

Abstract
The wo protein and its downstream gene, SlCycB2 have been demonstrated to regulate the trichome development in tomato. It was shown that only gain-of-function mutant form of wo, Wov (wo woolly motif mutant allele) could induce the increase of trichome density. However, it is still unclear the relationships between wo, Wov and SlCycB2 in trichome regulation. In this study, we demonstrated Nbwo (NbWov) directly regulated the expressions NbCycB2 by binding to the promoter of NbCycB2 and its genomic sequences. As a feedback regulation, NbCycB2 negatively regulates the trichome formation by repressing Nbwo activity at protein level. We further found that the mutations of Nbwo woolly motif could prevent repression of NbWov by NbCycB2, which results in the significant increase of active Nbwo proteins, trichome density and branches. Our results revealed a novel reciprocal mechanism between NbCycB2 and Nbwo during the trichome formation in Nicotiana benthamiana.
Highlight NbCycB2 is specifically expressed in trichomes of Nicotiana benthamiana and represses the Nbwo activity via a negative feedback loop in tobacco trichome developmemt.
Introduction
Trichomes are the specialized epidermal protuberances locating on aerial parts of nearly all terrestrial plants. They can be classified into various types by cell numbers and shapes -- unicellular/multicellular, and glandular/non-glandular. In Arabidopsis, it has been well-elucidated that the development of trichomes (unicellular and non-glandular), is regulated by the trimeric MYB-bHLH-WDR protein activators complex (GL1 (Oppenheimer et al., 1991)-GL3/EGL3 (Payne et al., 2000)-TTG1(Walker et al., 1999)). This transcriptional complex activates the expression of the homeodomain protein GLABROUS2 (GL2) to induce the formation of trichomes (Rerie et al., 1994; Grebe, 2012). In addition, it triggers the expression of the single repeat R3 MYBs (including TRY (Schnittger et al., 1999), CPC (Wada et al., 1997), ETC1, ETC2, ETC3 (Kirik et al., 2004; Wester et al., 2009) and TCL2 (Gan et al., 2011)) which act as negative regulators of GL3 or EGL3 by forming a repressor complex (GL3/EGL3-TRY/CPC-TTG1) in trichome development (Wang et al., 2008; Wester et al., 2009). Thus the control of trichome development in Arabidopsis requires a regulatory loop including both activators and repressors (Grebe, 2012; Pattanaik et al., 2014).
The trichomes are multicellular glandular (GSTs) structure in approximately 30% of all vascular plants (Glas et al., 2012). Since many phytochemicals and compounds with economical values can be synthesized and secreted by multicellular GSTs (Mauricio and Rausher, 1997; Hollósy, 2002; Valkama et al., 2003; Freeman and Beattie, 2008), multicellular glandular trichomes have considerable economic pontential (Sallets et al., 2014; Huchelmann et al., 2017). However, it has been demonstrated that the networks regulating unicellular trichomes did not work in the development of multicellular trichomes (Serna and Martin, 2006; Yang et al., 2011; Kang et al., 2016; Yan et al., 2016).
In tomato, a HD-ZIP IV transcriptional factor, wo protein has been demonstrated to regulate the trichome initiation (Yang et al., 2011). This HD-ZIP IV member contains four conserved domains including homeodomain domain (HD), the leucine zipper domain (LZ), the steroidogenic acute regulatory protein-related lipid transfer (START) and the START-adjacent domain (SAD). However, overexpression of wo failed to induce the change of trichome density, and only ectopic expression of its gain-of-function mutant alleles, Wov, could cause higher density of trichomes in tomato (Solanum lycopersicum) and tobacco (Nicotiana tabacum) (Yang et al., 2011; Yang et al., 2015). The Wov allele has two point mutations at the C-terminal domain (Since this motif was conserved in the most wo alleles, we name it as woolly motif in this study). The sequence analysis revealed that wo protein is more similar to PROTODERMAL FACTOR2 (PDF2) and the PDF2 redundant protein -- ARABIDOPSIS THALIANA MERISTEM L1 (ATML1), both of which are involved in shoot epidermal cell differentiation (Abe et al., 2001; Ogawa et al., 2015), than GL2 in Arabidopsis.
In Arabidopsis, the ectopic expression of a constitutive active B-type cyclin induced mitotic divisions and resulted in the increase of multicellular trichomes (Schnittger et al., 2005). SlCycB2, a hypothetical B-type cyclin, was reported to directly interact with wo to promote the development of type I trichome (Yang et al., 2011; Yang et al., 2015). Its homologous protein in Arabidopsis (AT5G06270) was also found to interact with GL2 or co-repressor TOPLESS proteins (Wu and Citovsky, 2017b, a). However, as reported in a recent study, overexpression of SICyc82 resulted in non-trichome phenotype, while suppression of SICyc82 promoted trichomes formation in tomato (Gao et al., 2017). These inconsistent results raise the important questions: what is the function of SlCycB2 in trichome formation and why the mutation of woolly motif can promote trichome formation?
Similar to tomato, trichomes in Nicotiana benthamiana are typically multicellular structures, and almost all of the trichomes in N. benthamiana are glandular (Fig. S1), making it a better system for studying glandular trichomes than tomato. In addition, the genome map of N. benthamiana has been constructed (Bombarely et al., 2012). Thus tobacco represents an excellent model plant to study the molecular mechanism of multicellular trichome formation (Goodin et al., 2008). In this study, we cloned the homologues of wo and SlCycB2 in N. benthamiana (named Nbwo and NbCycB2), and constructed a two-point mutantion Nbwo allele, NbWov. To investigate their biological functions in trichome development, we constructed overexpression and suppression transgenic lines of all genes. We demonstrated that Nbwo could positively regulate the expression of NbCycB2 through targeting to the cis-element in NbCycB2 promoter and its genomic DNA sequence. On the other hand, NbCycB2 could be a negative regulator of multicellular trichomes by directly binding and inhibiting Nbwo activity. The previous identified mutation in woolly motif (NbWov) blocked the interaction between NbCycB2 and Nbwo, removing the repression of Nbwo by NbCycB2 and resulting in increased trichome density. Our results revealed the mechanisms of the interaction between Nbwo and NbCycB2 in regulating the development of glandular trichomes.
Materials and Methods
Plant materials and growth conditions
Sterilized seeds of N. benthamiana were germinated and grown to seedlings on MS medium, which solidified with 0.8% (w/v) gellan gum under 26 °C, 14 h light/10 h dark conditions. Two-week-old plants were transferred to other sterilized bottle (for genetic transformation) or soil in pots to grow to maturity. All wild type and transgenic plants were grown in greenhouse under 26 °C, 14 h light/10 h dark condition.
Sequence analysis
The protein sequences of the homologues of wo and SlCycB2 were downloaded from NCBI database (http://www.ncbi.nlm.nih.gov/) and Sol genomic network (Fernandez-Pozo et al., 2015) (https://solgenomics.net/). In order to further analyze the grouping and relatedness, the aligned sequences were used to construct the phylogenetic trees in MEGA 5 by using the maximum-likelihood (ML) criterion with 100 bootstrap analysis. In addition, the relative conservation for each amino acid position in the protein sequences of Nbwo and NbCycB2 was evaluated via Weblogo (https://weblogo.berkeley.edu/) (Crooks et al., 2004), followed by the prediction of their conserved domains in SMART program (https://smart.embl-heidelberg.de/) (Letunic and Bork, 2018).
