

Abstract
Individuals with congenital amusia have a lifelong history of unreliable pitch processing. Accordingly, they downweight pitch cues during speech perception (even large, obvious ones), and instead rely on other dimensions like duration. We investigated the neural basis for this strategy. Using fMRI, individuals with amusia and controls (N=30) were scanned while they used pitch and duration cues to match auditory and visual sentences. A data-driven analysis procedure identified four ‘seed’ regions showing large Control > Amusic functional connectivity differences in lateral prefrontal cortex, which were examined with respect to the rest of the brain. Prominent decreases in functional connectivity were detected in the amusia group, between left prefrontal language-related regions (inferior and middle frontal gyrus/DLPFC) and right hemisphere pitch-related regions (auditory and anterior insular cortex). Our results suggest that individuals compensate for differences in the reliability of perceptual dimensions by regulating functional connectivity between task-relevant frontal and perceptual regions.
Introduction
Congenital amusia is a rare condition characterized by impaired perception of and memory for pitch (Peretz et al., 2002). Although amusia presents as an auditory condition, auditory cortical responses (Moreau et al., 2013; Norman-Haignere et al., 2016) are normal, as is subcortical encoding of pitch (Liu, Maggu, et al., 2015). The dominant view of amusia’s neural basis is that connectivity between right inferior frontal cortex and right auditory cortex is impaired, resulting in impaired conscious access to pitch information for guiding behavior (Hyde et al., 2011; Albouy et al, 2013; Leveque et al., 2016; Zendel, et al., 2015; see Peretz, 2016 for review). While congenital amusia is believed to be innate, there is evidence that recovery is possible through training (Whiteford & Oxenham, 2018).
Although pitch is usually associated with music, it is also important for cueing categories in spoken language (de Pijper & Sanderman, 1994; Streeter, 1978) and conveying emotion in speech (Frick et al., 1985). In highly-controlled laboratory tasks in which speech perception judgments must be made based on pitch alone, only minor deficits have been observed in amusia (Liu, Jiang, et al., 2015; Patel et al., 2008). In naturalistic speech perception contexts, people with amusia rarely report any difficulties (Liu et al., 2010). This may be because, in natural speech, pitch variation tends to co-occur with variation in other acoustic dimensions, such as duration and amplitude. Our lab has shown that in such cases where multiple redundant cues are available, English-speaking individuals with amusia tend to rely less on pitch than non-amusic controls. This suggests that they can calibrate their perception by down-weighting the cues that are less reliable for them (Jasmin et al., 2019).
It is unknown how decreased reliance on a particular acoustic cue during speech perception (such as pitch cues in amusia) is reflected in the brain. Previous neural studies of cue integration have focused on integration of multiple modalities, e.g. the “weighted connections” model of multisensory integration. In this model, the relative reliability of the modalities involved with perception of a stimulus is related to differential connectivity strength (Beauchamp et al., 2010; Rohe and Noppeney, 2018). For example, when participants simultaneously view and feel touches to the hand and reliability of visual and tactile perception is manipulated experimentally via introduction of noise, connection strength (effective connectivity measured with functional MRI and structural equation modeling) between unimodal and multimodal sensory areas adjusts accordingly. More concretely, when visual information is degraded, the connection strength between lateral occipital cortex (a visual area) and intraparietal sulcus (a multimodal area) decreases, and when tactile perception is made noisier, connection strength between secondary somatosensory cortex and intraparietal sulcus becomes weaker (Beauchamp et al., 2010). Similarly, effective connectivity between the (multimodal) superior temporal sulcus (STS) and visual and auditory areas has shown similar modulations during processing of audiovisual speech: connection strength between auditory cortex and the STS is weaker when noise has been introduced to the auditory speech, and conversely connection strength between visual cortex and STS is weaker if visual noise is introduced (Nath and Beauchamp, 2011).
Just as connectivity differences have been shown to reflect the precision of different dimensions during multisensory integration, an analogous phenomenon may be at work within a single modality during multidimensional integration. As mentioned, the acoustic speech signal carries multiple co-occurring acoustic dimensions (e.g. roughly described as pitch, duration, and amplitude), which often provide redundant cues to disambiguate a linguistic category (Patel, 2014; Winter, 2014; Jasmin et al., 2019a). Individuals with typical pitch perception have learned through a lifetime of experience with speech acoustics that vocal pitch is a useful and reliable cue. By contrast, individuals with amusia, who have unreliable perception of and memory for pitch (analogous to the ‘noise’ introduced in the multisensory integration studies cited above), would have learned that, for them, pitch is not a reliable cue for processing spoken language. Thus, by analogy to the multisensory weighting results described above, we hypothesize that amusics may exhibit decreased connectivity between language regions and pitch-related areas during speech processing.
