

ABSTRACT
Climate change is altering light regimes in lakes, which should impact disease outbreaks, since sunlight can harm aquatic pathogens. However, some bacterial endospores are resistant to damage from light, even surviving exposure to UV-C. We examined the sensitivity of Pasteuria ramosa endospores, an aquatic parasite infecting Daphnia zooplankton, to biologically relevant wavelengths of light. Laboratory exposure to increasing intensities of UV-B, UV-A, and visible light significantly decreased P. ramosa infectivity, though there was no effect of spore exposure on parasitic castration of the host. P. ramosa is more sensitive than its Daphnia host to damage by longer wavelength UV-A and visible light; this may enable Daphnia to seek an optimal light environment in the water column where both UV-B damage and parasitism are minimal. Studies of pathogen light sensitivity help us uncover factors controlling epidemics in lakes, which is especially important given that water transparency is decreasing in many lakes.
INTRODUCTION
Changing temperature and precipitation related to climate change are altering disease dynamics. One factor that plays a role is declining water transparency, since ultraviolet light penetration into lakes has germicidal effects (Williamson et al. 2017). Thus, by decreasing light penetration in lakes, climate change has the potential to promote epidemics.
However, while we know that many microbes are harmed by exposure to light, we also know some tolerate light remarkably well. Endospores, a resting stage found only in Gram positive bacteria of the group Firmicutes, are highly resistant to disinfecting techniques (Nicholson et al., 2000), including high levels of UV-C radiation (Newcombe et al., 2005). Surprisingly though, in the endospore form, the highly studied Bacillus showed decreased survival across a wide range of exposures, from UV-B to full sunlight (Xue and Nicholson, 1996). Other pathogens have shown a similar sensitivity to longer wavelengths of light. For example, an aquatic pathogen - the fungus Metschnikowia -was sensitive to solar radiation even in the absence of UV, and field surveys showed larger epidemics in less transparent lakes (Overholt et al., 2012).
Light can harm pathogens, and climate change is altering light regimes in lakes. Thus, investigations into how aquatic pathogens and their hosts respond to light are needed to better understand and predict disease dynamics. Here we test whether endospores of the virulent bacterial pathogen Pasteuria ramosa are sensitive to biologically relevant wavelengths of light, and if light exposure decreases its ability to lower fecundity in infected Daphnia hosts.
METHODS
We exposed P. ramosa to different environmentally realistic light conditions in the laboratory and measured subsequent pathogen infectivity and host reproduction in Daphnia dentifera. Shallow quartz dishes containing 25 mL aliquots of P. ramosa spores (2000 spores mL−1) were placed on a rotating wheel (2 rpm) for 12 h at 24°C in a UV-lamp phototron and exposed to different levels of biologically relevant UV-B, and photorepair radiation (PRR, comprised of UV-A and visible light), which stimulates repair of UV-damaged DNA (Williamson et al., 2001). In experiment 1, we examined the infectivity of P. ramosa under 10 intensities of PRR (8 replicates per treatment). Experiment 2 used a two-way factorial design to measure the effects of light wavelength and intensity on pathogen infectivity and host fecundity. Spores were exposed to either UVB and PRR or visible light only at nine intensity levels. For the exposure, spores (2000 spores mL−1) were aliquoted into five replicates of each treatment (intensity x light type). (See supplement for additional details.)
Following exposure in the phototron, dishes were removed and a single, week-old D. dentifera neonate was placed in each dish with the exposed P. ramosa spores. After three days, D. dentifera were transferred to 50 mL beakers and filled with 30 mL of spore-less, filtered lake water. In both experiments, offspring were removed during water changes. In experiment 2, neonates were quantified during water changes and eggs in the brood chamber counted on day 25. After 25 days, individuals were examined for infection.
We used a binomial logistic regression model to test the effects of increasing light intensity on infectivity for each light treatment. A two-way ANOVA was used to test the effect of light treatment and intensity on the number of neonates produced. Analyses were conducted using R version 3.4.4 (R Core Development Team).