RNA extraction and Real-time PCR
Total of RNA was extracted from various tissue of plants by using the Eastep®Super Total RNA Extraction Kit (Promega). The cDNA was synthesized from Dnase I treated total RNA using M-MLV 1st Strand Kit (Invitrogen). Real-time PCR (qRT-PCR) was determined by using SYBR Premix Ex Taq II (TaKaRa) and performed on ABI Stepone real-time PCR system (Applied Biosystems). L25 ribosomal protein (L18908) was used as an endogenous control (Schmidt and Delaney, 2010). Relative expression levels were determined as described previously (Guo et al., 2016). Primers are listed in Table S1.
Plasmid construction and N. benthamiana transformation
To investigate the biological functions of wo and SlCyc82 in N. benthamiana, the full-length coding sequences of two alleles of Nbwo and NbCycB2 were amplified with Nhel and BamHI, Xbal and BgIll cloning sites respectively from the general cDNA of leaves (All the primers used in current study are provided in Table S1). In addition, to confirm the function of the tomato Wov gene in tobacoo trichome formation (Yang et al., 2015), an allele NbWov with two point mutations at the locus 2084 (T was replaced with G, result in ile-697 changed to Arg) and 2092 (G was replaced with T, result in Asp-700 changed to Tyr, Fig. S2c) of Nbwo was generated in N. benthamiana by using a Mutagenesis Kit (Toyobo).
To construct the overexpression (OE) lines of Nbwo, NbWov and NbCycB2, these fragments were inserted into pCXSN-HA (Nbwo and NbWov fused with HA tag) and pCXSN-FLAG (NbCycB2 fused with Flag tag) vectors respectively under the control of CaMV 35S promoter (Chen et al., 2009). The suppression expression constructs of Nbwo and NbCycB2 were performed by recombining with the RNAi vector pH7GWIWGII with LR Clonase II enzyme (Invitrogen).
To comprehensively understand the expression sites of NbCycB2, approximate 2880bp upstream promoter fragments were amplified by PCR using the primers shown in Table S1. The promoter fragments were then inserted into the corresponding site of pH2GW7 vector to create the promoter-driven GFP-GUS transformation by using Pro-NbCycB2: GFP-GUS gene fusion (Cui et al., 2015).
All of these constructs were transferred into Agrobacterium tumefaciens strain GV3101, which were used to generate transgenic lines by using Agrobacterium-mediated transformation. The gene expression levels in the transgenic lines were examined by real-time PCR and western blot.
Subcellular localization and tissue distribution
The pCXDG vector (Chen et al., 2009) was used to analyse the subcellular localization of Nbwo, NbWov and NbCycB2, which were fused with GFP driven by the CaMV 35S promoter (p35s:GFP-Nbwo, p35s:GFP-NbWoV, p35s:GFP-NbCycB2). Transformants carrying these constructs were obtained as described above and then transiently transformed into 4-week-old N. benthamiana leaves. After cultivation in lowlight conditions for 48-72 h, GFP was observed using confocal microscopy (LSM 780, Carl Zeiss, Jena, Germany) with staining in DAPI solution (1 mg/ml) for 15 min before observation.
The tissue distribution assays were performed as described in the previous study (Jefferson et al., 1987). GUS staining was repeated at least three independent transgenic lines.
Yeast hybrid assays
Yeast one-hybrid assays were performed to test the specific function area of NbCycB2 promoter binding Nbwo. The promoter of NbCycB2, separated into 5 fragments (E: −1027 ∼ −831 bp, D: −830 ∼ −631 bp, C: - 630 ∼ −411 bp, B: −410 ∼ −201 bp, A: −200 ∼ −1bp, Fig. 2a), were amplified and inserted into the pHIS 2 vector (Clontech) (NbCycB2proE, NbCycB2proD, NbCycB2proC, NbCycB2proB, NbCycB2proA). Further investigation of the targeted sequences in NbCycB2 promoter were conducted by point mutations in the two L1-like boxes in the D fragment (NbCycB2proD-m1, mutant one L1-like box, changed 5’-GCAAATATTTACTC-3’ to 5’-GCGGGTGACTC-3’; NbCycB2proD-m2, mutant two L1-like boxes, changed 5’-GCAAATATTTACTC-3’ to 5’-GCGGGTGACTC-3’, and 5’-ATTTACTC-3’ to 5’-GGGACTCC-3’). To test the specific region of Nbwo genomic sequence binding itself, four genomic fragments of Nbwo (G1, −8 ∼ 251 bp, include T3 fragment; G2, 2169 ∼ 2522 bp, include T4 fragment; G3, 3485 ∼ 3780 bp, include T5 fragment; G4, 4333 ∼ 4660 bp, include T6 fragment, Fig. 7a), were amplified and inserted into the pHIS 2 vector (Clontech) (Nbwo-G1, Nbwo-G2, Nbwo-G3, Nbwo-G4). In addition, the CDSs of Nbwo and NbWov were inserted into vectors pGADT7 containing the GAL4 activation domain (AD) (Clontech). The plasmids were co-transformed into the Y187 yeast strain, empty AD vector was provided as the negative control, and cultivated on SD/-Leu/-Trp (-L-W) medium and tested on SD/-Leu/-His/-Trp (-L-W-H) with 60 mM 3-amino-1,2,4-triazole (Sangon Biotech (Shanghai) Co., Ltd) medium.

The trichome phenotypes of NbCycB2, Nbwo and NbWov transgenic seedlings (a), (b), (c), (d), (e), are the trichomes SEMs of wild type, NbCycB2-RNAi #7 T1, NbCycB2-OE #2 T1, NbWov-OE #1 T1, Nbwo-RNAi #2 T1 10d-old-seedlings respectively. The white bar is 500 µm. (f) The trichome density of the wild type, NbCycB2-RNAi #7 T1, NbCycB2-OE #2 T1, NbWov-OE #1 T1, Nbwo-RNAi #2 T1 10d-old-seedlings leaves were shown. “*” indicates a difference at P<0.05 by Student’s t test compared to WT. “**” represent significant difference against WT at P < 0.01. Error bars represent SD (n = 3).
