

Summary
Reef-building corals live in a mutualistic relationship with photosynthetic algae (family Symbiodiniaceae) that usually provide most of the energy required by the coral host. This relationship is sensitive to temperature stress; as little as a 1°C increase often leading to collapse of the association. This sensitivity has led to interest in the potential of more stress tolerant algae to supplement or substitute for the normal Symbiodiniaceae mutualists. In this respect, the apicomplexan-like microalga Chromera is of particular interest due to its greater temperature tolerance. We generated a de novo transcriptome for a Chromera strain isolated from a GBR coral (“GBR Chromera”) and compared to those of the reference strain of Chromera (“Sydney Chromera”), and to those of Symbiodiniaceae (Fugacium, Cladocopium and Breviolum), as well as the apicomplexan parasite, Plasmodium falciparum. By contrast with the Symbiodiniaceae, the two Chromera strains had a high level of sequence similarity evident by very low levels of divergence in orthologous genes. Although KEGG categories provide few criteria by which true coral mutualists might be identified, they do supply a molecular rationalization for the ubiquitous association of Cladocopium strains with Indo-Pacific reef corals. The presence of HSP20 genes may underlie the higher thermal tolerance of Chromera.
Introduction
The ecological success of reef-building corals is generally attributed to their ability to establish mutualistic relationships with specific photosynthetic algae – members of the dinoflagellate family Symbiodiniaceae (formerly, the genus “Symbiodinium”). However, as is now widely appreciated, this relationship breaks down under environmental stress, and coral reefs globally are under threat as a consequence of the increasing frequency of weather events that exceed the thermal thresholds of corals (Hughes et al., 2017). In addition to Symbiodiniaceae, a wide range of uncharacterized eukaryotes are associated with corals, including apicomplexan-related lineages (ARLs) (Clerissi et al., 2018) that can sometimes occur in high abundance (Kwong et al., 2019). Two of these ARLs isolated in association with corals, Chromera velia (Moore et al., 2008) and Vitrella brassicaformis (Obornik et al., 2012), constitute the newly defined phylum Chromerida. As the closest free-living relatives of the parasitic Apicomplexa, these photoautotrophic alveolates are of considerable scientific interest (Moore et al., 2008). Whole genome sequencing (Woo et al., 2015) has recently reaffirmed the close relatedness of Chromera and Vitrella.
The nature of the relationship between corals and Chromera has been a subject of debate. Given its photosynthetic ability (Moore et al., 2008) and its ability to colonize coral larvae (Cumbo et al., 2013), it was initially thought that Chromera might be an alternative coral mutualist, potentially bringing the benefit of higher thermal tolerance than most Symbiodiniaceae (Visser et al., 2012; Chakravarti et al., 2019). However, several lines of evidence now imply otherwise. It has recently been shown that the transcriptomic response of the coral host post Chromera uptake (Mohamed et al., 2018) differed markedly from that of the same coral to a mutualistic strain of Symbiodiniaceae (Mohamed et al., 2016), and resembled the response to incompatible (“incompetent”) Symbiodiniaceae strains (Voolstra et al., 2009). The apparently hostile responses of coral larvae to Chromera during infection suggested that Chromera is more likely to be a parasite or a commensal of corals rather than a mutualist (Mohamed et al., 2018). Other lines of evidence support this suggestion (Barott et al., 2011; Janouškovec et al., 2012, 2013), including a recent meta-analysis which implies that Chromera is near exclusively associated with coral biogenous sediments (Mathur et al., 2018).
The present work sought to address two specific issues, in both cases making use of a de novo transcriptome assembly generated for a strain of Chromera isolated from corals on the Great Barrier Reef (GBR). Whilst Chromera was originally isolated from a Sydney Harbor coral, it is known to have a wide distribution (Janouškovec et al., 2012; Visser et al., 2012), and the diversity within this monospecific genus has not been systematically explored. Given the metabolic diversity that is now known to exist (LaJeunesse et al., 2018) within what was previously known as “Symbiodinium”, the extent to which conclusions about the coral-Chromera interaction based on the GBR isolate are generalizable is unknown.
The first goal of the present study was therefore to estimate the degree of divergence between the GBR strain of Chromera and that originally isolated from Sydney harbor. Given the evidence that Chromera is unlikely to be a coral mutualist, the second goal was to investigate the repertoires of genes that are thought to play roles in symbiosis and environmental stress tolerance in Chromera and compare these with those of three members of the Symbiodiniaceae. Cladocopium goreaui (formerly Clade C1 Symbiodinium) was isolated from a colony of Acropora tenuis on the GBR (Howells et al., 2012), and is mutualistic with many Indo-Pacific corals, particularly Acropora species. Breviolum minutum (formerly Clade B Symbiodinium) was isolated from the Caribbean coral Orbicella faveolata, and is a mutualist of Caribbean corals. Fugacium kawagutii (formerly Clade F Symbiodinium) was originally isolated in association with the Hawaiian reef-building coral Montipora verrucosa and the initial whole-genome analyses followed the assumption that Fugacium is a coral mutualist (Lin et al. 2015). However, Fugacium failed to infect juvenile corals (Yuyama et al., 2016), and the consensus now is that Fugacium is probably a surface associate of corals rather than an endosymbiont (Liu el al., 2018; LaJeunesse et al., 2018; González-Pech et al., 2019). Thus, the expectation was that, with respect to metabolic repertoire, the Chromera strains would resemble Fugacium rather than the known coral mutualists, Cladocopium and Breviolum. Whilst the results suggest that small HSPs may account for the tolerance of Chromera to elevated temperatures, they were inconclusive with respect to the nature of relationships between corals and Chromera or Fugacium. The comparative analyses do, however, provide a molecular rationalization for the near ubiquitous association of Cladocopium with Indo-Pacific corals in general and with Acropora spp in particular.
Experimental procedures
Chromera culture and culturing conditions
A culture of Chromera (Mdig3 strain) from the University of Technology Sydney (Cumbo et al., 2013) was used in this study and referred to as “GBR Chromera”. The identity of the culture was confirmed both by microscopy and by using Chromera-specific PCR primers (Supplementary information Figure 1, Table 1). This Chromera strain was originally isolated from the stony coral Montipora digitata (Acroporidae) from Nelly Bay, Magnetic Island on the inner central part of the Great Barrier Reef. Cultures were maintained at 25 °C in Guillard’s f/2 medium on a 12 h/12 h day and night regime. Note that Chromera was subjected to a variety of treatments prior to RNA extraction in order to ensure that the transcriptome assembly captured as many genes as possible. Culture conditions included control, dark stress, cold shock, heat shock, motile and mixotrophic (for details see Supplementary Methods). In all cases, exponentially growing cultures were separated and subjected to the treatment condition and harvested at the end of the experimental treatment. During culturing no antibiotics were used to exclude any potential contribution of the antibiotic treatment to the mRNA expression in the cultures.