The neural foundations of perceptual weighting in speech have thus far not been investigated in atypical individuals. Indeed, only one previous functional neuroimaging study has examined the neural processing of spoken material in people with amusia. In this study no group differences were detected in task-related activation or functional connectivity during processing of speech (whereas group differences were observed during processing of tones; Albouy et al., 2018). However, this study used tasks for which pitch processing would not have been necessary (verbal memory), as well as a priori regions of interest derived from studies that used tonal/melodic (rather than linguistic) stimuli. It therefore may not be surprising that neural differences between amusics and controls did not emerge. It remains an open question how functional connectivity in amusic and non-amusic participants may differ during pitch-related language tasks with regions of interest selected with a whole-brain data-driven approach. Although we aim to focus on amusia here, our results may illuminate the neural basis of dimensional weighting in speech perception more generally.
As discussed, the relative reliability of senses in multisensory perception is reflected in neural connection strength—is reliability of dimensions within a sense reflected similarly? Given that amusics have unreliable perception of and memory for pitch and have low pitch cue weights during language processing (Jasmin et al., 2019a), do they also exhibit correspondingly decreased functional connectivity between regions typically found to be involved in pitch processing and frontal regions associated with speech and language? Here we set out to answer these questions. We used functional magnetic resonance imaging to scan 15 individuals with amusia and 15 controls. Participants matched spoken sentences with visually-presented ones on the basis of the position of intonational phrase boundaries. These intonation changes were conveyed differently, in three conditions: pitch cues only, duration cues only, or both these cue types together (Jasmin et al., 2019a, b). Functional connectivity was then examined using a data-driven approach that allowed us to identify the largest group differences, without the need for regions of interest to be selected a priori. The benefit of this approach is that any set of regions could emerge, not only ones reported in previous literature. Crucially, task performance was matched between the groups (based on prior behavioural testing (Jasmin et al., 2019a), ensuring that any neural differences did not simply represent an inability to perform the task.
Results
In-scanner Behavior
On each trial, participants read one visually presented text sentence, then heard two auditory versions of the sentence, only one of which contained an acoustically-conveyed phrase boundary in the same place as in the text sentence. Trials were scored as correct if a participant pressed the button associated with the auditory sentence that correctly matched the text sentence. Proportions of correct judgments were subjected to a repeated-measures Analysis of Variance. Overall, proportion correct across amusia and control groups was matched (main effect of Group, F(1,84) = 0.16, p = 0.69), interaction of Group by Condition (F(2,84) = 0.374, p = 0.96). This lack of interaction was predicted based on previous results obtained from a similar paradigm using out-of-scanner data but from the same participants (Jasmin et al., 2019a). There was a main effect of condition (F(2,84) = 3.32, p = 0.04. Follow-up post-hoc testing indicated that performance in the Combined condition (with pitch and duration cues simultaneously present) was more accurate than either Pitch-Alone (T(84) = 2.3, p = 0.02) or Time-Alone (T(84) = 2.1, p = 0.03), a result that was also predicted and which replicates the behavioral findings in Jasmin et al. (2019a).
