

ABSTRACT
Multipotent adult stem cells must balance self-renewal with differentiation into various mature cell types. How this activity is molecularly regulated is poorly understood. By using genetic and molecular analyses in vivo, we show that a small network of basic Helix-Loop-Helix (bHLH) transcription factors controls both stemness and bi-potential differentiation in the Drosophila adult intestine. We find that homodimers of Daughterless (Da, homolog to mammalian E proteins) maintain the self-renewal of intestinal stem cells and antagonise the activity of heterodimers of Da and Scute (Sc, homolog to ASCL and known to promote intestinal secretory differentiation). We find a novel role for the HLH factor Extramacrochaetae (Emc, homolog to Id proteins), titrating Da and Sc to promote absorptive differentiation. We further show that Emc prevents committed absorptive progenitors from de-differentiating, revealing the plasticity of these cells. This mechanism of interaction partner-switching enables the active maintenance of stemness, but primes stem cells for differentiation along two alternative fates. Such regulatory logic could be recapitulated in other bipotent stem cell systems.
INTRODUCTION
The regulation of stem cell fate decisions hinges on the control of transcription. Central to this control are gene regulatory networks, whose activity and dynamics steer cells along particular differentiation pathways (Graf & Enver, 2009; Levine & Davidson, 2005; Moris et al, 2016). These pathways can often be construed as a succession of binary steps that are regulated by the cross antagonism of transcription factor pairs (Graf & Enver, 2009; Simon et al, 2018). However, for multipotent stem cells, several fate options may be simultaneously available through higher-dimensional switches (Cinquin & Demongeot, 2002; 2005). To understand the choice between several alternative fates, knowledge of the interactions between transcriptional regulators is essential.
Intestinal stem cells (ISCs) are a clear example of multipotency in adult tissues. In Drosophila and mammals, ISCs face a triple choice between self-renewal and differentiation into one of two mature lineages: secretory or absorptive (reviewed in Jiang & Edgar, 2012; Crosnier et al, 2006; Philpott & Winton, 2014). The secretory lineage comprises several cell types in vertebrates, but only enteroendocrine (EE) cells in Drosophila. Absorptive cells are enterocytes (ECs). In Drosophila, ISCs produce lineage-specific precursors through distinct molecular triggers. High Notch signalling induces formation of enteroblasts (EBs), which will give rise to ECs. Expression of the bHLH (basic Helix-Loop-Helix) transcription factors Scute (Sc) or Asense (Ase), members of the achaete-scute Complex (AS-C, homologs of ASCL mammalian genes) induces the formation of EE precursor cells (pre-EEs) and their transition to mature EE cells (Micchelli & Perrimon, 2006; Ohlstein & Spradling, 2006; Bardin et al, 2010; Zeng & Hou, 2015; Chen et al, 2018). This argues for ISCs facing a multiple choice, rather than a sequence of binary decisions. However, very little is known about the molecular mechanisms that can simultaneously regulate both self-renewal and bi-potential differentiation.
Transcription factors of the bHLH family control fate in multiple developmental contexts, from sex determination and mesoderm specification, to neurogenesis and immune cell type specification (reviewed in Baylies et al, 1997; Murre et al, 1994; Murre, 2005; Bertrand et al, 2002; Ohtsuka & Kageyama, 2010). Their HLH motif mediates dimerization, while the preceding region, rich in basic amino acids, allows DNA binding (Massari & Murre, 2000). bHLH factors are subdivided into several classes, with class I (E proteins) comprising factors such as E2A, E2.2 and HEB (encoded by TCF3/4/12; a single gene, daughterless (da), in Drosophila), usually with broad expression patterns; and class II containing more restrictedly expressed factors such as MYOD, TWIST and ASCL (MyoD, Twi, and AS-C proteins in Drosophila). Generally, class II factors heterodimerise with a class I member to bind DNA, while class I factors can either homo- or heterodimerise (Murre et al, 1994). The class V comprises HLH factors lacking the basic domain, known as Inhibitors of DNA binding (Id proteins; a single gene, extra macrochaetae (emc), in Drosophila), as heterodimerisation with them renders dimers unable to bind DNA (Murre et al, 1994). Combinatorial, antagonistic or cooperative interactions confer unique properties to regulatory networks based on these factors (Amoutzias et al, 2008), potentially including multi-stable dynamical behaviour (Cinquin & Page, 2007; Cinquin & Demongeot, 2005).
Class II bHLH factors are known to regulate differentiation in the metazoan intestine (reviewed in Philpott & Winton, 2014; Hartenstein et al, 2017). In Drosophila, Sc and Ase can initiate EE differentiation (Bardin et al, 2010; Amcheslavsky et al, 2014; Chen et al, 2018; Guo & Ohlstein, 2015), while other bHLH genes maintain EE function (dimmed, homolog of NeuroD; Beebe et al, 2015), or promote their functional diversity (tap, homolog of Neurogenins; Hartenstein et al, 2017). On the other hand, Da is required for ISC maintenance, as ISCs mutant for da differentiate (Bardin et al, 2010; Lan et al, 2018). However, which interaction partner binds to Da to maintain stemness is not known, and how different bHLH factors dimerise to allow differentiation has not been explored. Here we identify the Da homodimer as the critical bHLH complex maintaining ISC self-renewal, elucidate the role of the HLH factor Emc in titrating Da and Sc to promote absorptive differentiation, and find a functional antagonism between Da:Da and Da:Sc dimers. Our results suggest that a network of bHLH factors acts as a three-way switch to regulate self-renewal and bipotential differentiation in the adult fly gut.
RESULTS
emc is expressed preferentially in the absorptive lineage
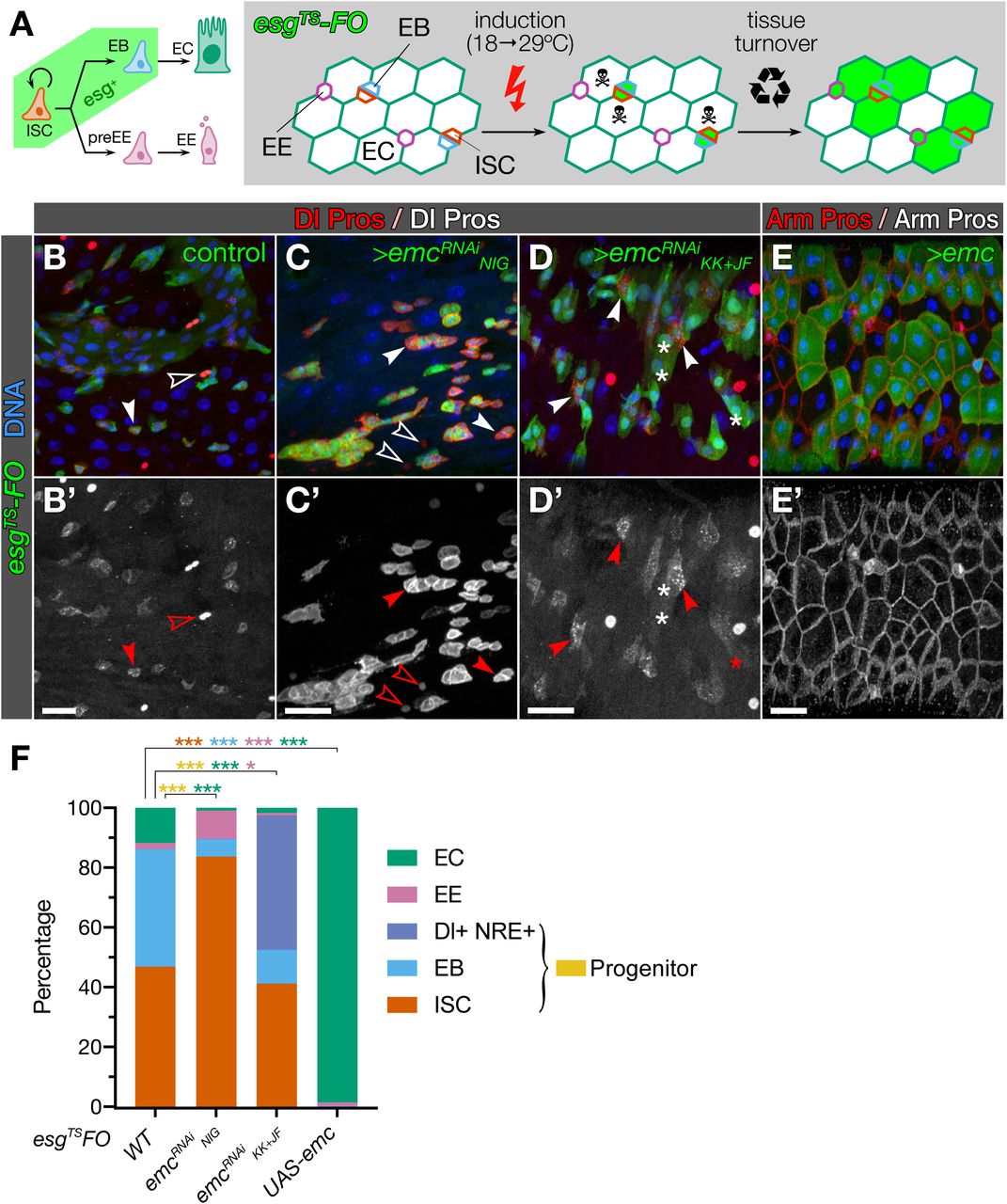
bHLH factors are essential for correct differentiation during adult intestinal homeostasis across metazoans (Philpott & Winton, 2014; Hartenstein et al, 2017). This suggests that emc, the only Drosophila Id factor (Garrell & Modolell, 1990; Campuzano, 2001), may have a regulatory function in the adult fly intestine, where it is strongly transcribed (Chintapalli et al, 2007; Brown et al, 2014; Dutta et al, 2015). We examined its expression pattern using the homozygous viable protein trap line emcCPTI-002740 (Lowe et al, 2014) and detected Emc in all EBs and most ECs in the posterior midgut (Fig 1A-B). Some EEs and ISCs showed very low levels of expression (Fig 1C-D). This expression pattern, with the described role of several bHLH proteins in intestinal homeostasis, suggested a function of emc in promoting EC differentiation.