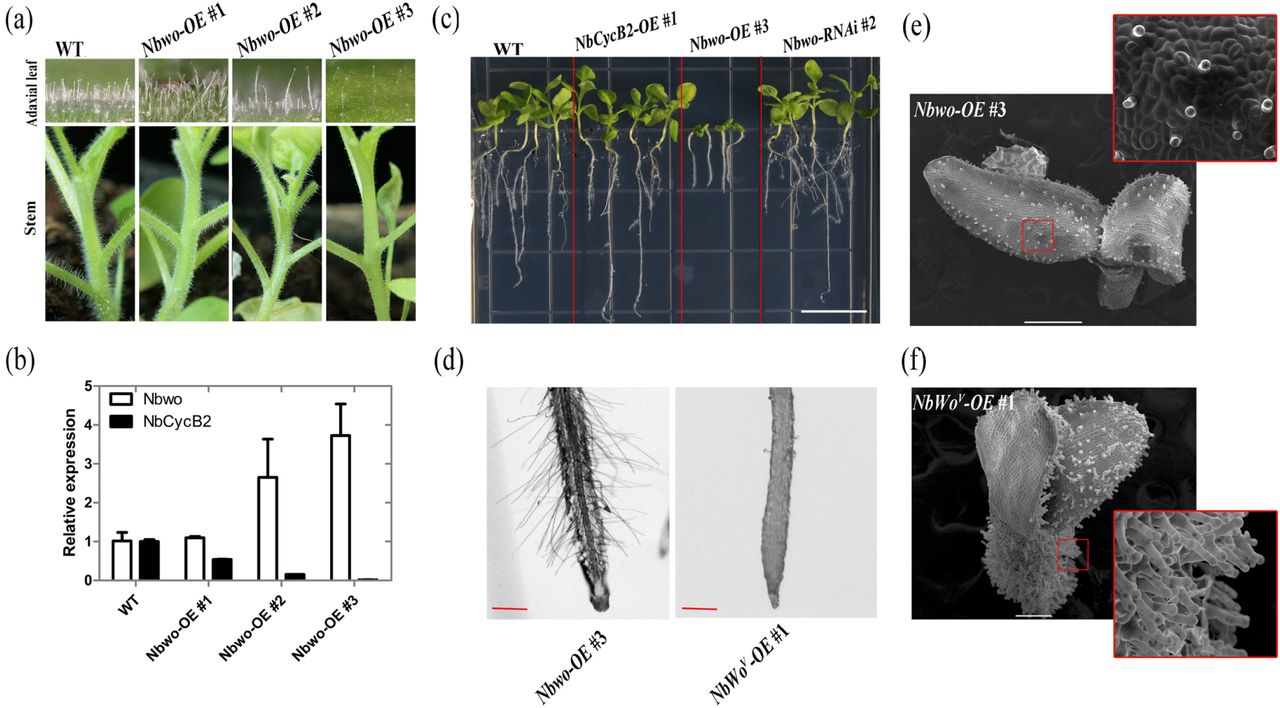
Over expression of Nbwo also induce the dwarf phenotype in N. benthamiana (a) Over expression of Nbwo was drivered by P35S promoter. The trichomes density reduced in the stems and leaves of transgenic lines. (b) The relative expression level of Nbwo andNbCycB2 in the transgenic lines measured by qRT-PCR. Error bars represent SD (n = 3). (c) The root lengths were measured in the Wild type, NbCycB2-OE #1 T1, Nbwo-OE #3 T1, Nbwo-RNAi #2 T1 two weeks-old-seedlings. The white bar is 1 cm. (d) The root hairs of Nbwo-OE #3 T1 and NbWov-OE #1 T1 two weeks-old-seedlings were detected by microscope. The red bar is 1 mm. (e), (f) are the SEMs of the Nbwo-OE #3 T1 and NbWov-OE #1 T1 10d-old-seedlings respectively. The white bar is 500 µm.
The yeast two-hybrid was performed to understand the interaction between Nbwo and NbCycB2. We truncated Nbwo into four segments containing: HD, LZ, START and SAD domains, and fused them into AD vectors to find the target region of the interaction in Nbwo. In addition, the CDS of NbWov was also amplified and inserted into AD vector. Each pair of AD and BD plasmids were co-transformed into the Y2HGold yeast strain. BD-53 and AD-T constructs were co-transformed in to Y2HGold as positive control, and BD-Lam and AD-T as negative control. The transformants were then cultivated on SD/-Leu/-Trp medium (DDO) and tested on SD/-Ade/-Leu/-His/-Trp with 40 mg/L X-a-Gal (QDO/X) or SD/-Ade/-Leu/-His/-Trp with 40 mg/L X-a-Gal and 400 µg/L Aureobasidin A medium (QDO/X/A).
Yeast three-hybrid was conducted to analyze the binding competition between Nbwo LZ domain (Nbwo-LZ) and NbCycB2 to Nbwo. The Nbwo-LZ was fused with BO (Clontech). The NbCycB2 was inserted into the downstream of methionine repressible promoter (PMet25: NbCycB2) in pBridge vector. The plasmid of BD-Nbwo-LZ was transferred with AD-Nbwo as positive control, and empty pBridge vector was transferred to with AD-Nbwo as negative control. The transformants were then tested on SD/-Ade/-Leu/-His/-Trp mediums with different concentrations of methionine (0, 250 µM).
Bimolecular fluorescence complementation Assay (BiFc)
In order to determine the interaction of NbCycB2 and Nbwo (or NbWov) in N. benthamiana protoplasts. The CDSs of Nbwo, NbWov and NbCycB2 were inserted into the pSAT6-cEYFP-C1-B vector (2×35s: YFPc-Nbwo, 2×35s: YFPc-NbWov) and the pSAT6-n(1-174)EYFP-C1 vector (2×35s: YFPn-NbCycB2) separately (Citovsky et al., 2006). Each pair of the two plasmids were then transiently transformed into the protoplasts via PEG–calcium transfection method as described in the previous study (Yoo et al., 2007).
Moreover, to determine the interactions between Nbwo and NbCycB2 in vivo, the CDSs of Nbwo, NbWov and NbCycB2 were fused separately with the C-terminal fragment of YFP in p2YC vector; Nbwo-LZ and NbWov were also fused with the N-terminal fragment of YFP in p2YN vector respectively (Shen et al., 2011). Different plasmid combinations were co-infiltrated into leaves of N. benthamiana as described in previous study (Shen et al., 2011).
The YFP fluorescence was observed by using confocal microscopy (LSM 780, Carl Zeiss, Jena, Germany). Before the observation, the transformed protoplasts were cultivation in 26 °C for 12 h, and the transformed leaves of N. benthamiana were cultivation in darkness for 48-72 h. Three biological repeats were observed independently for each samples.
Dual-luciferase (Dual-Luc) assay
The regulatory effectors of 2×35s: HA-Nbwo, 2×35s: HA-NbWov and 2×35s: Flag-NbCycB2 were generated by using the DNA sequences with Ncol (5’-end) and BgIll (3’-end) cloning sites.
The firefly luciferase reporters were created by inserting the B and D fragments of the NbCycB2 promoter, and the Renilla luciferase was driven by the 35S promoter in the pGreen-0800-II report vector (35S: REN-NbCycB2proB: LUC, 35S: REN-NbCycB2proD: LUC). The mutants of the NbCycB2 promoter D fragments were also constructed using a Mutagenesis Kit (35S: REN-NbCycB2proD-m2: LUC). The regulatory effector and the reporter were used at the ratio of 5:1 or 5:5:1 for the expression test of two or three plasmids.
lmmunoblotting, CO-IP and pull-down assay
The N. benthamiana leaves (∼0.5 g) were homogenized with liquid nitrogen and then solubilized with 0.4 ml of lysis buffer (25 mM Tris-HCI, 2.5 mM EDTA, pH 8.0, 0.05% v/v NP-40, 5% glycerol, 150 mM NaCl, 1 mM phenylmethylsulfonyl fluoride and 20 uM MG132) for 30 min at 4 °C. After solubilization, the protein extract was centrifuged at 13,000g for 10 min at 4 °c to separate the solubilized (supernatant) and non-solubilized material. Total protein (∼so µg) was then used for the immunoblotting assay. After SDS-PAGE separation, the proteins were electrophoretically transferred to a PVDF membrane for immunodetection.