Neuroimaging - whole-brain connectedness
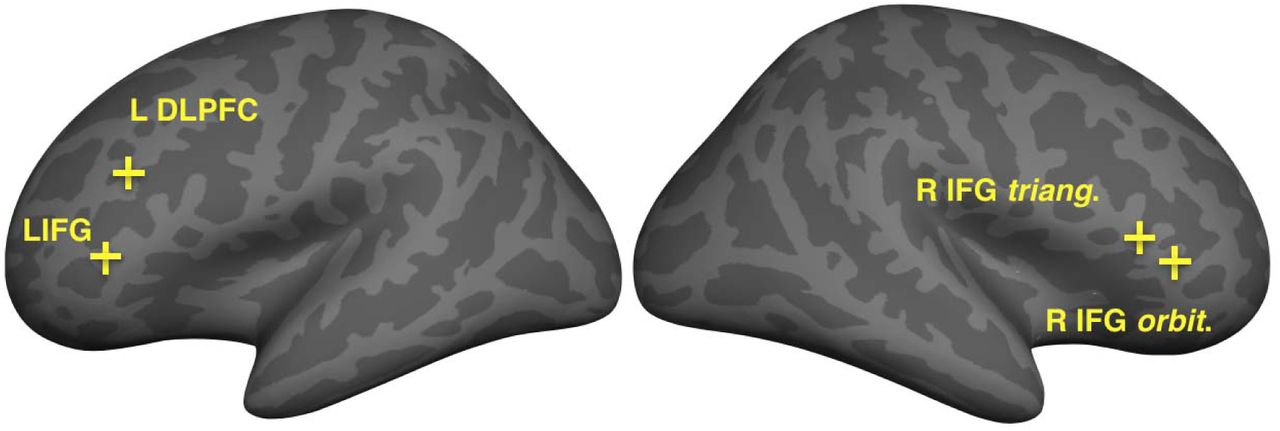
As discussed in the Methods, a data-driven approach was taken to identify brain regions with the largest group- and condition-related differences in functional connectivity. Comparing whole-brain connectedness values by group (Amusia vs. Controls) revealed four significant locations (where z of peak vertices > 4.61, FDR-corrected 0.05) that showed greater whole-brain connectedness for the control than amusia group (see Fig. 1). All group differences were located in the inferior frontal cortex: two left hemisphere vertices (inferior frontal gyrus p. triangularis, and dorsolateral prefrontal cortex); and two right hemisphere vertices (inferior frontal gyrus p. triangularis and p orbitalis). There were no areas where whole-brain connectedness differed by Condition, or showed an interaction of Group and Condition.

Inflated surfaces show the locations of False Discovery Rate-corrected group differences (Control > Amusia) in whole-brain connectivity, which were used as seeds in subsequent analyses. All four seed vertices were located in inferior frontal cortices (left inferior frontal gyrus; left DLPFC, right inferior frontal gyrus pars triangularis. and right inferior frontal gyrus orbitalis)
Follow-up seed-to-whole brain tests
Follow-up testing was conducted on the four significant regions (Control > Amusia, collapsed across the three conditions) identified above to characterize the specific cortical regions driving these group connectivity differences (Berman et al., 2016; Gotts et al., 2012; Jasmin et al., 2018; Song et al., 2015). Relative to control participants, amusic participants’ left inferior frontal gyrus seed region showed particularly notable decreases in connectivity with the right posterior superior temporal and inferior parietal cortex, as well as with the right posterior superior temporal sulcus (Fig. 2). Analysis of subcortical connectivity indicated that there was also weaker connectivity with the right nucleus accumbens.

Significant group differences (control > amusia) in functional connectivity with left hemisphere seeds, False Discovery Rate corrected, minimum Z=3.57). Warm colors indicate greater connectivity in the control than amusia participants. The largest decreases in connectivity in the amusia group were located in right superior temporal plane and gyrus, the posterior middle temporal gyrus onto the inferior bank of the superior temporal sulcus, and anterior insula.
The left dorsolateral prefrontal cortex in amusic participants showed decreased functional connectivity with the mid portions of the right superior temporal gyrus, posterior part of the right middle temporal gyrus extending into the inferior bank of the superior temporal sulcus, and the right anterior insula (Fig. 2). Several subcortical structures also showed significantly reduced (FDR-corrected) connectivity with the seed in amusics: bilateral caudate nucleus and putamen, bilateral pallidum, bilateral cerebellum, and bilateral thalamus.
The right pars triangularis seed showed Control > Amusic connectivity with right dorsolateral prefrontal cortex and left posterior superior temporal gyrus (Fig. 4). It also showed decreased connectivity with left nucleus accumbens. Right pars orbitalis showed decreased connectivity with right dorsolateral prefrontal cortex (Fig. 3). There was also decreased connectivity with the left thalamus.

Significant group differences (control vs amusia) in functional connectivity with right hemisphere seeds, False Discovery Rate corrected, minimum Z=3.57). Warm colors indicate greater connectivity in the control than amusia participants. Prominent decreases in connectivity with the right inferior frontal gyrus in individuals with amusia included superior temporal plane, and areas of occipital, frontal, and parietal cortex.

