

Abstract
The insula contributes to the detection and integration of salient events during goal-directed behavior and facilitates the interaction between motor, multisensory, and cognitive networks. Task-fMRI studies have suggested that experience with singing can enhance access to these resources. However, the long-term effects of vocal motor training on insula-based networks are currently unknown. In this study, we used resting-state fMRI to explore experience-dependent differences in insula co-activation patterns between conservatory-trained singers and non-singers. We found enhanced insula connectivity in singers compared to non-singers with constituents of the speech sensorimotor network, including the cerebellum (lobule VI, crus 2), primary somatosensory cortex, the parietal lobes, and the thalamus. Moreover, accumulated singing training correlated positively with increased co-activation in bilateral primary sensorimotor cortices in the somatotopic representations of the larynx (left dorsal anterior insula, dAI) and the diaphragm (bilateral dAI)—crucial regions for motorcortical control of complex vocalizations—together with the thalamus (bilateral posterior insula/left dAI) and the left putamen (left dAI). The results of this study support the view that the insula plays a central role in the experience-dependent modulation of sensory integration within the vocal motor system, possibly by optimizing conscious and non-conscious aspects of salience processing associated with singing-related bodily signals.
INTRODUCTION
Research over the last two decades has provided compelling evidence that the extensive practice and performance of high-level motor, sensory, and multimodal integration skills required for mastering a musical instrument can have profound effects on the functional and structural organization of the brain (Elmer et al., 2016; Klein et al., 2016; Schlaug, 2015; Vuust et al., 2014; Zatorre, 2013). Musical training has therefore been widely used as a framework for studying human neuroplasticity (Munte et al., 2002), demonstrating that adaptive changes in the brain can be linked to the rehearsed action, encompass quantifiable behavioral correlates (i.e., improved performance), occur in response to short and long-term training, while being pronounced when formal training began during a sensitive period before the age of seven years (for review, see Herholz and Zatorre, 2012). Trained singers, on the other hand, have rarely been studied in this context. They differ from instrumentalists as they (i) commence with formal training routinely later (Kleber et al., 2016; Penhune, 2011; Schirmer-Mokwa et al., 2015) and (ii) optimize a motor system for music production that has already undergone substantial maturation during speech and language development, and informal singing experience (Dalla Bella et al., 2007; Skeide and Friederici, 2016; Smith, 2006; Weiss-Croft and Baldeweg, 2015). As professional singers still require a large amount of deliberate practice to meet the musically prescribed acoustic standards, the current study used singers as a model for experience-dependent neuroplasticity within the speech-motor system (Kleber and Zarate, 2014; Zarate, 2013).
The neural pathways underlying basic vocalizations have been demonstrated by research in primates (Jurgens, 2002, 2009), whereas higher-level aspects of vocal motor production have been studied in the context of human speech (Price, 2012). Together, they revealed a hierarchically organized structure-function relationship in which basic vocal pattern are generated in the brain stem and the periaqueductal gray, which interact with limbic regions, whereas volitional sensorimotor control of complex vocalizations takes place in the cortex (Hage and Nieder, 2016; Simonyan and Horwitz, 2011). The presumed interactions between these regions are described and formalized by neurocomputational models in the context of speech motor processes (Bouchard et al., 2013; Guenther, 2016; Guenther and Hickok, 2015; Hickok, 2017; Houde and Chang, 2015; Parrell et al., 2017; Rodriguez-Fornells et al., 2009; Tian and Poeppel, 2010). Specifically, the DIVA (Directions Into Velocities of Articulators) model suggests that dorsal stream speech-motor control involve a sensory integration interface encompassing auditory, ventral somatosensory, and inferior parietal cortices, which interact via the arcuate fasciculus with the articulatory motor network located in ventral primary- and pre-motor regions (Guenther and Hickok, 2015). Moreover, cortico-thalamic loops including the supplementary motor area, the basal ganglia, and the cerebellum contribute to feedback and feedforward transformations supporting vocalizations (Dick et al., 2014; Hertrich et al., 2016).