The interaction of NbCycB2 and Nbwo dimers were determined by using co-IP assay in N. benthamiana protoplasts. In the expression vectors, the Nbwo protein was fused to the HA and Flag tags, respectively, and the NbCycB2 was fused to the GFP tag. Each pairs of the plasmids were transformed into the protoplasts via PEG–calcium transfection method as described in the previous study (Yoo et al., 2007). The total proteins were extracted by using lysis buffer and then incubated for 3 h at 4°C with 20 ul of Anti-HA Affinity Gel (Millipore). The immunoprecipitates were washed five times with lysis buffer. The isolated proteins were detected by immunoblotting with anti-Flag or anti-GFP antibodies.
In the pull-down assay, the CDSs of Nbwo, NbWov and NbCycB2 were respectively inserted into the pET22b and PGEX-4T-1 vectors to create the fusion proteins (His-Nbwo, His-NbWov and GST-NbCycB2), and then transformed into the Escherichia coli BL21 strain. The purified recombinant bait proteins (2 mg His-Nbwo or His-NbWov) and 2 mg of prey proteins (GST-NbCycB2) were then mixed with 1 ml binding buffer (SO mM Tris-HCI pH 7.5, 0.6% Triton and X-100, 100 mM NaCl,). After incubation at 4 °C for 2 h, 50 µl of glutathione agarose was added to the mixtures followed by the incubation for additional 1 h. The immunoprecipitates were washed five times with binding buffer. The isolated proteins were detected by immunoblotting with anti-His or anti-GST antibodies.
Chromatin immunoprecipitation (ChIP PCR) assay
Four weeks old NbWov-OE plants were used for ChIP assay as described in previous study (Gendrel et al., 2005). The NbWov proteins were precipitated by using HA antibody (Santa Cruz). Primers were designed to amplify 3 fragments (length ∼ 120 - 210 bp) within the 1.7k bp upstream sequence of the NbCycB2 transcription start site (Fig. 2a), and 7 fragments within ∼ 8.7k bp of genomic DNA sequence of Nbwo (Fig. 7a). After the immuno-precipitation, the purified DNA was analyzed by real-time PCR with the primers of NbCycB2 promoter and Nbwo genomic DNA sequence fragments (Table S1). Enrichment was calculated from the ratio of immuno-precipitated sequences.
Phenotype observation
Images of the transgenic plants stem were obtained using Nikon camera. The observation of abaxial leaves and root hair were used an Axioplan 2 microscope (Carl Zeiss AG).
Leaf of transgenic line and wild type seedlings, which used for scanning electron microscope analysis, were fixed with 2% glutaraldehyde (0.1M phosphate buffer, PH 7.4) at 4 □ for 12h. Then, the samples were dehydrated with a series of alcohol (10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% and 100%) for 20 min each time. Finally, the samples were dried in a critical point drying device (Leica EMCPD030), and coated with gold particles. The samples were observed by using a JSM-6390/LV scanning electron microscope.
Data deposition
The sequences reported in this article have been deposited in the Sol genomic network (Fernandez-Pozo et al., 2015) (https:ljsolgenomics.net/) with the accession numbers as follows: Nbwo (Niben101Scf07790g01007.1), Nbwo-allele (Niben101Scf00176g11005.1), NbCycB2 (Niben101Scf10299g00003.1), NbCycB2-allele (Niben101Scf10396g00002.1), NbML1 (Niben101Scf00703g00003.1), NbML1-allele (Niben101Scf01158g03010.1).
Results
Expression and cellular analysis of Nbwo and NbCycB2
We obtained 14 Nbwo and 8 NbCycB2 protein-related sequences from online databases (Fig. S2a, b). The full-length coding sequences of Nbwo and NbCycB2 obtained in the current study contained 2,199 bp and 333 bp, respectively. One allele of each of the Nbwo and NbCycB2 genes were identified via BLAST with 95.82% and 95.20% identity, respectively. The amino acid analysis determined the two-point mutations at 2,084 and 2,092 of NbWov could cause two amino acid replacements in the woolly motif (Ile to Arg, Asp to Tyr, Fig. S1c). Conserved domain analysis revealed that NbCycB2 contained a WD40-like domain in the N-terminal (NbCycB2-WD40, including an EAR like motif) and a RING-like domain in the C-terminal (NbCycB2-RING) (Fig. S2d). However, no conserved domain of B-Type Cyclin protein was found in NbCycB2 protein sequences.
The visualization of subcellular localization revealed that NbCycB2, Nbwo and NbWov all localized to the nucleus (Fig. S3a). As shown in the self-activation assay, clones of AD-Nbwo and AD-NbWov can grow on QDO/X/A medium compared to the positive control, which means they have strong self-activating ability, while the mutations of woolly motif did not affect Nbwo’s transactivation ability (Fig. S3b).
As shown in Fig. S4a, the spatial expression pattern assays of NbCycB2 and Nbwo indicated they were lowly expressed in roots but at a high level in the trichome containing organs. Further investigation revealed the expression of the NbCycB2 promoter-driven GFP-GUS transgenic lines was only detected in the trichomes of leaves and stems (Fig. S4b-c). However, GUS of the Nbwo promoter-driven GUS transgenic line was strongly expressed in the basal and venous regions of young leaves (Fig. S4d).
NbCycB2 negatively regulates trichome initiation
Most of NbCycB2 overexpression (OE) transgenic lines underwent a dramatic reduction of trichomes on leaves and stems (Fig. S5a, Fig. lc, lf). The root length and number of branch root significantly increased in NbCycB2-OE lines (Fig. S6). Western blot and qRT-PCR analysis showed the NbCycB2 transcripts significantly accumulated in NbCycB2-OE lines, while the expression levels of Nbwo and endogenous NbCycB2 were significantly reduced (Fig. S5b, c).
In contrast, the density of trichomes increased significantly on the leaves and stems of 16 NbCycB2 knockdown lines (NbCycB2-RNAi) (Fig. S5d, Fig. lb, lf). The qRT-PCR analysis showed the number of trichomes negatively correlated with the expression level of NbCycB2 (Fig. S5e), suggesting that NbCycB2 may play a negative role in trichome initiation.
NbWov positively regulates trichome initiation
To confirm the function of Nbwo, we also generated 22 Nbwo knockdown transgenic plants (Nbwo-RNAi). Compared with the wild type, trichome densities were clearly decreased on the leaves and stems of most Nbwo knockdown plants (Fig. S7a, e). The efficiency of RNAi mediated knockdown was confirmed by qRT-PCR in two independent lines, in which the expression of Nbwo and NbCycB2 was significantly reduced (Fig. S7b).
