

ABSTRACT
LIN-28 is a conserved RNA-binding protein known for its critical roles in the C. elegans heterochronic pathway and its capacity to induce proliferation and pluripotency as well as its oncogenic potential in mammals. LIN-28 binds to the precursor of the conserved – cellular differentiation-promoting – microRNA let-7 and inhibits its maturation. LIN-28 also binds to and regulates many mRNAs in various cell types. However, the determinants and consequences of LIN-28-mRNA interactions are not well understood. Here, we report that LIN-28 in C. elegans represses the expression of LIN-46, a downstream protein in the heterochronic pathway, via the (unusually conserved) 5’ UTR of the lin-46 mRNA. We found that both LIN-28 and the 5’UTR of lin-46 are required to prevent LIN-46 expression at the L1/L2 stages and that precocious LIN-46 expression, caused by mutations in the lin-46 5’UTR, is sufficient to skip L2 stage cell-fates, resulting in heterochronic defects similar to the ones observed in lin-28(lf) animals. We hypothesize that the lin-46 5’UTR mediates LIN-28 binding to the lin-46 mRNA, which results in the repression of LIN-46 expression. Our results show that precocious LIN-46 expression alone can account for lin-28(lf) phenotypes, demonstrating the biological importance of direct (let-7-independent) regulation of target mRNAs by LIN-28.
INTRODUCTION
Animal development involves complex cell lineages within which different cell-fates are executed in specific orders and at a pace that is in synchrony with overall developmental rate. Expressing symmetric cell-fates that allow cell proliferation or asymmetric cell-fates that enable both self-renewal and the generation of new cell types, a single totipotent cell (embryo) and its progeny generate the populations of specified cells that form the diverse tissues and organs of the animal body. Gene regulatory networks control the levels and spatiotemporal expression patterns of developmental genes so that proper cell-fates are acquired at the right time and place during development.
C. elegans develops through four larval stages (L1-L4). Each larval stage is comprised of an invariant set of cell division and cell fate specification events [1]. The order of cell-fates and the timing of cell-fate transitions within individual cell lineages are regulated by genes in the heterochronic pathway [2,3]. In this pathway, three major temporal regulatory transcription factors control the transitions from earlier to later cell fates. These transcription factors are directly or indirectly regulated by microRNAs and/or RNA binding proteins, which facilitate proper cell-fate transitions by controlling the developmentally dynamic expression of the heterochronic pathway transcription factors.
One of the transcription factors in the heterochronic pathway, Hunchback-like-1 (HBL-1), promotes L2-stage symmetric cell divisions and prevents progression to L3-stage asymmetric cell divisions [4,5]. HBL-1 is expressed at the L1&L2 stages and it is downregulated during the L2-to-L3 transition [4,6]. Proper temporal regulation of HBL-1 activity is characterized by specification of L2 cell-fates at the L2 stage and progression to L3 cell-fates concomitant with the L2-to-L3 stage progression. Mutations that cause reduced HBL-1 activity at the L1/L2 stages result in skipping of L2 cell-fates, whereas mutations that cause ectopic HBL-1 activity at the L3/L4 stages lead to reiterations of L2 cell-fates at these later stages. HBL-1 is regulated, during the L2-to-L3 transition, by let-7-family microRNAs (mir-48/84/241) [7] and in parallel by lin-28 [7,8], which acts on hbl-1 indirectly, via a protein coding gene lin-46 oi2+[9].
LIN-28 is a conserved RNA-binding protein discovered as a heterochronic gene product in C. elegans [2]. In C. elegans larvae lacking lin-28 activity, hypodermal stem cells skip L2-stage specific symmetric cell divisions, and precociously transition to later stage cell-fates, which eventually leads to terminal differentiation of the hypodermis while the rest of the larval tissues are still developing [2]. LIN-28 inhibits the maturation of the conserved microRNA let-7 [10], which, in C. elegans, is required for the terminal differentiation of hypodermal tissues at the end of the last larval stage [11]. Curiously, although let-7 is expressed precociously at the early stages in lin-28(lf) larvae and let-7 function is required for the precocious terminal differentiation of hypodermal cells in lin-28(lf) animals, let-7 function is not required for the skipping of L2 stage cell fates [8]. Instead, loss of a protein coding gene, lin-46, suppresses both early and late stage lin-28(lf) phenotypes [9], without repressing precocious let-7 expression [8]. This suggests that LIN-46 might be mis-regulated in lin-28(lf) animals, which could potentially be responsible for lin-28(lf) phenotypes. It has been shown that LIN-28 binds to the lin-46 mRNA [12], but the consequence of LIN-28 binding to the lin-46 mRNA or how (and if) LIN-28 regulates lin-46 is not known. Additionally, lin-46 encodes a protein related to bacterial molybdenum cofactor biosynthesis enzyme, and mammalian Gephyrin [13,14] and, although molecular functions of LIN-46 is not clear, our recent findings suggest that LIN-46 affects temporal cell-fates by inhibiting the nuclear accumulation of the transcription factor HBL-1 [manuscript-oi2]. Therefore, it is possible that LIN-28 could promote L2 fates by restricting the expression of LIN-46, and hence maintaining nuclear activity of the L2-fate factor HBL-1.
C. elegans LIN-28 and its homologs in mammals (Lin28) have conserved functions: LIN-28/Lin28 inhibits let-7 expression [10,15–17], binds to and regulates many mRNAs [12,18–20], and promotes proliferation and pluripotency [21–23]. Similar to C. elegans development, during mammalian embryogenesis LIN-28 is expressed at early or pluripotent stages and it is downregulated at later or more differentiated stages [21,24,25]. LIN-28 down-regulation in differentiating tissues allows let-7 microRNA to accumulate; and let-7 further promotes differentiation by repressing pluripotency and self-renewal promoting genes [26], including lin-28. Lastly, LIN-28 expression is associated with many types of cancers and poor prognosis [27,28], and conversely, let-7 is known to act as a tumor suppressor by repressing oncogenes [29–32].