A-D. Projected confocal stacks showing endogenous Emc:GFP expression. Emc is expressed in EBs (marked with NRE-lacZ, arrowheads in A, A’) and most ECs (positive for myoIA-lacZ, solid arrowheads in B, B’) but not all (empty arrowhead in B, B’). emc expression can also be detected in some ISCs (Dl+) and EEs (Pros+) (solid arrowheads and asterisks, respectively, in C-D’) but not all (empty arrowheads and asterisks in C-D’).
E-I. Differentiation is impaired in emc mutant clones. Control clones display more polyploid Dl−/Pros− ECs (asterisks in E, H) than clones homozygous of for the mutant alleles emcAP6 (F), emc1 (G) and emcLL02590 (I), which are enriched in Dl+ ISCs (solid arrowheads) and diploid Dl−/Pros− EBs (empty arrowheads) with respect to controls.
J. Stacked bars plots summarising the cell type composition of clones by genotype. Numbers (cells/clones) are: 194/106, 213/118, 310/117, 112/95 and 208/200 for control FRT2A, emcLL02590, control FRT80B, emc1, and emcAP6 respectively. p-values are <0.05, <0.01 and <0.001 for one, two or three asterisks (see Tables S1 and S2 for further details).
Scale bar: 20µm in all panels.
Emc is required for ISC differentiation
We examined the effect of emc loss in the adult posterior midgut by generating emc mutant MARCM clones (Lee & Luo, 1999). Clones of the null allele emcAP6 (Ellis, 1994; Fig S1A) were depleted in differentiated cells and enriched in ISCs, as revealed by expression of the Notch ligand, Delta (Dl) (Ohlstein & Spradling, 2007) (Fig 1E-F, J and Table S1). Clones for the strong hypomorphic allele emc1 and the predicted transcriptional null emcLL02590 (Fig S1A) showed similar enrichment in Dl+ ISCs respect to wild-type clones (Fig 1E, G-J). We then knocked-down emc in ISCs and EBs, and traced their offspring using the escargot flip-out (FO) approach (“esgTS-FO”; Jiang et al, 2009; Fig 2A). Loss of emc blocked the differentiation of ISCs and EBs, and sometimes the typically small nests of 1-3 esg+ cells (Ohlstein & Spradling, 2006; de Navascués et al, 2012) expanded into larger clusters (Fig 2C). This was suggestive of impairment of Notch signalling. However, these larger clusters did not display the unchecked growth and invasive behaviour observed when Notch signalling is deregulated in this tissue (Micchelli & Perrimon, 2006; Patel et al, 2015; see also Fig 4A). Moreover, knockdown of emc in the ISC/EB compartment led to the loss of many esg+ cell nests (identified by their expression of Hdc, an RNA binding protein expressed like esg; Resende et al, 2017; Fig S1B) (Fig S1C, compare with Fig 2B). This was due to apoptosis, since co-expression of the caspase inhibitor p35 and emc-specific RNAi prevents the loss of esg+ cells (Fig S1D). We conclude that emc is necessary to allow differentiation of ISCs and EBs, as well as for their survival.

A. The esgTS-FO method allows the temporal control of expression in progenitor cells (esg+ ISCs and EBs) and their lineage tracing to access their fate potential. UAS-GFP and UAS-FLP are expressed specifically in ISCs and EBs after induction by temperature switch. This leads to the progenitor-restricted expression of the (potentially) ubiquitous Actin5C-Gal4 by excision of a transcriptional terminator. Actin5C-Gal4 maintains UAS-GFP expression in the ISC/EB lineage when differentiation occurs and esg expression stops.
B-E. Emc is required for differentiation and sufficient for induce the EC fate. ISCs and EBs in control guts produce both EC (Dl−/Pros−) and EE (Dl−/Pros+) GFP+ cells (B, note the difference in intensity levels between Dl and Pros, respectively solid and empty arrowheads), while differentiation is reduced when ISCs/EBs express emc-specific RNAi, shown by increased number of Dl+/GFP+ cells (C, D). Reduction of emc also induces increased Dl expression: compare Dl-expressing with Pros-expressing cells (respectively solid and empty arrowheads in C, D, compare with those similarly marked in B). Forced expression of emc in ISCs/EBs forces differentiation into ECs (E).
F. Stacked bars plots summarising the cell type composition of esgTS-FO tissue. Numbers (total GFP+ cells / fields of view) are 1046/15, 104/21, 832/15 and 812/10 for control, UAS-emcRNAiNIG, UAS-emcRNAiKK+HMS, and UAS-emc, respectively. p-values are <0.05, <0.01 and <0.001 for one, two or three asterisks (see Tables S1 and S2).
Scale bar: 20µm in all panels.
emc maintains the commitment of EBs
The increased number of Dl+ cells in emc depletion conditions conflicted with emc expression being predominantly in EBs and ECs. To test whether the loss of emc in EBs could induce their expression of Dl, we expressed RNAi against emc specifically in EBs using NRE-Gal4, UAS-GFP, tub-Gal80TS (Zeng et al, 2010). Indeed, emc knockdown led to EBs reacquiring Dl expression, often while displaying reduced GFP signal (Fig 3A-B). This suggested reduced expression of the EB-specific NRE-Gal4 and that emc-depleted EBs were reverting their identity. To verify this, we used a flip-out lineage tracing approach to irreversibly label EBs (“NRETS-FO”; Fig 3C) and stained for the mitotic marker phospho-Histone3. In contrast with unmodified EBs, which are post-mitotic and only produce ECs (Zeng & Hou, 2015; Micchelli & Perrimon, 2006; Ohlstein & Spradling, 2006), emc-depleted EBs undergo cell division and generate Dl+, proliferative cells (Fig 3D-E). This suggests that emc-deficient EBs can not only activate Dl expression but also de-differentiate into functional ISCs, capable of cell division and EE differentiation.

