

Abstract
Mammalian Transcription Factor 4 (TCF4) has been linked to schizophrenia and intellectual disabilities like Pitt-Hopkins syndrome (PTHS). Here we show that similarly to mammalian TCF4, fruit fly orthologue Daughterless (Da) is expressed in the Drosophila brain structures associated with learning and memory, the mushroom bodies. Furthermore, silencing of da in mushroom body neurons impairs appetitive associative learning of the larvae and leads to decreased levels of the synaptic proteins Synapsin (Syn) and discs large 1 (dlg1) suggesting the involvement of Da in memory formation. Here we demonstrate that Syn and dlg1 are direct target genes of Da in adult Drosophila heads, since Da binds to the regulatory regions of these genes and the modulation of Da levels alter the levels of Syn and dlg1 mRNA. Silencing of da also affects negative geotaxis of the adult flies suggesting the impairment of locomotor function. Overall, our findings suggest that Da regulates Drosophila larval memory and adult negative geotaxis possibly via its synaptic target genes Syn and dlg1. These behavioural phenotypes can be further used as a PTHS model to screen for therapeutics.
Summary statement Human TCF4, a bHLH transcription factor, is associated with intellectual disability and schizophrenia. Here we propose a Drosophila model for human disease studies using TCF4 orthologue in fruit fly, Daughterless.
Introduction
Transcription Factor 4 (TCF4, also known as ITF2, E2-2, SEF2 etc.) belongs to the family of class I basic helix-loop-helix (bHLH) transcription factors, also called E-proteins (Murre et al., 1994). The E-proteins bind to the DNA Ephrussi box (E-box) sequence CANNTG as homodimers or heterodimers with class II bHLH transcription factors (Cabrera and Alonso, 1991). TCF4, Transcription factor 4, should be distinguished from T cell factor 4, also called TCF4 with official name TCF7L2, interacting with β-catenin and participating in WNT signalling pathway. TCF4 is essential for a range of neurodevelopmental processes including early spontaneous neuronal activity, cell survival, cell cycle regulation, neuronal migration and differentiation, synaptic plasticity, and memory formation (Chen et al., 2016; Crux et al., 2018; Forrest et al., 2013; Hill et al., 2017; Jung et al., 2018; Kennedy et al., 2016; Kepa et al., 2017; Li et al., 2019; Page et al., 2018; Thaxton et al., 2018). Genes involved in pathways including nervous system development, synaptic function and axon development are TCF4 targets (Forrest et al., 2018; Xia et al., 2018). Furthermore, TCF4 regulates the expression of ion channels NaV1.8 and KV7.1 (Ekins et al., 2019; Rannals et al., 2016). Recent insights into the mechanisms of activation of TCF4 show that TCF4-dependent transcription in primary neurons is induced by neuronal activity via soluble adenylyl cyclase and protein kinase A (PKA) signalling (Sepp et al., 2017). In addition to nervous system, TCF4 has been shown to function in immune system in plasmacytoid dendritic cells development (Cisse et al., 2008; Grajkowska et al., 2017).
Deficits in TCF4 function are associated with several human diseases. TCF4 haploinsufficiency causes Pitt-Hopkins syndrome (PTHS; OMIM #610954) (Amiel et al., 2007; Brockschmidt et al., 2007; Zweier et al., 2007). Patients with PTHS have severe intellectual disability, developmental delay, intermittent hyperventilation periods followed by apnea, and display distinct craniofacial features, reviewed in international consensus statement (Zollino et al., 2019). Currently there is no treatment for PTHS, but dissecting the functional consequences triggered by mutated TCF4 alleles could reveal attractive avenues for curative therapies for this disorder (reviewed in Rannals and Maher, 2017). Large scale genome wide association studies revealed SNPs in TCF4 among the top risk loci for schizophrenia (SCZ) (Talkowski et al., 2012). Consistently, TCF4 is involved in SCZ endophenotypes like neurocognition and sensorimotor gating (Lennertz et al., 2011a; Lennertz et al., 2011b; Quednow et al., 2011). Furthermore, many genes that are mutated in SCZ, autism spectrum disorder and intellectual disability patients are TCF4 target genes (Forrest et al., 2018). How deficits in TCF4 function translate into neurodevelopmental impairments and whether TCF4 plays an essential role in the mature nervous system is poorly understood.
We have previously demonstrated that TCF4 function can be modelled in Drosophila melanogaster using its orthologue and the sole E-protein in the fruit fly – Daughterless (Da). PTHS-associated mutations introduced to Da lead to similar consequences in the fruit fly as do the same mutations in TCF4 in vitro (Sepp et al., 2012; Tamberg et al., 2015). Furthermore, human TCF4 is capable of rescuing the lack of Da in the development of Drosophila embryonic nervous system (Tamberg et al., 2015). Da is involved in various developmental processes including sex determination, neurogenesis, myogenesis, oogenesis, intestinal stem cell maintenance, and development of the eye, trachea and salivary gland (Bardin et al., 2010; Bhattacharya and Baker, 2011; Brown et al., 1996; Castanon et al., 2001; Caudy et al., 1988; Cline, 1978; Cummings and Cronmiller, 1994; King-Jones et al., 1999; Massari and Murre, 2000; Smith et al., 2002; Wong et al., 2008). In the developing nervous system the role of Da is well established during neuronal specification as an obligatory heterodimerization partner for proneural class II bHLH transcription factors (Cabrera and Alonso, 1991; Powell et al., 2008). However, the functional role of Da following neurogenesis and nervous system maturation remains unknown.
Here we set out to characterize the expression of Da in the nervous system. To this end we created Drosophila lines where Da protein was endogenously tagged with either 3xFLAG or sfGFP epitope tags. We show that Da is broadly expressed in the larval CNS including mushroom body, the memory and learning centre of insects. To test whether Da is involved in learning and memory formation in the fruit fly we used appetitive associative learning paradigm in larvae (Michels et al., 2017). In this assay, reduced levels of Da in the mushroom body resulted in impaired learning and memory formation. Furthermore, silencing of da also resulted in decreased level of synaptic proteins Synapsin (Syn) and discs large 1 (dlg1). Therefore, we suggest that the knockdown of da in mushroom body neurons combined with appetitive associative learning paradigm is further applicable to screen for potential therapeutics for the treatment of PTHS as well putative genetic interactors of Da and by proxy, TCF4. We also demonstrate that Da binds to several areas in the dlg1 gene and to Syn promoter region in adult Drosophila heads and that overexpression of da increases Syn and dlg1 mRNA levels in the adult heads. Therefore, we have shown for the first time that Da is required to sustain elements of the synaptic proteome in a mature nervous system positing a post-developmental function for Da and possibly TCF4.