While certain phenotypes observed in Lin28-deficient mammalian cells can be attributed to increased let-7 expression and consequent repression of let-7 targets [17,33], there are let-7 independent functions of Lin28 [8,34,35], some of which could be explained by mis-regulation of specific mRNA targets of Lin28 [36–39]. Lin28 can regulate the translation of target mRNAs either positively [20] or negatively [18], perhaps in a cell-type specific manner. LIN-28 seems to have a tendency to bind to multiple sites on its mRNA targets, for example, Cho et al. found 38.5 sites per mRNA [18]. Therefore, testing the consequences of loss of LIN-28-mediated regulation of specific mRNAs, which would require mutating all potential LIN-28 binding sites on an mRNA, seems challenging. Moreover, although the LIN-28-bound mRNA regions are enriched for certain motifs (e.g. GGAG), these sequence motifs are neither required nor sufficient for LIN-28 binding to its targets [18,20]. In brief, the rules and consequences of LIN-28/lin-28 binding to mRNAs are not well understood.
Here we show that the critical target of LIN-28 in C. elegans, LIN-46, is expressed only at the L3&L4 stages in a temporal profile that is the inverse of LIN-28, which is expressed at the L1&L2 stages. We find that LIN-46 is expressed precociously at the L1&L2 stages in lin-28(lf) animals, supporting the idea that LIN-28 represses LIN-46 expression at these early larval stages. We also find that, similar to lin-28(lf), mutations in the 5’UTR of lin-46 result in precocious LIN-46 expression in the hypodermal seam cells, and that this ectopic LIN-46 expression at the L1/L2 stages is sufficient to result in skipping of L2 stage symmetric seam cell divisions, and precocious expression of L3-adult fates. Endogenously tagged LIN-46 is also expressed in the vulval precursor cells (VPCs) and LIN-46 is precociously expressed in the VPCs both in lin-28(lf) and lin-46 5’UTR mutants. Ectopic LIN-46 expression in the VPCs in lin-46 5’UTR mutants is sufficient to accelerate cell-fate transitions in these cells, which results in protruding vulva phenotypes similar to lin-28(lf) animals. Due to the phenotypic similarity between lin-28(lf) and lin-46 5’UTR mutants, we hypothesize that the lin-46 5’UTR mediates LIN-28 binding to the lin-46 mRNA, which results in the repression of LIN-46 expression from the lin-46 mRNA. Our results demonstrate that precocious LIN-46 expression alone, which is observed in lin-28(lf) animals and is sufficient to suppress L2 cell-fates and to induce precocious transition to L3 cell-fates, can account for majority of the lin-28(lf) phenotypes.
RESULTS
lin-28 represses LIN-46 expression at the early larval stages
The developmental expression patterns of lin-28 and lin-46 were previously identified using transcriptional reporters (transgenes expressing a fluorescent protein driven by the promoter of interest) and translational reporters (transgenes expressing the open reading frame of a gene of interest fused with a fluorescent protein driven by the promoter of the gene) [9,21]. The expression of such transgenes does not necessarily accurately represent the expression levels or spatiotemporal patterns of genes expressed from their endogenous loci. To more accurately determine the expression patterns of LIN-28 and LIN-46, we used CRISPR/Cas9 genome editing to tag lin-28 and lin-46 with fluorescent proteins at their endogenous loci (Figure 1A). Both of the endogenously tagged loci were determined to be fully functional. We found that endogenously tagged LIN-28::GFP expression is comparable to previous reports [21]: LIN-28 is highly expressed in the embryos and at the L1 and L2 stage larvae and it is diminished at the L3 and later stages (Figure 1B&C). However, the expression pattern of endogenously tagged LIN-46::mCherry differs from the pattern observed using transcriptional and translational reporters [9]: LIN-46::mCherry was not detected at all stages, but only at the L3&L4 stages, and most prominently in hypodermal seam cells (Figure 1C) and in the ventral hypodermal vulval precursor cells (See Table S1 for a detailed comparison). Based on the previous report that the lin-46 transcriptional reporter transgene is expressed at all larval stages [9], our finding that LIN-46::mCherry expression is restricted to the L3&L4 stages suggests that lin-46 is post-transcriptional regulated, perhaps on the level of mRNA translation.
Comparison of LIN-46 expression observed in transgene reporters versus endogenously tagged locus

A) Schematic views of the C. elegans lin-28 and lin-46 genes and the CRISPR-mediated integration of GFP (green) or mCherry (red) coding sequences at the C-termini. Thick bars represent the exons, thin bars represent the 5’UTRs (left) and 3’UTRs (right). Lines between the exons represent the introns. Ma426 denotes the endogenously GFP-tagged lin-28 allele and ma398 denotes the endogenously mCherry-tagged lin-46 allele. The ma398ma459 allele harbors the “GTAT” deletion in the 5’UTR of lin-46 that is also tagged with mCherry at the C-terminus. B) DIC and fluorescent images showing LIN-28 and LIN-46 expression in wildtype L2 (first row) and L4 (last row) stage larvae, and in lin-28(lf) and lin-46(gf/[ΔGTAT]) L2 stage larvae. Fluorescent images of different larvae are taken using the same microscopy setting. All images are then stitched together using the ImageJ software to adjust the brightness and contrast uniformly across the images for enhanced visualization. C) Schematic representation of the LIN-28 and LIN-46 expression observed during L1-L4 larval stages of wildtype, lin-28(lf), and lin-46(gf/[ΔGTAT]) animals. Note that LIN-46 is precociously expressed at the L1&L2 stages in both lin-28(lf) and lin-46(gf[ΔGTAT]) mutants.