RNA isolation and high-throughput sequencing
50 mL of Chromera cultures were pelleted by spinning the cultures at 3,000 x g for 5 min. Pellets were suspended in 1 ml 0.2 μm sterile FSW and centrifuged at 3,000 x g for 5 min. Pellets were snap frozen in liquid nitrogen and stored at −80°C until further treatment. Total RNA was isolated from ∼80 mg of the frozen Chromera pellets using the RNAqueous® Total RNA Isolation Kit (Ambion). The pellets were lysed twice for 20s at 4.0 ms−1 in Lysing Matrix D tubes (MP Biomedicals, Australia) containing 960 µL of lysis/binding solution plus 80 µL of the Plant RNA Isolation Aid (Ambion, USA) on a FastPrep®-24 Instrument (MP Biomedicals, Australia). RNA was bound to filter cartridges supplied with the kit and washed three times, finally RNA was eluted in 40 µL of the elution solution. RNA quantity and quality were assessed using a NanoDrop ND-1000 spectrometer, Qubit® 2.0 fluorometer and Agilent 2100 bioanalyzer. Messenger RNA (mRNA) was isolated from 1 µg of total RNA and 6 RNA-Seq libraries were prepared using the TruSeq RNA Sample Preparation Kit (Illumina). Libraries were sequenced on an Illumina HiSeq 2000 platform at the Australian Genome Research Facility (AGRF) in Melbourne, Australia. Sequencing produced a total of 189.5 million individual 100 bp paired-end reads (Table 1).
Illumina sequencing and mapping statistics: number of reads and bases of raw and processed data after quality control (adapter removal, trimming and filtering low-quality bases) for each library and percentages of reads successfully mapped onto the GBR Chromera transcriptome. Note that Chromera was subjected to a variety of treatments prior to RNA extraction in order to ensure that the transcriptome assembly captured as many genes as possible.
Processing of Illumina data
The raw Illumina reads were filtered and adapters were clipped using TRIMMOMATIC (v0.32) (Bolger et al., 2014). Reads were filtered based on quality and size as follows; both universal and indexed Illumina adapters were clipped, quality trimming was also performed by removing leading and trailing bases with Phred quality score < 25 and average Phred quality score was calculated in 4 bp sliding windows. Bases were trimmed from the point in the read where average Phred quality score dropped below 20 (i.e. the chances that a base is called incorrectly is 1 in 100) and reads of < 50 bp were also excluded.
De novo assembly and annotation of transcriptome
The trimmed/filtered Illumina reads were used for de novo transcriptome assembly using Trinity (r20140717 version). The assembly was carried out with the recommended protocol described in (Haas et al., 2013) and using options appropriate for de novo transcriptome assembly of strand specific RNA-Seq libraries. Minimum contig length of 500 and read normalization were specified. Trinity collects transcripts with shared sequence identity into clusters that are loosely related to genes. The longest isoform per cluster was selected using a custom Perl script from the assembled Trinity output “assembled transcriptome” for the purpose of annotation. Chromera contigs were annotated by similarity search using batch BLASTX conducted locally against the Swiss-Prot protein database downloaded in September 2014 (E-value cut off 10−3 and maximum 20 hits). Raw BLASTX outputs were imported to Blast2GO suite (version 2.6.5) (http://blast2go.com/b2ghome) for functional annotation and Gene Ontology (GO) assignment. KEGG analysis was also performed using the KEGG Automatic Annotation Server (KAAS) (Moriya et al., 2007) (http://www.genome.jp/kaas-bin/kaas_main) in order to obtain an overview of the associated metabolic pathways. The bi-directional best hit (BBH) method was used to obtain KEGG orthology (KO) assignments.
In order to validate the accuracy of the de novo assembly, reads were mapped back to the de novo-assembled transcriptome using the BOWTIE aligner version 0.12.7 (Langmead and Salzberg, 2012) with default mapping parameters. The percent of the mapped reads as proper pairs was used to assess the assembly quality. Moreover, BLASTN (E-value of ≤ 10−10) was performed against bacterial genomes downloaded from the GenBank, NCBI to determine the percentages of putative bacterial transcripts in the dataset. The completeness of the transcriptome assembly was assessed using BUSCO v3.1.0 making use of the function run_BUSCO.py (-- mode transcriptome) with the Eukaryota_odb9, alveolate_stramenophile and protists_ensembl data (retrieved 23 October 2019).
Phylogenomic analyses
A multi-gene phylogenetic analysis was performed to assess the relative phylogenetic distance between Chromera strains and infer their evolutionary relationship to other Apicomplexans and Symbiodiniaceae. The protein sequences of GBR Chromera were predicted by Transdecoder (Haas et al., 2013). Transcript nucleotide (CDS) and protein sequences for Chromera CCMP2878 strain “Sydney Chromera” were downloaded from CryptoDB (release-37; http://cryptodb.org/cryptodb/), and those for Plasmodium falciparum and Toxoplasma gondii from PlasmoDB and ToxoDB respectively (release-37; http://plasmodb.org/plasmo/ and http://toxodb.org/toxo/). Corresponding data for Symbiodiniaceae species were based on gene models from their respective genome sequencing projects; specifically, Fugacium kawagutii (Lin et al 2015) was obtained from the Symka Genome Database (http://web.malab.cn/symka_new/download.jsp), Cladocopium goreaui (Liu et al 2018) from ReFuGe 2020 site (http://refuge2020.reefgenomics.org/) and Breviolum minutum (Shoguchi et al., 2013) from the OIST Marine Genomics online resource (https://marinegenomics.oist.jp/symb/viewer/info?project_id=21).
The longest transcript for each gene was extracted for all seven species and used to infer orthologous clusters with OrthoFinder (Emms and Kelly, 2015). A total of 693 orthogroups were found to have representative genes in all species, and of these 110 consist of single-copy genes in each species; they represent strictly orthologous gene sets. Amino acid sequences for these 110 orthologous sets were aligned using MAFFT (Katoh and Standley, 2013) and converted into corresponding codon alignment by pal2nal (Suyama et al., 2006). Poorly aligned regions were removed using trimAl (Capella-Gutierrez et al., 2009). Finally, IQ-TREE (Nguyen et al., 2015) was used to perform a partitioned phylogenetic analysis allowing independent estimation of evolutionary model for each protein set. To summarise the effect of phylogenetic relatedness on overlap between gene repertoires, a Venn diagram was generated based on orthogroups present in four species (GBR-Chromera,, Sydney Chromera, Plasmodium falciparum and Fugacium). The Venn diagram was plotted using the R package VennDiagram (Chen and Boutros, 2011) based on orthogroup information generated with OrthoFinder (see above).