A-B. The NRE-Gal4 driver is expressed in EBs at different levels in normal intestines (solid and empty arrowheads in A for higher and lower-expressing EBs), but never co-express Dl. When emc is depleted with NRE-Gal4, many EBs co-express Dl (empty arrowheads, B) but not all (solid arrowheads, B). Also, EEs develop close to emc-depleted EBs (arrow, B).
C. Use of NRETS-FO allows the simultaneous manipulation and tracing of the EB lineage. NRE-Gal4, UAS-GFP is expressed specifically in EBs after induction and activates specifically in EBs the (potentially) ubiquitous Actin5C-Gal4, which in turn will maintain UAS-GFP expression if differentiation or decommitment occur.
D-E. Loss of emc in EBs leads to their decommitment. Activation of NRETS-FO in control guts leads exclusively to the labelling of EBs and ECs, with no co-expression of GFP and Dl or Pros (solid and empty arrowheads, respectively, in D; some ISCs and EEs in the micrograph seem inside the GFP territory due to the Z-projection of the confocal stack). By contrast, in NRETS-FO guts where emc is depleted, EB-derived cells express Dl and are capable of mitosis, as judged by phospho-Histone 3 staining (arrowhead in E).
Scale bar: 20µm in all panels.
emc acts in parallel to Notch to direct EC differentiation
We next assessed the capacity of emc to impose differentiation by overexpressing it with esgTS-FO. Indeed, all ISCs and EBs differentiated into ECs (Fig 2E), as previously reported (Lan et al, 2018). This establishes Emc as both necessary and sufficient to force differentiation into the absorptive fate.
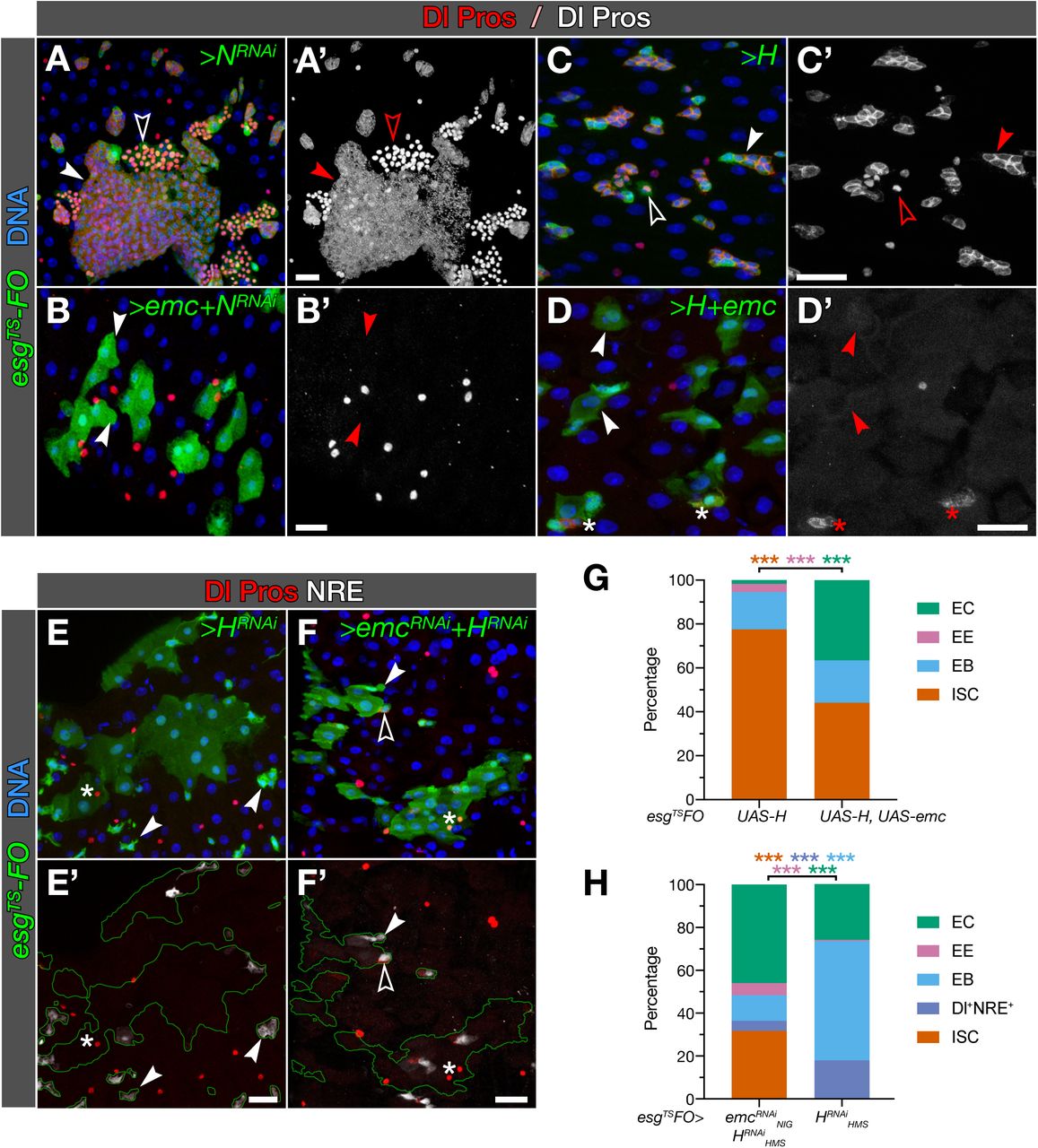
This phenotype coincides with that of over-activating the Notch pathway, a known regulator of ISC fate (Micchelli & Perrimon, 2006; Ohlstein & Spradling, 2006; Bardin et al, 2010). This suggests that emc might be mediating the effects of Notch signalling. To explore this possibility, we tested whether emc could force EC differentiation in the absence of Notch. Knockdown of Notch with esgTS-FO leads to an invasive expansion of Dl+ ISC-like and Pros+ EE-like cells (Micchelli & Perrimon, 2006; Patel et al, 2015; see Fig 4A). When we co-expressed emc and NRNAi together, all ISCs and EBs turned into ECs (Fig 4B), strongly suggesting that emc operates downstream of Notch. This phenotype, however, could also be due to the excess of Emc quickly driving cells irreversibly into differentiation, before the knockdown of Notch resulted in effective clearance of Notch protein. To reduce Notch signalling with the kinetics of protein synthesis, rather than the additive kinetics of RNAi biogenesis, RNA interference, and Notch decay, we overexpressed Hairless (H), the co-repressor associated to the nuclear effector of NICD, Su(H) (reviewed in Bray & Furriols, 2001), with esgTS-FO. As expected, this leads to expansion of Dl+ and Pros+ cells, with a strong reduction of EC differentiation (Fig 4C, G; see also Bardin et al, 2010). By contrast, co-expression of emc and H led to a sharp increase in the levels of EC differentiation (Fig 4D, G). This further indicates that Emc can induce differentiation in the absence of Notch signalling.

A-D. emc can direct EC differentiation in the absence of Notch signalling. EC differentiation is abolished or strongly impaired with knockdown of N (A) or overexpression of H (C), respectively, with accumulation of Dl+ or Pros+ cells (solid and empty arrowheads, respectively, in A, C). Simultaneous overexpression of emc induces EC differentiation in both cases (arrowheads in B, D), but to a lesser an extent in a background of H overexpression (asterisks in D, compare with B).
E-F. Notch signalling can direct EC differentiation in the absence of emc. Loss of EC differentiation caused by depletion of emc (see panel H and Fig. 2C-D, F) is rescued by simultaneous knockdown of H (F). Depletion of H can induce EC differentiation on its own (E).
G-H. Stacked bars plots summarising the cell type composition of esgTS-FO tissue. Numbers (total GFP+ cells / fields of view) are 1810/21 and 1292/27 for UAS-H and UAS-H+UAS-emc, respectively (G) and 984/15 and 126/16 for UAS-HRNAiHMS and UAS-HRNAiHMS+UAS-emcRNAiNIG, respectively (H). p-values are <0.05, <0.01 and <0.001 for one, two or three asterisks (see Tables S1 and S2).