The expression pattern of endogenously tagged LIN-46 reveals that LIN-28 and LIN-46 are expressed in a temporally mutually exclusive manner: LIN-28 is expressed early (L1&L2 stages) and LIN-46 is expressed late (L3&L4 stages). This mutually exclusive expression pattern suggested that LIN-28 could potentially repress LIN-46 expression during the L1 and L2 larval stages. To test this, we examined the effect of loss of lin-28 on the expression pattern of LIN-46. We indeed found that LIN-46 is expressed precociously at the L1 and L2 stages in lin-28(lf) animals (Figure 1B&C), consistent with the conclusion that LIN-28 represses LIN-46 expression at these early larval stages.
Mutations in the lin-46 5’UTR result in lin-28(lf)-like phenotypes and precocious LIN-46 expression
The finding that LIN-46 is expressed precociously at the L1 and L2 stages in lin-28(lf) animals suggests two non-mutually exclusive hypotheses; 1) that LIN-28 might directly repress LIN-46 expression, and 2) that precocious LIN-46 expression could contribute to the precocious developmental phenotypes of lin-28(lf). The latter hypothesis is supported by previous findings that lin-46(lf) suppresses the precocious development of lin-28(lf) [9]. However, although lin-46 activity was shown to be necessary for the precocious phenotypes of lin-28(lf), it was not known if (precocious) LIN-46 expression could be sufficient for precocious development. The possibility that LIN-28 could directly repressed LIN-46 expression by binding to the lin-46 mRNA is supported by CLIP experiments [12].
We sought to test the above two hypotheses by mutation of putative LIN-28-interacting sequences in the lin-46 mRNA sequence and assaying for precocious developmental phenotypes. Although the published CLIP tags map to both 5’ and 3’ UTRs and all five exons of the lin-46 transcript [12], nevertheless, we noticed that the lin-46 5’UTR exhibits unusually high sequence conservation among nematodes (Figure S1A) and contains a GGAG motif that is often associated with LIN-28 binding (Figure S1A&B). Therefore, we targeted the lin-46 5’UTR using a CRISPR guide (gR5U, Figure S1A&B), and observed frequent lin-28(lf)-like phenotypes in the F1/F2 progeny of the injected P0 animals (Figure S1C). We genotyped several of these F1/F2 progeny, and found a range of lin-46 5’UTR deletions varying in size (2-19 bp) in animals expressing lin-28(lf)-like phenotypes (Figure S1D).

A) Genome browser view of the C. elegans lin-46 gene (top) and the magnified 5’UTR sequence (bottom). Genome browser tracks: Ensemble Gene Predictions (top), PhyloP conservation (middle), Nematode Multiz Alignment (bottom). Note that the phyloP and Multiz tracks show the high conservation in the 5’UTR among the seven nematode species listed in the figure. The gR_5U guide cut-site and the GGAG in the 5’UTR of lin-46 are marked. B) lin-46 5’UTR flanked by the TTTCAG splice acceptor and the lin-46 ORF as well as the gR_5U and the GGAG are shown. C) Examples of F1/F2 animals among the progeny of gR_5U injected P0s that express precocious Pcol-19::gfp are marked with red circles. F1/F2 progeny displaying wildtype Pcol-19::gfp pattern are marked with blue arrowheads. D) Various mutations in the 5’UTR of lin-46 that are detected in the F1/F2 progeny of the gR_5U injected animals displaying precocious Pcol-19::gfp or protruding vulva phenotypes listed in the left column. Red fonts indicate the deleted nucleotides in each allele, and three different regions of the lin-46 5’UTR as shown in Figure 4. are highlighted with three different colors. Allele names given to these mutations and the P0s strains injected are given in the middle and right columns, respectively.
To determine the effects of the lin-46 5’UTR mutations on the expression of LIN-46 we injected the CRISPR mix containing the gR_5U guide into animals carrying the lin-28::gfp [lin-28(ma426)] and lin-46::mCherry [lin-46(ma398)] alleles (Figure 1A), and generated lin-46 5’ UTR deletion mutations, which resulted in precocious LIN-46 expression at the L1 and L2 stages (Figure 1A-C). This result shows that (an intact) lin-46 5’UTR is required to prevent precocious LIN-46 expression. Importantly, our CRISPR mutagenesis of the lin-46 5’ UTR did not affect the expression of LIN-28 (Figure 1B&C).; thus LIN-46 is expressed precociously in lin-46 5’UTR mutants despite the presence of LIN-28, indicating that the lin-46 5’UTR likely mediates LIN-28 binding to, and hence the repression of, the lin-46 mRNA.
It is noteworthy that the inhibition of LIN-46 expression by LIN-28 seems to be independent of, and in parallel to the previously-described lin-28-mediated inhibition of let-7 biogenesis, apparently by binding of LIN-28 to the let-7 primary transcript [10]. lin-46(lf) does not suppress the precocious expression of let-7 in lin-28(f) animals, indicating that LIN-46 is not responsible for precocious let-7 biogenesis.
Precocious LIN-46 expression causes precocious cell-fate transitions in hypodermal seam and vulval cell lineages
To compare in detail the phenotypes of lin-46 5’UTR mutants to those of lin-28(lf) mutants we assessed the number of seam cells, the timing of adult specific Pcol-19:gfp expression and the timing of adult alae formation, and protruding vulva morphology (Pvl) (Figure 2). We examined animals homozygous for three different 5’UTR mutations, ma461, ma467, and ma472, that are deleted for 6, 12, and 19 nucleotides, respectively (Figure 2A).