(A) Example spectrograms of the early closure (top) and late closure (bottom) stimuli for the Combined condition. Fundamental frequency contours are indicated with blue lines. The relative duration of the critical words are indicated with orange and green boxes. To the right, syntactic trees for the two sentences are shown to highlight the grammatical structure indicated by the phrase boundaries. (B) The time course of a single trial. Participants read a text version of the sentence from the screen, which was either early or late closure. This was followed by auditory presentation of the late and early closure versions. After both recordings were played, participants chose whether the first or second recording they heard matched the visually presentence sentence better. A single whole-brain volume was acquired after the button press, timed to capture the peak of the hemodynamic response.
Activation Results
Although we were concerned with functional connectivity rather than activation, we also tested for differences in activation levels between groups and conditions. False Discovery Rate correction was used to correct for multiple comparisons across both hemispheres for each test (Group, Condition and Group X Condition). No significant differences were detected for the main effects of group and condition, nor the interaction of those factors. Results images from these analyses are available online (see Data Availability Statement for details).
Discussion
We found that individuals with amusia, who have been previously shown to rely less on pitch than controls to process spoken language (Jasmin et al. 2019a), exhibit decreased functional connectivity between left frontal areas and right hemisphere pitch-related regions. In our task, participants matched spoken sentences with visually presented sentences based on pitch, duration, or both these acoustic dimensions together. Using a data-driven approach, we identified four regions in left and right inferior frontal cortex for which the amusic group exhibited decreased functional connectivity with several other sites in frontal, temporal and occipital cortex. The most prominent of these results was decreased connectivity between left frontal regions classically implicated in language processing (left IFG and DLPFC) and right hemisphere regions —in the superior temporal gyrus and sulcus, Heschl’s gyrus, and anterior insula—that have been implicated in pitch processing (Lee et al., 2011; Garcea et al., 2017; Warren et al., 2003). We suggest that this decreased connectivity between right hemisphere pitch and left hemisphere frontal cortices may relate to the unreliability of the amusics’ perception of and memory for pitch. This is similar to the “weighted connections” model of multisensory integration, where a more (or less) reliable modality is given a stronger (or weaker) weight. (Beauchamp et al., 2010).
Congenital amusia is often described as a disorder related to structural and functional connectivity within the right hemisphere, particularly between right inferior frontal cortices and right posterior temporal cortex (see Peretz, 2016 for review). Consistent with this proposal, we found in the present study that right inferior frontal cortex exhibited strongly decreased functional connectivity in the amusia group, and follow-up seed testing revealed that right auditory areas were involved as well. However, we also found that sites in left frontal cortex also showed large decreases in connectivity in amusia, also most prominently with right hemisphere auditory areas. Our results are consistent with an account that right hemisphere auditory areas are not only abnormally connected to right frontal areas (as observed during tonal tasks), but are less integrated with frontal left hemisphere regions when processing speech and language.
Our null results for group differences in activation are consistent with prior reports that amusics and controls do not differ in pitch representations within sensory regions. For example, the extent of pitch-responsive regions within auditory cortex has been shown to be similar in participants with amusia and controls (Norman-Haignere et al. 2016). Brainstem encoding of pitch in speech and musical stimuli is similarly unimpaired in individuals with amusia (Liu et al., 2015). Moreover, in oddball EEG paradigms, amusics show similar pre-attentive mismatch negativity responses to small pitch deviants, but impaired attention-dependent P300 responses (Moreau et al. 2009; Peretz et al. 2009; Goulet et al. 2012; Moreau et al. 2013). These findings, along with the fact that amusics show intact non-volitional behavioral responses (unconscious pitch shifts) when presented with pitch-altered feedback of their own voice (Hutchins and Peretz 2013), have been interpreted as evidence that amusia is a disorder of pitch awareness rather than one of low-level pitch processing (Peretz et al. 2009), with differences in structural connectivity as one possible foundation of this putative impaired pitch awareness (Hyde et al. 2006; Loui et al. 2009; but see Chen et al. 2015).