Comparative transcriptomics
Transcriptomes of Sydney Chromera, Fugacium, and the known mutualistic Symbiodiniaceae (Cladocopium and Breviolum) were also mapped against the KEGG database as previously performed for the GBR strain (using the bi-directional best hit (BBH) method). Genes mapped to different KEGG categories were calculated and compared. To better understand relationships between corals and both Chromera and Fugacium, repertoires of genes in categories that are important for symbiosis, such as ABC transporters, as well as those involved in processes such as nitrogen metabolism and stress tolerance, were compared. The overlap amongst these genes was plotted using the R package UpSetR https://github.com/hms-dbmi/UpSetR/ (Conway et al., 2017).
Results and discussion
GBR Chromera transcriptome assembly and annotation
After confirming the identity of Chromera cultures using novel Chromera-specific PCR primers, a transcriptome assembly was generated from 166 million paired-end Illumina reads (∼33 million per library; Table 2). The number of putative genes (39 457, based on the longest transcript isoform per Trinity gene cluster) identified in the GBR Chromera isolate (Table 2) is comparable to those predicted for various Symbiodiniaceae isolates (30 000-49 000) based on transcriptome and genome data (Bayer et al., 2012;Shoguchi et al., 2013;Rosic et al., 2015;Aranda et al., 2016; Liu et al. 2018 and Shoguchi et al., 2018), but is higher than the number predicted from the Chromera genome of the reference strain (26 112 excluding TEs; Woo et al., 2015). As only 19.4% of the GBR Chromera genes had significant BLASTX hits against the Swiss-Prot protein database, the majority of Chromera genes code for unknown functions. This level of novelty is to be expected for organisms such as chromerids that are evolutionarily distant from well-characterized species, and has previously also been observed with Symbiodiniaceae and other dinoflagellates (Lin et al., 2010;Bayer et al., 2012; Stephens et al., 2018). To assess the quality of the de novo assembled transcriptome, reads were mapped to the assembly and an average of 84% of the paired Illumina reads were mapped successfully (Table 1). The assembled transcriptome was judged to be relatively comprehensive on the basis of high percentages of reads mapping.
Summary statistics for the GBR Chromera de novo transcriptome assembly using Trinity and annotation based on Swiss-Prot (SP), Gene Ontology (GO) and KEGG databases
5225 of Swiss-Prot annotated genes (68.35%) were assigned to 38271 GO terms. Biological process (GO-BP) accounted for the majority of GO terms (22 205, 58.02%), followed by cellular component (GO-CC; 11 149, 29.1%) and molecular function (GO-MF; 4 917, 12.8%). Functions involved in cellular process and metabolic process (16% and 14%, respectively) were highly represented amongst GO-BP. In GO-MF, the most represented terms were catalytic activity (46%) followed by binding (36%). In GO-CC, the terms cell (39%) and organelle (33%) were highly represented (Supporting information Figure 2). Most of the KEGG-based annotations (34% of all assignments) were assigned to the metabolic pathway category, followed by the human disease category (18% of all assignments) (Supporting information Figure 3). Moreover, signal transduction and infectious diseases were the most highly represented pathways (Supporting information Figure 4, Table 2).
Several different approaches were used in order to assess the completeness of the assembled transcriptome. The KEGG annotation was searched for essential protein complexes/ pathways and the majority of genes for the pathways were found. Searched complexes included core cellular/ molecular protein complexes and pathways such as Ribosome biogenesis in eukaryotes, Ribosome, RNA polymerase, Spliceosome and Proteasome (Supporting information Table 3 and Figures 5-9). The use of BUSCO analysis resulted in a moderately high recovery (∼60%) of conserved eukaryotic genes and relatively low recovery (∼26-46 %) of conserved alveolate and protist genes, respectively, using default settings (Supporting information Table 4). In addition, 0.1% of the assembled Chromera sequences (contigs) had BLASTN hits to bacterial databases (E-value ≤ 10−10) indicating very low bacterial contamination.
Sequence and functional similarities of the two Chromera strains
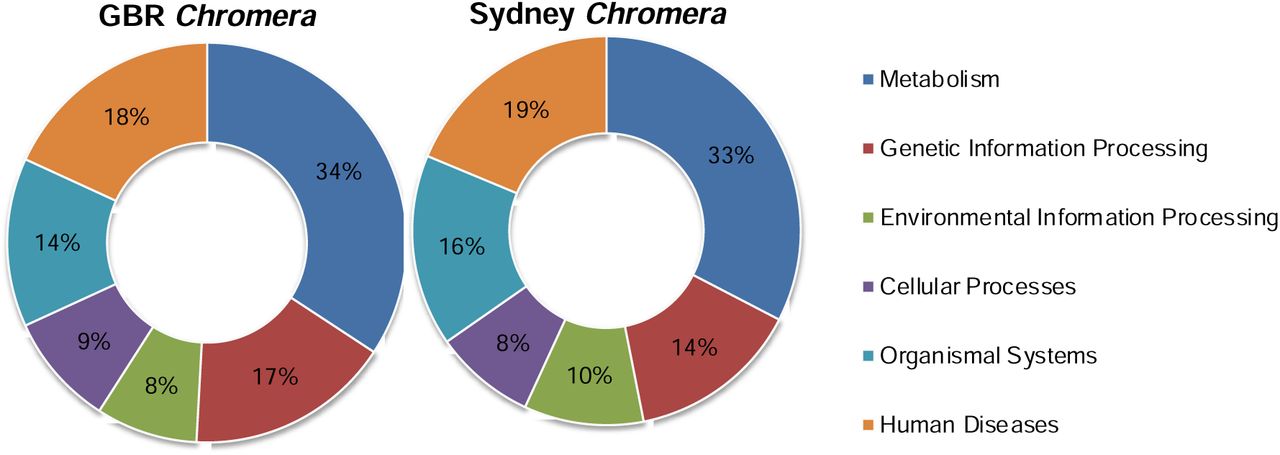
Phylogenomic analyses based on 110 orthologous single copy genes revealed a close relationship (branch length 0.07) between the GBR and Sydney strains of Chromera compared with divergences between different species within the Symbiodiniaceae (branch lengths 1.23 to 2.27) (Fig.1). Pairwise nucleotide similarity between the Sydney and GBR Chromera isolates was 96.2%, whereas the corresponding figures for the genera of Symbiodiniaceae were 72-78%. A close relationship between the Chromera isolates was further supported by the presence of a relatively large number (14645) of shared orthologous genes and an average nucleotide identity of 99.12% based on alignments between these one-to-one orthologs. The overall distributions of the six main KEGG categories were similar in the two Chromera strains, one third of KEGG-annotated genes being assigned to metabolism (Supporting information Figure 10). This might reflect broadly similar functions and lifestyles. The only attempt to understand the nature of the coral-Chromera association used the Sydney strain (Mohamed et al., 2018). Given the high level of similarity between the GBR and Sydney harbour strains, the responses of corals to them are unlikely to differ significantly.