The insular cortex is not typically a major player in these models but has long been considered to contribute to motor aspects of speech production (Dronkers, 1996; Price, 2000; Riecker et al., 2005), based on its involvement in vocalization-related autonomic and perceptual-motor processes (Craig, 2002, 2009), and its structural connectivity with constituents of the dorsal sensorimotor stream (Augustine, 1996; Eickhoff et al., 2009; Hickok, 2017; Remedios et al., 2009; Riecker et al., 2008). In a general context, the insular cortex is functionally and structurally connected to a vast number of cortical and subcortical brain regions subserving sensory, emotional, motivational and cognitive processes (Gogolla, 2017; Nomi et al., 2018). As an actively participating integration hub within these networks, its multifaceted functional roles range from conscious and non-conscious integration of bodily information (Craig, 2002, 2009; Critchley et al., 2004; Karnath and Baier, 2010; Olausson et al., 2002), the preservation of homeostatic states in autonomic systems that are critical to body regulation (Gu and FitzGerald, 2014; Strigo and Craig, 2016), to salience detection and valence assessment of incoming sensory inputs (Berret et al., 2019; Singer et al., 2009), as well as the coordination of large-scale network dynamics associated with higher-level cognitive processes (Menon and Uddin, 2010; Seeley et al., 2007; Uddin, 2015). Speech specific clinical and functional imaging data suggest a bilateral engagement for linguistic processes and a left-hemispheric dominance for non-linguistic aspects of speech-related perceptual and motor processes in the dorsal anterior insula (Ackermann and Riecker, 2010; Oh et al., 2014). Singing, on the other hand, shows a right-hemispheric dominance for perceptual and motor processes (Ackermann and Riecker, 2004; Jeffries et al., 2003; Riecker et al., 2000), possibly related to the processing of prosodic or melodic elements of vocalizations (Oh et al., 2014). This notion is supported by neuroimaging studies with classically trained singers, indicating a role of the right anterior insula in the stabilization of vocal pitch patterns during sensory feedback perturbations (Kleber et al., 2017; Kleber et al., 2013). However, long-term effects of vocal sensorimotor training on insula-based networks in singers are currently unknown.
The current study addressed this question by employing resting-state (rs-)fMRI to quantify experience-dependent differences in large-scale brain networks between trained singers and non-singers, following the notion that rs-fMRI reflects the record of repeated task-dependent synchronized activation between brain regions (Guerra-Carrillo et al., 2014). Specifically, we used a seed-based approach to explore temporally correlated spontaneous fluctuations of blood oxygen level–dependent (BOLD) between a priori defined regions of interests (ROI) with all other brain regions (Biswal et al., 1995; Fox and Greicius, 2010; Fox and Raichle, 2007). Given the insula’s involvement in the brain’s language and speech system (Ardila et al., 2014) and in the coordination of large-scale brain networks (Uddin et al., 2014), we have chosen insula ROIs following a tripartite subdivision model (Deen et al., 2011). This model is based on structural and functional co-activation patterns across task domains, which suggest both unique and overlapping functional profiles encompassing the posterior insula (PI), the dorsal anterior insula (dAI), and the ventral anterior insula (vAI) (Deen et al., 2011; Nomi et al., 2018; Uddin et al., 2014). We expected to find enhanced insula connectivity in singers compared to non-singers (Kleber et al., 2017; Kleber et al., 2013), specifically between the AI with regions of the dorsal sensorimotor speech stream involved in vocal tract coordination, analogous to task-related findings with musicians trained in playing an instrument (Zamorano et al., 2017). A high level of task-related structure function specificity would provide converging evidence for a central involvement of the insula in vocal motor control and may furthermore help delineating experience-dependent changes within sensory integration networks.
METHODS
Participants
A total of twenty-five right-handed subjects without reported history of neurological or psychiatric disease participated in this study. Participants were subdivided in two groups based on their expertise in singing. Twelve professional singers (6 female, 32.7 ± 8.6 yrs) with music conservatory education participated in this study. Trained singers took their first formal singing lesson at the average age of 16 years (± 6.7) and accumulated an average of 12957 hours (range: 1456-38220) of singing experience. Thirteen non-singers (6 female, 28.1 ± 7.3 yrs) participated from the University of Tübingen medical school. Neither of them received singing training nor reported involvement in occasional singing activities (e.g., choirs, informal rock bands etc.). All participants were informed about the details of the study and provided written consent. The study was conducted under a protocol approved by the research ethics board of the University of Tübingen.
Image Acquisition
Magnetic resonance imaging was performed using a 3-Tesla whole body MRI Scanner (Siemens MAGNETOM Prisma™ 3T, Erlangen, Germany). For each subject, 225 echo-planar volumes were acquired (repetition time, 2000 ms; echo time, 30 milliseconds; matrix dimensions, 64 × 64; field of view, 1260 mm; 30 transversal slices; slice thickness, 4 mm; flip angle, 90 degrees) over a period of 7.5 minutes with the eyes closed. The structural imaging data consisted of T1-weighted images (mprage, repetition time, 2300 milliseconds; echo time, 4.18 milliseconds; matrix dimensions, 512 × 512; field of view, 256 × 256 mm; 176 slices; slice thickness, 1 mm; flip angle, 9 degrees).
Pitch-matching accuracy
Participants’ ability to sing on pitch was quantified using a pitch-matching paradigm. Musical interval are presented via headphones and participants are prompted to immediately reproduce the pitch height of each tone with their singing voice. A total of 36 musical intervals were presented (17 ascending, 15 descending, and four same tone) using Max/MSP software to control the experiment (Cycling 74, San Francisco, California, USA). The first tone always started at the fundamental frequency of 311.13 Hz for females (D#4 in musical notation) and 155.565 Hz for males (D#3 in musical notation). Each tone was played with a duration of 900-ms, separated by a 200-ms gap. The second tone differed from the first by a value between zero and ± nine semitones (one semitone corresponds to a frequency ratio of 1.0595 or 100 cents). Vocal performance was recorded and stored in wave format for offline automated analyses of pitch-matching accuracy.
