of tissue cell polarity and vein patterning are, at least in part, provided by TIR1/AFB2- and AXR1-mediated auxin signaling. This conclusion is suggested by the ability of simultaneous reduction in auxin transport and signaling to phenocopy defects in coordination of tissue cell polarity, auxin response and vein patterning of gn; it is also supported by the epistasis of the vein pattern defects of gn to those of axr1, which is consistent with genetic analysis placing GN upstream of auxin signaling in the formation of apical-basal polarity in the embryo (Mayer et al., 1993).
Though it is unclear how GN controls auxin signaling during vein patterning, the most parsimonious account is that GN controls the coordinated localization of proteins produced in response to auxin signaling. Auxin signaling has indeed been shown to control the production of proteins that are polarly localized at the plasma membrane of root cells (e.g., (Scacchi et al., 2009; Scacchi et al., 2010; Yoshida et al., 2019)), and at least some of these proteins act synergistically with PIN-mediated auxin transport in the root (e.g., (Marhava et al., 2018)); however, it remains to be tested whether such proteins have vein patterning activity, whether their localization is controlled by GN, and whether they mediate GN function in auxin signaling during vein patterning.
Alternatively, because cell wall composition and properties are abnormal in gn (Shevell et al., 2000), GN could control the production, propagation or interpretation of a mechanical signal that has been proposed to be upstream of both auxin signaling and transport in the shoot apical meristem (Heisler et al., 2010; Nakayama et al., 2012); however, whether a mechanical signal controls vein patterning and whether such signal acts downstream of GN remains to be tested.
Irrespective of the mechanism of action, our results reveal synergism between auxin transport and signaling, and their unsuspected control by GN, in the coordination of tissue cell polarity during vein patterning, a control whose logic is unprecedented in multicellular organisms.
Materials & Methods
Notation
In agreement with (Crittenden et al., 1996), linked genes or mutations (<2,500 kb apart, which in Arabidopsis corresponds, on average, to ~10 cM (Lukowitz et al., 2000)) are separated by a comma, unlinked ones by a semicolon and homologous chromosomes by a slash.
Plants
Origin and nature of lines, genotyping strategies and oligonucleotide sequences are in Tables 1, 18 and 19. Seeds were sterilized and sown as in (Sawchuk et al., 2008). Stratified seeds were germinated and seedlings were grown at 22°C under continuous fluorescent light (~80 μmol m-2 s-1). Plants were grown at 25°C under fluorescent light (~110 μmol m-2 s-1) in a 16-h-light/8-h-dark cycle. Plants were transformed and representative lines were selected as in (Sawchuk et al., 2008).
Chemicals
N-1-naphthylphthalamic acid and phenylboronic acid were dissolved in dimethyl sulfoxide and water, respectively; dissolved chemicals were added to growth medium just before sowing. Indole-3-acetic acid was dissolved in melted (55°C) lanolin; the IAA-lanolin paste was applied to first leaves 4 days after germination and was reapplied weekly.
RT-PCR
Total RNA was extracted as in (Chomczynski and Sacchi, 1987) from 4-day-old seedlings grown as in (Odat et al., 2014). RT-PCR was performed as in (Odat et al., 2014) with the “GN_qFb” and “GN_qRb” oligonucleotides (Table S19), and with the “ROC1 F” and “ROC1 R” oligonucleotides (Beeckman et al., 2002) (Table S19).
Imaging
Developing leaves were mounted and imaged as in (Sawchuk et al., 2013), except that emission was collected from ~2.5-μm-thick optical slices. Light paths are in Table S20.
Mature leaves were fixed in 3 : 1 or 6 : 1 ethanol: acetic acid, rehydrated in 70% ethanol and water, cleared briefly (few seconds to few minutes) — when necessary — in 0.4 M sodium hydroxide, washed in water, mounted in 80% glycerol or in 1 : 2 : 8 or 1 : 3 : 8 water: glycerol: chloral hydrate and imaged as in (Odat et al., 2014). Grayscaled RGB color images were turned into 8-bit images, look-up-tables were applied, and brightness and contrast were adjusted by linear stretching of the histogram in the Fiji distribution (Schindelin et al., 2012) of ImageJ (Schneider et al., 2012; Schindelin et al., 2015; Rueden et al., 2017).
