

Abstract
The corrinoid B12 is synthesised only by prokaryotes yet is widely required by eukaryotes as an enzyme cofactor. Microalgae have evolved B12 dependence on multiple occasions and we previously demonstrated that experimental evolution of the non-requiring alga Chlamydomonas reinhardtii in media supplemented with B12 generated a B12-dependent mutant (hereafter metE7). This clone provides a unique opportunity to study the physiology of a nascent B12 auxotroph. Our analyses demonstrate that B12 deprivation of metE7 disrupted C1 metabolism, caused an accumulation of starch and triacylglycerides and a decrease in photosynthetic pigments, proteins and free amino acids. B12 deprivation also caused a substantial increase in reactive oxygen species (ROS), which preceded rapid cell death. Surprisingly, survival could be improved without compromising growth by simultaneously depriving the cells of nitrogen, suggesting a type of cross protection. Significantly, we found further improvements in survival under B12 limitation and an increase in B12 use-efficiency after metE7 underwent a further period of experimental evolution, this time in coculture with a B12-producing bacterium. Therefore, although an early B12-dependent alga would likely be poorly adapted to B12 deprivation, association with B12-producers can ensure long-term survival whilst also providing the environment to evolve mechanisms to better tolerate B12 limitation.
Introduction
Over 50% of algal species require an exogenous source of B12 for growth (1), yet large areas of the ocean are depleted of this vitamin (2, 3). Eukaryotic algae cannot synthesise B12, but must instead obtain it from certain prokaryotes that can (1). Indeed, whilst dissolved B12 concentrations are positively correlated with bacterioplankton density (4, 5), they have been found to negatively correlate with phytoplankton abundance (6, 7). Furthermore, nutrient amendment experiments suggests B12 limits phytoplankton growth in many aquatic ecosystems (8–10). Despite this, understanding of the physiological and metabolic adaptations that B12-dependent algae employ to cope with B12 deprivation is rather limited.
In many algae B12 is required as a cofactor for the B12-dependent methionine synthase enzyme (METH) (11), although some algae encode a B12-independent isoform of this enzyme (METE) and do not require B12 for growth. Bertrand et al (12), showed that the B12-dependent marine diatom Thalassiosira pseudonana, which encodes only METH, responds to B12 scarcity by increasing uptake capacity and altering the expression of enzymes involved in C1 metabolism. Heal et al (13) found that despite these responses B12 deprivation disrupted the central methionine cycle, transulfuration pathway and polyamine biosynthesis. Phaeodactylum tricornutum, a marine diatom which uses but does not depend on B12 (encoding both METE and METH), responds similarly to T. pseudonana (12) but can also rely on increasing expression of METE to maintain the production of methionine. Phylogenetic analysis of the METE gene among diatoms shows no simple pattern of gene loss or gain, as indeed is the case across the eukaryotes (14, 15), but there is a clear link between the lack of a functional copy of the METE gene and B12-dependence (11, 16).
As with the diatoms, the phylogenetic distribution of METE within the Volvocales (a family of green freshwater algae) points to gene loss on several independent occasions. The genomes of two volvocalean algae, V. carteri and G. pectorale, contain METE pseudogenes indicating that B12 dependence has evolved relatively recently in these species (11). Chlamydomonas reinhardtii is a related alga that encodes a functional copy of METE and so is B12-independent. Helliwell et al. (17) generated a METE mutant of C. reinhardtii by experimental evolution in conditions of high vitamin B12 concentration, demonstrating that sustained levels of B12 in the environment can drive METE gene loss. This mutant, which contained a Gulliver-related transposable element in the 9th exon of the METE gene, was completely reliant on B12 for growth but in the presence of the vitamin it was able to outcompete its B12-independent progenitor. In the absence of B12 the METE mutant would sometimes revert to B12 independence and resume growth. Reversion was found to be due to excision of the transposon to leave behind a wild-type METE gene sequence, but there was a single case where 9 bp fragment of the transposon was left behind resulting in a stable B12-dependent strain hereafter called metE7.