Neuroimaging data preprocessing
Following our previous approach (Zamorano et al., 2017; Zamorano et al., 2019), functional image were preprocessed with the Data Processing Assistant for Resting-State fMRI (DPARSF; Chao-Gan and Yu-Feng, 2010), based on the Statistical Parametric Mapping software package (SPM8; http://www.fil.ion.ucl.ac.uk/spm) and the Data Processing & Analysis of Brain Imaging toolbox (DPABI; http://rfmri.org/DPABIDPARSF_V3.1_141101). The first 10 volumes from each data set were discarded prior to preprocessing. Following slice-time correction and co-registration, gray and white matter were segmented from co-registered T1 images using the unified segmentation model (Ashburner and Friston, 2005). The resulting parameter file was used to normalize the functional images (3mm3 voxel size) to standard Montreal Neurological Institute (MNI) stereotactic space, which were subsequently smoothed with an isotropic Gaussian kernel (FWHM: 6mm3). Nuisance regression parameters included white matter, CSF, and the six head motion parameters. WM and CSF masks were generated using SPM’s tissue probability maps (empirical thresholds: 90 % for WM mask and 70 % for CSF mask). No global signal regression was performed to avoid introducing distortions of BOLD signal (Murphy et al., 2009). Head motion was below 2.0 mm maximum displacement or 2.0° of any angular motion across all participants. A temporal filter (0.006–0.1 Hz) was applied to reduce low frequency drifts and high frequency physiological noise.
Functional connectivity analysis
Statistical analyses were performed in SPM8 to assess voxel-wise connectivity maps of posterior, ventral anterior and dorsal anterior insula ROIs in each hemisphere. The six insula ROIs consisted of parcellated insular subdivisions (posterior, PI; dorsal anterior, dAI; and ventral anterior insula, vAI) in MNI stereotactic space based on the clustering of functional connectivity patterns during resting state. These template images were kindly provided by Deen and colleagues (2011).
First, the main connectivity patterns across participants (N=25) were determined by entering z-transformed connectivity maps into one-sample t-tests for each insula ROI (Figure 1). Significance threshold for voxel-wise statistics was set to p<0.05 familywise error corrected (FWER), in order to validate the results against previously published patterns (Deen et al., 2011; Uddin et al., 2014).

Main patterns of functional connectivity across all participants for ventral anterior insula (vAI), dorsal anterior insula (dAI), and posterior insula (PI) subdivisions. Significance thresholds were set at P<0.05, familywise error (FWER) corrected. Connectivity maps for left hemisphere seeds are shown on the left; maps for right hemisphere seeds are on the right.
Subsequently, voxel-wise differences between insula-connectivity maps were computed between singers and non-singers using independent two-sample t-tests for each insula ROI. A cluster-extent based thresholding method was employed to increase sensitivity for detecting true activations in studies with moderate sample sizes (Friston et al., 1994; Smith and Nichols, 2009; Woo et al., 2014). This methods accounts for the fact that individual voxel activations are not independent of the activations of their neighboring voxels in spatially smoothed data (Friston et al., 2000; Heller et al., 2006; Wager et al., 2007) and hence used to effectively reduce the possibility of obtaining false positive clusters while improving the degree of confidence in inferences about specific voxels (Woo et al., 2014).
Monte Carlo simulation was as implemented in DPABI’s instantiation (Song et al., 2011; Yan et al., 2016) of AlphaSim (Cox, 1996) was used in combination with the DPABI gray matter mask to determine cluster-extent thresholds at P < 0.05 FWER correction for multiple comparisons. Following the recommendations for cluster-extent thresholding in fMRI analyses, a stringent primary voxel-level threshold of P< 0.0001 was applied using smoothness estimation based on the spatial correlation across voxels to avoid inaccurate FWER correction (Woo et al., 2014). Clusters surviving the FWER probability threshold were used for statistical inference. T-values of significantly activated peak-voxels within clusters are presented as MNI coordinates. Effect sizes using Cohen’s d (d = 2t / sqrt(df)) were computed and posteriorly adjusted [unbiased Cohen’s d; dunbiased = d (1 – (3 / 4df -1))] to control for overestimation of the effect size due to the moderate sample size (Cohen, 2013).
The Automated Anatomical Labeling atlas of Tzourio-Mazoyer (aal, Tzourio-Mazoyer et al., 2002) was used to determine anatomical regions. For regions already cytoarchitectonically mapped, we used the Anatomy Toolbox (Eickhoff et al., 2005).
Regression analysis
Two regression analyses were performed in SPM8 to correlate insula ROI connectivity maps with (i) pitch-matching accuracy across all participants and (ii) the accumulated musical training in trained singers. Cluster extent FWER correction was applied as detailed above.