Results
Da is expressed in all developmental stages of the fruit fly
While the expression of Da protein has been studied in fruit fly embryos, ovaries, larval optic lobes and imaginal discs using various anti-Da antibodies (Andrade-Zapata and Baonza, 2014; Bhattacharya and Baker, 2011; Bhattacharya and Baker, 2012; Brown et al., 1996; Cronmiller and Cummings, 1993; Li and Baker, 2018; Tanaka-Matakatsu et al., 2014; Yasugi et al., 2014), its expression during adulthood remains largely uncharacterized. Therefore, we first aimed to study Da expression throughout the development of the fruit fly using immunoblot analysis. Since there are no commercial antibodies available that recognize Da we used the CRISPR/Cas9 system to create transgenic flies where Da is N-terminally tagged with 3xFLAG epitope. The resulting 3xFLAG-da line is maintained as homozygotes indicating that the tagged Da protein is functional as both da null mutations and da ubiquitous overexpression lead to embryonic lethality (Caudy et al., 1988; Giebel et al., 1997). We then characterized Da expression throughout development and in adult Drosophila heads of the 3xFLAG-da line by performing immunoblot analysis with anti-FLAG antibodies. During development, we compared Da expression from embryonic to late pupal stages (Figure 1A and 1C). In adults, we analysed the Da levels from the heads of one, four and seven day old males and females (Figure 1B and 1D). We found Da to be expressed throughout development, with the highest levels detected in the 3rd instar larvae and early pupae while the lowest levels were observed in the 2nd instar larvae and late pupae (Figure 1C). During adulthood, Da expression was highest in the heads of one day old females and decreased thereafter in both males and females (Figure 1D). Also, one day old females had significantly higher Da expression in the head than one day old males (Figure 1D).

A and B - 3xFLAG-Da fusion protein is expressed throughout the fruit fly development. Western blot analysis was carried out using anti-FLAG antibody, w1118 wild type - wt serves as negative control, numbers on the right side indicate molecular weights of proteins in kDa. C and D - results of densitometric analysis of Western blot, 3xFLAG-Da signals were normalized using β-tubulin signals. The mean results from three independent Western blots are shown. Error bars show standard errors. Statistical significance is shown with asterisks between the groups connected with lines. *P<0.05, **P<0.01, ***P<0.001, Student t-test.
3xFLAG-Da retains transactivational capability of Da in HEK293 cells
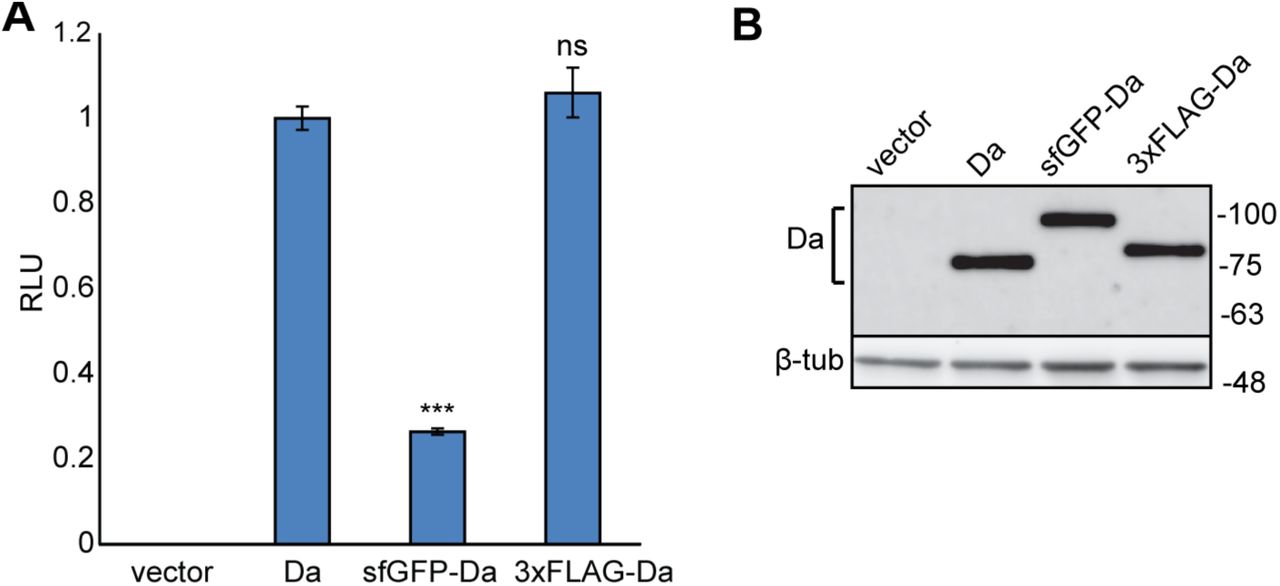
In addition to 3xFLAG-da we also created sfGFP-da flies, where da is tagged with superfolder green fluorescent protein (sfGFP) in the same N-terminal position. To determine whether N-terminal tagging of Da proteins influences their transactivation capability we used the luciferase reporter system where the expression of the luciferase gene is controlled by E-boxes with a minimal promoter. For this, we cloned the 3xFLAG-tagged or sfGFP-tagged da from the genomes of the tagged lines into mammalian expression vector pcDNA3.1 and overexpressed these constructs in HEK293 cells. Luciferase reporter assay showed that compared to wild type (wt) Da the transactivational capability of 3xFLAG-Da was unchanged (Figure 2A). In contrast, the transactivational capability of sfGFP-Da was significantly reduced (Figure 2A). Both 3xFLAG-da and sfGFP-da constructs were expressed at equal levels as revealed by Western blot analysis (Figure 2B). This suggests that the 3xFLAG tag does not interfere with the transcriptional activity of Da.

A - HEK293 cells were co-transfected with constructs encoding wild type da, tagged da, or empty vector, firefly luciferase construct carrying 12 µE5 boxes with a minimal promoter, and Renilla luciferase construct without E-boxes for normalization. Luciferase activities were measured and data are presented as fold induced levels compared to the signals obtained from cells transfected with wild type da encoding construct. The mean results from six independent transfection experiments performed in duplicates are shown. Error bars show standard errors. Statistical significance is shown with asterisks relative to wt Da expressing cells ***P<0.001, ns - not significant, Student t-test; RLU - relative luciferase unit. B - Western blot from transfected HEK293 cells using anti-Da antibody dam109-10. Wild type Da, sfGFP- and 3xFLAG-tagged Da are all expressed at equal levels. Numbers on the right side indicate molecular weight of proteins in kDa.
Da is expressed in the brain structures associated with learning and memory
Next we used the 3xFLAG-da line to characterize expression of Da in the third instar larval brain. Da was expressed throughout the larval central nervous system (CNS), with stronger expression detected in the optic lobes (Figure 3A’’ - 3C’’). Since mutations in or deletion of one of the TCF4 alleles lead to PTHS in humans and one of the hallmarks of PTHS is severe learning disability, and since TCF4 is expressed in the adult mammalian hippocampus (Jung et al., 2018), we aimed to determine whether TCF4 homolog Da is expressed in the mushroom body, the brain structure of insects responsible for learning and memory. For this, we deployed the UAS-Gal4 binary expression system (Brand and Perrimon, 1993) by combining the 3xFLAG-da line with different driver lines with expression in the mushroom body. Resulting lines with 3xFLAG-da and Gal4 were then combined with nuclear targeted UAS-nls-GFP. The GMR12B08-Gal4 line directed expression of Gal4 under the control of the single intron of da in most regions of the brain, including the mushroom body (Figure 3A). The two other drivers, 30Y-Gal4 and 201Y-Gal4, were both mushroom body-specific (Figure 3B and 3C). We observed that the expression of 3xFLAG-Da and GMR12B08>nlsGFP overlapped in many areas of the third instar larval brain (Figure 3A’), including the mushroom body (Figure 3B’ and 3C’). In contrast, with 30Y-Gal4 and 201Y-Gal4, the mushroom body-specific driver lines, 3xFLAG-Da showed partial co-expression in cells contributing to the third instar mushroom body. Thus, Da is expressed broadly in the CNS of 3rd instar larvae including mushroom body.