Scale bar: 20µm in all panels.
Next, we tested whether emc mediates the effects of Notch signalling. Activating the pathway by knocking-down H leads to an increase in EC differentiation (Bardin et al, 2010; Fig 4E) and to the expression of the NRE reporter in all the progenitor, diploid cells, including those expressing Dl (Fig 4H); this is likely due to an increased baseline expression of the NRE reporter, as expected in the absence of H (Furriols & Bray, 2001). Simultaneously knocking down H and emc did not reduce the increase in EC differentiation induced by overexpression of H-specific RNAi, but actually enhanced it. (Fig 4F-H). Taken together, our data suggest that both Emc and Notch signalling can induce EC differentiation independently of each other.
Emc antagonises self-renewal by disrupting Daughterless homodimers
The molecular function of Emc consists in titrating transcriptional activators. Archetypal targets of this inactivation are the class I and II bHLH factors such as Da or Sc, respectively, to which Emc binds directly in vitro and in vivo (Van Doren et al, 1991; Cabrera et al, 1994; Alifragis et al, 1997; Giot et al, 2003; Shokri et al, 2019). Therefore, the intestinal phenotypes we observed with the loss of emc may be due to increased activity of Da and its dimerization partners. Indeed, overexpression of da with esgTS-FO recapitulated precisely the effect of emc knockdown in esg+ cell nests. Some of these nests (labelled by hdc expression) were expanded (Fig 5A-B, I), and many were lost, their survival rescued by co-expression of p35 (Fig 5B and S2C). Conversely, loss of da leads to EC differentiation (Fig S2A-B and 5I; as previously described (Bardin et al, 2010; Lan et al, 2018) and mimics the excess of Emc (Fig 2E). This suggests that the role of Emc is largely to repress one or more Da-containing bHLH dimers, which in turn maintain stemness and survival. Epistasis experiments further confirmed this, as the simultaneous loss of Emc and Da led to the differentiation of ISCs and EBs into ECs (Fig 5E-H).

A-D. Da homodimers maintain ISCs/EBs undifferentiated. Solid arrowheads: ISCs/EBs; empty arrowheads: ECs/EEs. Overexpression of Da with esgTS-FO prevents formation of ECs, though it allows EE differentiation (B, compare with A). Expression of a forced Da:Da dimer in esg+ cells prevents all differentiation, as evaluated with Hdc/NRE-lacZ (C) or Dl (D).
E-H. da is epistatic over emc. Solid arrowheads: ECs; empty arrowheads: Dl+ ISCs. MARCM clones mutant for da10 differentiate as ECs (E). This phenotype is recapitulated when these clones simultaneously express emc-specific RNAi (F). Likewise, impaired differentiation in emcLL02590 clones (G) is rescued by simultaneous knockdown of da, which leads to EC formation (H).
I. Stacked bars plots summarising the cell type composition of esgTS-FO tissue. Numbers (total GFP+ cells / fields of view) are 1135/15, 549/14, 693/13, 803/15 and 1086/19 for control, UAS-da, UAS-da:da, UAS-daRNAiJF and UAS-daRNAiHMS, respectively. P-values are <0.05, <0.01 and <0.001 for one, two or three asterisks (see Tables S1 and S2).
Scale bar: 20µm in all panels.
We sought to determine the identity of the Da partners involved in maintaining stemness. Most Drosophila class II bHLH factors (Moore et al, 2000) are expressed in the intestine at very low levels (Fig S2D); several of them are already known not to have an essential role in ISC maintenance (Bardin et al, 2010; Beebe et al, 2015; Hartenstein et al, 2017). However, Da can form homodimers (Cabrera et al, 1994; Cabrera & Alonso, 1991) with demonstrated or suspected functional relevance in several contexts, and often antagonised by Emc (Tapanes-Castillo & Baylies, 2004; Tanaka-Matakatsu et al, 2014; D’Rozario et al, 2016; Bhattacharya & Baker, 2011; Andrade-Zapata & Baonza, 2014; Troost et al, 2015). To test whether Da:Da dimers were involved in preventing differentiation, we overexpressed forced Da homodimers using the UAS-da:da tethered construct (Tanaka-Matakatsu et al, 2014; Neuhold & Wold, 1993) with esgTS-FO. This blocked the differentiation of ISCs and EBs, which accumulated in larger clusters (Fig 5C-D; Table S2). Expression of Da:Da did not lead to esg+ cell death (Fig 5D, compare nest density with 5B). This shows that Da homodimers prevent differentiation; it does not rule out that other Da-containing complexes are also necessary to maintain self-renewal. To test this, we took advantage of the UAS-daRNAi transgene P{TRiP.JF02488}; this targets the 5’UTR of the endogenous da transcript, absent in the UAS-da:da construct (Fig S2E). Thus, we co-expressed da:da and daJF02488 in ISCs and EBs to evaluate the capacity of Da:Da to prevent differentiation in the absence of endogenous, monomeric Da. The effects were identical to the overexpression of UAS-da:da alone (Fig S2F, compare with Fig 5D). This shows that Da can promote ISC self-renewal solely as homodimers, which are antagonised by Emc to allow EC differentiation. Furthermore, our results suggest that Da binds to an unknown partner, to form complexes that are titrated by Emc and whose excess promotes cell death in esg+ cells.
Basal levels of Sc in esg+ cells confer ISC properties
While the maintenance of self-renewal corresponds to Da homodimers, the loss of emc mimics best the overexpression of monomeric Da, which increases all Da-containing complexes. As emc loss induces the elevation of Dl levels, another Da partner, such as Scute, could be mediating this effect. Detectable levels of Sc have recently been found in esg+ cells (Chen et al, 2018; Doupé et al, 2018), and members of the AS-C are proposed to promote Dl expression during neurogenesis (Kunisch et al, 1994; Hinz et al, 1994). Therefore, we tested whether the increased levels of Dl observed upon emc loss (Figs 2C-D) could be caused by elevated activity of Sc or any of the other bHLH factors belonging to the AS-C. We induced emc knockdown in MARCM clones, which were either wild-type or homozygous for scB57, a deficiency that removes the entire AS-C, and compared their respective levels of Dl expression. Indeed, we observed an average four-fold decrease in Dl levels in emc-depleted clones when they were homozygous for scB57, indicating that basal Sc levels (and possibly other members of the AS-C) can boost Dl expression in ISCs (Fig 6A-C).

A-C. Emc antagonises Sc in inducing Dl expression. Loss of the AS-C using the scB57 deficiency reduces the increase in Dl expression observed with knockdown of emc (B, compare with A). Dl levels are quantified by comparison with those of Pros (see Appendix). Numbers (Dl+ cells/Pros+ cells) are 210/146 and 718/210 for UAS-emcRNAiNIG and UAS-emcRNAiNIG, Df(1)scB57, respectively. P-values are <0.05, <0.01 and <0.001 for one, two or three asterisks (Mann-Whitney test).
D-F. Antagonism/collaboration between Sc and Da. Solid arrowheads: Dl+/Pros+ pre-EEs; empty arrowheads: Dl+ ISCs; asterisks: Pros+ EEs. Overexpression of Sc leads to a dramatic increase of Dl+/Pros+ pre-EEs and Pros+ EEs, but maintains a population of Dl+ ISCs (D). The population of ISCs progressively increases by co-expression of monomeric Da (E) and dimeric Da:Da (F), at the cost of EE differentiation (E, F) and pre-EE formation (F). Quantified in H.
G. Da:Da antagonises EE formation. While reduction of Notch signalling leads to an excess of EEs, simultaneous overexpression of Da:Da and N-specific RNAi results in very few Pros+ cells (solid arrowheads, compare with Fig 4A).
H. Stacked bars plots summarising the cell type composition of esgTS-FO tissue. Numbers (total GFP+ cells / fields of view) are 1241/6, 2131/10 and not determined/11 for UAS-sc, UAS-sc, UAS-da, and UAS-sc, UAS-da:da, respectively. p-values are <0.05, <0.01 and <0.001 for one, two or three asterisks (see Tables S1 and S2).