C. reinhardtii is a well-researched model organism that has been instrumental in improving our understanding of algal photosynthesis, ciliogenesis, and responses to fluctuating nutrient environments (18–20). We wanted to use the metE7 mutant of C. reinhardtii to study how recently acquired B12 auxotrophy impacts an organism’s fitness and physiology, and to provide insight into the metabolic challenges that other B12 dependent algae might have faced when they first evolved. In this work we characterized the responses of metE7 to different vitamin B12 regimes and compared them to the responses of its ancestral B12-independent strain, and to a closely related, naturally B12-dependent alga Lobomonas rostrata. The responses of metE7 to B12 deprivation were quantified by measuring changes in gene expression, cellular composition, photosynthetic activity and viability, and were contrasted against changes under nitrogen deprivation. To assess whether a recently evolved algal B12 auxotroph could improve its survival during B12 deprivation relatively quickly, we subjected metE7 to a further experimental evolution period of several months in limited B12 or coculture with a B12 producing bacterium and characterised the resulting lines.
Materials and Methods
Strains
Mesorhizobium loti (MAFF 303099) was a gift from Prof. Allan Downie at the John Innes Centre, Norwich, UK. Algal strains used in this study are shown in Table S1 and include Lobomonas rostrata (SAG 45/2), as well as several Chlamydomonas reinhardtii strains derived from strain 12 of wild type 137c or the cell wall-deficient strain cw15. The stable B12-dependent metE7, the unstable B12-dependent (S-type) as well as the B12-independent revertant line (R-type) all evolved from the strain 12 of wild type 137c (Ancestral) as described by Helliwell et al. (2015). Another B12-dependent mutant (metE4) was generated by targeted (CRISPR/Cpf1) knockout of the METE gene in the UVM4 strain using the protocol described in Ferenczi et al. 2017 (21),
Culture conditions and growth measurements
Algal colonies were maintained quarterly on Tris-acetate phosphate (TAP) + 1000 ng·l−1 cyanocobalamin (B12) agar (1.5%) in sealed transparent plastic tubes at room temperature and ambient light. Cultures were grown in TAP or Tris min medium under continuous light or a light-dark period of 16hr-8hr, at 100 µE·m−2·s−1, at a temperature of 25°C, with rotational shaking at 120 rpm in an incubator (InforsHTMultitron; Switzerland). For nutrient starvation experiments the pre-culture TAP media contained 200 ng·l−1 of B12, and when cell densities surpassed 1*106 cells·ml−1 or an OD730 nm of 0.2, cultures were centrifuged at 2,000 g for 2 minutes, followed by supernatant removal and resuspension of the cell pellet in media.
Algal cell density and optical density at 730 nm were measured using a Z2 particle count analyser (Beckman Coulter Ltd.) with limits of 2.974-9.001 µm, and a FluoStar Optima (BMG labtech) or Thermo Spectronic UV1 spectrophotometer (ThermoFisher) respectively. Mean cell diameter was also quantified on a Z2 particle analyser (Backman Coulter Ltd.). Dry mass was measured by filtering 20 ml of culture through pre-dried and weighed grade 5 whatmann filter paper (Sigma-Aldrich WHA1005090), drying at 70°C for 24 hours, followed by further weighing on a Secura mass balance (Sartorius). Algal and bacterial CFU·ml were determined by plating on solid media.
Measurement of photosynthetic parameters
200 µl of cultures with an OD730 nm>0.1 were transferred to a 96 well plate which was then incubated at 25°C in the dark for 20 minutes. F0 was measured prior to, and Fm during, a saturating pulse at 6172 µE·m−2·s−1. The light intensity was increased to 100 µE·m− 2·s−1 and the cells allowed to acclimate for 30 seconds prior to another set of fluorescence measurements before and during a saturating pulse. From these fluorescence measurements the CF imager software calculated non-photochemical quenching (Fm/Fm’-1), PSII maximum efficiency (Fv’/Fm’), and the coefficient of photochemical quenching (Fq’/Fv’) at each light intensity.