A) Number of seam cells observed in young adults of wildtype, lin-28(lf), and three lin-46 5’UTR mutants (ma461, ma467, and ma472). Each dot in the plot on the right represents the number of seam cells observed on one lateral side of a single worm. Black bars indicate the average number of seam cells observed for each genotype. B) DIC and fluorescent images of L4 and adult stage wildtype and lin-46(ma461) animals. The adult specific Pcol-19::gfp (maIs105) is normally expressed in the hypodermal seam and hyp7 cells at the adult stage of both wildtype and lin-46(ma461) animals. However, unlike in wildtype larvae, pcol-19::gfp is also precociously expressed in the seam cells of L4 stage lin-46(ma461) larvae. C) DIC images showing the (precocious) adult-specific cuticle structure called alae (upper panel) on the cuticle of a larva at the L4 stage indicted by the developmental stage of the gonad (lower panel). D) Percent protruding vulva (Pvl) phenotype observed in wildtype, lin-28(lf), and three lin-46 5’UTR mutants and DIC images showing normal vulva or Pvl morphology observed in each genotype.
All three lin-46 5’UTR mutants displayed an average of fewer than sixteen seam cells (Figure 2A), which indicates that lin-46 5’UTR mutations, presumably as a consequence of consequent precocious LIN-46 expression, result in skipping of L2-stage symmetric seam cell divisions. The severity of this precocious seam cell phenotype is different in three different mutants and lin-28(lf) displays a stronger phenotype than all three 5’UTR mutants (Figure 2A). The variability in the number of seam cells is due to variation in cellfate decisions across the seam cells of each larva; namely, in lin-28(lf) animals almost all seam cells skip L2 cell-fates whereas in the 5’UTR mutants of lin-46 not all but only some (and a varying number of) seam cells skip L2 stage cell-fates, resulting in number of seam cells varying between ten (all the seam cells skipped L2 fates) and sixteen (none of the seam cells skipped L2 fates). This variation in the number of seam cells that skip L2 fates in lin-46 5’UTR mutants could reflect variability in the timing or level of LIN-46 expression across seam cells.
Similar to lin-28(lf) mutants, in lin-46 5’UTR mutants seam, cells precociously express adult fates during larval stages, demonstrated by the precocious expression of a GFP transgene driven by an adult onset collagen promoter, Pcol-19::gfp, (Figure 2B) and by the expression of an adult cuticle structure called alae in L4 stage larvae (Figure 2C).
Lastly, we quantified the percent animals that display protruding vulva (Pvl) phenotypes in young adults (a characteristic of lin-28(lf) animals): all three lin-46 5’UTR mutants displayed Pvl phenotypes similar to lin-28(lf) animals (Figure 2D).
LIN-46 is expressed in the vulval precursor cells (VPCs) and precocious LIN-46 expression leads to precocious onset of vulva development
During C. elegans larval development, stem cells of the ventral hypodermal lineages P3-P8 divide during the L1 stage and give rise to the six P3.p-P8.p vulval precursor cells (VPCs) [1]. After their birth in the L1 stage, the VPCs temporarily arrest in the G1 stage of the cell cycle [40] until the L3 stage when they undergo a single round of cell division. Concomitant with this cell division, three of the six VPCs (P5.p, P6.p, and P7.p) become induced to undergo additional rounds of cell divisions, giving rise to twenty-two cells that progressively differentiate and form the adult vulva. The timing of the first VPC divisions in the mid-L3 stage is controlled by genes in the heterochronic pathway, including lin-28 [2,40]. In lin-28(lf) mutants, the first VPC divisions precociously take place in the L2 stage; the VPC progeny subsequently continue to precociously divide and differentiate, resulting in precocious vulva development, evidenced by an abnormally formed, protruding vulva (hence the Pvul phenotype) in lin-28(lf) L4 animals. Loss of lin-46 suppresses the Pvl phenotype caused by lin-28(lf). However, because LIN-46 expression in the VPCs had not been previously detected using transgenic reporters [9], it was not clear how loss of lin-46 could affect the Pvl phenotype of lin-28(lf) animals.
We found that endogenously tagged LIN-46 is expressed in the VPCs at the L3 and L4 stages (Figure 3A), which coincides with the period when the VPCs develop into adult vulva. In lin-28(lf) mutants, LIN-46 is precociously expressed in the L2-stage VPCs, which coincides with precocious development of these VPCs in lin-28(lf) animals (Figure 3B, left). Moreover, in lin-46 5’UTR mutants, LIN-46 is precociously expressed in the VPCs at the L2 stage and VPCs develop precociously (Figure 3B, right), which indicates that precocious LIN-46 expression is sufficient to alter the timing of vulva development. These results show that precocious LIN-46 expression is likely responsible for precocious vulva development in lin-28(lf) animals.

A) Wildtype larval stages and the cell lineage diagram of the vulval precursor cells (VPC), and DIC and fluorescent showing LIN-46 expression the VCPs. B) Larval stages (according to gonad morphology), precocious LIN-46 expression and VPC development in lin-28(lf) and lin-46(gf[ΔGTAT]) animals are shown.
Altogether, our results show that the LIN-28 target LIN-46 suppresses L2 cell-fates and promotes transition to L3 cell-fates in both seam cells and vulval precursor cells, and precocious LIN-46 expression causes precocious cell-fate transitions, which is likely responsible for the major phenotypes observed in lin-28(lf) animals.