(A) Unrooted phylogenetic tree based on maximum-likelihood phylogenetic analysis of 110 orthologous nucleotide sequences from Sydney Chromera, GBR Chromera, P. falciparum, F. kawagutii, B. minutum and C. goreaui. All nodes in the tree were fully supported (100% of bootstraps) based on 1000 bootstraps by ultrafast bootstrap in IQtree. The sizes of filled circles at some nodes in the tree indicate numbers of shared othologs. (B) Heat map summarizing Hamming distances between taxa based on nucleotide similarity in the 110 orthologous sequences used for phylogeny reconstruction.
Focus on genes implicated in the symbiotic lifestyle
To gain additional perspectives on the coral-Chromera interaction, the assembled transcriptomes for the Chromera strains were compared to those for Fugacium (which is assumed to be non-mutualistic) and to those of Cladocopium and Breviolum, which are mutualistic Symbiodiniacae strains, focusing on categories of genes likely involved in symbiosis, particularly nitrogen metabolism, transport and stress.
Metabolic exchanges in coral-algal symbioses
Nutrient exchange between the symbiotic partners is of major importance in coral-Symbiodinium mutualisms (Meyer and Weis 2012, Davy et al., 2012, Lin et al., 2015, Aranda et al., 2016). However, the nature of the translocated material(s) and mechanisms underlying exchanges between the coral host and algal symbionts are still unclear. To assess the potential for nutrient exchange between Chromera and coral hosts, the representation of the KEGG pathways Nitrogen metabolism and ATP-binding cassette (ABC) transporters were investigated in the two Chromera strains and compared to those in the three members of the Symbiodiniaceae (Fugacium, Cladocopium and Breviolum).
The Nitrogen metabolism category was investigated on the basis that nitrogen cycling or conservation appears to be critical to the coral-Symbiodinium mutualism. Nitrogen is thought to be a growth limiting factor in nutrient-poor tropical waters, and many marine microbes have the ability to assimilate inorganic nitrogen (Pernice et al., 2012), for a recent review see Radecker at al. (2015). The results of a survey of genes captured under the KEGG pathway identifier KO00910 (Nitrogen metabolism) are presented in Supporting information Table 4. Note that this KEGG pathway does not include ammonium transporters, although it has previously been reported that the genomes of Symbiodiniaceae encode multiple ammonium transport proteins (Aranda et al., 2016).
In ocean waters, nitrate is generally the most abundant form of available nitrogen, concentrations often being at least an order of magnitude higher than those of ammonium and nitrite. However, in hospite, ammonium is likely to be the dominant nitrogen source available to intracellular symbionts. Thus, facultative symbionts (as most Symbiodiniaceae are thought to be) must not only be nutritionally versatile, but also able to regulate genes involved in the transport and assimilation of different N-sources. As might be expected, all of the algae surveyed encode nitrate /nitrite transporters as well as enzymes required for assimilation of nitrogen in these forms. Although nitrate/nitrite transporters (NRTs) were present in all of the algae surveyed, some differences were apparent. Whereas the NRTs of Breviolum, Fugacium and Chromera were the MFS-type (K02575), this type was not detected in Cladocopium; rather, in this organism, components (K15577 and K15579) of a distinct ABC-type NRT were found. This difference may be significant; in cyanobacteria, the MFS-type has high affinity for both nitrate and nitrite, whereas the ABC-type has a much higher affinity for nitrate (Maeda and Omata, 2009).
ATP-binding cassette (ABC) transporters
The ABC class includes the largest number of transporters involved in either or both uptake and export of a wide range of substrates, including inorganic ions, carbohydrates and lipids. ABC transporters have been implicated in translocation of nutrients and metabolites in cnidarian symbioses (Davy et al., 2012; Mathews et al., 2017; Mohamed et al., 2019), and hence were an obvious focus for comparative analyses.
As with the nitrogen metabolism category, surveying ABC transporter complements (Fig. 2) was largely unsuccessful in providing general molecular criteria by which known mutualists (Breviolum and Cladocopium) can be distinguished from commensals or parasites. One potentially significant difference, however, is that both Breviolum and Cladocopium encode K10111 members, which are nominal transporters of a variety of different sugars and which may be involved in carbohydrate translocation in hospite, whereas proteins of this type were not detected in either Fugacium or Chromera (Supporting information Table 5). Unfortunately, the survey provides few other grounds for speculation about photosynthate translocation. Whilst a number of candidates for roles in sterol or lipid translocation were detected, these were generally not restricted to the coral mutualists. For example, components of the mla/lin type transport system for phospholipids/sterols/gamma-HCH (K02065 and K02066) were detected in Fugacium as well as Cladocopium, and the Chromera ABC repertoire does include possible sterol transporters (K05683, K05681, K08712).

UpsetR plot (which allows comparative visualisation of the presence/absence of genes of a pathway in different taxa) illustrating the intersection between genes of two KEGG pathways: (A) Nitrogen metabolism (KO00910) and (B) ABC transporters (KO02010) in the two Chromera strains, Fugacium, mutualistic Symbiodinianceae (Cladocopium and Breviolum) and the parasite P. falciparum. Each plot is generated from a binary matrix with unique KEGG identifiers for each pathway and each taxon in a column. The number of genes per taxon is represented by the length of the orange bars. The black dots represent presence/absence. The purple columns show the number of genes shared amongst taxa or exclusive to one of them. So, reading the upper plot from the left there are 8 different genes (each gene is represented once) that are exclusively found in Cladocopium and not the others, second column shows 3 genes that are found in the 3 symbiodiniaceae and the 2 Chromera, but not present in P. falciparum, the third column shows 3 genes that are found in all taxa considered, etc. The intersection size reflects the number of unique or shared genes.
Despite these analyses being uninformative on the issue of general characteristics of mutualists, they do provide grounds for speculation about the near-ubiquitous association of Cladocopium with Indo-Pacific corals (LaJeunesse et al., 2018) and particularly with Acropora spp.. The ABC repertoire of Cladocopium by far exceeded those of all of the other organisms surveyed (Fig. 2), and included many genes identified only in this organism, amongst which were transporters for sugars, lipids and amino acids as well as inorganic nutrients. A range of candidate amino acid transporters were restricted to Cladocopium, one of which (HisP) is of particular interest in the context of coral symbioses, as it encodes a histidine transport ATP-binding protein (Supporting information Table 6). The Cladocopium strain included here was originally isolated from Acropora tenuis (Liu et al., 2018), a member of the complex coral superfamily, and whereas robust corals (members of the other coral superfamily) have a complete histidine biosynthetic pathway, complex corals (like all other animals) must acquire this either from the diet or from their resident endosymbionts (Ying et al., 2018). Also intriguing in the context of mutualism is the detection only in Cladocopium of three components required for cystine transport (K02424, K10009, K10010); the interest in these stems from the fact that, amongst corals, members of the genus Acropora lack one of the enzymes required for cysteine biosynthesis (Shinzato et al., 2011). In this respect, Acropora appears to be unique; all other corals surveyed had a compete cysteine pathway. The presence of an efficient cystine transport system may therefore underlie the near ubiquitous association of Cladocopium C1 with Acropora spp.