Measurement of cellular biochemical composition
Lipids were extracted from the cell pellet from 10 ml of culture using the chloroform/methanol/water method and triacylglycerides (TAGs), polar lipids and free fatty acids in the total lipid extract and total fatty acid methyl esters (FAMEs) were analysed by GC-FID and GC-MS, as described in Davey et al. (2014) (22). A 1 ml aliquot of algal culture was used for pigment and starch quantification as described in Davey et al. (2014), and a 10 ml aliquot for protein quantification using a Bradford assay and amino acids by HPLC as described in Helliwell et al. (2018) (23).
Reactive oxygen species quantification
2 µl of 1 mM 2’,7’ Dichlorofluorescein diacetate (Sigma-Aldrich) dissolved in DMSO was added to 198 µl of cell culture in a black f-bottom 96 well plate (Greiner bio-one) and incubated at room temperature in the dark for 60 minutes before recording fluorescence at 520 nm after excitation at 485 nm in a FluoStar Optima Spectrophotometer (BMG labtech). Fresh cell culture media devoid of any cells was used as a blank.
SAM and SAH quantification
10 ml of samples were centrifuged at 2,000 g for 2 minutes, supernatant removed, and cell pellet lyophilised at <-40°C and <10 pascals for 12-24 hours. 300 µl of 10% methanol (LC-MS grade) spiked with stable isotope-labelled amino acids (L-amino acid mix, Sigma-Aldrich, Co., St. Louis, MO, USA) was added to each sample. They were vortexed 3 times, every 10 min, before sonicating for 15 min in an iced water bath then centrifuging (16,100 x g) for 15 min at 4°C. Quantitative analysis was performed on 150 µl of supernatant using an Agilent 6420B triple quadrupole (QQQ) mass spectrometer (Agilent Technologies, Palo Alto, USA) coupled to a 1200 series Rapid Resolution HPLC system. Details of the HPLC-MS are given in the supplementary information.
Transcript quantification
Total RNA extraction was performed on the cell pellet from 10 ml of algal culture using the RNeasy® Plant Mini Kit (QIAGEN). DNase treatment was carried out using TURBO DNA-free™ kit (Ambion), and cDNA synthesis using SuperScript®III First-Strand synthesis system for RT-PCR (Invitrogen) according to the manufacturer’s instructions. RT-qPCR was performed as described by Helliwell et al. 2018 (24), using primers listed in Table S2
Artificial Evolution setup
A culture of metE7 cells was plated on TAP +1000 ng·l−1 B12 agar, then 8 colonies picked and resuspended in TAP + 200 ng·l−1 B12 in a 96 well plate. Each well was split into 3 wells, each in a different 96 well plate containing 200 µl of a different media: TAP +1000 ng·l− 1 B12, TAP +25 ng·l−1 B12, and TP medium. M. loti was prepared in a similar manner to metE7, except preculturing was performed in TP + 0.01% glycerol. M. loti was added to the TP culture containing metE7 at a density roughly 20 times greater than the alga. The 96 well plates were incubated at 25°C, under continuous light at 100 µE·m−2·s−1, on a shaking platform at 120 rpm. Each week the cultures were diluted: Those in TAP +1000 ng·l−1 B12 were diluted 10,000-fold, TAP +25 ng·l−1 B12 = 100-fold, and TP = 5-fold. Every three weeks 10 µl of serial dilutions of each culture was also spotted onto TAP agar + Ampicillin (50 µg·ml−1) and Kasugamycin (75 µg·ml−1) and TAP agar + 1000 ng·l−1 B12 to check for B12-independent C. reinhardtii, or bacterial contaminants and to act as a reserve in the case of contamination. If cultures were found to be contaminated, then at the next transfer they were replaced by colonies from the same well that had grown on the TAP agar plates. At four points during the 12-month evolution period all cultures were transferred to TAP agar plates where they were stored for 2 weeks during an absence from the lab, meaning that the total time in liquid culture was 10 months. See Fig S9 for an illustration of the experimental evolution setup and the tests of B12 dose response and viability during B12 deprivation that were performed on the evolved lines.