The architecture and conservation of the lin-46 5’UTR
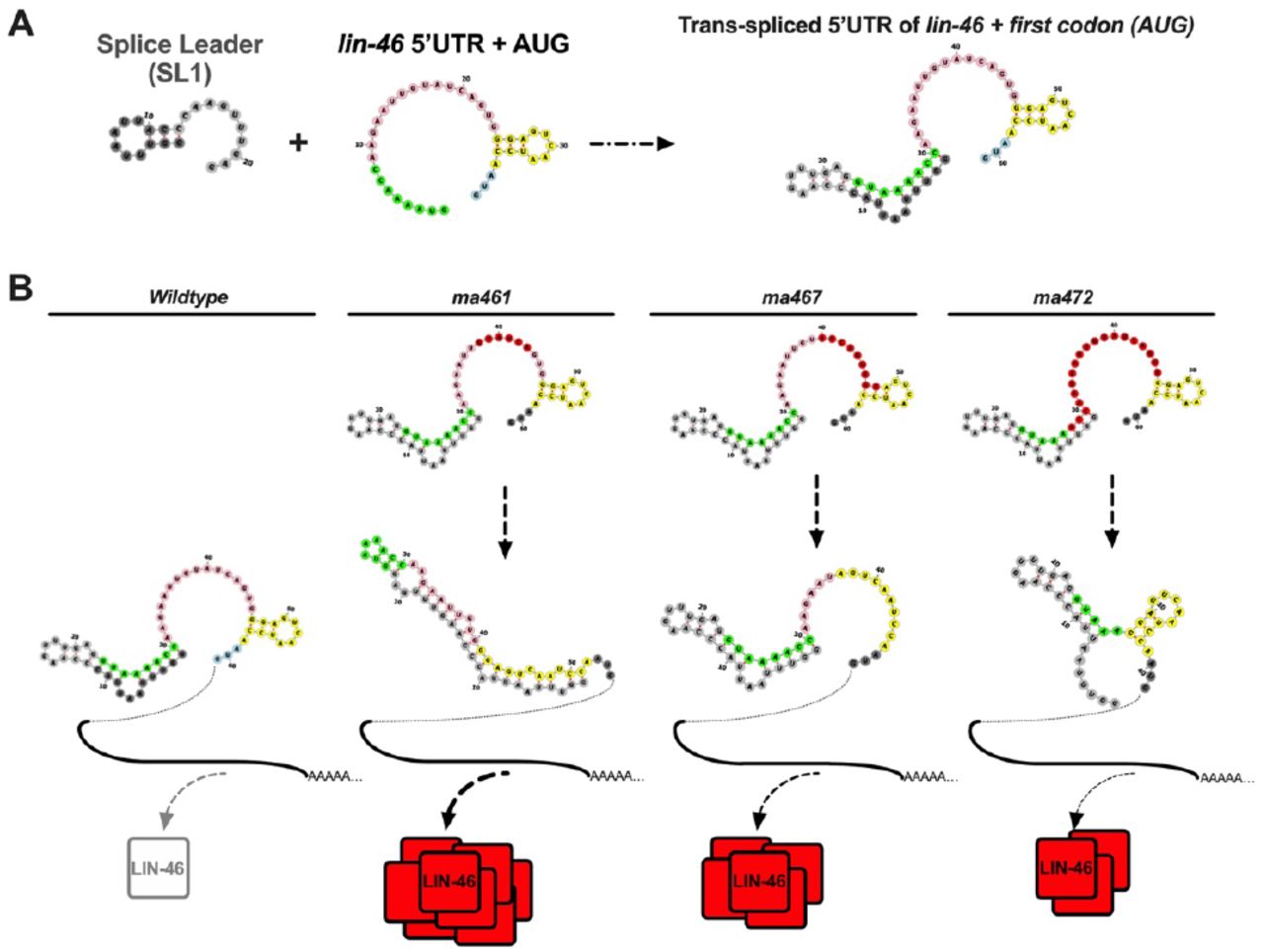
Most C. elegans transcripts are trans-spliced [41], which results in the fusion of a 22-nt spliceleader (SL) RNA to the 5-prime end of the transcripts. The presence of an upstream splice acceptor “TTTCAG” (Figure S1B and 4) and expressed sequence tag (EST) clones that contain the SL1-lin-46-5’UTR fusion sequence, such as GenBank: FN875625.1., indicate that lin-46 mRNA is trans-spliced. We used the RNAfold Webserver [42] to predict the structure of the 5-prime end of the lin-46 mRNA upstream of the first (AUG) codon, which contains a fusion of the SL1 RNA to the lin-46 5’UTR (Figure S2A).

A) Predicted folding of the splice leader (SL1) and lin-46 5’UTR before and after trans-splicing. B) Predicted changes in the folding of the trans-spliced lin-46 5’UTR in three different mutants and schematic representation of LIN-46 expression inferred from the phenotypes (e.g. Figure 2A) observed in animals carrying each mutation.

A) The predicted structure of the transspliced lin-46 5’UTR and the annotation of distinct structural regions. Red arrowheads indicate nucleotides that are mutated in nematodes where 5’UTR of lin-46 is conserved; numbers denote the number of times a mutation is found at the position indicated. B) An alignment of the genomic sequences encoding the lin-46 5’UTR in various Caenorhabditis species. The lin-46 5’UTR is highly conserved and the conservation pattern is consistent with the conservation of the base-pairing interactions in the predicted structure: base-pairing nucleotides are more conserved or nucleotide changes preserve base-pairing.