As in the previous study, dramatic increases in the density and branching of trichomes were found on the leaves and stems of NbWov-OE plants (Fig. 1d, 1f, S7d, S7h). Our qRT-PCR assay showed the expression levels of the NbWov, endogenous Nbwo and NbCycB2 genes were all significantly upregulated in transgenic lines (Fig. S7f-g). The dwarfism phenotypes were observed in the T1 plants of NbWov-OE (Fig. S8).
Over-expression of Nbwo also induces the dwarfism
Twenty transgenic plants with overexpression of Nbwo (Nbwo-OE) were generated. Interestingly, the density of trichomes was shown to negatively correlate with the expression level of Nbwo in the T0 of Nbwo-OE plants (Fig. 2a, b). Additionally, the expression levels of NbCycB2 were significantly reduced in trichome reduced plants (Fig. 2b). However, the decreased trichome phenotypes were not observed in T1 plants, but the higher expression level of exogenous Nbwo (for example, Nbwo-OE #3) also resulted in dwarfism similar to NbWov-OE lines (Fig. 2c). Except for the dwarfism, these two lines, however had a great difference in both trichomes and root hairs. In the Nbwo-OE #3, the development of glandular trichomes and the development of root hairs were normal. In NbWov-OE #1, the density of glandular trichomes as opposed to root hairs increased significantly (Fig. 2d-f).
Nbwo and NbWov directly targeted the L1-like box of the NbCycB2 promoter
The expression of NbCycB2 was significantly upregulated in the NbWov-OE lines, and decreased in the Nbwo-RNAi lines (Fig. S7b, f). These results indicated that NbCycB2 was positively regulated by Nbwo and NbWov. To test whether Nbwo directly bound to the promoter of NbCycB2, NbWov-OE (NbWov fused with HA tag) plants were analyzed by ChIP qRT-PCR assay with an HA antibody. Strong enrichment of NbWov was observed in the P2 region of the NbCycB2 promoter in NbWov-OE plants (Fig. 3a, b).

Nbwo and NbWov protein can bind NbCycB2 promoter in vitro and vivo (a) The fragments of NbCycB2 promoter were used to CiHP (P1, P2 and P3) and yeast one-hybrid assays (A, B, C, D and E). The numbers indicate positions of the NbCycB2 promoter truncations. (b) Graphs show the ratio of bound promoter fragments (P1-P3) versus total input detected by qRT-PCR after immuno-precipitation in HA-NbWov by HA antibodies. Data shown are mean ± SE (n = 3). (c) One-hybrid (Y1H) assays were used to determine the interaction of NbCycB2 promoter fragments (A, B, C, D, E) bait constructs and AD-Nbwo or empty pGADT7 constructs in Y187 yeast strains. (d) The schematic diagram of the effectors and reporters constructs were used in the LUC assay. (e) The relative reporter activities were measured in N. benthamiana protoplasts after transiently transformed the effector and reporter constructs. The relative LUC activities normalized to the REN activity are shown (LUC/REN). The difference between combinations were detected by Student’s t test. “**” present that the LUC activity is significantly different (P< 0.01). Error bars represent SD (n = 3).
To further determine the specific area of the NbCycB2 promoter binding to Nbwo, we performed yeast one-hybrid (Y1H) assays. The five truncated fragments of NbCycB2 promoter were shown in Fig. 3a. The yeast colonies containing NbCycB2-proD-pHIS 2 and AD-Nbwo constructs were grown on the selection medium with 3-aminotriazole (60 mM) (Fig. 3c).
To investigate whether Nbwo and NbWov directly affect the expression of the D fragment in vivo, we performed dual-Luc assays. The reporters 35S: REN-NbCycB2proD: LUC, 35S: REN-NbCycB2proB: LUC and the effectors were shown in Fig. 3d. As shown in Figure 3e, each pair of reporter and effector was transiently co-expressed in N. benthamiana protoplasts. Compared to the B fragment, when the Nbwo or NbWov was transiently co-expressed, the D fragment-driven LUC expression accumulated significantly in the protoplasts, indicating that the D fragment of the NbCycB2 promoter should be a specific site for Nbwo and NbWovbinding.
Further analysis of the targeting sequence of NbCycB2proD revealed this sequence contained two L1-like boxes (5’-ATTTACTC-3’) (Fig. 4a). In the Y1H assay and the in vivo LUC assay, when two L1-like boxes were mutated (NbCycB2proD-m2), the interaction with the Nbwo protein is abolished (Fig. 4b, c). Based on these results, we inferred the L1-like boxes may be the binding target of the Nbwo and NbWov proteins.
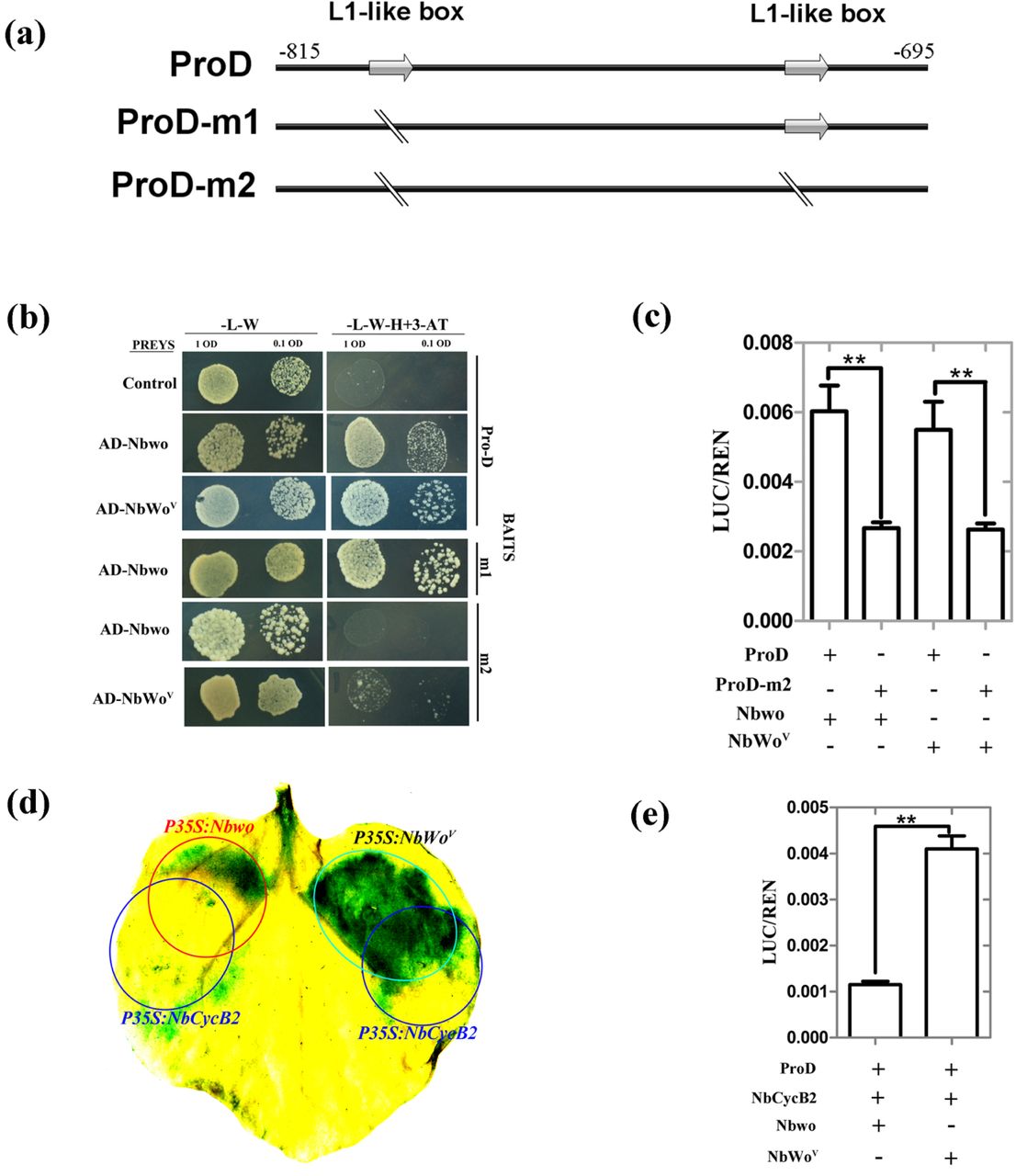