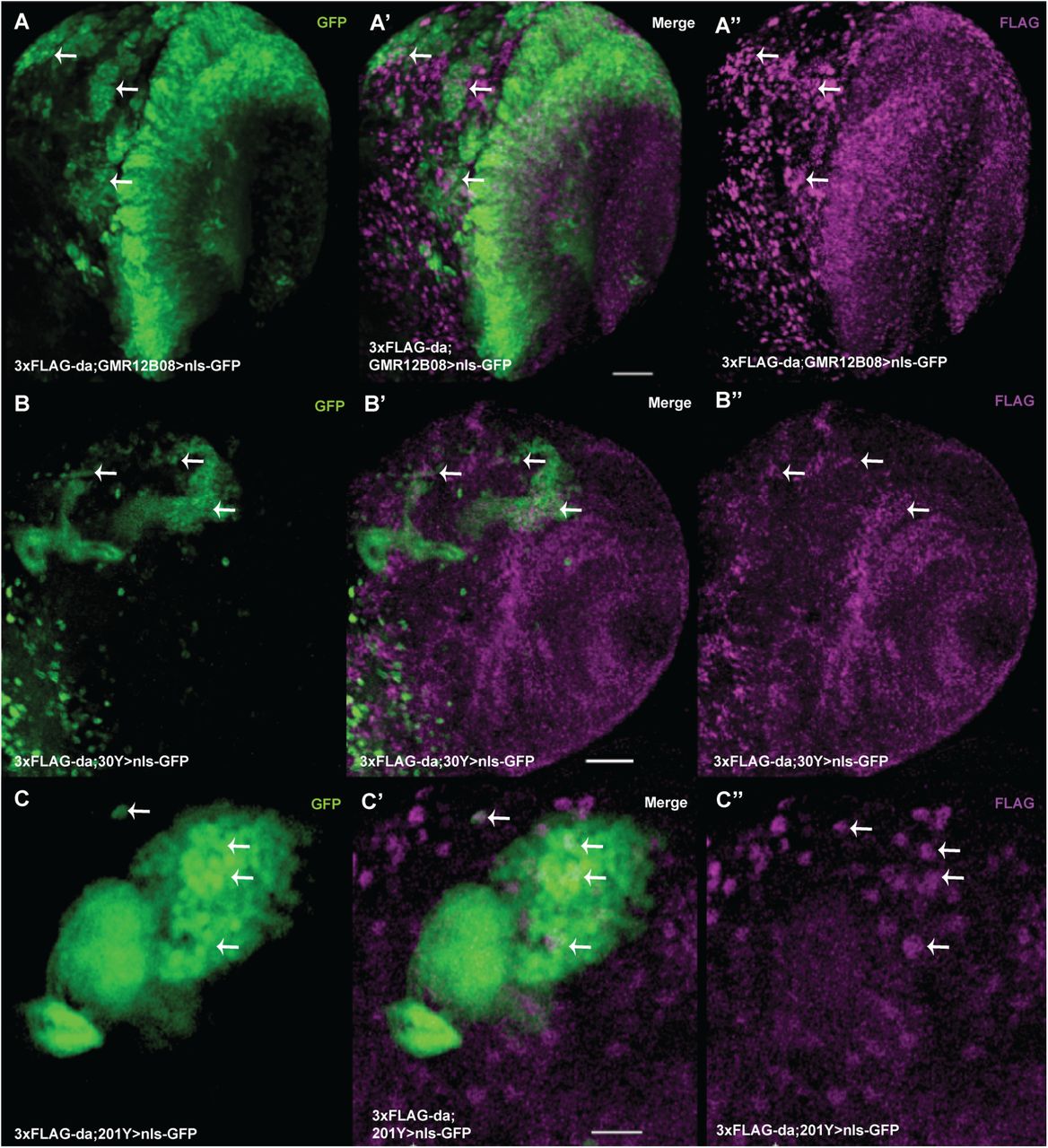
Single slices of laser confocal microscopy presented. Da is coexpressed with GMR12B08-Gal4 in many areas of the larval brain lobe (A-A’’). Da is expressed in the mushroom body, which is marked by nlsGFP driven by 30Y-Gal4 (B-B’’) and by 201Y-Gal4 (C-C’’). A, B, and C - nlsGFP expression shows driver expression pattern. A’’, B’’ and C’’ - expression of 3xFLAG-Da. Some sites of coexpression of nlsGFP and 3xFLAG-Da are shown by arrows. Scale bars on A’ and B’ represent 30 µm and on C’ 20 µm.
Silencing of da in the CNS leads to impaired memory of the larvae
Heterozygous mutations in TCF4, the orthologue of da, lead to the PTHS syndrome, characterized by intellectual disability. This fact, and the observation that Da is expressed in the mushroom body implies that it might be involved in learning and memory in flies. To test this, we decided to take advantage of the ease of assaying appetitive associative learning and memory in the Drosophila larvae (Michels et al., 2017). However, this assay showed that learning ability is not impaired in da heterozygous mutants (Figure 4A), which could be due to da upregulation by autoregulation (Smith and Cronmiller, 2001). Since sfGFP-Da showed diminished transactivation capability in vitro (see above), we also tested homozygous sfGFP-da larvae, and found no impairment of learning ability (Figure 4A). Thus, we next investigated whether knockdown of da with concurrent enhancement by Dicer-2 (Dcr2) expression (Dietzl et al., 2007) in the Drosophila CNS could impact memory and learning ability. To silence da in the CNS, we used several CNS-specific Gal4 lines. We found that appetitive associative learning was impaired in flies when using three drivers - GMR12B08-Gal4 (Figure 4B) and mushroom body specific lines 30Y-Gal4 (Figure 4C) and 201Y-Gal4 (Figure 4D). For control experiments, we used both the UAS-daRNAi line and the UAS-Dcr2 driven by the CNS-specific Gal4 line. All of the transgenes (UAS-Dcr2, UAS-daRNAi and the driver Gal4) were homozygous for lowered learning ability in the case of GMR12B08-Gal4 and 201Y-Gal4 (Figure 4B and 4D). In the case of UAS-Dcr2;3xFLAG-da,UAS-daRNAi;GMR12B08-Gal4 line (where 3xFLAG-da, UAS-daRNAi and GMR12B08-Gal4 were all in a homozygous state), Da levels in the larval brains were reduced by approximately 25% and 35% when compared to UAS-Dcr2;3xFLAG-da;GMR12B08-Gal4 and 3xFLAG-da larval brains, respectively (Suppl. figure 1A and 1B). We were unable to get homozygotes using mushroom body specific driver 30Y-Gal4, possibly because knockdown of da with this driver in homozygous state is too strong and causes lethality. The learning ability of heterozygous larvae where da was silenced with the 30Y-Gal4 driver was notably but non-significantly impaired (Figure 4C). Larvae with impaired learning were also tested for their ability to taste and smell. All three lines showed preference towards fructose (Suppl. fig 2A) and amyl-acetate (Suppl. fig 2B) which indicates that the larvae can sense taste and smell and that their locomotor activity is not affected. Expressing only Dcr2 under neuron-specific elav-Gal4, as a control for da silencing using the same driver, as such already caused learning impairment (data not shown), so learning ability of elav-Gal4>Dcr2;daRNAi larvae was nonsignificantly different from the control larvae elav-Gal4>Dcr2. Knockdown of da with glia-specific repo-Gal4 did not alter learning and memory (data not shown). This suggests that for normal larval appetitive associative memory appropriate Da levels are needed in the brain structures specified by GMR12B08-Gal4, 30Y-Gal4 and 201Y-Gal4.