RESULTS
Voxel-wise functional connectivity of insula subdivisions
Main connectivity patterns across all participants
Whole-brain connectivity of PI, dAI, and vAI (left and right, respectively) across participants are shown in Figure 1. The resulting connectivity patterns were consistent with those reported earlier (Cauda et al., 2011; Deen et al., 2011; Uddin et al., 2014).
The posterior insula (PI) connectivity pattern was strongly centered on regions involved in sensory and motor processes, including the bilateral insulae and adjacent operculae, sensorimotor (S1, S2, and M1) and pre-motor cortices, the supplementary and pre-supplementary motor area (SMA and pre-SMA), posterior and mid cingulate cortex (PCC and MCC), the parietal lobe (supramarginal gyrus, superior and inferior parietal gyrus, and the precuneus), the temporal lobe (superior temporal, Heschl’s, and fusiform gyrus), the inferior frontal gyrus (Broca’s area), occipital lobe (calcarine, lingual, and cuneus), and the cerebellum.
The dorsal anterior insula (dAI) connectivity pattern was centered on the bilateral insulae and both adjacent rolandic and frontal opercula, the primary and secondary somatosensory regions (S1 and S2), the motor cortices (primary, pre-motor, and SMA); the occipital lobe (calcarine, lingual, and cuneus); the temporal and parietal lobes (superior temporal gyrus, Heschl’s and fusiform gyrus, temporal pole; supramarginal, superior and inferior parietal gyrus, and the precuneus), dorsolateral prefrontal ortex (DLPFC) and inferior frontal cortex, cingulate cortex (posterior, PCC; middle, MCC, and anterior, ACC), as well as the cerebellum.
The ventral anterior insula (vAI) connectivity pattern was centered on the bilateral insulae and adjacent rolandic and frontal operculae, the frontal lobe (DLPFC, inferior frontal gyrus, IFG) including frontal limbic cortices (ventro anterior prefrontal cortex, VAPFC; orbitofrontal gyrus, OFC), cingulate cortex (ACC and MCC), SMA and pre-SMA, temporal cortex (e.g., Heschl’s gyrus, temporal pole, superior, middle and inferior temporal gyrus), and the parietal lobe (supramarginal and angular gyrus, precuneus).
Subcortically, all three insula sub-regions showed co-variation patterns with the basal ganglia (putamen, pallidum and caudate) and the thalamus.
Differences in insula connectivity between trained singers and non-singers
T-contrasts of connectivity maps between singers and non-singers revealed increased functional connectivity for each insula subdivision (Figure 2 and Table 1). Reversed comparisons yielded no significant differences.

T-maps showing significant group differences in functional insula connectivity during resting-state. Significance thresholds for between-group differences were set at p>0.05 (FWER), using a cluster-extent based thresholding method. Detailed information is provided in Table 1. Abbreviations: IPL, inferior parietal lobe; SPL, superior parietal lobe; S1, primary somatosensory cortex; IV-VI and Crus II, cerebellar lobules.
The left and right PI (Figure 2A and Table 1) showed increased functional connectivity with lobule VI of the cerebellum.
The left dAI (Figure 2B, Table 1) showed increased functional connectivity with bilateral inferior parietal and primary somatosensory cortex, the cerebellum (lobule VI) and the thalamus. The right dAI showed increased functional connectivity with the cerebellum (lobule VI and Crus 2) and the right S1. Connectivity with the right inferior frontal gyrus failed to reach significance (Broca’s homologue, Area 45).
The left vAI (Figure 2C, Table 1) showed increased functional connectivity with the left superior and inferior parietal cortex. The right vAI did not reveal significant differences.
Regression Results
Multiple regression analyses were performed to determine correlations between insula ROI connectivity maps and two behavioral variables: (i) pitch-matching accuracy and (ii) accumulated singing training (Table 2).
Pitch-matching accuracy across all participants revealed no significant correlations with insula connectivity maps.