I. Sc can induce decommitment of EBs. Expression of Sc with NRETS-FO leads to reacquisition of ISC properties (Dl expression, mitosis assessed with phospho-Histone 3, empty arrowhead) and pre-EE formation (Pros expression and mitosis, solid arrowhead).
Scale bar: 20µm in all panels.
These observations prompted us to consider whether Sc could be contributing to additional stem cell properties in esg+ cells. Expression of Sc with esgTS-FO clearly leads to increased differentiation into Pros+ EEs (Bardin et al, 2010; Chen et al, 2018) and Fig 6D, H). To uncover a role in inducing stem properties, we overexpressed Sc with the EB-specific driver NRETS-FO. Sc-expressing EBs were capable of division, and their offspring included both Pros+ and Dl+ expressing cells, indicating the acquisition of ISC-like properties (Fig 6I, compare with 3D). These functions of Sc depend on the formation of Sc:Da heterodimers, as the overexpression of Sc in da-depleted cells prevents the induction of EE differentiation and of Dl expression (Fig S3E).
Sc:Da and Da:Da antagonise each other in the ISC-to-EE transition
We have shown so far that Da homodimers maintain ISC self-renewal, with EC differentiation simply triggered by the Emc-mediated titration of Da. EE differentiation, however, requires the transition from the transcriptional program of Da:Da to that of Da:Sc. Two main scenarios are possible. Sc:Da could initiate a program that silenced, or was epistatic to, that maintained by Da:Da (“succession”). Alternatively, the relative strengths of the two programs could determine the fate outcome for the cell (“antagonism”). To distinguish between these alternatives, we evaluated the ability of an excess of Da monomer to suppress EE differentiation induced by overexpression of Sc with esgTS-FO. Overexpression of sc in ISCs and EBs leads to a quick expansion of Pros+ cells (Fig 6D) (Bardin et al, 2010; Chen et al, 2018). These were often mitotic (Fig S3A, D), and the number of GFP+ cells generated in only 3 days of lineage tracing was much higher than those generated during normal tissue replacement (Fig 6D, compare with 2A and 5A). This suggests that some of these cells have been trapped into a pre-EE state, where cells express Pros and Dl and are also capable of mitosis (Zeng & Hou, 2015; Chen et al, 2018). However, the co-expression of Da with Sc greatly reduced the number of extra Pros+ Dl− cells and led to an increase in Dl+ cells (Fig 6E, H), while maintaining mitotic figures (Fig S3B, D). Under these conditions, the amount of Sc:Da is expected to be either similar or higher than with the overexpression of Sc alone (as endogenous da is weakly expressed (Bardin et al, 2010), likely in limiting amounts), with a simultaneous increase in Da homodimers. Thus, the reduction of EE differentiation and increase of ISC population is indicative of the antagonism, rather than the succession, of their respective transcriptional programs. If that was true, we would expect that the co-expression of tethered Da:Da and Sc would result in even less EE differentiation, as this construct cannot contribute Da monomers to bind to Sc. This was the case, resulting in an even higher amount of Dl+ cells at the expense of Pros+ cell formation (Fig 6F, H), with a concomitant increase in mitotic figures (Fig S3C, D).
To further test the capacity of Da homodimers to antagonise EE differentiation, we co-expressed da:da and NotchRNAi with esgTS-FO, as the reduction of Notch leads to the formation of abundant Pros+ EE-like cells (Ohlstein & Spradling, 2006; Bardin et al, 2010). Indeed, Da:Da opposed the formation of extra Pros+ cells under conditions of Notch loss (Fig 6G, compare with Fig 4A). This further shows that Da:Da can oppose EE differentiation, and suggests that Sc:Da dimers need to overcome the activity of Da:Da dimers to induce the EE fate.
Da:Da promotes ISC identity while Da:Sc confers mixed ISC/EE properties
Our data indicate that Da:Da and Sc:Da activate distinct and antagonistic transcriptional programs to maintain self-renewal and initiate EE differentiation, respectively. However, during the development of the peripheral nervous system, both Da homodimers and Sc:Da heterodimers seem to be able to direct the same cell fate (sensory organ precursor) (Troost et al, 2015). To determine the transcriptional signature induced by these factors, we performed RNAseq analysis of the overexpression of either Da, Da:Da or Sc, in ISCs, EBs and ECs simultaneously.
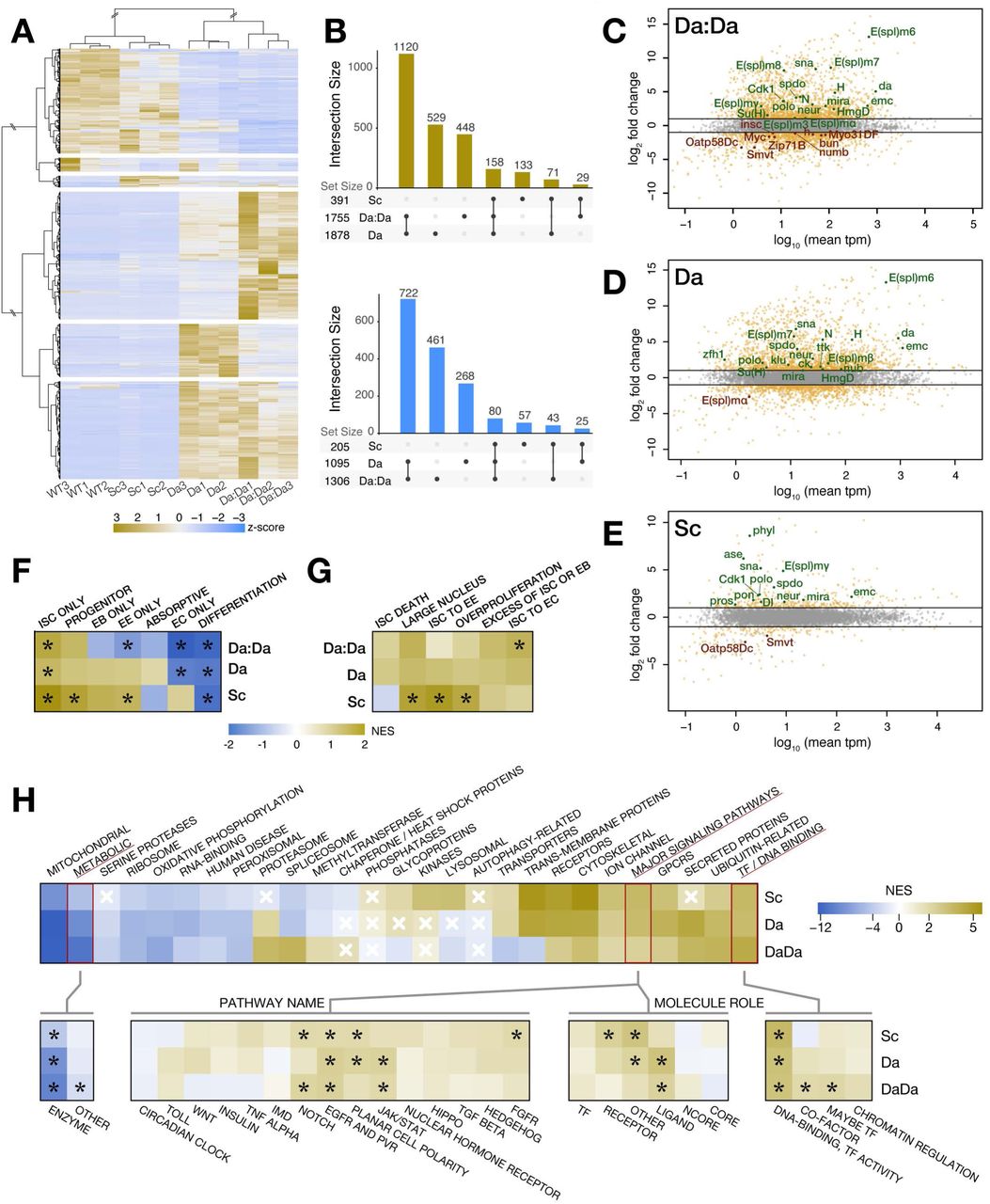
Each overexpression experiment gave a distinct signature (Fig 7A, Fig S4A and Table S3), with Sc inducing modest changes in gene expression, and Da and Da:Da affecting far more genes (Fig 7B). Interestingly, the three conditions had a modest overlap, and monomeric Da induced gene expression changes in many genes unaffected by either Da:Da or Sc, suggesting that it may participate in additional complexes (Fig 7B). We first considered whether the genes upregulated in the three conditions were located in the vicinity of regulatory elements containing the conserved binding site of the bHLH factor, the E-box. Indeed, using the target and enhancer prediction tool i-cisTarget (Herrmann et al, 2012; Imrichova et al, 2015) we observed that E-boxes were overrepresented in the predicted regulatory elements of the genes with elevated expression after Da, Da:Da or Sc overexpression (Tables S4, S5 and S6), suggesting that a good fraction of these genes might be primary targets.