A - Heterozygous da mutation does not cause reduction of appetitive associative learning, and both FLAG-da and GFP-da larvae show olfactory learning. B - Appetitive associative learning was impaired when da was silenced using GMR12B08-Gal4, C - 30Y-Gal4 and D - 201Y-Gal4. UAS-Dcr2, UAS-daRNAi and Gal4 - all were homozygous when driven by GMR12B08-Gal4 (B) or 201Y-Gal4 (D). In the case of 30Y-Gal4 homozygotes never emerged and the effect on learning was smaller (C). PI-s (performance index) are visualized using box-whisker plots, which show the median, the 10% - 90% quantiles, and the 25% - 75% quantiles. For statistical analysis one-sample sign test and Mann-Whitney U test with Bonferroni correction were used. * p<0.025, ** p<0.005.
Reduced level of Da in the larval CNS leads to decreased expression of synaptic proteins Synapsin and discs large 1
To investigate the putative mechanisms underlying learning and memory deficits in larvae with lowered levels of Da in the nervous system we used the driver line GMR12B08-Gal4 for silencing da since it had the broadest expression. For this we compared the expression levels of several known synaptic proteins in the 3rd instar larval brains under both da knockdown and overexpression conditions using the GMR12B08-Gal4 line (Figure 5). We quantified the expression levels of presynaptic protein bruchpilot (brp) (Figure 5A), postsynaptic protein discs large 1 (dlg1) (Figure 5B), presynaptic Synapsin (Syn) (Figure 5C) which is important for learning and memory (Michels et al., 2005), and pan-neuronally expressed neuronal specific splicing factor embryonic lethal abnormal vision (elav) (Figure 5D). We found that the levels of both dlg1 and Syn were reduced in 3rd instar larval brains with lower levels of Da (Figure 5B and 5C respectively). On the other hand, Da overexpression did not result in increased levels of these proteins. The levels of elav and brp were not significantly changed by knockdown or overexpression of da (Figure 5A and 5D, respectively). The finding that elav levels were not affected by Da suggests that reducing Da levels does not affect the number of neurons and the observed learning impairment might rather stem from lowered expression levels of synaptic proteins or, alternatively, from reduced number of synapses.

Western blot was carried out using larval brains where da was silenced with GMR12B08-Gal4. A-D - results of densitometric analysis of Western blot, protein signals were normalized using β-tubulin signals. The mean results from four independent Western blots are shown. Error bars show standard errors. Statistical significance is shown with asterisks between the groups connected with lines. *P<0.05, **P<0.01, ***P<0.001, Sudent t-test. Overexpression of Da does not alter bruchpilot, discs large 1, Synapsin or embryonic lethal abnormal vision levels (A-D). discs large 1 and Synapsin expression levels are lower when Da is silenced (B and C respectively). Silencing of Da does not change bruchpilot and embryonic lethal abnormal vision expression levels (A and D respectively). E – Representative Western blot using 3rd instar larval brains. Numbers indicate molecular weight of proteins in kDa. Control - GMR12B08>Dcr2 larval brains, Da overexpression - GMR12B08>Da larval brains, daRNAi - GMR12B08>Dcr2,daRNAi larval brains.
Larval appetitive associative learning and memory test can be used for screening drugs for PTHS treatment
Our finding showing that larval appetitive associative learning and memory becomes impaired upon da silencing indicates that these fly lines could be used for modelling certain aspects of PTHS in Drosophila and testing potential drug candidates. For instance, various drugs or drug candidates could be tested for their capacity to rescue this behavioural impairment. Previously, it has been shown in our group that resveratrol improves transactivational capability of TCF4 in primary neuronal cultures (unpublished data). Also histone deacetylase inhibitor suberoylanilide hydroxamic acid (SAHA) has been shown to rescue memory impairment in the mouse model of PTHS (Kennedy et al., 2016). We therefore decided to test these two substances in the appetitive associative learning experiments. We observed that while the da knockdown larvae fed with 400 µM resveratrol or 2 µM SAHA showed increased associative memory, the rescue of the learning deficit was nonsignificant when compared to the controls (Figure 6A and 6B). Nevertheless, the impaired memory and learning of Drosophila larvae with specific da knockdown can be further used for screening pharmaceuticals for potential treatment of PTHS.

A and B - Adding 400 µM resveratrol or 2 µM SAHA to the larval growth media improves appetitive associative memory. PI-s are visualized using box-whisker plots, which show the median, the 10% - 90% quantiles, and the 25% - 75% quantiles. For statistical analysis one-sample sign test and Mann-Whitney U test with Bonferroni correction were used. Median PI (performance index) of these larvae are significantly different from zero so these larvae show associative memory although the PI-s are not significantly different compared to the controls.
Suppressing Da using 30Y-Gal4 leads to impaired negative geotaxis of adult flies
Negative geotaxis has been successfully used to evaluate climbing ability indicative of motor dysfunction in the Drosophila model for Angelman syndrome that has similar symptoms to PTHS (Wu et al., 2008). Thus, we next used this assay to evaluate locomotion in adult flies where da knockdown had been achieved by the same drivers as used for the larval learning test. We found that negative geotaxis was unchanged in homozygotes where da knockdown had been achieved by the broad neuronal driver GMR12B08-Gal4 or mushroom body specific driver 201Y-Gal4 (Figure 7A or 7C respectively). Interestingly, both female and male heterozygotes whose da was silenced by the 30Y-Gal4 driver had severely impaired negative geotaxis (Figure 7B). Next we visualized Da expression in the adult heads using 3xFLAG-da line and confirmed its coexpression with 30Y-Gal4. Da was expressed widely in the adult Drosophila brain including the central brain and optic lobes (Figure 7D-D’’), and coexpressed with 30Y-Gal4 in many cells in the mushroom body (Figure 7E-E’’). 30Y-Gal4 is expressed in all mushroom body αβ, α’β’ and γ lobes but 201Y-Gal4 is not expressed in α’β’ Kenyon cells (Aso et al., 2009). It has previously been shown that when mushroom body α’β’ Kenyon cells are activated then negative geotaxis is decreased (Sun et al., 2018). Silencing da by 201Y-Gal4 or GMR12B08-Gal4 did not cause decreased negative geotaxis probably because these drivers are not expressed in brain areas responsible for this behaviour, for example mushroom body α’β’ lobes. We also made an attempt to rescue this phenotype by administrating resveratrol or SAHA either to larvae during development or to adult flies in the food substrate, but we could not get an improvement (data not shown).