Our interpretation of differences in functional connectivity between amusics and controls diverges somewhat from these previous approaches: we argue that down-weighting of pitch information during perceptual categorization in both speech and music is adaptive, inasmuch as amusics have learned that pitch is an unreliable source of evidence relative to other perceptual dimensions. The evidence above suggesting that encoding of pitch in the brainstem and auditory cortex and pre-attentive responses to pitch changes are unaffected in amusia can be interpreted as suggesting that the fundamental deficit in amusia may not be increased perceptual noise or decreased pitch awareness but difficulties with retention of pitch information in memory. This interpretation is consistent with evidence suggesting that amusics have difficulty with pitch sequence processing tasks even when discrimination thresholds are accounted for (Tillmann et al. 2009), as well as the finding that delaying the time interval between standard and comparison tones exacerbates pitch discrimination impairment in individuals with amusia (Williamson et al. 2010). Moreover, the pitch awareness account of amusia cannot explain the Jasmin et al. (2019a) finding that pitch cues are downweighted only during longer-scale suprasegmental speech perception, while pitch weighting is not different between amusics and controls during shorter-scale segmental speech perception, despite pitch cues being arguably more subtle in the segmental condition. However, this finding can be explained by the pitch memory account, as the suprasegmental task requires detection of and memory for pitch patterns within a complex sequence, while the segmental task does not. Furthermore, an account of amusia which suggests that the disorder primarily stems from differences in structural connectivity cannot account for the recent finding that functional connectivity patterns do not differ between amusics and controls during a verbal memory task (Albouy et al. 2018). We suggest, therefore, that amusics neglect pitch because they have implicitly learned that their memory for pitch is unreliable, and that this down-weighting of pitch is reflected in decreased functional connectivity between right auditory areas and downstream task-relevant areas which integrate information from perceptual regions. One way to test this hypothesis would be to examine functional connectivity during perceptual categorization of consonant-vowel syllables as voiced versus unvoiced based on a pitch cue (F0 of the following vowel) and a durational cue (voice onset time). We predict, based on our previous findings (Jasmin et al. 2019a), that functional connectivity will not differ between amusics and controls on this task, a finding which would not be predicted by the pitch awareness account of amusia.
Several other future directions are suggested by our results, particularly for examining cue weighting during auditory/speech perception. In the multimodal integration studies mentioned above (Beauchamp et al., 2010; Nath and Beauchamp. 2011), reliability of two different sensory modalities was manipulated experimentally, resulting in changes in connectivity. Similarly, aspects of speech could be selectively masked with noise in order to make them less reliable, which in turn could cause corresponding changes in functional or effective connectivity. Indeed, behavioral work has indicated that when fundamental frequency (pitch) or durational aspects of speech are manipulated to be unreliable cues, categorization behavior shifts such that participants place less relative weight on the dimension that has been made less reliable (Holt & Lotto, 2006). Certain groups, such as tone language speakers, are known to have fine-grained pitch perception abilities, and tend to place greater weight on pitch even when processing speech from a second, non-tonal language that they have learned (e.g. English; Yu, et al., 2010; Zhang et al, 2010, Zhang et al., 2008; Qin et al., 2017). Given the increased reliability of their pitch perception, tone language speakers may exhibit correspondingly high connectivity strength between right hemisphere auditory regions and left hemisphere ‘language regions’ when pitch cues are present (more so than native non-tonal language speakers).
Materials and Methods
Participants
Participants, 15 individuals with amusia (10 F, age = 60.2 ± 9.4, range = 43–74) and 15 controls (10 F, age = 61.3 ± 10.4, range = 38–74), were recruited from the UK and were native British English speakers. All participants gave informed consent and ethical approval was obtained from the relevant UCL and Birkbeck ethics committees. Amusia status was obtained using the Montreal Battery for the Evaluation of Amusia (MBEA). Participants with a composite score (summing the Scale, Contour and Interval tests scores) of 65 or less were classified as having amusia (Peretz et al., 2003).
Stimuli
The stimuli were 42 compound sentences that consisted of a pre-posed subordinate clause followed by a main clause (see Fig. 4 for an example, and Jasmin et al., 2019a,b for details). There were two versions of each sentence: (1) an ‘early closure’ version, where the verb of the subordinate clause was used intransitively and the following noun was the subject of a new clause [“After Jane dusts, the dining table [is clean]”]; and (2), ‘late closure’, where the verb was transitive and took the following noun as its object, moving the phrase boundary to a slightly later position in the sentence [“After Jane dusts the dining table, [it is clean]”]. Crucially for the task, the words in both versions of the sentence were identical from the start of the sentence until the end of the second noun (“After Jane dusts the dining table …”).