A-B. Da:Da, Da and Sc induce specific but overlapping transcriptional signatures. Hierarchical clustering of differentially expressed genes (fold change > 4) (A) shows groups of genes specifically increased or reduced in Da, Da:Da and Sc respect to the wild-type. UpSetR plots (B) indicate the degree of overlap between these signatures, which is highest between Da and Da:Da.
C-E. MA plots (log2 fold change vs mean expression in transcripts per million, tpm) for the overexpression of Da:Da, Da and Sc, respectively. Some genes of interest are identified because of their role or expression pattern.
F-H. Heatmaps of gene set enrichment scores for sets of genes that are: expressed in specific cell types (Dutta et al, 2015) (F), known to be functionally relevant in different aspects of intestinal homeostasis (Zeng et al, 2015) (G) or annotated because of their broad molecular function in Drosophila (Hu et al, 2015). Black asterisks indicate the enrichment scores with p-values < 0.05 in that heatmap. White crosses indicate the enrichment scores with p-values > 0.05 in that heatmap; the rest have p-values < 0.05.
Overexpressing Da or Da:Da led to increased expression of many genes known to be expressed specifically in ISC/EBs and/or required for their maintenance and proliferation (Table S7) like spdo, polo, Cdk1, mira, HmgD, klu, zfh1, sna, ttk and multiple components of the Notch pathway (N, Su(H), H, neur, and several members of the E(spl) complex of HLH/BFM transcription factors (E(spl)-C), typical targets and effectors of the pathway); it also led to decreased expression of known mediators of differentiation into EEs (pros, numb) or ECs (Myo31DF) (Fig 7C-D). By contrast, overexpression of Sc led to increased expression of genes known to promote EE differentiation (ase, pros, phyl), as well as of genes expressed specifically in ISCs and/or being required for ISC function, such as Dl, neur, spdo, polo, Cdk1, pon, sna, mira and HmgD (Fig 7E). This is all in agreement with our phenotypic observations. However, Da:Da or Da overexpression also induced increased expression of genes required for differentiation and/or specifically expressed in differentiated cells (nub/pdm1, ck) while reducing expression of other ISC and/or EB marker genes (Smvt, Oatp58Dc, Myc, bun, insc, Zip71B) (Fig 7C-D), which shows that the response of individual genes is insufficient to characterise the molecular phenotypes of Da:Da, Da and Sc.
To characterise the molecular signature induced by Da:Da, Da and Sc more systematically, we used Gene Set Enrichment Analysis (GSEA; Subramanian et al, 2005) against previously described transcriptional profiles of midgut cell types (Dutta et al, 2015) and genes reported to be required for normal homeostasis in the Drosophila gut (Zeng et al, 2015). Genes upregulated by Da:Da, Da and Sc were enriched in ISC-specific signatures, in line with the genetic requirements of Da as well as our observation that Sc contributes to conferring ISC properties (Fig 7F and Fig S4B-C). Genes regulated by Da and Da:Da also showed depletion of the transcriptional signature of ECs and genes common to differentiated cells, while Da:Da showed specifically a reduction in EE-specific genes (Fig 7F and Fig S4D-E). In turn, Sc-regulated genes showed enrichment in genes specifically expressed in EEs (Fig 7F and Fig S4F) and required for their differentiation (Fig 7G and Fig S4B-C).
We next analysed the transcriptional signatures induced by Da, Da:Da and Sc more generally, to determine whether they indicated changes in specific cellular processes. Using the Gene List Annotation for Drosophila (GLAD) resource (Hu et al, 2015), the transcriptional signatures of Da:Da, Da and Sc overexpression showed enrichment in categories involved in regulatory mechanisms (receptors, transcription factors and DNA binding, GCPRs and signalling pathways, with Notch and EGFR/receptor tyrosine kinases (RTKs) singled out) (Fig 7H and Fig S4I). Interestingly, they also showed depletion of genes involved in metabolism, ribosome, mitochondrial function and oxidative phosphorylation (Fig 7H and Fig S4G-H). This is unlikely to reflect a switch towards glycolytic metabolism, as most glycolytic genes (including Pfk, which codes for the rate-limiting enzyme) are generally either not affected or mildly downregulated under these conditions (Table S8). These two features (higher decision-making pathways and lower metabolism) were confirmed with a GSEA of Gene Ontology terms (Biological Process), which show an enrichment of terms related to fate specification and morphogenesis (specifically Notch and RTK pathways), and depletion of metabolic and mitochondrial-related genes. Interestingly, this analysis also shows an enrichment of genes regulating cell cycle and cytokinesis (Table S9 and Fig S4J); in the case of Sc, this is in agreement with its recently described role as a mitotic inducer in the intestine (Chen et al, 2018).
These results suggest that Da and Sc, while enhancing largely overlapping developmental and cell cycle regulatory processes, induce distinct signatures which actively promote the ISC and EE identities, respectively. However, Sc can also induce ISC-specific genes. In parallel, Da and Sc promote a state of lower metabolic signature.
Da:Da is antagonised by Notch and maintains self-renewal in parallel to Esg
Esg, a Zinc finger factor of the Snail family, is required to prevent ISC/EB differentiation, mostly through transcriptionally repressing differentiation genes (Korzelius et al, 2014; Loza-Coll et al, 2014). Da:Da seems to have a similar effect, through the activation of ISC-identity genes. Therefore, we sought to test whether they may be acting in combination to maintain stemness. Knockdown of esg in ISCs and EBs leads to differentiation, with an overrepresentation of EEs (Fig S5C and 8F), as expected from the described antagonism between Esg and Sc at the pros locus (Li et al, 2017). Simultaneous overexpression of UAS-esgRNAi and UAS-da:da prevented most differentiation, and specifically reduced EE cell production (Fig 8D and 8F), indicating that Da:Da can maintain ISCs and oppose EE differentiation independently of esg. In turn, overexpression of esg blocked differentiation irrespective of the presence of da (Fig 8C, F and Fig S5B), indicating that Da:Da and Esg largely act independently in ISC maintenance. Furthermore, expression of da and esg are independent of each other (Fig S5D, E).

Solid arrowheads: ISCs (Hdc+/NRE− or Dl+/NRE−); empty arrowheads: EEs (Pros+).
A-B. da is epistatic over Notch signalling. Loss of da leads to EC and EE differentiation when Notch signalling is impaired by overexpression of H (A), while overexpression of da:da maintains cell undifferentiated even when Notch signalling is exacerbated by knockdown of H (B).
C-D. da and esg function in parallel to prevent differentiation. Overexpression of esg can prevent differentiation and compensate for the loss of da (C), while overexpression of da:da can similarly compensate for the loss of esg, including prevention of EE formation (D).