Negative geotaxis was not affected when Da was suppressed using GMR12B08-Gal4 (A) or 201Y-Gal4 (C). The climbing height of the flies was significantly lower when Da was silenced using 30Y-Gal4 (B). Average climbing heights are visualized using box-whisker plots, which show the median, the 10% - 90% quantiles, and the 25% - 75% quantiles. For statistical significance pairwise U-tests were used. * p<0.05, ** p<0.01. D-D’’ – Da is expressed widely in the adult Drosophila brain including the central brain and optic lobes. 3xFLAG-Da is green on D and D’ and nuclei are visualized using DAPI on D’ and D’’. Scale bar represents 50 µm (D’). E-E’’ – Da coexpresses with 30Y-Gal4 in adult Drosophila mushroom body. 30Y-Gal4 expression is visualized with nlsGFP (E and E’) and 3xFLAG-Da is magenta (E’ and E’’). Some parts of coexpression is shown by arrows. Scale bar represents 15 µm (E’).
Synapsin and discs large 1 are Da target genes
Since Syn and dlg1 levels were reduced in third instar larval brains when da was silenced and it has been shown that Da binds to both Syn and dlg1 gene locus in embryonic stage 4-5 (MacArthur et al., 2009) we sought to investigate whether Da binds to these areas in adult heads too. For this we used chromatin immunoprecipitation assay (ChIP) in 3xFLAG-da adult heads using anti-FLAG antibodies. As a control we used white1118 fly line with no FLAG tag. For quantitative PCR (qPCR) from immunoprecipitated chromatin we designed primers to amplify Syn and dlg1 gene areas containing E-boxes where Da binds in early embryos (MacArthur et al., 2009). In addition to previously shown Da binding site in Syn gene we also tested Da binding to Syn promoter region (Figure 8A). For dlg1 we designed four primer pairs, since Da has been shown to bind four areas in that gene (Figure 8B) (MacArthur et al., 2009). As a negative control we used primers for achaete (Andrade-Zapata and Baonza, 2014) since achaete is a proneural protein essential for neuronal development and should not be expressed in adult heads. As a positive control we used peptidylglycine alpha-hydroxylating monooxygenase (PHM) gene first intron where Da binds as a heterodimer with dimmed to activate transcription (Park et al., 2008). qPCR from immunoprecipitated chromatin using Syn primers resulted in enrichment of Syn promoter area (primer pair SynI) while previously reported Da binding site was not enriched in adult heads (primer pair SynII) (Figure 8C). All dlg1 primers resulted in enrichment of previously reported Da binding areas (Figure 8C). This means that Da does not bind to the locus at the 3’ end of Syn gene but binds to Syn promoter and all four dlg1 gene areas that we selected in the adult heads. To validate Syn and dlg1 as Da target genes we carried out RT-qPCR analysis in adult Drosophila heads under da silencing and overexpression conditions. Although upon da silencing using elav-Gal4 only Syn mRNA levels were decreased (Figure 8D), da overexpression using elav-Gal4 increased mRNA levels of both Syn and dlg1 (Figure 8E). This indicates that both Syn and dlg1 are direct targets of Da in the Drosophila nervous system.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A – JBrowse view of Syn gene, four annotated transcripts are shown. B – JBrowse view of dlg1 gene, 21 annotated transcripts are shown. Red boxes indicate areas, where primer pairs Syn1, SynII, Dlg1I, Dlg1II, Dlg1III and Dlg1IV amplify DNA. C - qPCR results from chromatin immunoprecipitation experiment from 3xFLAG-da and white1118 wild type adult heads using anti-FLAG antibody. ac – achaete gene locus as a negative control; PHM - peptidylglycine alpha-hydroxylating monooxygenase gene first intron as a positive control; SynI – promoter region of Synapsin, SynII – 3’ end of Synapsin gene locus; dlg1I, dlg1II, dlg1III, dlg1IV – discs large 1 gene locus. D, E – RT-qPCR results showing the effects of da silencing (D) or overexpression (E) using elav-Gal4 on da, ac, PHM, Syn and dlg1 mRNA levels. da silencing reduces da and Syn, and increases ac mRNA levels (D). da overexpression increases da, Syn and dlg1 mRNA levels (E). C, D, E - Results from three biological replicates are shown, error bars indicate standard errors. *P<0.05, **P<0.01, ***P<0.001, Student t-test.
Discussion
Here we characterized the expression of Da in the Drosophila melanogaster larval and adult brain. Da was expressed in many areas of the brain including the mushroom body, which is the centre for learning and memory in the fruit fly and carries out a role that is comparable to the mammalian hippocampus. Orthologue of da, TCF4 is expressed not only in the adult mammalian hippocampus but also in cortical and subcortical structures (Jung et al., 2018).
We created N-terminally tagged 3xFLAG-da and sfGFP-da fly strains. Both strains are homozygous viable and fertile, indicating that the overall functionality of Da in vivo is not altered by the molecular tag. However, in luciferase reporter assay in mammalian HEK293 cells the sfGFP tag reduced transcription activation capability of Da. E-proteins activate transcription preferably as heterodimers with class II bHLH proteins, but can also act as homodimers (Cabrera and Alonso, 1991). In mammalian HEK293 cells Da has to activate transcription as a homodimer, since there are no heterodimerization partners like class II bHLH proteins expressed (Sepp et al., 2012). This suggests that sfGFP tag could interfere with Da function as a homodimer in the luciferase assay but not as a heterodimer in vivo. We also compared appetitive associative learning ability of 3xFLAG-da and sfGFP-da larvae and both of the lines had no learning impairment in this assay. This provides additional evidence that the 3xFLAG tag does not affect Da function and sfGFP tag reduces its transactivational capability probably by interfering with Da homodimer function.