NbCycB2 can suppress the function of Nbwo, but has no effect on NbWov (a) The DNA sequences of wild type NbCycB2 promoter (ProD) and mutant fragments were shown. L1-like boxes were indicated by using gray arrow. The mutated bases in the m1 and m2 sequences are shown by a dividing line. (b) One-hybrid (Y1H) assays were used to detected the interaction between mutant D fragments of NbCycB2 promoter bait constructs and AD-Nbwo or AD-NbWov in Y187 yeast strains, the empty pGADT7 construct as the control. (c) Relative reporter activity was measured in N. benthamiana protoplasts after transiently co-transformed the effector and reporter constructs. The relative LUC activities normalized to the REN activity are shown (LUC/REN). “**” present that the LUC activity has significant difference between the D and m2 fragments reporter when their co-transformed with Nbwo or NbWov effector respectively (P< 0.01, Student’s t test). Error bars represent SD (n = 3). (d) The GUS staining of the proNbCycB2: GFP-GUS transgenic line leaves, when co-expressing P35S: Nbwo and P35S: NbCycB2, P35S: NbWov and P35S: NbCycB2. The area of red circle was injected with P35S: Nbwo GV3101 strain; the area of Green circle was injected with P35S: NbWov GV3101 strain; the area of blue circles were injected with P35S: NbCycB2 GV3101 strain. (e) The LUC activity of co-transforming the effector and reporter constructs were measured. “**” present that the LUC activity is significantly different between Nbwo and NbWov effector when they co-transformed with NbCycB2 effector respectively (P< 0.01, Student’s t test). Error bars represent SD (n = 3).
NbCycB2 represses the activity of Nbwo rather than NbWov
Since the NbCycB2-OE and Nbwo-RNAi transgenic lines shared the non-trichome phenotype, and the expression of Nbwo was not enhanced in the NbCycB2-RNAi lines (Fig. S5e), we suspect that NbCycB2 could affect Nbwo transactivation ability at protein levels. To confirm this, constructs of overexpression Nbwo, NbWov and NbCycB2 were transiently co-expressed in the leaves of the NbCycB2pro: GFP-GUS transgenic line using the Agrobacterium-mediated method (Fig. 4d). The GUS induced by the transient expression of Nbwo-OE was only detected in the area where there was no NbCycB2-OE Expression. However, the transient expression of NbWov caused a strong GUS staining which is independent of NbCycB2-OE expression. These results were further supported by the LUC assay, in which co-expression with NbCycB2 could clearly repress the transactivation activity of Nbwo, but did not affect the NbWov protein (Fig. 4e). These results suggest that NbCycB2 may act as a negative regulator of Nbwo rather than NbWov.
NbCycB2 suppresses the transactivation ability of Nbwo via the direct interaction
The interaction between NbCycB2 and Nbwo was reported in a previous study (Yang et al., 2011). To explore which domain was involved in the physical interaction between NbCycB2 and Nbwo, four truncated fragments of Nbwo containing HD, LZ, START and SAD were used (Fig. 5a). Y2H assays suggested that the LZ domain of Nbwo interacted with NbCycB2 (Fig. 5b). The BiFC assay was further used to verify the interaction between the Nbwo LZ domain and NbCycB2 in vivo (Fig. 5c).

NbCycB2 protein interact with LZ domain of the Nbwo protein in vitro and vivo (a) Schematic diagrams of Nbwo protein domain constructs. The numbers indicate positions of the first and the last amino acid of the Nbwo truncations. (b) Interaction between NbCycB2 and domains of Nbwo protein were determined by YH2 system. (c) The interaction between Nbwo and NbCycB2 or itself were demonstrated by BiFC assay. Each indicated pair of constructs were co-infiltrated into leaves of N. benthamiana (Bar, 50 µm).
To further examine if NbCycB2 also interacts with NbWov, we used yeast two-hybrid (Y2H) assays. Our results indicate that NbCycB2 physically interacts with Nbwo but did not interact with NbWov, suggesting that the interaction between NbCycB2 and Nbwo can be reduced by mutations in the woolly motif (Fig. 6a). Consistent with the Y2H result, the interaction between NbCycB2 and Nbwo or NbWov was further confirmed by the pull-down assay (Fig. 6b) and the bimolecular fluorescence complementation (BiFC) assay (Fig. S10a).

Mutation of woolly motif reduce the interaction between Nbwo and NbCycB2 (a) Interaction between NbCycB2 and Nbwo or NbWov proteins were determined by YH2 system. Blue clones grown on the QDO/X/A medium indicates positive protein-protein interactions. (b) The pull down assay between Nbwo or NbWov with NbCycB2 proteins. Only recombinant HIS-Nbwo protein can co-precipitate with GST-NbCycB2 protein. (c) The competitive binding between NbCycB2 and LZ domain, LZ domain and Nbwo were determined by Yeast three-hybrid assays. (d) The co-IP assay between NbCycB2 and Nbwo dimers. The total protein immunoprecipitation was performed by using anti-HA beads.
Nbwo was restrained by NbCycB2 through forming a homodimer
It is known that the HD-Zip proteins bind to DNA as dimers by the LZ domain (Ariel et al., 2007). To verify whether Nbwo can be dimerized via the LZ domain, we first demonstrated in the Y2H that the LZ domain could bind to the Nbwo protein (Fig. S10b). In addition, BiFC assays were used to confirm the interaction between the LZ domain and Nbwo (or NbWov) proteins in vivo (Fig. 5c).
To test whether the NbCycB2 could bind at the Nbwo homodimer, we conducted the yeast three-hybrid (Y3H) and co-IP assays. As shown in Fig. 6c-d, NbCycB2 could interact with the Nbwo dimers.