Results
B12 deprivation causes substantial changes to C1 metabolism in the metE7 mutant
Methionine synthase plays a central role in the C1 cycle (Fig. 1A), and thus facilitates nucleotide synthesis and production of the universal methyl donor S-adenosylmethionine, which is essential for many biosynthetic and epigenetic processes (25, 26). Wild-type (WT) C. reinhardtii can operate these cycles in the absence of B12 using the methionine synthase variant METE, but metE7 relies solely on the B12-requiring METH isoform. Before investigating the effect of B12 deprivation on C1 metabolism in metE7 we first wanted to eliminate the possibility that other mutations in the experimentally evolved metE7 line might account for its B12 dependent phenotype. We therefore generated an independent METE mutant line (metE4) using CRISPR/Cpf1 (21). This mutant has an in-frame stop codon (Fig. S1) and, as predicted, exhibits B12-dependence. We therefore proceeded to investigate the effect of B12 on C1 metabolism in the metE7 line, since its origin is a closer reflection of how B12 auxotrophy would have arisen naturally in other algae.

C1 cycle metabolites and transcripts increase during B12 deprivation of metE7. (A) Metabolic map of a portion of the C1 cycle centred around METE and METH, with enzyme abbreviations in black, metabolite abbreviations in grey, and arrows depicting enzyme-catalysed reactions. (B) Abundance of six transcripts for enzymes of the C1 cycle measured by RT-qPCR on RNA extracted from the ancestral line and metE7 after 30 hours of incubation in mixotrophic conditions with (1000 ng·I−1) or without B12. (C) Abundance of Met, SAM, and SAH metabolites measured by HPLC-MS on the same samples as above. Metabolite and transcript abundances are expressed as levels in B12-deprived conditions relative to B12-replete conditions and presented on a log2() scale. Error bars = sd, n=3-4, ‘ns’=not significant, *=p<0.05, **=p<0.01, ***=p<0.001, Welch’s t test. WT = ancestral B12-independent strain, metE7 = experimentally evolved B12-dependent line. See also figure S2.
Both the WT ancestral line and metE7 were precultured in TAP medium in continuous light with adequate (200 ng·l−1) B12 to maintain a low cellular quota of the vitamin. The cells were then pelleted, washed and transferred to B12 replete (1000 ng·l−1) or B12 deprived (no B12) TAP medium at 5×105 cells/ml and grown for 30 hours. Steady state transcript levels of six enzymes in the C1-cycle were then investigated by RT-qPCR (Fig. 1B). In the WT, three transcripts (METE, SAH1, and MTHFR) were significantly (p<0.05) upregulated by B12 deprivation, while in metE7 all six (including METH, METM, and SHMT2) increased. Levels of the methionine cycle metabolites methionine, SAM and SAH were quantified by HPLC-MS. In the WT there was no difference in methionine, SAM or SAH levels in the two conditions (Fig. 1C). However, in metE7 cells under B12 deprivation methionine levels were raised 6-fold, which was somewhat unexpected given that methionine synthase activity was impeded. SAH levels were also significantly elevated, whereas there was no effect on SAM. Consequently, the SAM:SAH ratio decreased by 10-fold to 3:1 under B12 deprivation. We then studied the dynamics of these changes by measuring metabolites and RNA abundance at several points during 3 days of B12 deprivation and then for 2 days following add-back of 1000 ng·l−1 B12. The transcripts for all six tested C1 cycle genes increased rapidly in the first 6 h and then plateaued; reintroduction of B12 led to an immediate reduction to near initial amounts (Fig. S2A). Similar profiles were seen for the metabolites SAM and SAH, although the peak occurred later at 24 h (Fig. S2B). Methionine levels were more variable, but nonetheless there was a similar trend of a peak 24 h after removal of B12. More significantly, the SAM:SAH ratio fell sharply from 30 to less than 1 within 24 h. A subsequent gradual increase occurred over the next 2 days, and resupply of B12 increased this ratio further over the following 2 days. The likelihood therefore is that many cellular processes would be impacted in B12-deprived metE7 cells.