A native British English speaking male (trained as an actor) recorded early closure and late closure versions of each sentence in a sound-proofed room. The recordings were cropped such that only the portions with the same words remained, and silent pauses after phrase breaks were removed. Synthesized versions of these sentences were created with STRAIGHT voice-morphing software (Kawahara and Irino, 2005). First, the two versions of the sentence were manually time-aligned by marking corresponding ‘anchor points’ in the two recordings. Then, morphed speech was synthesized by varying the degree to which the early closure and late closure recordings contributed duration and pitch information. We synthesized pairs of stimuli in three conditions: (1) In the Pitch condition, the stimulus pair had exactly the same durational properties (that is, the length of phonemes, syllables, and words was the average between the two original recordings) but the vocal pitch indicated early or late closure at a morphing level of 80%; (2) in the Time condition, vocal pitch in the stimulus pair was identical (at 50% between both versions) but the durational characteristics indicated early or late closure at a morphing level of 80%; (3) in the Combined condition, both pitch and time cued early or late closure simultaneously at 80%. The morphed speech varied only in duration and pitch, while all other aspects of the acoustics (such as amplitude and spectral characteristics other than pitch) were the same, held constant at 50% between the two original recordings during morphing. This stimulus set is freely available (Jasmin et al., 2019b).
MRI data collection
Subjects were scanned with a Siemens Avanto 1.5 Tesla magnetic resonance imaging scanner with a 32-channel head coil, with sounds presented via Sensimetrics S14 earbuds, padded around the ear with NoMoCo memory foam cushions. Functional data were collected using a slow event-related design with sparse temporal sampling to allow presentation of auditory stimuli in quiet. We used an echo planar image sequence, with 40 slices, slice time 85 ms, slab tilted to capture the entire cerebrum and dorsal cerebellum, ascending sequential acquisition; 3×3×3 mm voxel size; silent stimulus and response period = 8.7s, volume acquisition time = 3.4 s, total inter-trial interval = 12.1s, flip angle = 90 degrees, bandwidth = 2298 Hz/pixel, echo time (TE) = 50ms. After collecting functional runs, a high-resolution T1-weighted structural scan was collected (MPRAGE, 176 slices, sagittal acquisition, 2x GRAPPA acceleration, 1 mm isotropic voxels, acquisition matrix = 224 × 256).
Procedure (see schematic in Fig. 4)
Each run began with three dummy scans to allow magnetic stabilization. Each trial (repetition time) lasted 12.1 seconds. The start of each trial was triggered by a pulse corresponding to the start of a volume acquisition (which acquired neural data from the previous trial, at a delay). At t=1 s into the trial, the sentence appeared on the screen; before scanning participants were instructed to read each sentence silently to themselves. At t=5 seconds (plus or minus a random 100 ms jitter) participants heard a spoken version of the first part of the sentence. At t = 7.4 seconds (plus or minus 100 ms jitter) the second version was presented. The two spoken versions contained the same words but their pitch and/or timing characteristics cued a phrase boundary that occurred earlier or later in the sentence. Following this, there were approximately 2 seconds of silence during which the participant responded with the button box, before the scanner began acquiring the next volume at t=12 s. Participants performed three blocks of 42 trials (14 each of Pitch, Time, and Combined) with 8 Rest trials interspersed within each block.
MRI pre-processing
Image preprocessing was performed with FreeSurfer 6.0.0 (Fischl, 2012) and AFNI-SUMA 18.1.18 (Cox, 1996). Anatomical images were registered to the third echo planar image of the first run using Freesurfer’s bbregister and processed with FreeSurfer’s automated pipeline for segmenting tissue types, generating cortical surface models, and parcellating subcortical structures. Masks of inferior colliculi were obtained by manually examining individual subjects’ anatomical images and selecting a single EPI voxel located at its centre, bilaterally. Freesurfer cortical surface models were imported to AFNI with the @SUMA_Make_Spec_FS program. Then a standard pre-processing pipeline using AFNI’s afniproc.py program was used: all echo planar image volumes were aligned to the third repetition time of the first run using AFNI’s 3DAllineate, intersected with the cortical surface with SUMA, smoothed along the surface with a 2D 6-mm-FWHM kernel, and converted to a standard mesh (std.141) for group analyses, separately for each hemisphere, where each vertex in the mesh (198812 per hemisphere) is aligned to the ‘same’ location in the cortex across subjects, using curvature-based morphing.