Since Da is expressed in larval brain structures associated with learning and memory, and PTHS is caused by heterozygous mutations in TCF4 we tested learning ability of da heterozygous mutant larvae. These larvae had no memory impairment, which could be due to da upregulation by autoregulation (Smith and Cronmiller, 2001). Learning and memory of larvae was impaired when da was knocked down in the mushroom body. Da mammalian orthologue TCF4 is also associated with learning and memory, since when TCF4 is downregulated in mouse hippocampus, pathways associated with neuronal plasticity are dysregulated (Kennedy et al., 2016) and silencing of TCF4 in human pluripotent stem cell-derived neurons results in downregulated signalling pathways important for learning and memory (Hennig et al., 2017). In TCF4 conditional knock-out mice the neurons in the cortex and hippocampus have reduced numbers of dendritic spines, which also suggests that synaptic plasticity is altered (Crux et al., 2018). In multiple PTHS mouse models spatial learning is defective probably through hippocampal NMDA receptor hyperfunction (Thaxton et al., 2018). Furthermore, many genes that code for synaptic proteins and have been linked to autism, intellectual disability, or psychiatric diseases are direct targets of TCF4 (Forrest et al., 2018; Hennig et al., 2017).
Here we show that when da is silenced using driver with broad expression in the Drosophila larval brain, expression levels of synaptic proteins disc large 1 (dlg1) and Synapsin (Syn) are downregulated. dlg1 is a member of the membrane-associated guanylate kinase (MAGUK) protein family. Several vertebrate homologs of dlg1 have been shown to be important for learning and memory. Discs Large MAGUK Scaffold Protein 3 (DLG3), also called Synapse-Associated Protein 102 (SAP-102) knock-out mice have spatial learning deficit (Cuthbert et al., 2007) and in human DLG3 mutations which cause dysfunctional NMDA receptor signalling have been associated with X-linked mental retardation (Tarpey et al., 2004; Zanni et al., 2010). We also found that Da is a direct regulator of dlg1, since in the adult Drosophila heads Da binds to multiple areas in dlg1 gene and dlg1 expression is upregulated when da is overexpressed. Gene coding for Discs Large MAGUK Scaffold Protein 2 (DLG2) also called Postsynaptic Density Protein 93 (PSD-93) which is a homolog of Drosophila dlg1, is a direct target of TCF4 (Hennig et al., 2017), which indicates that Da and TCF4 share at least some common mechanisms in regulating learning and memory.
Synapsins are presynaptic phosphoproteins that regulate synaptic output (reviewed in Diegelmann et al., 2013). There are three genes coding for vertebrate Synapsins but only one Syn gene in Drosophila (Klagges et al., 1996). Using knockout experiments in mice it has been shown that Synapsins are involved in learning and memory (Gitler et al., 2004; Silva et al., 1996) and SYN1 has been implicated in human neurological diseases such as learning difficulties and epilepsy (Garcia et al., 2004). Likewise, Drosophila syn97 mutant larvae have impaired appetitive associative learning (Michels et al., 2005). The fact that memory of syn97 larvae can be rescued by expressing Syn in the mushroom bodies (Michels et al., 2011) is consistent with our findings that lower Da levels affect Syn expression levels and that appropriate Da levels in mushroom body are required for proper memory formation. This means that Da and Syn-dependent memory trace is formed in the mushroom bodies. Syn-dependent memory is likely formed by its phosphorylation by Protein kinase A (PKA) (Michels et al., 2011). When Syn is phosphorylated at its PKA/CamK I/IV (Protein kinase A/ Ca2+/calmodulin-dependent protein kinase I/IV) sites its affinity for actin is reduced and synaptic vesicles from the reserve pool can be exocytosed (reviewed in Benfenati, 2011). We found that Syn is likely a direct target of Da since Da binds to Syn promoter, and both silencing and overexpression of da changes Syn levels.
We also sought out to rescue the learning phenotype caused by da silencing. For this we fed the larvae with resveratrol or SAHA. In luciferase reporter assay in primary neuronal cultures resveratrol improves transactivational capability of TCF4 significantly (unpublished data from our lab). Resveratrol inhibits cAMP-degrading phosphodiesterases which leads to elevated cAMP levels (Park et al., 2012) and TCF4-dependent transcription upon neuronal activity is activated by cAMP-PKA pathway mediated phosphorylation of TCF4 (Sepp et al., 2017). It is plausible that Da could also be regulated by phosphorylation by PKA and therefore resveratrol could improve Da transactivational capability. Indeed, resveratrol had a moderate positive effect on learning and memory of the da knockdown larvae. Whether this effect is through cAMP-PKA pathway is yet to be verified. SAHA is a histone deacetylase inhibitor which improves learning and memory in TCF4(+/−) mice through the normalization of synaptic plasticity (Kennedy et al., 2016). Here we show that feeding SAHA to Drosophila larvae had also a moderate effect on learning.
We also made an attempt to rescue the impaired geotaxis of 30Y>Dcr2;daRNAi flies using resveratrol or SAHA, but we could not get significant improvement. We administrated the drugs in the food substrate either during development or to adult flies. Getting resveratrol or SAHA during development in the larval stage and the amount ingested by the adults is probably not enough to rescue phenotypes caused by lowered levels of Da in the adults. If administrating the drug in the food substrate during development and to adults prior to testing is insufficient, other administration methods could be used like possibly starving the adult flies to increase the amounts of drug ingested (Pandey and Nichols, 2011). In a recent study in Drosophila melanogaster where genes associated with autism spectrum disorders and intellectual disability were suppressed, the knockdown of Da resulted in impaired habituation (Fenckova et al., 2018). Rescuing this habituation phenotype could be also tested for improvement with drugs.
In conclusion, this study demonstrates that the levels of TCF4 homolog Da are important for learning and memory of Drosophila larvae and that Da directly regulates the expression of Syn and dlg1. This gives novel insights into the mechanisms of PTHS and this learning deficit model can be used for screening therapeutics for PTHS.
Materials and methods
Drosophila stocks
All Drosophila stocks and crosses were kept on malt and semolina based food with 12 h light and dark daily rhythms at 25°C with 60% humidity. Drosophila strains used in this study were CantonS (a gift from dr. Bertram Gerber), 201Y-Gal4 (Bloomington Drosophila Stock Center (BDSC #4440) and 30Y-Gal4 (BDSC #30818) (Yao Yang et al., 1995) were gifts from Mark Fortini. GMR12B08-Gal4 (BDSC #48489) (Pfeiffer et al., 2008), elav-Gal4 (BDSC #8760) (Luo et al., 1994), repo-Gal4 (BDSC #7415) (Sepp et al., 2001), UAS-Dcr2;Pin1/CyO (BDSC #24644) (Dietzl et al., 2007), UAS-nlsGFP (BDSC #4776), UAS-daG (BDSC #37291), da mutant line da10 (BDSC #5531) (Caudy et al., 1988) from Bloomington Stock Center at Indiana University, USA. The following transgenic lines were generated in this study: 3xFLAG-da2M4 and sfGFP-da4M1.