Accumulated musical training in trained singers revealed significant correlations with insula connectivity maps in the following regions (Figure 3, Table 2):

{kind=link}
{kind=link}
{kind=link}
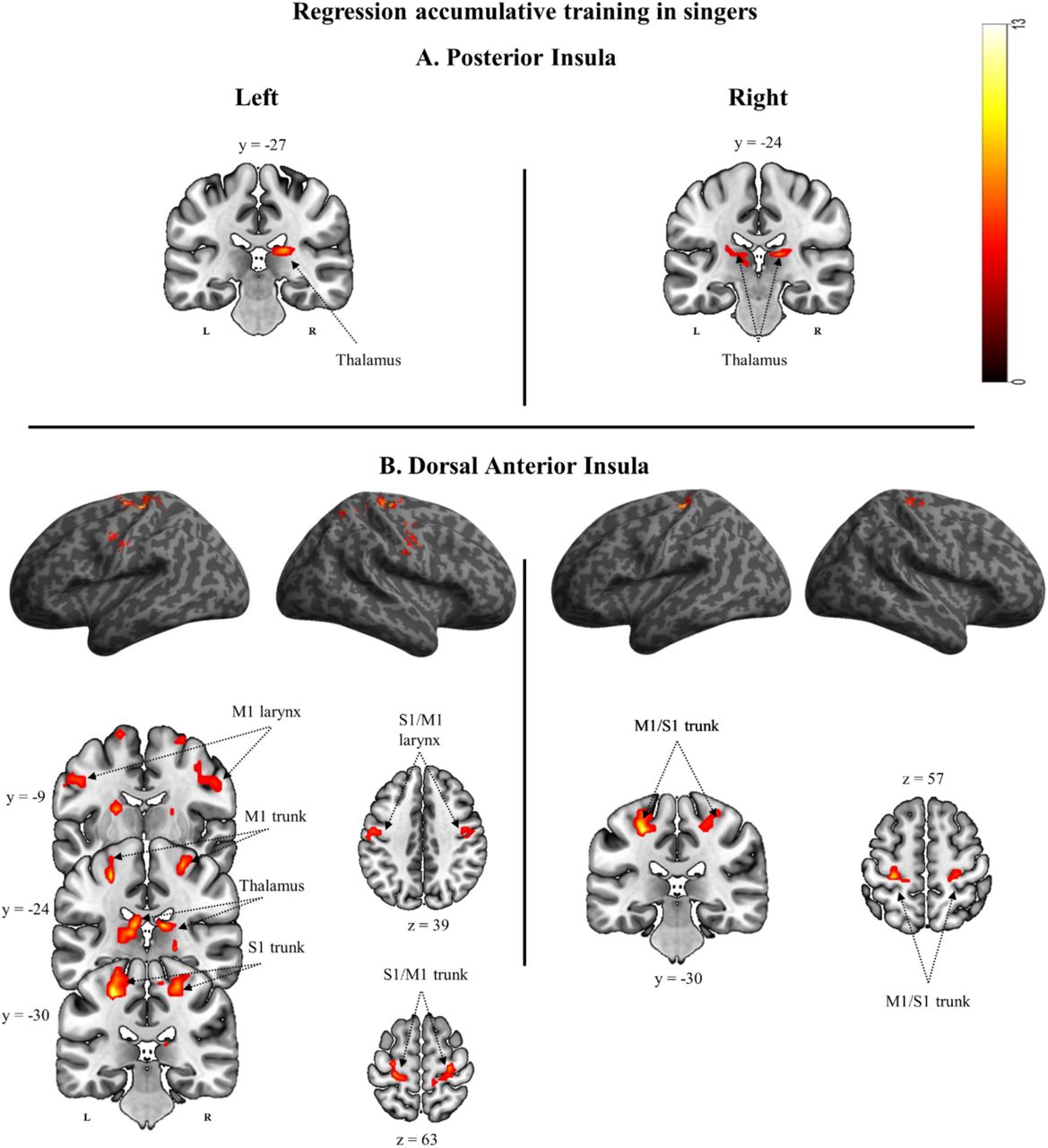
Results from regression analyses for each insular ROI to assess connectivity maps in relation to the amount of accumulated years of vocal training in singers. Only results surviving a cluster-extent based threshold of p<0.05 (FWER correction) are shown. Detailed information is provided in Table 2. Abbreviations: M1, primary motor cortex; S1, primary somatosensory cortex.
The left and right PI connectivity maps (Figure 3A) showed a positive correlation with accumulated training in the thalamus.
The left dAI maps showed positive correlations with accumulated training in the bilateral representation of the larynx and the diaphragm, both within primary motor and primary somatosensory cortices, as well as in the bilateral thalamus and the left putamen. The right dAI maps showed positive correlations with accumulated training in the bilateral representation of the diaphragm within primary motor and primary somatosensory cortex (Figure 3B).
DISCUSSION
In this fMRI resting-state study, we compared temporally correlated spontaneous blood oxygen level–dependent (BOLD) fluctuations between conservatory trained singers and non-singers. Direct comparisons revealed bilaterally increased connectivity in posterior and anterior insula subdivisions in singers with constituents of the sensorimotor speech network. Moreover, accumulated singing training correlated positively with increased insula connectivity, particularly between the dorsal anterior insula with primary motor and somatosensory cortices in the somatotopic representations of the larynx and the diaphragm, as well as with the thalamus and the putamen. These results indicate neuroplastic changes within insula-based networks as a function of singing expertise.