Motion
The magnitude of transient head motion was calculated from the six motion parameters obtained during image realignment and aggregated as a single variable using AFNI’s @1dDiffMag to calculate a Motion Index (Berman et al., 2016; Gotts et al., 2012; Jasmin et al., 2019c). This measure is similar to average Frame Displacement over a scan (Power et al., 2012) and is in units of mm per repetition time. The difference in average motion between the groups was small (amusia group mean motion = .31mm/TR; control group mean = .28mm/TR) and amounted to 32 micrometers (∼1/30th of a millimeter) per TR. The mean and distribution of motion did not differ statistically between groups (two sample t-test P = 0.7, two-tailed).
Beta series analysis of context-modulated functional connectivity
Given the previous reports (described above) of changes in connection strength between unimodal and multimodal areas in response to noise (Beauchamp, et al., 2010; Nath and Beauchamp, 2011), we chose a connectivity-based analysis approach for our study. Beta series correlation (Rissman et al., 2004) is a technique for examining functional connectivity and its modulation by task, using correlations in trial-by-trial responses. It has been shown to be more powerful than alternatives such as generalized psycho-physiological interaction (gPPI) for event-related designs (Cisler et al., 2014). In a beta series analysis, one beta weight is calculated for each trial in the experiment (rather than for each condition). All of the trial-wise betas associated with a given condition are then ordered to form a “beta series”. Finally, using the beta series in the same way as a standard BOLD fMRI time series, functional connectivity (measured as Pearson correlations) is calculated between seed regions of interest and the rest of the brain. Differences in functional connectivity can then be examined by comparing groups, comparing conditions, or examining the interaction of these factors.
Obtaining trial-wise beta weights
Our experiment used a slow event-related design with a long repetition time (12.1s) and sparse temporal sampling (with volume acquisition separated by silent periods). Therefore, the time between acquisitions was long enough for the haemodynamic response to return to baseline, and each echo planar image acquisition corresponded to exactly one trial (Fig. 4). For this reason, we did not convolve the echo planar image time series with a basis function during subject-level statistical analysis (Hall et al., 1999). In the design matrix for obtaining trial-wise betas, 126 column regressors were used (one for each non-rest trial). Each column vector was of length 150 (corresponding to all trials, including rest trials) and had a single “one” in the position where the trial associated with that column occurred, while zeros were located in every other position. Polynomials up to second degree were also included in the model, on a run-wise basis, to remove the mean and any linear or quadratic trends. Fitting the trial regressors on a subject-wise basis resulted in cortical surface models of beta weights for each of the 126 trials, at each vertex on the reduced-vertex icosahedral cortical surface, with beta weights reflecting the neural response associated with that trial. As noted above, trial-wise betas were then serially ordered to form beta series for each of the three experimental conditions (Pitch, Time, and Combined) (Rissman et al., 2004). In total, 90 beta series were created for each voxel, corresponding to each of the three task conditions (Pitch, Time, Combined) from all 30 participants.
Defining seed regions of interest
Beta series analysis requires initial seed voxels, vertices, or regions to be identified, whose trial-to-trial changes in activity are then compared to those of the rest of the brain. Rather than choose a priori seeds derived from the literature, which used mainly musical tasks or resting state, we used a data-driven approach to search for the largest group and condition differences in functional connectivity (Berman et al., 2016; Cole et al., 2010; Gotts et al., 2012; Jasmin et al., 2019c; Meoded et al., 2015; Song et al., 2015; Steel et al., 2016; Stoddard et al., 2016; Watsky et al., 2018). To do this, we first calculated the “whole-brain connectedness” (similar to ‘centrality’ in graph theory) of each cortical vertex (a procedure available in AFNI as the 3dTCorrMap function). The whole-brain connectedness of a given vertex is defined as the Pearson correlation of activity within that vertex/voxel and the average signal across all neural gray matter in the rest of the brain. Mathematically, this is equivalent to calculating thousands of Pearson correlations, of a given vertex/voxel series and every other vertex/voxel series in the brain, and then taking the mean of those correlations (Cole et al., 2010), then repeating the process for every individual voxel/vertex. As such, it represents the global connectedness (or ‘global correlation’) of a vertex/voxel.