Fugacium – an evolutionary recidivist?
The ABC transporters identified in Fugacium include several candidates for roles in nutrient exchange – for example, K05641 encodes an ABCA1 protein known as the cholesterol efflux regulatory protein (CERP) that mediates efflux of cellular cholesterol and phospholipids (reviewed by Zhao and Lappalainen, 2012). Note that an ABCA1 gene was up-regulated in adult Acropora millepora colonies in the light, i.e. when transport of photosynthate from alga to host occurs (Bertucci et al.,2015). K02065, which was present in all three Symbiodiniaceae, encodes a phospholipid/cholesterol/gamma-HCH transport system ATP-binding protein, again with a potential function in lipid/sterol translocation. Indeed, some ABC family A transporters (ABCA7 and ABCA3) were upregulated in Cladocopium during establishment of symbiosis with coral larvae (Mohamed et al., 2019). Thus, although Fugacium is now considered not to be a coral mutualist, the comparative analyses presented here do not rule this out. Whilst the nature of the relationship between Fugacium and corals is still unclear, the apparent contradictions could, however, be rationalized by consideration of the phylogenetic position of Fugacium. The ability to become endosymbiotic is considered to be a defining characteristic of the family Symbiodiniaceae (LaJeunesse et al., 2018); as a member of a derived clade within the Symbiodiniaceae, Fugacium may have undergone (or be undergoing) secondary loss of symbiotic potential but its genome still bear vestiges of this ancestral condition.
The small heat shock protein HSP20 might explain stress tolerance in Chromera
Molecular chaperones are important for refolding damaged proteins (Vierling, 1991), thus their involvement in stress tolerance is inevitable. Amongst these chaperones we looked at heat shock proteins in both Chromera and two symbiotic members of Symbiodiniaceae, Cladocopium and Breviolum. Despite genes coding for chaperones in the KEGG Orthology 03110 Chaperones and folding catalysts (including HSP90/70) being present in similar numbers in all of these genera, gene(s) coding for two HSP20 isoforms were found in both Chromera strains to the exclusion of the three Symbiodiniaceae algae (Fig. 3). Consistent with a potential role in stress tolerance the HSP20 was reported recently to be correlated with stress tolerance in reef-building corals where coral species containing more HSP20 genes were more stress tolerant (robust corals) than those with smaller numbers of HSP20 genes (complex corals) (Ying et al., 2018).

Comparison of the repertoires of genes encoding chaperones and folding catalysts (KO03110) of Chromera strains and Symbiodiniaceae. (A) A five-way Venn diagram of the gene overlap amongst the two Chromera strains, mutualistic Symbiodiniaceae (Cladocopium and Breviolum) and Fugacium. (B) A bar graph illustrating numbers of HSP20, HSP70 and HSP90 genes identified in the five algal datasets.
Conclusions
This paper describes de novo assembly and annotation of a transcriptome for a Chromera strain isolated from a GBR coral, which was generated in order to understand diversity within the species and to provide a better understanding of the metabolic capabilities and lifestyle of this photosynthetic apicomplexan alga. The GBR strain has a high degree of similarity with the Sydney (culture collection) strain, hence infection studies based on the former (e.g. Mohamed et al., 2018) can be generalized to the latter. Comparisons between Chromera, the Symbiodiniaceae algae Fugacium, Cladocopium and Breviolum and the apicomplexan parasite P. falciparum based on specific categories of genes were inconclusive with respect to common molecular characteristics of mutualists, but suggest that HSP20 genes may underlie the higher thermal tolerance of Chromera compared to Symbiodiniaceae. The presence of specific genes implicated in mutualism suggest that Fugacium may have secondarily lost the ability to establish symbioses, and the comparative analyses provide a molecular rationale for the near-ubiquitous association of Cladocopium with Indo-Pacific corals.
Conflict of Interest
The authors declare that they have no conflict of interest.
DATA AVAILABILITY
Data reported in this study have been submitted to the NCBI Gene Expression Omnibus (GEO) under accession number GSE139820 including raw illumina data, transcriptome assembly and annotations.
Author contributions
AM and DM conceived and designed the study. AM conducted the experiments and generated the RNA-Seq data. AM performed bioinformatics required for transcriptome assembly, annotation and comparative analyses with guidance from CXC and MR. JZ and IC performed the phylogenomic analysis. AM, EB and DM interpreted that data and wrote the manuscript. All authors read the article and approved the final version.
Supplementary Methods
Identity check of Chromera cultures
Microscopic check of Chromera culture
The starting Chromera culture was checked with an inverted microscope for protist and bacterial contamination before small aliquots were subjected to genetic identification, growth and further application of the experimental treatments.
Genetic Identification
Chromera gDNA extraction
gDNA was extracted from 50 ml of exponentially growing culture. Cultures were centrifuged at 9000 rpm for 5 minutes at 4 °C, the Chromera pellet was resuspended in 1ml fresh f/2 medium, centrifuged at maximum speed for 5 minutes at 4°C and stored at −80 °C until further treatment. The ISOLATE II Plant DNA Kit (BIOLINE) was used for DNA extraction according to the manufacturer’s instructions. DNA was eluted in 100 μl of elution buffer in a 1.5 ml tube. DNA was checked by running onto an agarose gel and a Nanodrop ® ND-100 Spectrophotometer (Wilmington, U.S.A) was used to estimate the concentration and quality of the DNA obtained from the DNA extractions. Milli-Q water was used to blank the instrument. 1.5 μl of sample was placed directly onto a fibre optic measurement surface where a retention system using surface tension held the sample in place. DNA concentrations, absorbance at 230 (λ230) and the ratio 260/280 were recorded.
Polymerase chain reaction (PCR) Primer Design
Chromera large subunit ribosomal RNA gene, partial sequence, GenBank: EU106870.1, (Moore et al., 2008) and C. velia clone JS497 18S ribosomal RNA gene, partial sequence; internal transcribed spacer 1, 5.8S ribosomal RNA gene, and internal transcribed spacer 2, complete sequence; and 28S ribosomal RNA gene, partial sequence, GenBank: JN935835.1 (Morin-Adeline et al., 2012) were retrieved from GenBank and used as templates for designing the PCR primers. The following primers (Table 1) were used to check the identity of the starting culture.
Amplification of Chromera ribosomal genes using PCR
Amplification of Chromera ribosomal genes was undertaken using specific primers (above) to obtain a PCR product ranging between 416 to 778 bp in size. PCR reaction was conducted in 50 μl using 1 μl of Chromera gDNA (approx. 100ng of DNA) as template. 1 μl of GoTaq® DNA polymerase and 2X GoTaq® reaction buffer and, 5 μl of each primer were used and finally sterile MQW was added to the reaction mixture to make a total volume of 50 μl. The PCR profile was one cycle for 2 min at 94 °C for initial denaturation followed by 34 cycles of 30 sec at 94 °C, annealing for 30 sec at 47 °C/ 51 °C and extension for 2 min at 72 °C. The final extension was at 72 °C for 10 min. The obtained amplicons were run on 1.5% Agarose gel and visualized using a UV trans-illuminator.