B12 deprivation significantly impacts cell physiology and biochemical composition
Our data demonstrate a substantial impact of B12 limitation on the expression of C1 metabolic genes as well as the abundance of C1 metabolites. To elucidate downstream consequences of perturbed C1 metabolism we also characterised broader physiological responses to B12 deprivation. As has been documented previously (17), growth of metE7 cells was significantly impaired in B12-deprived conditions (Fig. S3A). However, by day 2 the B12 deprived cells had a 36% larger diameter resulting in a 150% increase in volume (Fig. 2A and Fig. S3B), indicating that cell division was more restricted than overall growth. Moreover, cell viability, which was assayed by the ability of cells to form colonies when plated on B12-replete TAP agar, decreased to below 25% within 4 days of B12 limitation (Fig. S3C). This was preceded by a reduction in photosystem II maximum efficiency (Fv/Fm) (Fig. S3D), an often-used indicator of algal stress (27, 28).

B12 deprivation of metE7 causes cell enlargement and significant changes in macromolecular composition. (A) Microscope photographs taken at 1000× magnification of metE7 cells grown in TAP medium in B12 replete (1000 ng·l−1) or B12 deprived (0 ng·l−1) conditions over a period of 4 days (B) Macromolecular composition of B12-deprived cells on day 2 and day 4 of the growth period expressed as mass of those compounds normalised to total cell dry mass and then expressed relative to the amounts in B12 replete conditions. Error bars = sd, n = 5. **=p<0.01, ***=p<0.001, Welch’s t test.
The biochemical composition of C. reinhardtii cells is altered considerably and similarly under various nutrient deprivations and so we hypothesised that B12 limitation would also induce broadly the same responses (20, 29, 30). Therefore, metE7 cells were precultured as before in 200 ng·l−1 B12, then washed and resuspended in TAP with (1000 ng·l−1) or without B12 and cultured mixotrophically for 4 days. Cultures were visually inspected by microscopy (Fig. 2A) and the amounts of various cellular components were measured on day 2 and 4 (Fig. 2B). Chlorophyll levels declined considerably under B12 deprivation so that by day four the cells had a bleached appearance with an 85% lower concentration than the B12 replete cells. Similarly, free fatty acids (FFA), polar lipids and proteins were at least 50% lower under B12 deprived conditions on day 4. Starch content on the other hand, showed the largest absolute increase from B12 replete to B12 deprived cells (Fig S3), and triacylglycerides were 10-fold higher in B12-deprived cells (Fig 2B), which effectively balanced the loss of polar lipids and free fatty acids so that overall lipid levels were roughly 8-10% of dry mass in both treatments. To look in more detail, quantification of free amino acids and fatty acid composition of all lipid classes was carried out (Fig S4). By day 4 most of the amino acids decreased significantly under B12 deprivation. Particularly noteworthy is the reduction in methionine, in contrast to its elevation at an earlier timepoint, and the increase in glutamine, the only amino acid to be more abundant in B12 deprived cells. Overall the degree of fatty acid saturation was higher under B12 deprivation, due mainly to an increase in the dominant saturated fatty acids palmitate (16:0) and stearate (18:0) (Fig. S5B), although levels of several unsaturated fatty acids, in particular 16:2, 16:3(7,10,13), 18:1 and 18:2, were also elevated.
Responses to nitrogen deprivation improve survival under B12 deprivation
Our results demonstrate that B12 deprivation of metE7 causes several changes in biochemical composition akin to those exhibited following nitrogen deprivation of WT C. reinhardtii. To further investigate this comparison we measured growth, viability, and photosynthetic efficiency under both conditions over a timecourse (Fig. S6). metE7 culture density increased more under B12 than nitrogen deprivation (Fig. S6A), but started to decline after day 2, unlike under nitrogen deprivation where growth continued more slowly over 4 days. For cell viability, both conditions caused a decline, but while loss of viability continued in B12 deprived cells, under nitrogen deprivation the initial loss was followed by recovery (Fig. S6B). Maximum photosynthetic efficiency of photosystem II, however, did not recover under either condition, and its decline was more rapid in nitrogen-deprived cells (Fig. S6C).