The predicted structure of the SL1-lin-46-5’UTR chimeric RNA shows base-pairing between SL1 and the first eight nucleotides at the 5-prime end of the lin-46 5’UTR (Figure 4A, “SL1-complementary”).
These eight nucleotides are highly conserved across nematodes; and a nucleotide variation at position four that is found in five different species preserve the predicted base-pairing (A-U to G-U) with the SL1 RNA, which supports the biological relevance of the predicted structure. Perturbing this base-pairing in addition to mutating more upstream sequences in the 5’UTR resulted in a weaker phenotype (ma461 vs ma472 in Figure 2A&D and S2), which suggests that these first eight nucleotides or their base-pairing with the SL1 sequences has a positive impact on LIN-46 expression.
Sixteen nucleotides that follow the SL1-complementary region constitute a “single-stranded stretch” region (Figure 4A) and all sixteen nucleotides are 100% conserved among all nematode species analyzed here, with the exception of C. sp34 that has a single nucleotide change in this region (Figure 4B). This single-stranded stretch was the region primarily targeted by our CRISPR guide, gR_5U (Figure S1B).Mutations of various sizes (Figure S1D) in this region alone displayed precocious LIN-46 expression (Figure 1A-C, ma459) and strong lin-28-like phenotypes (Figure 2A, ma461). Interestingly, in certain lin-46 5’UTR mutants, such as the ma459, that result in strong precocious phenotypes, the predicted RNA structure is entirely altered (Figure S2B), which may indicate a causative relationship between loss of all structural elements in the lin-46 5’UTR and strong LIN-46 expression.
The last twelve nucleotides in the lin-46 5’UTR contains a GGAG sequence that is located in the stem of a predicted stem-loop structure (Figure 4A, stem-loop). The sequence conservation pattern in this region supports the biological relevance and significance of the predicted structure: 1) a C to T nucleotide change in the stem preserves base-pairing (G-C to G-U), and 2) mutations in the nucleotides in the loop region, which are not contributing to the hairpin stability, seem to be more tolerated (Figure 4B). The GGAG motif is found to be enriched in LIN-28 bound RNA regions [12], however, here the GGAG sequence or the stemloop in the lin-46 5’UTR alone is not sufficient to confer repression of LIN-46 expression (see ma472 in Figure 2A and Figure S2). Moreover, perturbing the GGAG sequence in addition to the single-stranded stretch sequences did not enhance but weakened the precocious phenotypes (ma467 vs ma467 in Figure 2A&D and S3), which suggests that this GGAG-containing loop, rather than an having an inhibitory role, can positively affect LIN-46 expression.
DISCUSSION
Our results provide insights into how the conserved RNA-binding protein LIN-28 regulates its critical mRNA target, lin-46, in C. elegans, and demonstrate that lin-46 mis-regulation is likely responsible for the phenotypes observed in lin-28-null animals. Our results suggest that LIN-28 controls temporal cellfate progression by regulating LIN-46 expression via the 5’UTR of lin-46 mRNA.
The temporally mutually exclusive expression pattern between LIN-28 and LIN-46 (revealed by the endogenously tagged alleles of lin-28 and lin-46) and the effect of loss-of-function of lin-28 on the LIN-46 expression led us to conclude that lin-28 represses LIN-46 expression at early stages (Figure 5). Our results also suggest that the 5’UTR of lin-46 prevents LIN-46 expression, which is likely via mediating LIN-28 binding to and repression of the lin-46 mRNA. We showed that LIN-46 is precociously expressed in lin-28(lf) animals; and with the help of the lin-46 5’UTR mutants, which uncouple precocious LIN-46 expression from the loss-of-function of lin-28, we showed that precocious LIN-46 expression alone is sufficient to suppress L2 cell-fates and to promote precocious transitions to L3 cell-fates. Lastly, endogenously tagged LIN-46 is expressed in the vulva precursor cells (VPCs), which had not been reported before, and lin-28 and lin-46 5’UTR are required to repress LIN-46 expression also in the VPCs. Precocious onset of LIN-46 expression in the VPCs is sufficient to stimulate the precocious onset of vulva development. These results demonstrate that precocious LIN-46 expression in seam cells and VPCs in lin-28(lf) mutants is responsible for the two major heterochronic phenotypes observed in the lin-28(lf) animals; skipping of L2 stage seam cell proliferation and precocious onset of vulva development.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
At the L2 and L3 stages LIN-28 is highly expressed and although the lin-46 mRNA is transcribed it cannot be translated due to LIN-28-mediated inhibition of its translation. Inhibition of LIN-46 expression at the L2 stage permits HBL-1 to function: HBL-1 promotes L2 cell-fates and prevents L3 cellfates. At the L2 to L3 molt, LIN-28 expression is diminished, and this allows LIN-46 expression. LIN-46 opposes HBL-1 activity, thereby prevents expression of L2 cell-fates at the L3 stage.