Nbwo could bind to its own genomic DNA
The endogenous expression level of Nbwo reduced in NbCycB2-OE lines (Fig. S5b) and increased in NbWov-OE plants (Fig. 1d), indicated that Nbwo may be able to regulate self expression. In order to prove this hypothesis, ChIP assay was carried out to check whether Nbwo could bind to its genomic DNA sequence in the leaf of NbWov-OE transgenic line. Interestingly, enrichment of NbWov was found in the T5 fragment in NbWov-OE plants (Fig. 7a, b). This result was further demonstrated by using Y1H assay. Only the clones with AD-Nbwo (or AD-NbWov) and Nbwo-G3-pHIS 2 constructs could grow on the resistant medium, suggesting that Nbwo or NbWov could bind to the G3 fragments (include T5 fragment, Fig. 7a) of its own genomic DNA sequences (Fig. 7c).

Nbwo and NbWov can bind at itself genomic DNA sequences (a) The fragments of Nbwo genomic sequences were used to CHiP (T1, T2, T3, T4, TS, T6 and T7) and yeast one-hybrid assays (G1, G2, G3 and G4). The bar present 1 kb DNA sequences. (b) Graphs show the ratio of bound genomic fragments (T1-T7) versus total input detected by real-time PCR after immuno-precipitation in NbWov-OE lines by HA antibodies. Data shown are mean ± SE (n = 3). (c) One-hybrid (Y1H) assays were used to determine the interaction of Nbwo genomic sequence fragments (G1, G2, G3, G4) bait constructs and AD-Nbwo, AD-NbWov or empty pGADT7 constructs in Y187 yeast strains. The clones grown on the SD/-Leu/-His/-Trp (-L-W-H) with 60 mM 3-AT medium indicates the interaction between DNA fragment and Nbwo or NbWov proteins.
Overexpression of NbCycB2 can reduce the dwarf phenotype of Nbwo-OE plants
To determine whether NbCycB2 can inhibit the transactivation activity of Nbwo in vivo, we crossed NbCycB2-OE #2 T1 plant to Nbwo-OE #3 T0 plants. As shown in Fig. 8 a-b, the dwarf and short-root phenotypes of Nbwo-OE #3 were indeed reduced by the NbCycB2-OE #2. The crossed F1 plants were tested using PCR (Fig. 8 c), and the expression of NbCycB2 and Nbwo was also verified by qRT-PCR. Compared with T1 Nbwo-OE #3 plants, the balance between NbCycB2 and Nbwo expression was restored in NbCycB2-OE #2 and Nbwo-OE #3 crossed F1 plants (Fig. 8 d).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Hybridization between Nbwo-OE and NbCycB2-OE plants (a), (b) The phenotype of NbCycB2-OE #2 and Nbwo-OE #3 hybridization F1 two weeks old (a) and mature (b) plants were shown. “I” present wild type N. benthamiana, “II” present NbCycB2-OE/WT hybrid T1 plants, “III” present NbCycB2-OE #2 and Nbwo-OE #3 hybridization F1 plants, “IV” present Nbwo-OE #3/WT hybrid T1 plants. (c) F1 plants were tested by using PCR. Wild type as a negative control, Nbwo-OE #3 and NbCycB2-OE #2 as positive controls. No DNA band was detected in the wild type N. benthamiana. A ∼330 bp DNA band was detected in NbCycB2-OE #2 lines. A ∼2199 bp DNA band was detected in Nbwo-OE #3 lines. In contrary, two bands were detected in NbCycB2-OE #2 and Nbwo-OE #3 hybridization F1 plants. (d) The relative expression levels of Nbwo and NbCycb2 were measured by qRT-PCR in F1 plants. Data are given as means SD (n = 3).
Discussion
NbCycB2 negatively regulates trichome initiation
It is well known that ectopic expression of a constitutive active B-type cyclin promoted the unicellular trichome differentiate to multicellular in Arabidopsis (Schnittger et al., 2002, 2005). The SlCycB2 gene has been reported to be a hypothesis B-type cyclin gene participating in trichome formation in tomato (Yang et al., 2011; Gao et al., 2017). However, the function of SlCycB2 during trichome development has not been well-studied.
In this study, we found that overexpression of NbCycB2 caused a non-trichome phenotype, whereas inhibition of NbCycB2 significantly increased the density of trichomes instead of branching on stems and leaves (Fig.1b, S5). Consistent with the qRT-PCR results (Fig. S4a), GUS staining assay indicated NbCycB2 is specifically expressed in the trichomes of leaves and stems (Fig. S4b-c), and Nbwo is expressed in the basal and venous regions of young leaves (Fig. S4d). These results suggested NbCycB2 serves as a negative regulator of trichome initiation. None of B-type cyclin conserve domains were found in the SlCycB2 and NbCycB2 protein sequences (Fig. S2d). Thus, whether SlCycB2 has the function of B type cyclin protein requires further study.
NbWov and Nbwo directly regulate the expression of NbCycB2 through binding to the Ll-like boxes in the promoter
In previous study, SlCycB2 has been reported to be indirectly regulated by Wov (Yang et al., 2011; Yang et al., 2015). The expression level of NbCycB2 was upregulated in overexpression of NbWov plants and downregulated in Nbwo-RNAi lines (Fig. S7), indicating that NbCycB2 may be the downstream gene of Nbwo. Additionally, the D fragments of the NbCycB2 promoter have been shown to be the binding target of Nbwo and NbWov through ChIP, Y1H and LUC assays (Fig. 3). Mutation of the two L1-like box sequences in NbCycB2proD inhibited the binding of Nbwo or NbWov in vitro and vivo (Fig. 4b). Therefore, we proved that NbCycB2 was directly regulated by Nbwo or NbWov through the binding of L1-like boxes in the NbCycB2 promoter (Fig. 4a-c). In addition, we also proved that Nbwo and NbWov can self-regulate the endogenous expressions by binding to its own genomic DNA sequence by ChIp and YIH assay (Fig. 7).
The increase of trichome density and plant dwarfism was regulated by Nbwo through different pathways
The Wo homozygous plants have been shown to cause the embryo lethality in seeds (Yang et al., 2011). We also found the overexpression of Nbwo or NbWov genes causes abnormal embryonic development and results in dwarf phenotype in its offspring (Fig. 2c, S6). However, the trichome phenotype of Nbwo-OE and NbWov-OE was completely different (Fig. 2e and 2f), suggesting that Nbwo was involved in both the regulation of the development of trihcome and embryo. The trichomes’ density was decreased with the expression of Nbwo in T0 of Nbwo-OE lines (Fig.2a and 2e), which suggested that high expression of Nbwo in wild type background could repress trichome development. Thus, we suspected the woolly motif represses Nbwo activity. However, the trichome density was not increased when the SAD domain deleted Nbwo CDS (including woolly motif) was overexpressed in the wild type plants (Fig. S9), which suggesting that woolly motif did not have repressing activity. Therefore, the decreased trichome density in high expression of wo gene plants requires further investigation.