Insula connectivity patterns across groups
Previous studies identified a vast number of functional and structural connections between the insula and both cortical and subcortical brain regions subserving sensory, emotional, motivational and cognitive processes (Gogolla, 2017; Nomi et al., 2018). Specifically, functional segregation based on resting-state co-activation patterns have suggested a tripartite organization of insula subdivisions that contain both unique and overlapping functional profiles (Deen et al., 2011; Uddin et al., 2014). These profiles encompass the posterior insula (PI), the dorsal anterior insula (dAI), and the ventral anterior insula (vAI) and have been supported by the insula’s structural organization (Nomi et al., 2018). In the current study, we replicated previous results (Deen et al., 2011; Uddin et al., 2014; Zamorano et al., 2017; Zamorano et al., 2019), demonstrating a predominantly “cognitive” frontoparietal connectivity pattern of the dAI, a stronger co-activation pattern of the vAI with regions involved in affective processing, and a sensorimotor co-activation pattern for the PI. We furthermore confirmed an overlap between co-activated areas across insula subdivisions (Nomi et al., 2018). Direct group comparisons showed significantly enhanced connectivity across all insula subdivisions in trained singers relative to non-singers, but not for the reversed comparison.
Enhanced PI connectivity in singers versus non-singers
We found increased connectivity in conservatory trained singers between bilateral PI and lobule VI of the cerebellum, a somatotopically organized sensorimotor region that receives somatosensory afferents from the vocal tract and sends out efferents to motor areas (Grodd et al., 2005; Grodd et al., 2001; O’Reilly et al., 2010; Stoodley and Schmahmann, 2010), including those engaged in speaking and singing (Brown et al., 2004; Brown et al., 2006). This is consistent with the unique profile of the PI, which is predominantly associated with sensorimotor processes and the interoceptive representation of visceral and sensory feelings from the body (Berret et al., 2019; Chang et al., 2013; Craig, 2002; Olausson et al., 2002; Pugnaghi et al., 2011; Segerdahl et al., 2015), including orofacial regions (Ikenoue et al., 2018; Sato et al., 2017; Tsutsumi et al., 2018; Yoshida et al., 2017).
Lesions of the PI (Baier et al., 2011) as well as the cerebellar lobule VI give rise to speech motor disorders (Ackermann et al., 2007; Ackermann et al., 1992). Lobule VI shows moreover overlapping functional activation for both the production and perception of song and speech (Ackermann, 2008; Callan et al., 2007; Mathiak et al., 2002). A role in perceptual processes links the cerebellum to predictive mechanisms in both motor and non-motor (cognitive) aspects of language (Argyropoulos, 2016; Bastian, 2006; Friston and Herreros, 2016; Guenther, 2006; Guenther et al., 2006; Knolle et al., 2012; Shadmehr et al., 2010). Based these functions, we suggest that the observed increased PI-cerebellar connectivity in trained singers may be related to perceptual optimization and prediction processes (Baumann et al., 2015; Moberget and Ivry, 2016), which has been associated with enhanced behavioral performance in musicians (Baer et al., 2015; Kleber et al., 2017; Lu et al., 2014) and smooth sensorimotor tuning (Ackermann, 2008; Fujioka et al., 2015).
Enhanced dAI connectivity in singers versus non-singers
In the current study, we found increased functional connectivity in singers between the left dAI with the bilateral inferior parietal lobe (IPL), adjacent S1 (area 2), and the cerebellum (lobule VI). The right dAI showed increased functional connectivity with right S1 (area 2) and the cerebellum (lobule VI and crus 2). Crus 2 has been linked to executive control, whereas lobule VI has also been associated with salience processing (Baumann et al., 2015; Habas et al., 2009), a task that requires top-down attention and cognitive control in the context of planned actions (Uddin, 2015). Enhanced connectivity within the salience network during resting state has moreover been found in professional musicians (Luo et al., 2014; Zamorano et al., 2017; Zamorano et al., 2019).
Although strongly interconnected with the PI (Kurth et al., 2010), the unique functional profile of the dAI is associated with salience detection and high-level cognitive processes, such as task-related attention switching, inhibition, and error awareness (Klein et al., 2013; Menon and Uddin, 2010; Uddin, 2015), in line with results in the current study. The typical co-activation pattern of the dAI encompasses functionally diverse brain regions, including those that pertain to phonological and speech processes (Uddin et al., 2014), which links the anterior insula to higher-order aspects of speech motor control (Ackermann and Riecker, 2004, 2010). Specifically, it has been suggested that the dAI may act as a relay between more cognitive aspects of verbal communication and motor coordination, thus contributing to serial processing within a core speech network that involves BA44, the basal ganglia, and the cerebellum (Adank, 2012; Eickhoff et al., 2009; Oh et al., 2014). This is consistent with a role of the insula in sound- to-speech transformations within the dorsal stream speech network (Cloutman et al., 2012; Hickok, 2017).