We hypothesize that lin-46 5’UTR contains a LIN-28-binding element that is required for LIN-28-mediated repression of LIN-46 expression from the lin-46 mRNA. The evidence that supports this hypothesis include: 1) The phenotypic similarities between lin-28(lf) and the 5’UTR mutants of lin-46 reported here; 2) LIN-28 binding to the lin-46 mRNA (including the lin-46 5’UTR) reported previously [12], 3) The existence of a putative LIN-28 interacting sequence, the “GGAG”, in the 5’UTR of lin-46 (Figure 4). However, because previously LIN-28 was shown to interact with the lin-46 mRNA at multiple sites across the entire lin-46 mRNA in addition to the 5’UTR [12], it is surprising that mutations of the lin-46 5’UTR are sufficient to cause a phenotype that is consistent with an almost total loss of LIN-28-mediated regulation of the lin-46 mRNA. Nonetheless, at least two models could reconcile a potential total loss of LIN-28-mediated repression of LIN-46 expression by mutating the 5’UTR of lin-46 alone while leaving other LIN-28 binding sites on the lin-46 mRNA intact. The first model is that the binding of LIN-28 to the lin-46 5’UTR would be required for inhibiting LIN-46 expression whereas LIN-28 binding to other sites on the lin-46 mRNA would not inhibit LIN-46 expression. It is known that translation initiation is highly regulated [43]; and the 5’UTRs harbor sequence elements, such as upstream open reading frames (uORFs), or structural elements, such as highly structured RNA (including G-quadruplexes and pseudoknots) or specific RNA structures that serve as binding sites for RNA-binding proteins [44], which can interfere with or inhibit the translation initiation [45,46]. In the second model, among all the LIN-28 binding sites on the lin-46 mRNA, the lin-46 5’UTR (and particularly the single-stranded stretch region) might have the highest affinity for LIN-28 and might be required to initiate a sequential binding of multiple LIN-28 proteins to the lin-46 mRNA, leading to the formation of a repressive LIN-28-lin-46-mRNA mRNP (messenger ribonucleoprotein) complex. In support of this model, in in vitro assays, LIN28 is shown to preferentially bind to single stranded RNA and more than one LIN28 proteins are shown to bind to long (longer than 30-nts) RNA in a sequential manner after the first LIN28 binds to a predicted single-stranded loop [20].
Precocious cell-fate transition phenotypes observed in lin-46 5’UTR mutants are not as strong as the phenotypes observed in lin-28(lf) mutants. Moreover, the severity of the lin-28(lf)-like phenotypes vary among different lin-46 5 ‘UTR alleles. The severity of these phenotypes does not correlate with the size of the lin-46 5’UTR deletions; and in some cases, larger deletions result in not stronger but more moderate phenotypes (Figre 2). These findings are consistent with a model where the lin-46 5’UTR harbors multiple cis-regulatory elements that can either positively or negatively affect LIN-46 expression. Accordingly, mutants that inactivate a negative regulatory element (the presumed LIN-28 binding site) without disturbing a positive regulatory element result in higher LIN-46 expression and hence stronger precocious phenotypes.
A putative positive regulatory element in lin-46 5’UTR could be the first eight nucleotides of the 5’UTR that is predicted to base-pair with the SL1 sequence (SL1-complementary, Figure 4). The lin-46 mRNA is trans-spliced, which results in the fusion of the SL1 sequence to the 5-prime end of the transcript (Figure S2A and 4). A small stem-loop structure in the SL1 sequence (Figure S2A) has been shown to enhance translation in nematodes [47]. In the predicted folding of the SL1-lin-46-5’UTR chimeric RNA (Figure 4), the nucleotides that form the stem-loop in the SL1 alone base-pair with the first eight nucleotides of the lin-46 5’UTR (Figure S2A). This SL1-5’UTR base-pairing is lost in the lin-46(ma472) (Figure S2B) and the phenotype of this larger deletion is weaker than the two other, smaller deletions (Figure 2A), which is consistent with the idea that base-pairing of SL1 to the lin-46 5’UTR has a positive impact on the translatability of the lin-46 mRNA.
In addition to preventing precocious LIN-46 expression at the L1/L2 stage, LIN-28-mediated regulation of lin-46 mRNA translation might play a role in controlling the level of LIN-46 accumulation at the early L3 stage, when LIN-46 has been shown to compensate for environmentally-induced reduction in let-7-family levels [6]. The C. elegans lin-28-lin-46 pathway acts in parallel to let-7 family microRNAs [7,8] and regulate the nuclear localization, hence the activity, of a critical let-7-family target and a transcription factor, Hunchback-like-1 (HBL-1) [manuscript-=oi2]. Remarkably, precocious LIN-46 expression conferred by the lin-46 5’UTR mutants can fully compensate for the loss of all let-7- complementary sites in the hbl-1 3’UTR [manuscript-oi2], which otherwise causes a severe extra seam cell phenotype due to ectopic HBL-1 activity at L3/L4 stages [manuscript-oi2]. Additionally, lin-46 activity becomes more important in preventing L3-/L4-stage HBL-1 activity at low temperatures [9], or when animals develop through a temporary diapause [48], or merely when animals experience an extended L2 (L2d) in the presence of diapause-inducing pheromones or starvation stress [6]. L2d/Dauer-inducing conditions also result in the repression of let-7-family microRNAs mediated by the nuclear hormone receptor DAF-12 [49,50]. Our recent findings suggest that this repression of let-7-family microRNAs is coupled to a DAF-12-mediated activation of an alternative lin-46-mediated program for HBL-1 downregulation in seam cells [6]. One way to compensate for reduced let-7-family levels by LIN-46 activity would simply be the induction of LIN-46 expression in response to environmental conditions that repress let-7-family microRNAs. Such an induction mechanism that regulates LIN-46 levels at the L3 stage to match the level of let-7-family repression at the L2 stage may utilize LIN-28/5’UTR-mediated regulation of LIN-46 expression as a gate to uncouple lin-46 mRNA accumulation from LIN-46 accumulation, which can provide a control over the rate of LIN-46 accumulation at the early L3 stage (Figure 5B). In this hypothetical model, during a lengthened L2d, lin-46 mRNA may accumulate in proportion to the length of the L2d stage (which is thought to correlate with the severity of the environmental conditions as well as the degree of let-7-family repression). At the L2/L2d stage lin-46 mRNA cannot be translated due to LIN-28-mediated inhibition; however, at the L3 stage, when LIN-28 expression is diminished, the L2/L2d-accumulated pool of lin-46 mRNA would be translated. Thus, LIN-46, expressed from a lin-46 mRNA pool whose size negatively correlates with the reduction in let-7-family levels, can accumulate fast enough to sufficiently inhibit residual or ectopic HBL-1 activity at the post-L2d L3 stage, perfectly compensating for the failure to inhibit the synthesis of HBL-1 at the L2d stage due to repressed let-7 family microRNAs.