E-F. Stacked bars plots summarising the cell type composition of esgTS-FO tissue. Numbers (total GFP+ cells / fields of view) are 1510/21, 1810/21, 613/8 and 984/15 for UAS-H, UAS-H+UAS-daRNAiJF, UAS-HRNAiHMSand UAS-HRNAiHMS+UAS-da:da, respectively (E) and 850/9, 1236/12, 628/18 and 939/19 for for UAS-esg UAS-esg+UAS-daRNAiJF, UAS-esgRNAiHMS and UAS-esgRNAiHMS+UAS-da:da, respectively (F). p-values are <0.05, <0.01 and <0.001 for one, two or three asterisks (see Tables S1 and S2 for further details). Scale bar: 20µm in all panels.
We next sought to determine whether Da:Da operates as part of a pathway with the known regulators of ISC fate. Our previous epistatic analysis with emc and Notch signalling suggested that Da:Da is required independently of the Notch pathway. Indeed, when we knocked-down both N and da using esgTS-FO, all ISCs/EBs became ECs (Fig S5A). Similar results were obtained by simultaneously knocking-down da and inhibiting Notch signalling by overexpressing H (Fig 8A, compare with Fig 4C, and Fig 8E). However, expressing UAS-da:da prevented the differentiation induced by increasing Notch signalling by knockdown of H (Fig 8B, compare with Fig 4E, and Fig 8E), suggesting that Da:Da is antagonised by Notch signalling. Together, our data indicate that Da homodimers operate downstream of Notch and in parallel to Esg to maintain ISC self-renewal.
DISCUSSION
Multipotent adult stem cells must be able to change their transcriptional state from self-renewal to several alternative differentiation programs. bHLH transcription factors seem well placed to coordinate these changes molecularly; they have been described to operate synergistically, antagonistically or combinatorially (Amoutzias et al, 2008; Bhattacharya & Baker, 2011; Troost et al, 2015; Sallee et al, 2017); their capacity for dimerization allows reduction in molecular noise (Bundschuh et al, 2003). Here we show that a bHLH factor network controls three alternate fates in the adult Drosophila intestine. Class I homodimers (Da:Da) promote the progenitor state of ISCs/EBs. Progenitor cells will acquire the absorptive fate by titration of Da by the HLH factor Emc, or the secretory fate by expression of the class II bHLH Sc, which heterodimerises with Da to initiate EE differentiation. Moreover, Emc is required in EBs to maintain their committed state, while low levels of Sc seem to contribute to boost the ISC transcriptional program. Our results argue for a central and conserved role of bHLH factors in the acquisition and maintenance of multiple fates in the metazoan intestine.
Three fates regulated by a dimerization network
Our data show that Da homodimers maintain ISCs and EBs undifferentiated, but this situation can be easily swayed towards either EC or EE differentiation by Da:Da swapping partners with Emc or Sc (Fig 9). Similar networks involving class I, II and V bHLH factors have been found to regulate the development of the Drosophila retina (Bhattacharya & Baker, 2011) and the peripheral nervous system (PNS) (Cubas et al, 1991; Van Doren et al, 1991; Troost et al, 2015). However, in these cases only one new fate is accessed (neural), and Da:Da dimers seem to have a relatively ‘accessory’ role. In the retina, Da:Da dampens the titration by Emc of the bHLH proneural factor Ato (Bhattacharya & Baker, 2011; Li & Baker, 2018). In the PNS, Da:Da cooperate with Da:Ac/Sc heterodimers to overcome Emc inhibition and induce neural fate, which both Da:Ac/Sc and Da:Da can induce (Troost et al, 2015). By contrast, in the adult midgut Da:Da and Da:Sc support distinct fates (progenitor and secretory, respectively) while Emc titrates both dimers to allow EC differentiation, enabling the network to regulate a three-way fate decision. This is possible because Da:Da and Da:Sc induce specific but partially overlapping transcriptional profiles (Fig 7) which yet seem to be able to act antagonistically (Fig 6). By contrast, Emc simply prevents the DNA binding of either Da or Sc (Van Doren et al, 1991; Cabrera et al, 1994), which suggests that EC is the default fate when Da:Da dimers do not operate (Fig 5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Orange arrows: Da homodimers maintain ISCs and EBs undifferentiated, antagonising EC differentiation and pre-EE formation by inducing expression of ISC-specific genes. Pink arrows: An increase of Sc expression induces formation of Da:Sc heterodimers, which steer the transcriptional program towards pre-EE formation; Sc can also induce expression of the ISC-specific signature, potentially leading to EB decommitment if its activity is not prevented by titration by Emc. Green arrows: Emc increased expression in EBs maintains commitment for EC differentiation and can direct terminal EC differentiation by interruption of Da:Da and Da:Sc transcriptional activities. This occurs in parallel to Esg prevention of differentiation and Notch signalling being upstream and opposing Da:Da to induce EC differentiation.
Interestingly, bHLH transcription factors of classes I-II-V (or E/A/Id) are also active in the mammalian intestine. Class II bHLH factors promote EE differentiation (reviewed in Philpott & Winton, 2014) and ISC maintenance (van der Flier et al, 2009). However, class I factors E2a and Heb are specifically expressed in crypt cells in the mouse intestine (van der Flier et al, 2009). Our work suggests that they may function as class I dimers as well as dimerization partners for class II factors. Meanwhile, Emc homologs Id2 and Id3 are expressed specifically in the intestinal differentiated cells (Wice, 1998), suggesting similar functions. By contrast, Id1 is required in ISCs for regeneration (Zhang et al, 2014) and can act as an oncogene (Wice, 1998), which is the opposite behaviour to that of Emc.
A balance of bHLH factors regulates intestinal stem cell fate
Our observations indicate that additional work on the mechanisms that regulate the transcription of both emc and sc would be very informative. The cis-regulatory region of emc seems to be complex and spreads across a large genomic region (Li & Baker, 2019), so it is likely that is subject to multiple regulatory inputs. An obvious candidate for regulation of emc transcription is Notch signalling, as shown in other contexts (Baonza et al, 2000; Baonza & Freeman, 2001; Adam & Montell, 2004; Bhattacharya & Baker, 2009; Spratford & Kumar, 2015), and considering the importance of Notch for EB establishment and EC differentiation (Micchelli & Perrimon, 2006; Ohlstein & Spradling, 2006; Bardin et al, 2010). However, our epistasis analyses suggest that Emc and Notch signalling can induce EC differentiation independently of each other, observation that makes Notch an unlikely regulator of emc in the midgut. A major regulator of Sc is the transcriptional repressor Ttk69 (Wang et al, 2015), whose stability is in turn regulated by the E3 ubiquitin ligase Sina and its adaptor Phyl (Yin & Xi, 2018). Therefore, Phyl expression could be the trigger to allow the accumulation of Sc and initiate EE differentiation, but how phyl is regulated is not known yet. On the other hand, Chen et al. (2018) observed that Sc activates itself as well as E(spl)m8, a member of the Enhancer of split Complex (which comprises several transcription factors, many of which are often both targets and effectors of the Notch pathway; reviewed in Delidakis et al, 2014). As E(spl)m8 is able to repress Sc, the authors proposed that a cell-autonomous oscillator involving Sc and E(spl)-C members would create pulses of Sc expression which would set the pace of EE production (Chen et al, 2018). However, while we too observe induction of E(spl)-C genes upon Sc forced expression, so we do as a result of Da:Da and Da overexpression (Fig 7). This suggests that the regulation of E(spl)-C (and therefore of sc) may be more complicated. Additional work will be necessary to determine the sequence and timing of regulatory inputs that initiate fate-inducing levels of Sc or Emc.