The co-activation patterns of lobule VI suggest that it also contributes to cognitive aspects of language processes together with frontoparietal regions (Stoodley et al., 2012; Vias and Dick, 2017). The inferior parietal lobe (IPL) and S1 (area 2) are tightly interconnected (Borich et al., 2015) and serve as an interface to process proprioceptive signals and body-related information (Prevosto et al., 2011; Tsakiris et al., 2007). They also support the preparation and coordination of speech-related vocal tract movements (Bouchard et al., 2013; Golfinopoulos et al., 2011; Nasir and Ostry, 2006; Tremblay et al., 2003), contributing nearly 7 times more to the vocal motor system in humans than in non-human primates (Kumar et al., 2016). Previous studies found that trained singers show increased activation in ventral S1, IPL, and the cerebellum during normal singing (Kleber et al., 2010) as well as increased gray-matter volume in S1/IPL relative to non-singers (Kleber et al., 2016). Functionally, the IPL and the cerebellum interact with the dAI to dynamically adapt for sensory feedback perturbations during singing, allowing greater pitch-matching accuracy through enhanced sensorimotor feedback and feedforward control mechanisms in singers (Jones and Keough, 2008; Kleber et al., 2017; Kleber et al., 2013; Zarate and Zatorre, 2008). Such functions have been supported by studies investigating the neural correlates of sensorimotor control of speech and song production (Behroozmand and Sangtian, 2018; Correia et al., 2015; Finkel et al., 2019; Guenther and Vladusich, 2012; Hickok, 2017; Houde and Chang, 2015; Parkinson et al., 2012; Shum et al., 2011).
Taken together, enhanced connectivity with the dAI in singers may indicate enhanced sequencing and sensorimotor mechanisms (Ackermann, 2008; Bohland and Guenther, 2006; Ziegler and Ackermann, 2017) as well as enhanced cognitive control pertaining to the execution of planned actions (Eickhoff et al., 2009; Ito, 2008; Pisotta and Molinari, 2014; Stoodley et al., 2012), which may facilitate the organization of motor patterns in the context of musically defined performance goals (Zatorre and Baum, 2012).
Enhanced vAI connectivity in singers versus non-singers
The vAI is more strongly associated with valence attribution, body awareness, and affective processing (Craig, 2009, 2010). In the current study, trained singers showed increased co-activation with the left vAI involved the superior parietal lobe, intraprietal sulcus, and the right prefrontal lobe (BA10/Frontal Pole). The latter is involved in mediating limbic reward and learning processes (Denny et al., 2014; Kufahl et al., 2005). However, connection patterns of right frontal pole (FP1) suggest that it also plays a critical role in supporting high-level cognitive processes, such as language and working memory (Gilbert et al., 2006; Liu et al., 2013; Ray et al., 2015). This is in line with a functional specialization of BA10, in which lateral regions may act as a gateway to shift attention from external (sensory) monitoring to internal activation of behavioral responses (Howe et al., 2013). Such cued response representations are a typical feature of musical performances, for example when the conductor signals the singer the precisely timed (vocal) entrance after a period of rest.
Although the observed increased connectivity with the SPL in the current study does not comfortably fit within its unique profile, studies have associated the vAI also with task-level control and focal attention (Nelson et al., 2010), perhaps based on its structural connection to the parietal lobe via the inferior frontooccipital fasciculus (Nomi et al., 2018). The parietal lobe is moreover part of a frontoparietal attentional control system (Uddin, 2015), which contributes to task performance when stimulus-response associations have been well trained and can be prepared in advance (Corbetta and Shulman, 2002). SPL activation in musicians has also been associated with heightened sensorimotor control, increased attention, and working memory load (Foster and Zatorre, 2010; Pallesen et al., 2010; Stewart et al., 2003; Strait et al., 2010), suggesting that increased vAI connectivity with frontoparietal areas in trained singers in the current study could reflect attentional processes related to task-specific multisensory awareness (Janata et al., 2002; Nelson et al., 2010; Simmons et al., 2013).
Insula connectivity as a function of accumulated singing training
The insula contributes to the spinothalamocortical pathway, in which the lamina I projects afferent information from body tissues via the thalamus to both the somatosensory area 3a and the posterior insula, the so-called interoceptive cortex (Craig, 2002, 2009, 2010). These signals are re-represented, integrated, and affectively evaluated in the insula along a posterior-to-anterior progression scheme, generating a comprehensive representation of one’s feelings. In the current study, we found enhanced connectivity as a function of singing experience among constituents of this pathway. Specifically, between (i) the bilateral PI with the thalamus, (ii) the dAI (bilaterally) with the sensorimotor representation (S1/M1) of the diaphragm, and (iii) the left dAI furthermore with the bilateral sensorimotor representation (S1/M1) representation of the larynx, bilateral thalamus, and the left putamen. We propose that the observed enhanced functional connectivity among these regions in the current study reflects a role of the insula as a major integration site for task-relevant bodily information based on experience (Critchley et al., 2004; Kleber et al., 2017; Schirmer-Mokwa et al., 2015; Zamorano et al., 2017; Zamorano et al., 2019).