To calculate whole-brain connectedness, first, the average of trial-wise betas in gray matter across the brain was calculated in volume space, separately for each subject and for each condition (Pitch, Time, Combined) by running first-level (subject) models. The statistical models were identical to those conducted on the cortical surface, described above, but were performed on volumetric Talairach images instead of the cortical surfaces. The reason for this choice was so that voxels in cortex and subcortex would contribute equally to our measure of global (whole-brain) connectivity. First, average gray-matter beta value was calculated for each trial by intersecting each image in the beta series with a whole-brain gray matter mask (which excluded white matter and ventricles) and calculating the average beta value within the mask (Gotts et al., 2012; Jasmin et al., 2019c). Next, this gray matter average was correlated with each cortical surface vertex’s beta series, separately for each subject and condition, to obtain whole-brain connectedness maps. These values were then subjected to a statistical analysis based on our 2 (Group) × 3 (Condition) experimental design. Linear mixed effects models (AFNI’s 3dLME) (Chen et al., 2013) were constructed whose dependent variables were the vertex-wise whole-brain connectedness maps from each beta series. Group and Condition and their interaction were included as fixed effects. Participant was treated as a random intercept. Results of this step were corrected vertex-wise for multiple comparisons with False Discovery Rate (q < 0.05), separately for each test (Main Effect of Group; Main Effect of Condition; Interaction of Group by Condition) by pooling the p-values from both hemispheres’ cortical surfaces. This False Discovery Rate threshold corresponded to uncorrected p < 4×10−6 for the Main Effect of Group. Four significant results (contiguous significant vertices) survived this threshold and were taken forward for the next analysis step. For the Main Effect of Condition and Interaction of Condition x Group, no results survived statistical correction FDR (q < 0.05).
A similar procedure was performed for subcortical structures. Beta series were obtained for each subject, structure, and experimental condition, from their standard Freesurfer subcortical parcellations by masking the EPI data within each structure and calculating the average of the voxels. Each structure’s beta series was then correlated with the whole-brain gray matter beta average, separately for each condition, and the resulting values were subjected to linear mixed effects models with the same factors as above. Tests for Main Effect of Condition, of Group, and the Interaction of these factors was performed. All p-values were greater than p > 0.001 and no results survived an FDR-correction calculated over them.
Follow-up seed-to-whole-brain testing
The first analysis step (seed definition, described above) identified which, if any, brain areas showed the largest connectivity differences between groups. However, this step is insufficient to localize the other specific regions driving this pattern. An analogy is in Analysis of Variance, where a significant omnibus test indicates a difference exists, but follow-up testing is required to determine where in the model differences exist (Gotts et al., 2012). Thus, to locate the regions driving this pattern, we undertook a second step: follow-up seed-to-whole-brain testing (Cole et al., 2010; Gotts et al., 2012; Jasmin et al., 2019c). Each seed region was examined with respect to its connectivity pattern with every cortical vertex and subcortical structure.
For each of the 90 beta series (30 subjects by three conditions), values within the seed vertices was averaged and then correlated with the beta series for every vertex in the brain. These correlations were Fisher z-transformed and used as the dependent variables in linear mixed effects models (3dLME) with the same fixed and random effects as above. For each of the seeds, we tested for the group difference (Amusia vs Control) in connectivity. Results were False Discovery Rate corrected to (q < 0.05) across all eight follow-up tests [4 seeds × 2 hemispheres] corresponding to a threshold of p < 0.00035. Similar, for the subcortical structures, each seed beta series was correlated with subcortical structure beta series, with resulting values subject to statistical testing. An FDR correction over all tests involving subcortex was applied. For display in figures, the data were converted from SUMA’s standard mesh (std.141) to Freesurfer’s standard surface (fsaverage) using AFNI’s SurfToSurf program and mapping values from the closest nodes (i.e. vertices).
Analysis of activation
A standard General Linear Model comparing activation strength (rather than connectivity) was also conducted. As in the General Linear Model for obtaining beta weights, no basis function was used, and polynomials up to second degree were included in the models.
Data availability
The data that support the findings of this study are openly available in the Birkbeck repository.
Funding
This study was funded by Seed Grant No. 109719/Z/15/Z from The Wellcome Trust to A.T.T., a Reg and Molly Buck Award from the Society for Education, Music and Psychology Research to K.J., and an Early Career Fellowship from the Leverhulme Trust (ECF-2017-151) to K.J.
Competing Interests
The authors have no competing interests to declare.
Acknowledgments
We would like to thank our study participants.
Footnotes
Methods moved to end of manuscript. Title and abstract shortened.





{kind=link}
{kind=link}
{kind=link}
{kind=link}