Endogeneous tagging of Da by CRISPR/Cas9
The coding sequence for 3xFLAG- or sfGFP-tag was inserted to the 5’ coding region of da gene using CRISPR-Cas9 technology. Genomic sequence around the tag was following: 5’ ATGGCGACCAGTG|ACGATGAGCC 3’ (PAM sequence shown as bold and the cut site marked with |). For the higher mutagenesis rate a specific fruit fly line for guide RNA production was created. Partially overlapping oligonucleotides 5’-CTTCGTGCATCGGCTCATCGTCAC-3’ and 5’-AAACTGGACGATGAGCCGATGCAC-3’, designed to target the N-terminus of Da protein, were cloned downstream the Polymerase III U6:2 promoter in the pCFD2-dU6:2gRNA plasmid (Addgene #49409). Transgenic flies expressing gRNA-s were created by injecting the generated plasmid into PBac{yellow+-attP-9A}VK00027 (BDSC #9744) fly strain embryos. For donor plasmid generation, pHD-3xFLAG-ScarlessDsRed (a gift from Kate O’Connor-Giles (DGRC)) or pHD-sfGFP-ScarlessDsRed (a gift from Kate O’Connor-Giles (DGRC)) and Gibson cloning was used. The following primer pairs were used for amplification of upstream and downstream homology arms: upst F5 CGGCCGCGAATTCGCCCTTGGTTGTGAATCAGGTGTAGAAACA and upst_R GCCGGAACCTCCAGATCCACCACTGGTCGCCATTTCAGCA, dwns_F TTCTGGTGGTTCAGGAGGTTACGATGAGCCGATGCACTTG and dwns_R GTTTAAACGAATTCGCCCTTAACGCCCTGGAACACCGAGG.
After verification, the obtained donor plasmids pHD-da-3xFLAG-ScarlessDsRed and pHD-da-sfGFP-ScarlessDsRed were injected into F1 embryos from a cross between da-gRNA (our gRNA expressing transgenic strain) and y1M{w+mC=nos-Cas9.P}ZH-2A w* (BDSC #54591) fly strains. All embryo injections were ordered from BestGene Inc. (USA).
dsRed cassette was removed from selected progeny by crossing to PB transposase line Herm{3xP3-ECFP,αtub-piggyBacK10}M10 (BDSC #32073) (Horn et al., 2003). The obtained 3xFLAG-da and sfGFP-da lines were verified by sequencing.
RNA isolation and cloning
RNA from 3xFLAG-da or sfGFP-da Drosophila embryos was isolated using RNeasy Mini Kit (Qiagen) according to the manufacturer’s protocol. cDNA was synthesized using 2µg of RNA. Primer sequences for cloning were 5’ ACTAGTTGAAGTCGACTGGAC 3’ and 5’ CCAGGTCCTCCAATTCCACC 3’. PCR products containing either 3xFLAG-da or sfGFP-da cDNA sequences were sequenced and cloned into pCDNA3.1 expression vector (Tamberg et al., 2015) using BcuI (SpeI, 10 U; Thermo Scientific) and BstII (Eco 91I, 10 U; Thermo Scientific) restriction enzymes. The pcDNA3.1 constructs encoding Da, and reporter vectors pGL4.29[luc2P/12µE5/Hygro], pGL4[hRlucP/PGK], pGL4.83[hRlucP/EF1α/Puro] have been described previously (Sepp et al., 2011; Sepp et al., 2012; Tamberg et al., 2015).
Cell culture and transfections and luciferase reporter assay
Human embryonic kidney cells HEK-293 obtained from ATCC (LGC Standards GmbH, Wesel, Germany) and routinely tested for contamination were grown in MEM (Capricorn Scientific) supplemented with 10% fetal bovine serum (PAA Laboratories), 100 U/ml penicillin, and 0.1 mg/ml streptomycin (Gibco). For transfection 0.375 µg of DNA and 0.75 µg of PEI (Sigma-Aldrich) were used per well of a 48-well plate or scaled up accordingly. For cotransfections, equal amounts of pGL4.29[luc2P/12µE5/Hygro], pGL4[hRlucP/min/Hygro], and effector constructs were used. Luciferase assays were performed as described previously (Sepp et al., 2011) using Passive Lysis Buffer (Promega) and the Dual-Glo Luciferase Assay (Promega). Cells were lysed at 24 h after transfection. For data analysis, background signals from untransfected cells were subtracted and firefly luciferase signals were normalised to Renilla luciferase signals. The data was then log-transformed, auto-scaled, means and standard deviations were calculated and Student t-tests were performed. The data was back-transformed for graphical representation.
Protein electrophoresis and Western blotting
For SDS-PAGE embryos, larvae, pupae, adult heads or larval brains were lysed in 2x SDS sample buffer. Equal amounts of protein were loaded to gel.
The following mouse monoclonal antibodies were obtained from the Developmental Studies Hybridoma Bank (created by the NICHD of the NIH and maintained at The University of Iowa, Department of Biology, Iowa City, IA 52242): β-tubulin E7 (developed by Klymkowsky, M., dilution 1:3000); Synapsin SYNORF1 3C11 (developed by Buchner, E., 1:1000); discs large 1 4F3 (developed by Goodman, C., 1:2000), elav 9F8A9 (developed by Rubin, Gerald M., 1:1000), Bruchpilot nc82 (developed by Buchner, E., 1:100).
Other antibodies and dilutions used: mouse anti-Da dam109-10 1:10 (a gift from C. Cronmiller), mouse anti-FLAG M2 HRP-conjugated 1:6000 (Sigma-Aldrich), goat anti-mouse IgM HRP-conjugated secondary antibody (MilliporeSigma).
Immunohistochemical staining
The anterior parts of 3rd instar larvae were dissected in PBS and fixed using 4% paraformaldehyde in PBS. Adult flies were first fixed in 4% paraformaldehyde in PBS and then dissected. Primary antibody labelling was performed over 72 hours on overhead rotator at 4°C in PBS with 0.5% TritonX-100. Used antibodies were as follows: mouse anti-FLAG M2 1:1000 (Sigma-Aldrich) and goat anti-mouse Alexa594 1:1000 (ImmunoResearch Laboratories). Secondary antibodies were preadsorbed to wt tissues before use. Incubation with secondary antibodies was performed for 3 h on overhead rotator at room temperature in PBS with 0.1% TritonX-100. The labelled larval brains were dissected and mounted in Vectashield mounting medium (Vector Laboratories). For image collection, Zeiss LSM 510 Meta confocal microscope with Pln Apo 20×/0.8 DICII objective (Carl Zeiss Microscopy) was used. Suitable layers were selected using Imaris (Bitplane Inc.) software.
Appetitive associative learning assay
Appetitive associative memory assay in the Drosophila larvae was performed as previously described (Michels et al., 2017). Shortly, the larvae were trained three times for 5 minutes on Petri dishes, where one odor - amyl acetate (AM) was presented with plain agar and the other odor – octanol (OCT) with agar containing fructose as a reward. Then the larvae were placed in the midline of a plain agar plate and given a choice between the two odors placed on separate halves of the Petri dish and after three minutes larvae were counted on each half of the Petri dish. Then reciprocal training was performed with AM and fructose and OCT with plain agar. Using data from two reciprocally trained tests the performance index (PI) was calculated PI = (PREF AMAM+/OCT - PREF AMAM/OCT+) / 2. The odors and the reward were presented in four different orders to eliminate any non-specific preferences. All together 12 training and test cycles were conducted per genotype, each time with new larvae, PI-s were calculated and used for statistical analysis. The PI-s were visualized as box-whisker plots, which show the median, the 10% - 90% quantiles, and the 25% - 75% quantiles. For statistical analysis inside one genotype a one-sample sign test was applied with an error threshold of smaller than 5%. For pairwise U-tests Bonferroni correction was used. SAHA was dissolved in dimethyl sulfoxide (DMSO) and same concentration of DMSO (0.1%) was used in the food substrate for a control. Resveratrol was dissolved in 96% ethanol and 1% ethanol in the food was used for the control.