This interpretation is supported by the specific functional involvement of the co-activated areas, which process visceral and somatosensory information underlying voluntary respiratory, laryngeal, and articulatory sensorimotor control (Banzett et al., 2000; Bouchard et al., 2013; Brown et al., 2008; Conant et al., 2014; Conant et al., 2018; Khalsa et al., 2009; Pfenning et al., 2014). Vocalization related movements are accompanied by proprioceptive, kinesthetic, and interoceptive consequences, which in turn are integrated with the motor system to guide song and speech production (Bernardi et al., 2015; Gozaine and Clark, 2005; Nasir and Ostry, 2006; Simonyan and Horwitz, 2011; Smotherman, 2007; Tremblay et al., 2003; Wyke, 1974). For example, every movement specifically stimulates a corresponding somatosensory region in S1 (Giraud and Poeppel, 2012; Schomers and Pulvermuller, 2016), which is intrinsically connected to ipsilateral M1 and IPL (Borich et al., 2015; Bouchard et al., 2013), involves the thalamus (Ackermann and Riecker, 2010; Hwang et al., 2017), and becomes tightly associated with corresponding auditory-motor transformations (Ito et al., 2016; Ito and Ostry, 2010, 2012; Ito et al., 2015; Ito et al., 2009). Moreover, M1 sends its strongest subcortical projections to the putamen (Simonyan and Horwitz, 2011), which shows increased activations in more proficient singers (Kleber et al., 2010; Segado et al., 2018; Zarate and Zatorre, 2008).
Importantly, the insula is substantially involved in the integration of sensory and visceral inputs with motor modalities to coordinate respiratory and vocal tract musculature (Ackermann and Riecker, 2010; Dronkers, 1996; Oh et al., 2014; Simonyan et al., 2007), based on its connectivity with cortical areas associated with different aspects of sensorimotor and cognitive control of speech (Battistella et al., 2018). At the cortical level, the voluntary control of expiratory, laryngeal, and articulatory activity is somatotopically overlapping in ventral sensorimotor cortex (Belyk and Brown, 2017; Kumar et al., 2016; Loucks et al., 2007). Conversely, the voluntary regulation of inspiratory patterns, such as taking a deeper than normal breath before a long musical phrase (Ackermann and Riecker, 2010; Leanderson et al., 1987; Smotherman et al., 2010; Watson and Hixon, 1985), is more dorsally located within the somatotopic representation of the trunk and diaphragm (Eickhoff et al., 2008). Both regions are activated during pitch-matching tasks in trained singers (Kleber et al., 2017; Kleber et al., 2013). Increased connectivity strength with these areas as a function of singing experience in the current study may therefore indicate that the insula contributes increasingly to body awareness in singers, thus supporting optimal vocal performance via enhanced integration of task-related bodily sensations (Kleber et al., 2017; Kleber et al., 2013; Schirmer-Mokwa et al., 2015; Zarate, 2013).
Limitations
Limitations of this study include the fact that resting state fMRI exposes participants to continuous scanner noise, which may reduce the robustness and the replicability of functional connectivity findings within the somatosensory, auditory, and motor networks (Andoh et al., 2017). However, robust experience-dependent differences in S1/M1 in vocalization areas in the current study replicate previous findings with classical instrumental musicians, in which greater experience yielded greater insular connectivity with the sensorimotor hand area (Zamorano et al., 2017). Together, this supports a critical role of insula-based networks in body awareness underlying music production. However, taking into account that participants in the current study were conservatory trained singers, we cannot rule out the possibility that co-activation patterns differ in amateur-level singers. Finally, future studies should replicate these results in bigger samples to improve the generalizability of these results.
Conclusions
This study has examined the impact of singing training on insula-based network connectivity. The results of this research support the idea that the insula plays a central role in modulating sensory integration within the vocal system (Cloutman et al., 2012; Nieuwenhuys, 2012; Parkinson et al., 2012), perhaps by regulating conscious and non-conscious aspects of salience processing associated with singing-related bodily processes (Allen et al., 2016; Chong et al., 2017). This may optimize the processing, manipulation, and transformation of sensory information based on experience (Zarate, 2013; Zarate and Zatorre, 2005). Together, our data add to the growing body of literature, suggesting that trained singers may increasingly rely on somatosensory feedback processing to support vocal motor control (Jones and Keough, 2008; Kleber et al., 2017; Kleber et al., 2010; Kleber et al., 2013; Mürbe et al., 2004; Zarate and Zatorre, 2008). It also accords with earlier observations, demonstrating heightened sensitivity to interoceptive and exteroceptive stimuli in musicians and dancers compared to laymen (Christensen et al., 2017; Schirmer-Mokwa et al., 2015; Zamorano et al., 2014). As changes in temporally correlated spontaneous fluctuations of blood oxygen level–dependent (BOLD) signals can be associated with both the type of training and the corresponding behavioral performance (Baldassarre et al., 2012; Fauvel et al., 2014; Luo et al., 2012; Palomar-Garcia et al., 2017; Vahdat et al., 2011; Ventura-Campos et al., 2013; Zamorano et al., 2017), our findings provide converging evidence for a central role of the insula in speech and music processing.
References