Acknowledgements
We thank the Arabidopsis Biological Resource Center for emb30-8, gn-13, pin1-1, eir1-1, pin6, pin8-1, pgp1-100, mdr1-101, ucu2-4, aux1-355, lax1-064, axr1-3 and axr1-12; Hidehiro Fukaki for fwr and gnSALK_103014; Sandra Richter and Gerd Jürgens for gnB/E and gnR5; Satoshi Naramoto and Hiroo Fukuda for van7/emb30-7; Eva Benková and Jiří Friml for PIN1::PIN1:GFP; Jian Xu and Ben Scheres for PIN1::PIN1:YFP and PIN2::PIN2:GFP; Jian Xu, Miyo Morita and Masao Tasaka for PIN3::PIN3:GFP; Ikram Blilou and Ben Scheres for Atpin1::En134, pin3-3, pin4-2 and pin7En; Venkatesan Sundaresan for toz-1, Thomas Berleth for mpG12; Markus Geisler and Jiří Friml for ABCB1::ABCB1:GFP and ABCB19::ABCB19:GFP; Cris Kuhlemeier for aux1-21;lax1;2-1;3; Michael Prigge and Mark Estelle for tir1-1;afb2-3; and Marcus Heisler and Elliot Meyerowitz for DR5rev::nYFP. This work was supported by Discovery Grants of the Natural Sciences and Engineering Research Council of Canada (NSERC) to E.S. C.V. was supported, in part, by a University of Alberta Doctoral Recruitment Scholarship. M.G.S was supported, in part, by an NSERC CGS-M Scholarship and an NSERC CGS-D Scholarship.
Footnotes
Word duplication in text eliminated; formatting of special characters in names of researchers acknowledged corrected -- with our apologies; incomplete references -- well... -- completed.





{kind=link}
{kind=link}
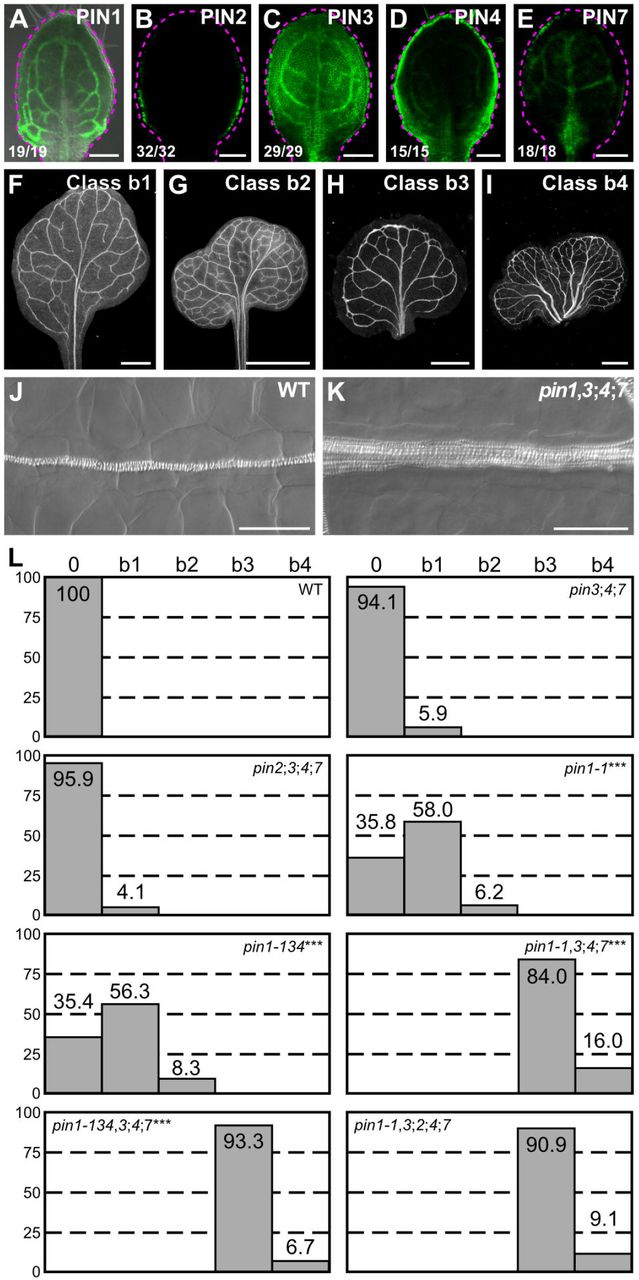{kind=link}
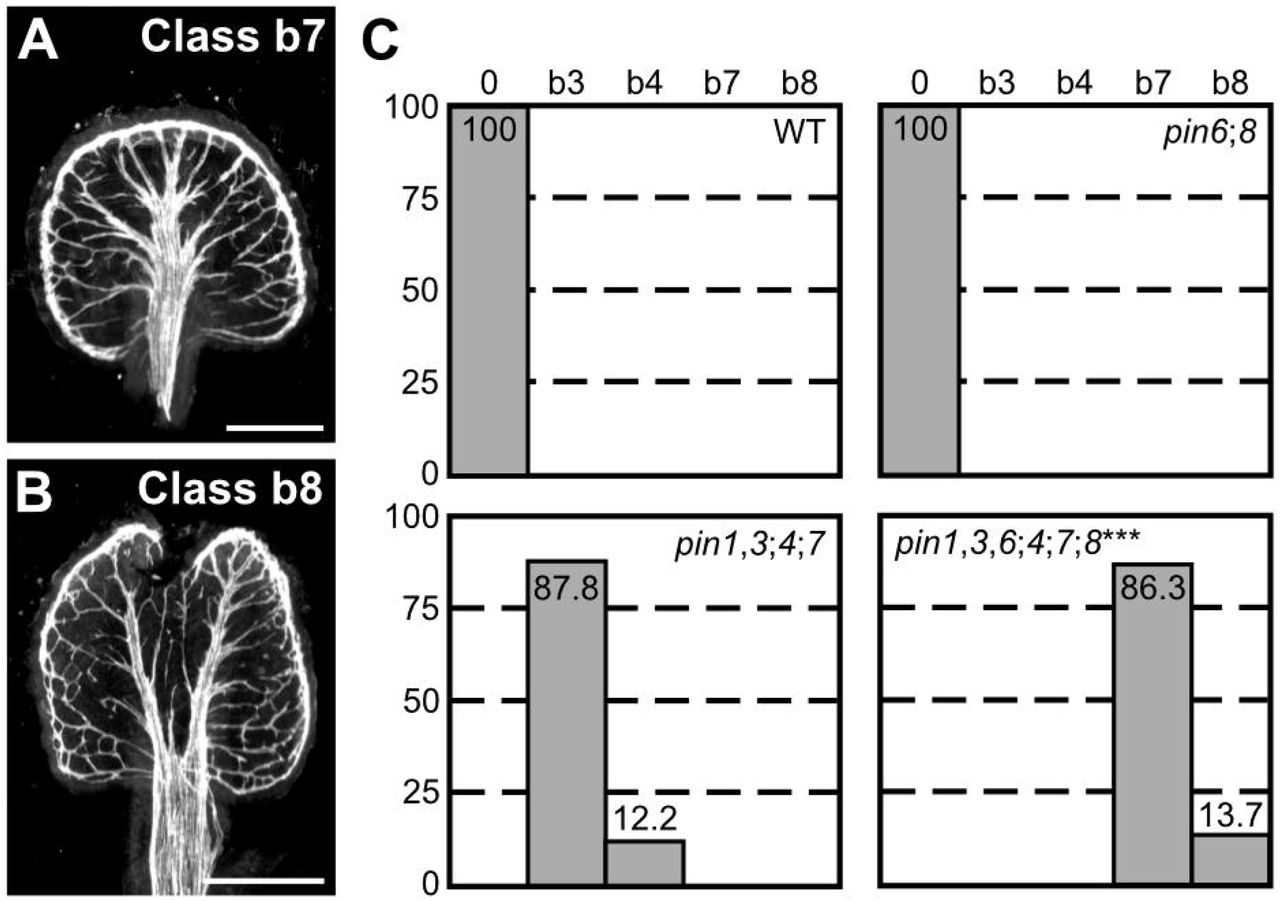{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}