Chromatin immunoprecipitation
Chromatin preparations were carried out as previously described (Chanas et al., 2004). Approximately 150 mg of adult heads were collected on dry ice and homogenized in buffer A1 (60 mM KCl, 15 mM NaCl, 4 mM MgCl2, 15 mM HEPES pH 7.6, 0.5% Triton X-100, 0.5 mM DTT, 10 mM sodium butyrate, 1 × EDTA-free protease inhibitor cocktail (Roche)) + 1.8% formaldehyde in room temperature using first KONTES pellet pestle followed by three strokes using Dounce homogenizer with a loose pestle. Homogenate was incubated 15 minutes and glycin was added to 225 mM followed by 5 minutes incubation. Homogenate was then centrifuged 5 minutes at 4000 g at 4°C and supernatant discarded. Pellet was washed three times with 3 ml A1 followed by a wash with 3 ml of lysis buffer (14 mM NaCl, 15 mM HEPES pH 7.6, 1 mM EDTA, 0.5 mM EGTA, 1% Trition X-100, 0.5 mM DTT, 0.1% sodium deoxycholate, 0.05% SDS, 10 mM sodium butyrate, 1 × EDTA-free protease inhibitor cocktail (Roche)). Cross-linked material was resuspended in 0.5 ml of lysis buffer + 0.1% SDS and 0.5% N-lauroylsarcosine and incubated 10 minutes at 4°C on a rotator followed by sonication using SONICS VibraCell on 70% amplitude 15 seconds intervals 30 times. Cross-linked material was then rotated 10 minutes at 4°C and centrifuged 5 minutes at room temperature at maximum speed. Supernatant was transferred to a new tube and 0.5 ml of lysis buffer was added to the pellet followed by rotation and centrifugation. Supernatants were combined and centrifuged 2×10 minutes at maximum speed. Chromatin extract was transferred to Microcon DNA Fast Flow Centrifugal Filter Units (Merck Millipore) blocked with 1mg/ml BSA in PBS and purified using lysis buffer. The volume of chromatin extract was brought to 1 ml using lysis buffer. Protein concentrations were determined using BCA assay (Pierce).
After removing equal amounts of inputs chromatin extracts were diluted 10x using dilution buffer (1% Triton X-100, 150 mM NaCl, 2 mM EDTA pH 8.0, 20 mM Tris-HCl pH 8.0, 1 × EDTA-free protease inhibitor cocktail (Roche)) and added to 50 µl of Dynabeads Protein G (Invitrogen) beads that were previously incubated with 5 µg of monoclonal anti-FLAG M2 antibody (Sigma-Aldrich) in 0.05% PBS Tween20 overnight. Chromatin immunoprecipitation (ChIP) was carried out overnight at 4°C. Beads with chromatin were then washed in wash buffer (1% Triton X-100, 0.1% SDS, 150 mM NaCl, 2 mM EDTA pH 8.0, 20 mM Tris-HCl pH 8.0, 1 × EDTA-free protease inhibitor cocktail (Roche)) using a magnetic rack for 10 minutes three times at 4°C on a rotator followed by final wash buffer (1% Triton X-100, 0.1% SDS, 500 mM NaCl, 2 mM EDTA pH 8.0, 20 mM Tris-HCl pH 8.0, 1 × EDTA-free protease inhibitor cocktail (Roche)). Chromatin was eluted using 3×50 µl of elution buffer (1% SDS, 100 mM NaHCO3, 1 mM EDTA) 3×10 minutes at 37°C. The volume of inputs was brought to 150 µl with elution buffer. For decrosslinking 8 µl of 5 M NaCl was added and the samples were incubated at 65°C overnight. Then 2 µl of RNase A (10 mg/ml) was added and incubated at 37°C for 30 minutes followed by incubation with 2 µl of EDTA (0.5M) and 4 µl Proteinase K (10 mg/ml) at 45°C for 30 minutes. DNA was extracted using QIAquick PCR Purification Kit (Qiagen).
Quantitative PCR
For RT-qPCR 15 heads were collected from 2-3 days old adult flies on dry ice. RNA was extracted using RNeasy Mini Kit (Qiagen). cDNA was synthesized with Superscript IV reverse transcriptase (Invitrogen) and oligo(dT)20 primers. Quantitative PCR was performed using LightCycler 480 II (Roche) with Hot FIREPol EvaGreen qPCR Mix Plus (Solis Biodyne). Primer sequences are shown in Supplementary table 1.
Negative geotaxis assay
10 females and males were separated to fresh vials 48 hours before the assay to allow recovering from anesthesia. Prior to the test males and females from control and da silencing group were transferred to empty vials without anesthesia and closed with another upside down vial using sticky tape. The flies were knocked down three times on the table and a photo was taken after 10 seconds. The height of the vial was divided into 10 equal parts and the number of flies in each compartment was counted and average height was calculated. The experiment was repeated five times, each time with new flies. Average climbing heights were visualized using box-whisker plots, which show the median, the 10% - 90% quantiles, and the 25% - 75% quantiles. For statistical significance pairwise U-tests were used.
Competing interests
No competing interests declared.
Funding
This study was supported by Estonian Research Council (institutional research funding IUT19-18), European Union through the European Regional Development Fund (Project No. 2014-2020.4.01.15-0012) and H2020-MSCA-RISE-2016 (EU734791), Pitt Hopkins Research Foundation and Million Dollar Bike Ride Pilot Grant Program for Rare Disease Research at UPenn Orphan Disease Center (grants MDBR-16-122-PHP and MDBR-17-127-Pitt Hopkins).
Author contributions statement
Conceptualization: L.T., M.P.; Methodology: L.T., M.J., M.P.; Formal analysis: L.T., A.Sh., C.S.K.; Investigation: L.T., M.J., K.S., A.Sh., C.S.K., M.P.; Resources: T.T.; Writing – original draft preparation: L.T.; Writing – review and editing: L.T., A.S., T.T., M.P.; Visualization: L.T., M.P.; Supervision: L.T., A.S., M.P.; Project administration: T.T., M.P.; Funding acquisition: T.T;
Acknowledgements
We thank Mark Fortini, Bertram Gerber and Claire Cronmiller for sharing fly stocks and reagents. We thank Epp Väli and Jan Erik Alliksaar for technical assistance. We are grateful to Jürgen Tuvikene, Allan-Hermann Pool, and Richard Tamme for critical comments on manuscript. Stocks obtained from the Bloomington Drosophila Stock Center (NIH P40OD018537) were used in this study. We acknowledge the Drosophila Genomics Resource Center supported by NIH grant 2P40OD010949.
Footnotes
The Supplemental figures and table was added
References