Chromera culture and culturing conditions
Cultures growing in the mid exponential (log) phase (+11 days after inoculation) were harvested at the middle of the cultures’ daytime phase and labeled as “control”. In order to maximize the variety of expressed genes, the cultures were subjected to a variety of treatments before RNA isolation and preparation of cDNA libraries. Cultures were subjected to dark stress (24 hour dark period), cold shock (4°C for 4 hours) and heat shock (36°C for 4 hours). Cultures growing in the control conditions +8 days after inoculation cultures were harvested at the middle of the cultures’ daytime phase and labeled as “motile” as cultures showed both Chromera life forms. In addition, cultures were also grown in f/2 media autotrophically while supplemented with exogenous organic compounds at final concentration of total 0.1%(w/v) including; Galactose (D+)
(D00201; Sigma-Aldrich), sodium acetate (D00385; ICN Biomedicals) and Glycerol (D00217; Sigma-Aldrich) and labeled as “mixotrophic”. In all cases, exponentially growing cultures were separated and subjected to the treatment condition and harvested at the end of the experimental treatment. During culturing no antibiotics were used to exclude any potential contribution of the antibiotic treatment to the mRNA expression in the cultures.
Illumina data quality check
Illumina raw reads from each paired end file were visualized using FASTQC version 0.11.2 in order to determine the quality of the data. In addition, reads were inspected for adapter contamination by searching for the IIlumina universal and indexed adapters.
Supplementary Figures

Amplification of Chromera ribosomal genes using newly-designed Chromera-specific PCR primers. M refers to the marker or DNA ladder. A refers to positive control reaction (Symbiodinium gDNA and Symbiodinium-specific primers), while B refers to negative control reaction (MQ water as a template). C, D, E and F are Chromera gDNA tested with the four primer pairs at two different annealing temperatures of 47°C and 51°C.

Gene Ontology (GO) assignment (2nd level GO terms) of the GBR Chromera transcriptome. Biological processes (A) constituted that majority of GO assignment of contigs (22,205 counts, 58.02%), followed by cellular components (C) (11,149 counts, 29.1%) and molecular function (B) (4,917 counts, 12.8%).

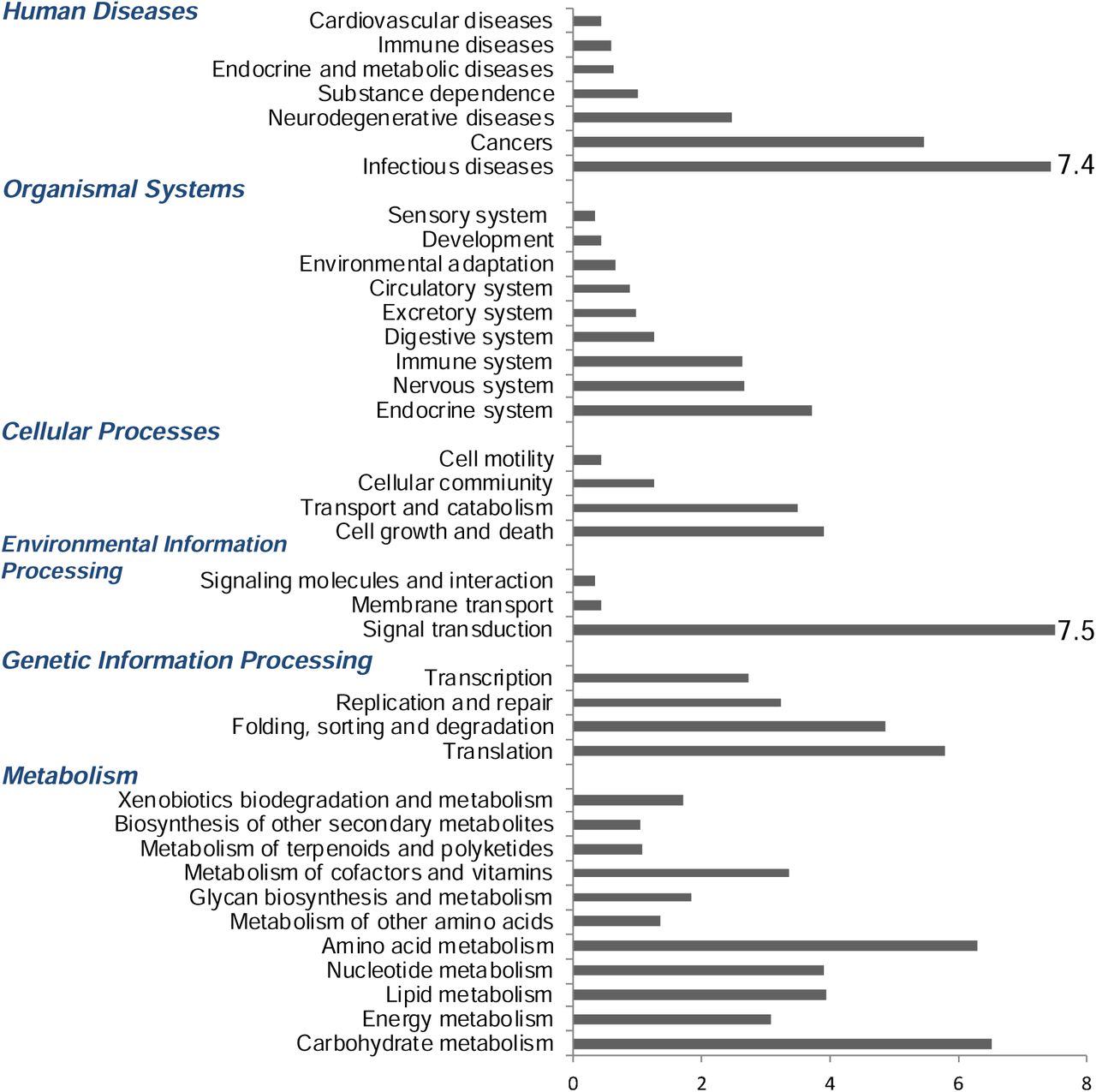
Main KEGG pathway category representation and percentages in the case of the GBR Chromera strain. Numbers above the bars give percent of annotated sequences in each category.

Distribution of KEGG pathways in transcriptome of the GBR Chromera strain. The chart shows the percentage of sequences assigned to each category.