RESULTS
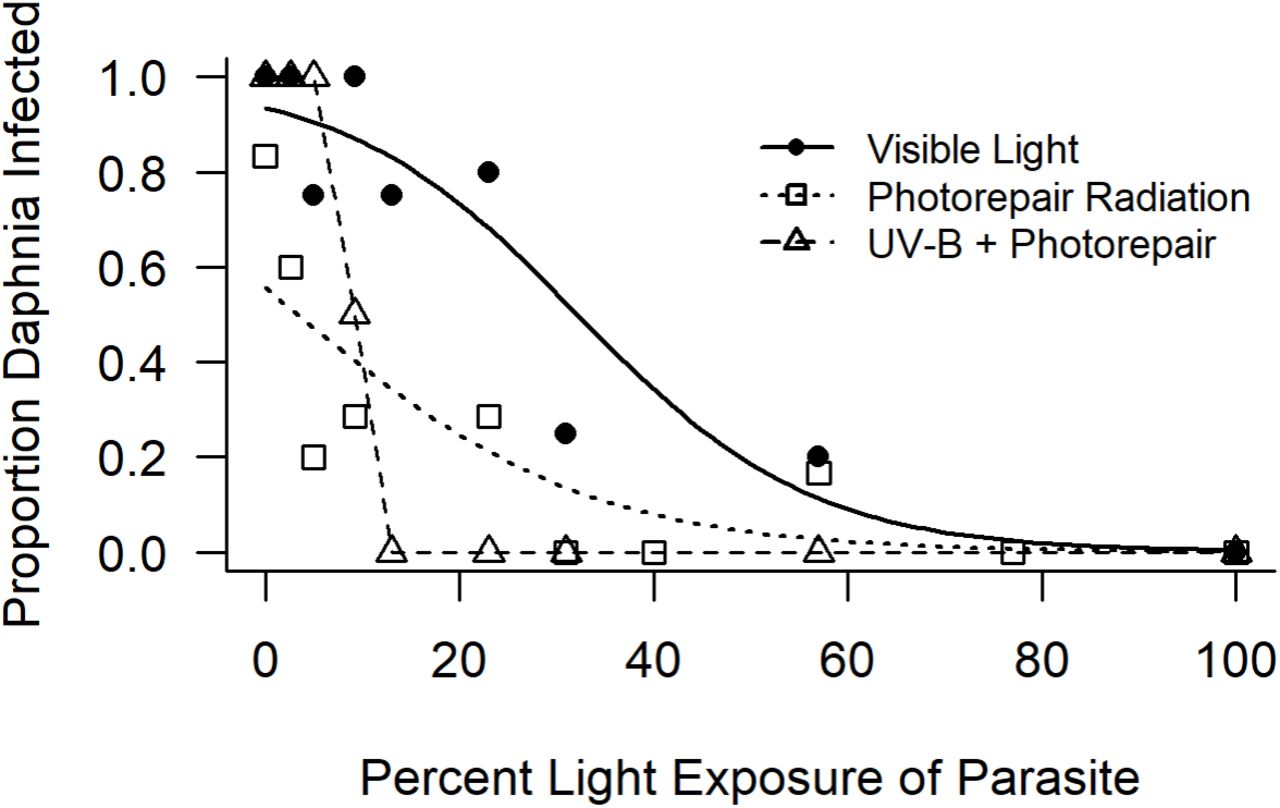
Light exposure greatly decreased parasite infectivity: In both experiments, the highest rates of infection were in the dark (0% light) and decreased with increasing light exposure of the pathogen (Fig. 1). The fecundity of Daphnia also increased with increasing exposure of P. ramosa to light, but, for hosts that became infected, there was no difference in the number of neonates produced (Fig. 2).

Proportion of Daphnia infected at each exposure level of the pathogen to visible light, photorepair radiation, or UV-B + photorepair. Lines represent the fitted logistic regression model for the visible light and photorepair radiation treatments. When UV was blocked, the proportion of D. dentifera infected significantly decreased with increasing exposure of the pathogen (experiment 2 Visible Light, p < 0.01); the logistic model fit indicated that there was an 8.0% decreased probability of infection for every 1% increase in light exposure. We also found a significant negative relationship between light intensity and infectivity when the pathogen was exposed to increasing levels of photorepair radiation (experiment 1 Photorepair Radiation, p < 0.01); the logistic model indicated that for every 1% increase in PRR exposure the probability of D. dentifera infection decreased by 6.5%. When exposed to the full spectrum of light (experiment 2 UV-B + Photorepair), there was also a decline in infectivity; this model would not converge due to sharp cut off in proportion infected, so the line does not represent the fitted model, but instead simply connects the points.

Fecundity (total neonates plus eggs on day 25) of infected (triangles) and uninfected (circles) Daphnia in experiment 2. Overall, Daphnia fecundity increased when spores were exposed to higher intensities (intensity p < 0.001, light treatment p = 0.03), but, for hosts that became infected, there was no difference in the number of neonates produced in the different treatments (intensity p = 0.37; light treatment p= 0.57). The lines represent linear regressions with shared 95% confidence intervals for infected and uninfected individuals.
DISCUSSION
We found that endospores of P. ramosa are surprisingly susceptible to longer wavelength UV-B, UV-A and even visible light. This influence of light on spores benefitted Daphnia via impacts on infection: hosts that were exposed to spores that had been exposed to more light were less likely to become infected and, therefore, produced more offspring. However, when just infected hosts were considered, spore light exposure did not alter host reproduction.