In summary, we provide evidence indicating that LIN-28 represses the expression of its critical mRNA target in C. elegans, an intact lin-46 5’UTR is required LIN-28-mediated repression of lin-46 expression, and precocious LIN-46 expression alone is likely responsible for the majority of lin-28(lf) phenotypes. Our findings highlight the biological importance of the mRNA targets of LIN-28 (C. elegans LIN-28 and its orthologs), which may have important functions in regulating pluripotency, reprogramming, or oncogenesis in humans and various other organisms.
3’UTR- and microRNA-mediated mechanisms and their roles in controlling temporal dynamics of gene expression have extensively been studied in the context of the C. elegans heterochronic pathway. However, the involvement of 5’UTRs in the heterochronic pathway was not known and the identities and roles of cis-regulatory elements in the C. elegans 5’UTRs are largely unknown. Here, we identified a critical role for the lin-46 5’UTR in preventing precocious cell-fate transitions in seam and vulval precursor cell lineages. Conservation in the 5’UTRs is not widespread, but interestingly, the 5’UTRs of many heterochronic and developmental genes in C. elegans appear to be evolutionary conserved, which may provide a platform to further explore the functions of mRNA cis-regulatory elements and the roles of transacting RNA binding proteins in regulating stage specific gene expression and developmental progression.
MATERIALS AND METHODS
C. elegans culture conditions
C. elegans strains used in this study and corresponding figures in the paper are listed in Table S2. C. elegans strains were maintained at 20°C on nematode growth media (NGM) and fed with the E. coli HB101 strain.
C. elegans strains used in this study.
Assaying extra seam cell and Pvl phenotypes
The worms were scored at the young adult stage (determined by the gonad development) for the number of seam cells using fluorescence microscopy with the help of the maIs105 [pCol-19::gfp] transgene that marks the lateral hypodermal cell nuclei and/or for protruding vulva phenotype (Pvl) by examining the vulva morphology (as given in Figure 2D).
Each circle on the genotype versus number of seam cells plots shows the observed number of seam cells on one side of a single young adult worm. A minimum of 20 worms for each genotype are analyzed and the average number of seam cells (denoted by lateral bars in the genotype versus number of seam cell plots); percent Pvl values are calculated and represented using a bar graph. The Student’s t test is used to calculate statistical significance when comparing different genotypes. The GraphPad Prism 8 software is used to plot the graphs and for statistical analysis.
Microscopy
All DIC and fluorescent images are obtained using a ZEISS Imager Z1 equipped with ZEISS Axiocam 503 mono camera, and the ZEN Blue software. Prior to imaging, worms were anesthetized with 0.2 mM levamisole in M9 buffer and mounted on 2% agarose pads. The ImageJ Fiji software is used to adjust the brightness and contrast of the images to enhance the visualization of the fluorescent signal. All images are taken using the same microscopy settings and a standard exposure time for all larval stages and genetic background for each reporter (lin-28::gfp and lin-46::mCherry). To enhance the visualization of the fluorescent signals in the figures and to allow comparison of signal intensities in larvae of different genetic backgrounds, fluorescent images of larvae from different backgrounds are stitched together using the ImageJ software and the brightness and contrast of these montaged images were adjusted (in Figure 1B and 3).
Tagging of lin-28 and lin-46 using CRISPR/Cas9
A mixture of plasmids encoding SpCas9 (pOI90, 70 ng/μL), and single guide RNAs (sgRNAs) targeting the site of interest (60 ng/μL of pSW65 for lin-28 or pOI113 for lin-46) and the unc-22 gene (pOI91, 30 ng/μL) as co-CRISPR marker, a donor plasmid (20 ng/μL of pOI173 for lin-28 or pOI167 for lin-46) containing the gfp or mCherry sequence flanked by gene-specific homology arms, and a rol-6(su1006) containing plasmid (pOI124, 30 ng/μL) as co-injection marker was injected into the germlines of ten young adult worms. F1 roller and/or twitcher animals (100-200 worms) were cloned and screened by PCR amplification (Table S3) for the presence of the expected homologous recombination (HR) product. F2 progeny of F1 clones positive for the HR-specific PCR amplification product were screened for homozygous HR edits by PCR amplification of the locus using primers that flanked the HR arms used in the donor plasmid (Table S3). Finally, the genomic locus spanning the HR arms and gfp or mCherry DNA was sequenced using Sanger sequencing. A single worm with a precise HR edited lin-28 or lin-46 locus was cloned and backcrossed twice before used in the experiments.
PCR Primers used in this study.
CRISPR/Cas9-mutagenesis of the lin-46 5’UTR
A mixture of plasmids encoding SpCas9 (pOI90, 70 ng/μL), and gR_5U single guide RNA (sgRNAs) targeting the lin-46 5’UTR (Figure S1; pOI193 60 ng/μL) was injected into the germlines of young adult worms expressing the adult onset gfp transgene (Table S2; VT1357). F1 or F2 animals displaying precocious cell-fate phenotypes, which were consisted of precocious Pcol-19::gfp expression in the seam cells (Figure S2C) and protruding vulva morphology (Figure 2D), were cloned and genotyped for in-del events at the gR_5U targeting site (Figure S2D).
REFERENCES