Moreover, the Emc/Da/Sc network clearly does not work as a simple boolean switch. The three genes are expressed, and seem to be functionally relevant, in both ISCs and EBs (Bardin et al, 2010; Chen et al, 2018; Doupé et al, 2018; this work). This argues against a mechanism that simply initiates Emc or Sc expression to induce the EC or EE fates. Moreover, the functions of Emc and Sc seem to be more complex than acting as mere fate switches. Adequate levels of Emc are required in the EB, as its depletion induces de-differentiation into ISCs (Fig 3), but its overexpression accelerates EB terminal differentiation into ECs (Fig 2). Intriguingly, it has recently been proposed that the Emc homolog Id1 protects pluripotency specifically during the peri-implantation transition (Malaguti et al, 2019), which suggests that Emc could be part of a timing mechanism in the transition between ISC and EC. As for Sc, our data suggests that low levels of expression in ISCs (Chen et al, 2018; Doupé et al, 2018) induce the expression of ISC-specific genes (Fig 7) without eliciting a conversion into pre-EE cell. Higher levels would be required to initiate EE differentiation. Therefore, the amounts of Da, Sc and Emc must be maintained in balance. In the case of Emc, this is likely to be achieved through post-transcriptional mechanisms. During neurogenesis, Emc is stabilised when bound to Da, and degrades when Da frees itself to bind a proneural factor (Bhattacharya & Baker, 2011; Li & Baker, 2018). This may be the reason why Emc shows higher levels of accumulation in EBs and ECs (Fig 1), where Da is expressed but other bHLH factors are at minimal levels.
Da and Sc cooperate to endow cells with ISC properties
Our data shows a critical requirement for Da:Da in ISC maintenance, but also that Sc can contribute to enhance ISC features (proliferation, expression of Dl). This is likely an active promotion of ISC fate, rather than just prevention of differentiation, as our transcriptomic analysis shows an enrichment of the ISC transcriptional signature when overexpressing either Da:Da, Da or Sc, together with a depletion of differentiation genes (except for EE-specific genes and Sc). This active role is further supported by the capacity of Sc to induce EBs to de-differentiate into ISCs and re- or transdifferentiate into EEs (Fig 6). The transcriptional changes induced by Sc, Da and Da:Da expression also show an enrichment in receptors and ligands of signalling pathways (Fig 7). This is in agreement with the recent proposal that ISCs are sources of autocrine signals required for their own maintenance (Doupé et al, 2018). We also find evidence that mitochondrial function is reduced by expression of Sc, Da and Da:Da (Fig 7), which fits well the observation that ISCs display an ‘empty mitochondria’ phenotype under the electron microscope (Hung et al, 2018). Together, this suggests that low levels of Sc:Da dimers contribute non-critically to the maintenance of the ISC identity and function, and we surmise that Emc expression in EBs might be required to dampen this Sc activity.
Regulation of the committed state of the enteroblast
Under normal conditions, EBs will only give rise to mature ECs without further division (Zeng & Hou, 2015; Wang et al, 2015; Yin & Xi, 2018). Depletion of Emc specifically in EBs coupled to lineage tracing results in cells proliferating and expressing Dl (Fig 3), which likely represents a de-differentiation process. This may partially reflect the need to control Sc activity in the EB, as the expression of Sc can induce the same effects (Fig 6). Intriguingly, EB-specific depletion of Ttk69 or Klu or overexpression of Phyl seem to induce trans-differentiation, as these conditions produce Pros+ cells (Wang et al, 2015; Yin & Xi, 2018). This difference could be because Emc is also influencing the activity of multiple Da-containing complexes in the EB, or because of the activity of additional targets of Klu and the Phyl/Ttk69 pathway than Sc. The reacquisition of proliferation and Delta expression is also controlled by other factors, such as Sox21a (Zhai et al, 2015), or the global co-repressor Gro, acting downstream of Notch (Guo et al, 2019) (though surprisingly Notch itself does not seem to be required to maintain the EB; Siudeja et al, 2015). Together with our observations, this indicates a previously unappreciated plasticity of the EB, which is consistent with its similarity to the ISC in transcriptome (Hung et al, 2018) and chromatin accessibility landscape (Aughey et al, 2018). Importantly, this resembles the behaviour of EC precursors in the mammalian intestine, which can dedifferentiate and repopulate the intestinal crypt during regeneration (Tetteh et al, 2016).
MATERIALS AND METHODS
Drosophila culture and genetics
Flies were raised and maintained on standard cornmeal/yeast medium at 25°C unless specifically indicated. Experiments were conducted in mated females, collected daily and maintained in fresh vials with added yeast until 4-7 days old. For experiments using Gal80TS, flies were reared and aged to gut maturity (∼7 days) at 18°C, then transferred to 29 °C. For induction of MARCM and flip-out clones, 4-7 days old flies were treated at 37°C for 60 or 15 min, respectively. Flies were aged for 7 days after induction treatment before dissection, unless otherwise indicated. See Appendix for fly strains.
Immunohistofluorescence, confocal microscopy and image analysis
Stainings were performed as in Bardin et al, (2010). See Appendix for details on primary and secondary antibodies. DNA dye was Hoechst (Sigma-Aldrich B2261 used 1:5000 from a stock solution at 10 mg/ml). Mounting medium was 4% N-propyl-galate, 80% glycerol. Confocal stacks were obtained in a Zeiss LSM 710 with an EC Plan-Neofluar 40X oil immersion objective (numerical aperture 1.3). All stack positions were acquired in the posterior midgut. Typically, three positions along the anterior-posterior axis of the posterior midgut were acquired for each organ. In MARCM clone experiments, stacks were acquired from all clones in the posterior midgut. Quantification of anti-Dl staining is detailed in Images and figures were assembled using Adobe Photoshop and Illustrator CS6.
Cell counts and statistics
For evaluating the proportion of cell types in GFP+ clones or labelled tissue, confocal stacks were maximum-intensity projected using FIJI/ImageJ (Schindelin et al, 2012), cells of the relevant types were counted with the Cell Counter plugin. In the experiments overexpressing UAS-sc and UAS-da:da, the associated increase in proliferation generated large, highly densely populated cell clusters which could not be counted with single-cell precision. Therefore, for this genotype we estimated the proportion of each cell population in each field of view separately. See the Appendix for details on cell type identification.
Statistical tests were performed in Prism 8 or in R 3.5.1 (R Core Team, 2018) using RStudio 1.1.383. Statistical significance of changes in proportions of cell types was assessed by binomial logistic regression in basic R. We used Firth’s bias reduced logistic regression (package logistf; Heinze and Ploner, 2013) in experiments with zero observations in one or more of the genotypes considered, to avoid the nonsensical results arising from the ‘complete separation’ of data (Albert & Anderson, 1984).
RNA-seq
Flies bearing either UAS-da:da, UAS-da or UAS-sc as well as esg-Gal4, Myo1A-Gal4 and tub-Gal80TS were reared at 18°C until 4-7 days old, transferred to 29°C for 24h and their posterior midguts dissected, collected in lysis buffer and frozen at −80°C, then thawed and extracted for total RNA. Libraries from three biological replicates per condition were prepared and ∼26 million reads per library were generated using Illumina technology. See the Appendix for additional details.
AUTHOR CONTRIBUTIONS
Conception of the study and supervision: JdN. Data acquisition, analysis and interpretation of data: AP and JdN. Experimental design; writing, revising and approval of the manuscript: JdN and AP.
CONFLICT OF INTEREST
The authors declare no competing interest.
ACKNOWLEDGEMENTS
We thank Nicholas Baker, Allison Bardin, Antonio Baonza, Sonsoles Campuzano, Sangbin Park, Bruce Edgar, Mike Taylor, Shinya Yamamoto, the Bloomington Drosophila Stock Center, the Vienna Drosophila Resource Center, the Drosophila Genetics Resource Center (Kyoto) and the National Institute of Genetics (Japan) for providing fly stocks, and the Developmental Studies Hybridoma Bank (University of Iowa) for supplying antibodies.
We would like to thank Juan Modolell, Sonsoles Campuzano, Catherine Hogan, Fernando dos Anjos-Afonso, Florian Siebzehnrubl and Terrence Trinca for critical comments on the manuscript, Sonia López de Quinto, Helen White-Cooper, Mike Taylor and Wynand van der Goers van Naters for useful discussions during the development of the project, and the data analysis team of the College of Biomedical and Life Sciences and the Genomics facility of the School of Biosciences of Cardiff University for technical assistance. This work was supported by Cardiff University.
Footnotes
The previous submissions had the main figures duplicated.
REFERENCES