NbCycB2 represses the transactivation activity of Nbwo at protein level
The NbCycB2 gene was proven to be directly regulated by Nbwo in our study. However, the similar non-trichome phenotypes of NbCycB2-OE and Nbwo-RNAi transgenic lines were found (Fig. S5a, S7a). Additionally, the expression of Nbwo was not increased in NbCycB2-RNAi plants (Fig. S5e). These results suggested that NbCycB2 might repress the transactivation activity of Nbwo at protein levels. The GUS activity of the NbCycB2pro: GFP-GUS transgenic line was upregulated by the expression of Nbwo and inhibited by the co-expression of NbCycB2 (Fig. 4d), and the same result was found in the LUC assay (Fig. 4e). Additionally, Hybridization with NbCycB2-OE can attenuate the dwarf phenotype of T1 Nbwo-OE (Fig. 8a-b). Further studies revealed that the expression of endogenous NbCycB2 and Nbwo were reduced in the NbCycB2-OE lines (Fig. S5b). They were shown to be downstream regulatory genes of Nbwo (Figures 3 and 7). These results altogether supported our hypotheses that NbCycB2 may act as a negative regulator of Nbwo at protein levels.
In a previous study, SlCycB2 was reported to interact with wo protein (Yang et al., 2011). Further investigation of the interaction between Nbwo and NbCycB2 revealed the dimerized LZ domain of Nbwo binds to NbCycB2 (Fig. 5b, c). Through Y2H, BiFC, Y3H and co-IP assays, we also found that Nbwo could form a homodimer through the LZ domain, and the NbCycB2 protein could bind to LZ domain of Nbwo dimers (Fig. 5b-c, 6 c-d, S10b). These results indicated that NbCycB2 may bind to Nbwo protein via its LZ domain to form a complex, which inhibits its transactivation ability. However, further study is required to determine whether NbCycB2 functions similarly to its homologue gene --AT5G06270.1 which also interacts with the co-repressor TOPLESS (Long et al., 2006; Szemenyei et al., 2008; Pauwels et al., 2010; Wu and Citovsky, 2017a).
The interaction between Nbwo and NbCycB2 was blocked by the mutation in the Nbwo woolly motif
In Arabidopsis thaliana, the feedback loop regulation mechanisms of the R3 MYB (TRY, CPC and so on) is through competitively binding to GL3/EGL3 to form a non-functional trimeric protein complex (MYB-bHLH-WDR), inhibiting the formation of trichomes (Wang et al., 2008; Wester et al., 2009). Feedback loop regulation has been reported as an effective strategy to maintain normal organism development by many HD-ZIP proteins (Ohgishi et al., 2001; Williams and Fletcher, 2005; Kim et al., 2008; San-Bento et al., 2014). However, the trichome formation was not repressed by the high expression level of NbCycB2 in the NbWov-OE plants (Fig.1d, S7d, f), which suggest that the negative effect of NbCycB2 could be eliminated by the mutation in NbWov. The LUC assay and GUS activity assay in the leaves of NbCycB2pro: GFP-GUS transgenic lines also supported this conclusion (Fig. 4d, e).
Further investigation demonstrated the interaction between NbCycB2 and Nbwo could be blocked by the mutation of woolly motif in NbWov protein in vitro and in vivo (Fig. 6a-b, S10a), indicating that NbWov abolishes interaction with NbCycB2 to prevent inhibition of NbCycB2. The high expressions of NbCycB2 and endogenous Nbwo in NbWov-OE lines also supported this conclusion (Fig. S7f).
In summary, through this study, we found NbCycB2 is specifically expressed in the trichomes of N. benthamiana and negatively affects trichome formation. Further study revealed the Nbwo and NbWov were demonstrated to directly regulate NbCycB2 and Nbwo expressions by binding to the L1-like box in the NbCycB2 promoter and its own genomic DNA sequences. In addition, the NbCycB2 protein may via binding to the LZ domain of Nbwo dimers, which represses the activity of Nbwo and reduces the expression of Nbwo downstream genes, eventually leading to the inhibition of trichome initiation. By contrast, the interaction between NbCycB2 and Nbwo could be blocked by the mutation in woolly motif (NbWov), which prevents the repression by NbCycB2, and results in the dramatic increase of trichome density and branching. In previous studies, since SlCyc82 is highly expressed in the Wo (wo gain-of-function mutant alleles) and Wov lines and underexpressed in the Wo-RNAi lines, SlCycB2 is believed to promote the development of type I trichomes in tomato (Yang et al., 2011). However, the function and detailed molecular mechanisms of NbCycB2 in regulating trichome development have not been studied. Our findings provide further insights into the regulatory network of NbCycB2 and Nbwo or NbWov in multicellular trichomes development of N. benthamiana (Fig. S11).
Author contributions
Liang Chen, Hong Cui, Shuang Wu and Minliang Wu conceived and designed the experiments. Minliang Wu, Yuchao Cui, Li Ge, Lipeng Cui, Zhichao Xu, Hongying Zhang, Zhaojun Wang and Dan Zhou performed the experiments and analysed the results. Minliang Wu and Shuang Wu wrote the main manuscript text. Liang Chen supervised the project.
Supplemental Data
Supplemental Fig. S1: The Scanning electron micrographs (SEMs) of trichomes in the leaf of N. benthamiana.
Supplemental Fig. S2: Sequence analysis of Nbwo, NbCycB2 and their similar proteins.
Supplemental Fig. S3: The subcellular localization and auto activation test of NbCycB2, Nbwo and NbWov.
Supplemental Fig. S4: The expression pattern of NbCycB2 and Nbwo in N. benthamiana.
Supplemental Fig. S5: Overexpression of NbCycB2 and RNA interference of NbCycB2 in N. benthamiana.
Supplemental Fig. S6: The root phenotypes of wild type, NbCycB2-RNAi #7 T1, NbCycB2-OE #2 T1, NbWov-OE #1 T1, Nbwo-RNAi#2 T1 seedlings
Supplemental Fig. S7: RNA interference of Nbwo and overexpression of NbWov in N. benthamiana.
Supplemental Fig. S8: The phenotype of NbWov-OE lines.
Supplemental Fig. S9: The phenotype of overexpressing Nbwo-SAD-mutant in N. benthamiana.
Supplemental Fig. S10: The interaction between Nbwo and NbCycb2, Nbwo and Nbwo LZ domain.
Supplemental Fig. S11: A simplified model for regulation between Nbwo and NbCycb2.
Supplemental Table S1: Primers used in this study.
Acknowledgements:
We would like to thank Prof. Xi Huang for kindly providing pBridge and pGreen-0800-II report vectors and thank Dr. Yongjia Zhong, Fujian Agriculture and Forestry university, for kindly providing p2YN and p2YC vectors, and thank Prof. Tao Huang, Prof. Yi Tao and Prof. Hongrui Wang for the help in the implement of experiments. The authors declare no competing financial interests. This study was supported by the State Tobacco Monopoly Administration of China [grant No. 110201401003 (JY-03)], XMU Training Program of Innovation and Enterpreneurship for Undergraduates (2016Y0635) and Guizhou Science and Technology Major Project [Grant No: (2019)3001-2].
Reference