Other aquatic pathogens are sensitive to longer wavelengths of light as well. For example, Cryptosporidium cysts lost infectivity following exposure to UV (Connelly et al., 2007; King et al., 2008) and visible light (Connelly et al., 2007). Natural sunlight caused additional sublethal effects on the protein secretion required by Cryptosporidium for attachment to its host (King et al., 2010). The fungal parasite Metschnikowia, which can be found in the same lake systems as P. ramosa, was also sensitive to both short wavelength UV-B as well as longer wavelength UV-A and visible light in both laboratory and field studies (Overholt et al., 2012). Another field study suggested that Pasteuria was susceptible to solar radiation; however sensitivity to visible light was not specifically tested (C. L. Shaw, unpublished data).
In our study, P. ramosa exhibited decreased infectivity even under long wavelength UV-A and visible light, in the absence of UV-B, though shorter wavelengths overall caused the greatest decrease in infectivity. In contrast, these longer wavelengths benefit the host, Daphnia, by stimulating photorepair of DNA damage (Sancar, 1994). Since the same wavelengths that damage the pathogen can benefit the host, Daphnia may be able to find a refuge from disease and damaging UV-B at intermediate depths in the water column where UV-A and visible light levels are high, but damaging UV-B is less intense.
CONCLUSIONS
Lakes in many regions are experiencing lower intensity light regimes due to increased dissolved organic matter inputs and/or eutrophication (Monteith et al., 2007; Solomon et al., 2015; Strock et al., 2017; Williamson et al., 2017; Williamson et al., 2015). Our finding that the common bacterial pathogen P. ramosa is sensitive to both UV and visible light suggests that decreases in lake transparency through “browning” and/or “greening,” may allow for increased P. ramosa prevalence.
SUPPLEMENTARY DATA
Supplementary data will be available online after publication.
FUNDING
This work was supported by the US National Science Foundation [grant number NSF-DEB 1305836 to M.A.D.] and Miami University.
SUPPLEMENTAL METHODS
D. dentifera were isolated from Bishop Lake (Livingston County, MI, USA) and maintained on a diet of Ankistrodesmus falcatus at 24°C on a 16:8 light: dark cycle throughout the experiment. A single clone of the host species was used to standardize the susceptibility of the host organism to the bacterial P.ramosa strains.
PRR was supplied by two 40-W UV-A bulbs (Q-Lab QUV UV-A, Q-Lab, Cleveland, OH) and two fluorescent, 40 W, cool white tube bulbs. Light intensity was manipulated using mesh screens as neutral density filters. A total of ten intensity treatments, with eight replicates each, were used in experiment 1 (0%, 2.6%, 5%, 9.2%, 23%, 31%, 40%, 57%, 77%, 100%).
In experiment 2, PRR and UV-B were crossed factorially. Covering the UV-B lamp (Spectronics XX15B, Rochester, New York) with acetate removed wavelengths less than 295 nm, allowing only biologically relevant wavelengths to be transmitted. The addition of light filters on individual dishes either transmitted UV (creating the “UV-B + photorepair” treatment) or blocked all UV (creating the “visible light” treatment). In the UV-B + photorepair treatment, a plastic filter (printed acetate, transmitting 91% photosynthetically active radiation (PAR) 400–700 nm, 87% UV-A 320–399 nm, and 70% UV-B 295–319 nm, in air) allowed full-spectrum exposure while keeping the starting intensity between UV-B + photorepair and visible light treatments similar. In the visible light treatment, Courtgard (CP Films, Inc., Martinsville, VA, USA) transmitted visible light, but blocked most UV (in air, transmits 87% of 400–700 nm PAR, 0% UV-B 295–319 nm and 6% of UV-A 320– 399 nm). Similar to experiment 1, mesh screens acting as neutral density filters were again used to create nine intensity levels (0%, 2.6%, 5%, 9.2%, 13%, 23%, 31%, 57%, 100%).
When D. dentifera were exposed to spores, all dishes were placed in an incubator at 24°C on a 16:8 light (cool white bulbs): dark cycle. Individual D. dentifera were fed A. falcatus (1 x 103 cells mL-1) daily for three days. After three days, individuals were moved to clean, spore-free water. Individuals were then fed daily, and the water was changed every three days.
ACKNOWLEDGMENTS
We thank Rebecca Healy for preliminary Pasteuria work.
Footnotes
Email: overhoep{at}miamioh.edu, duffymeg{at}umich.edu, meeksmp{at}miamioh.edu, leachth{at}miamioh.edu, and craig.williamson{at}miamioh.edu





{kind=link}
{kind=link}