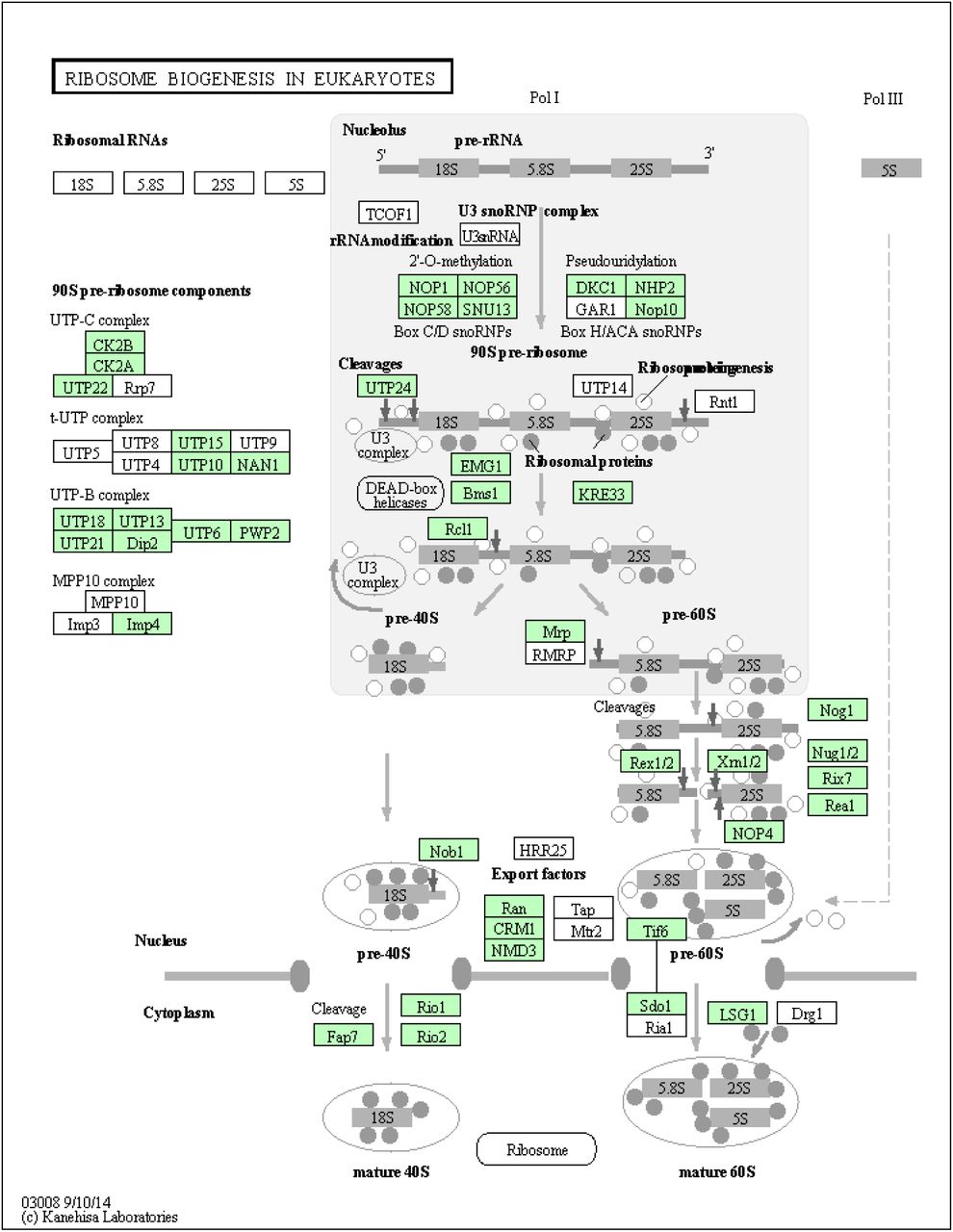
Ribosome biogenesis (eukaryotes) pathway (KO03008) identified in the Chromera transcriptome. KEGG pathways analysis shows Chromera orthologs involved in ribosome biogenesis (highlighted in green).

Ribosome pathway (KO03010) identified in the Chromera transcriptome. KEGG pathways analysis shows Chromera orthologs involved in ribosome (highlighted in green).
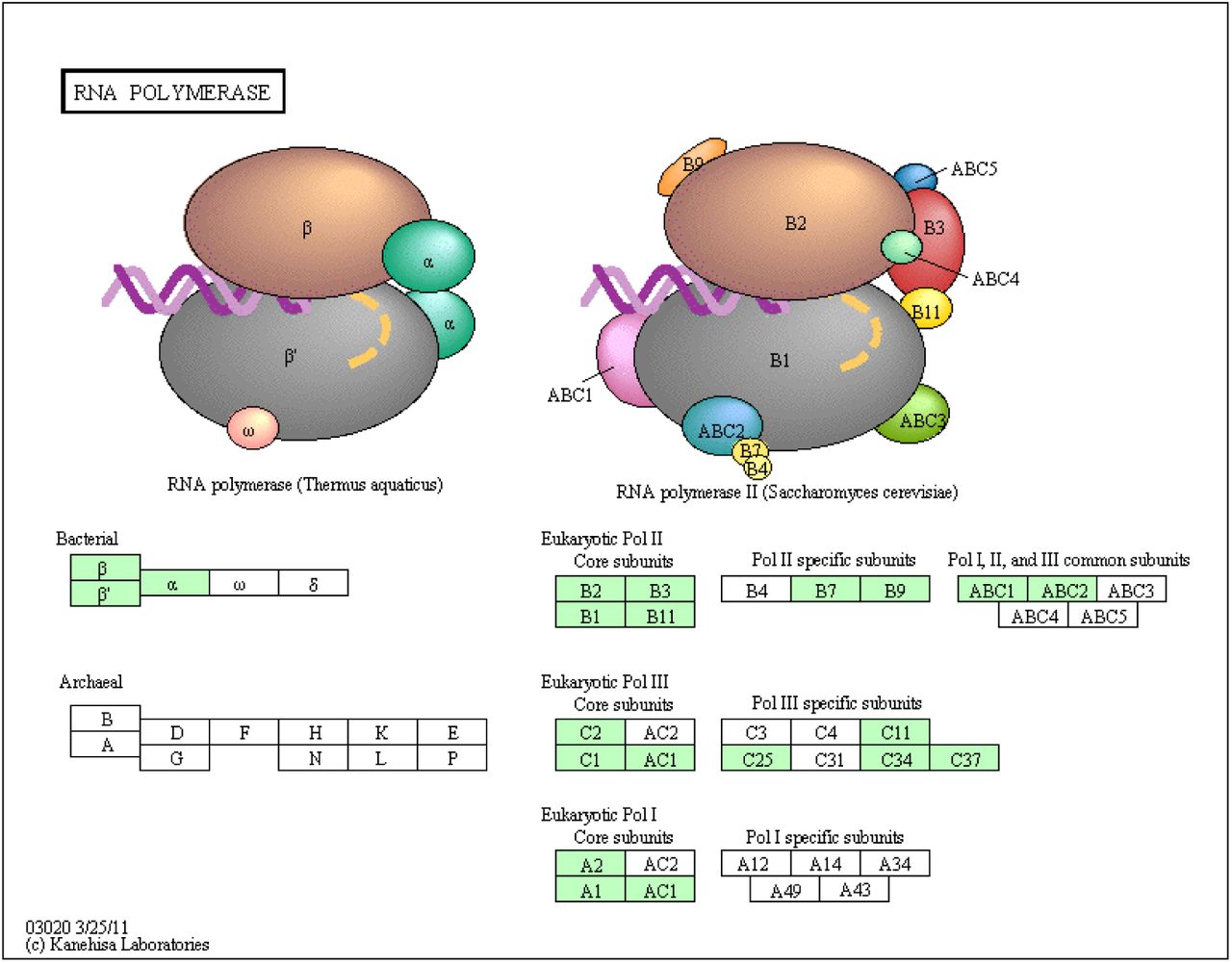
RNA polymerase pathway (KO03020) identified in the Chromera transcriptome. KEGG pathways analysis shows Chromera orthologs involved in RNA polymerase (highlighted in green).

Spliceosome pathway (KO03040) identified in the Chromera transcriptome. KEGG pathways analysis shows Chromera orthologs involved in spliceosome (highlighted in green).
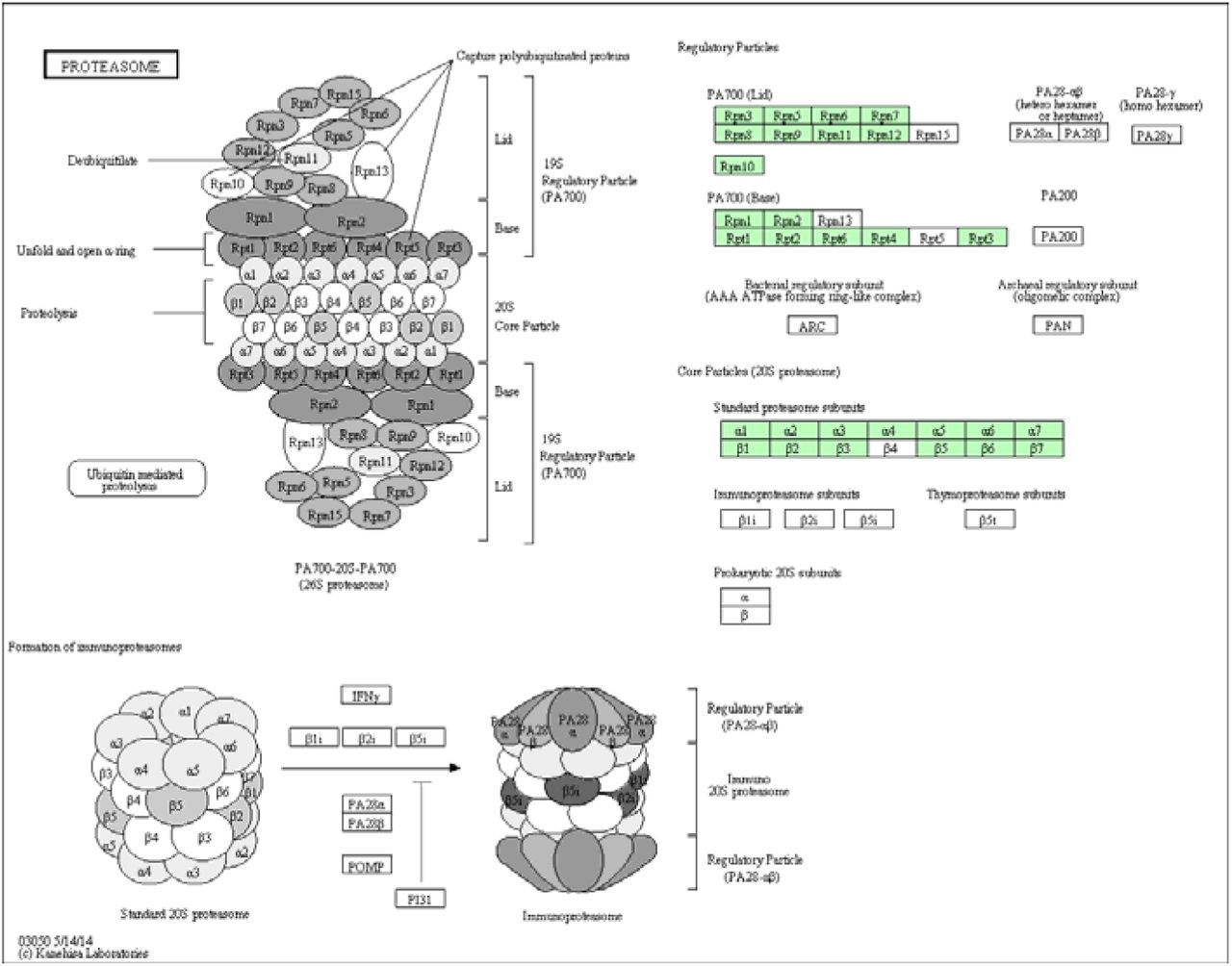
Proteasome pathway (hsa03050) identified in the Chromera transcriptome. KEGG pathways analysis shows Chromera orthologs involved in proteasome (highlighted in green).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Overall distribution of the main KEGG categories in GBR and Sydney Chromera. The doughnut charts show the percentages of the sequences assigned to the six KEGG categories.
List of Chromera velia specific PCR primers used for verifying the identity of the cultures. Primers were designed using the NCBI Primer-Blast tool (http://www.ncbi.nlm.nih.gov/tools/primer-blast/)
Summary of KEGG orthology data for the GBR Chromera strain
Selected KEGG pathways/protein complexes identified in the GBR Chromera transcriptome
Recovery of BUSCO genes in de novo transcriptome of GBR Chromera
Table 5. Genes mapped to the KEGG pathway Nitrogen metabolism “KO00910” in in GBR-Chromera, Sydney Chromera, Cladocopium, Breviolum and P. falciparum. Provided as excel file
Table 6. Genes mapped to the KEGG pathway ABC transporters “KO02010” in GBR-Chromera, Sydney Chromera, Cladocopium, Breviolum and P. falciparum. Provided as excel file
Acknowledgments
The research was supported by the Australian Research Council through Grant CE140100020 to DM via the ARC Centre of Excellence for Coral Reef Studies at James Cook University. AM was supported by PhD scholarships provided by the Cultural Affairs and Mission sectors of the Egyptian Ministry of Higher Education and Scientific Research and James Cook University Postgraduate Research Scholarship (JCUPRS) and through the AIMS@JCU joint-scheme. The authors gratefully thank Dr Vivian Cumbo at Macquarie University for supplying Chromera cultures.
References