The increased viability of metE7 under nitrogen compared with B12 deprivation suggested to us that either the metabolic role of B12 would make it intrinsically more difficult to cope without or that the evolutionary naivety of metE7 to B12 dependence would mean it had little time to evolve protective responses to B12 limitation. We therefore tested whether responses to nitrogen deprivation could afford some protection against B12 deprivation. Viability measurements were monitored over several days, and cultures lacking nitrogen or B12 behaved as previously (Fig 3A). However, metE7 cells deprived of both nitrogen and B12 simultaneously were more similar to those starved on nitrogen: there was an initial decrease in viability followed by recovery to a level significantly higher than in B12 deprivation alone. As total growth in B12 and nitrogen deprivation was not significantly different from B12 deprivation alone (Fig. S7) this apparent protective mechanism in response to nitrogen deprivation is not simply a result of inhibiting growth and hence avoiding severe B12 starvation.

metE7 survives better and produces lower levels of reactive oxygen species (ROS) when limited for both N and B12 than just B12 alone. (A) Percentage of cells that could form colonies (a measure of viability) on nutrient replete agar when removed at different timepoints from nutrient deprivation conditions (Indicated in panels above the graphs). (B) Reactive oxygen species (ROS) measured by dichlorofluorescein diacetate (DCFDA) fluorescence and normalised both on a per cell basis and to the nutrient replete treatment (+B12+N). Error bars = sd, n = 3-6.
In C. reinhardtii, as in many photosynthetic organisms, the absorption of light energy in excess of that required for metabolism can increase the production of reactive oxygen species (ROS) (31). To investigate whether the cell death observed under B12 deprivation of metE7 could be due to ROS, the general ROS-sensitive dye dihydrodichlorofluorescein diacetate was incubated with cells at different timepoints during nutrient deprivation. We found that ROS levels increased in all nutrient deprived conditions in the first two days but were highest in those cells deprived of B12 alone (Fig. 3B). This peak coincided with the start of the substantial decline in cell viability (Fig. 3A). The combination of B12 and nitrogen deprivation reduced ROS levels to similar amounts to those seen in the nitrogen-deprived cells, and so may be a factor behind reduced cell death.
Natural B12 auxotroph Lobomonas rostrata fares better under B12 limiting conditions than metE7
Considering that metE7 quickly lost viability in the absence of B12 while nitrogen starvation invoked protective responses independent of B12 status, it is possible that as a novel auxotroph metE7’s response to B12 deprivation is simply underdeveloped. To test this we compared the B12 physiology of metE7 with Lobomonas rostrata, a naturally B12-dependent member of the same Volvocaceae family of chlorophyte algae (32, 33). Cell viability was significantly greater in L. rostrata cells compared to the metE7 line after 2-4 days of B12 deprivation despite also growing to a greater density (Fig S7A). Moreover, a B12 dose-response experiment, in which the two species were each cultured mixotrophically in a range of B12 concentrations, revealed that L. rostrata reached a higher optical density than metE7 at all B12 concentrations below 90 ng·l−1, while the inverse was true above 90 ng·l−1 (Fig 4A). This indicates that L. rostrata has a lower B12 requirement than metE7.

L. rostrata grows better than metE7 in coculture with a B12 producing bacterium, in part due to its lower demand for B12. (A) Cultures were grown mixotrophically (TAP medium in continuous light), B12 concentrations ranged from 0 to 200 ng·l−1 and precultures of the algae, which were grown with 200 ng·l−1 B12, were washed thrice and inoculated at a density of roughly 100 cells·ml−1. Culture density was measured as optical density at 730 nm after 5 days of growth for the C. reinhardtii strains and 9 days for L. rostrata. (B) Cultures were grown photoautotrophically (Tris minimal media in 16h:8h light:dark cycles) in axenic culture (with 100 ng·l−1 B12) or coculture (with the B12-producing bacterium M. loti) over a period of 16 days with measurements of cell density every 1-2 days. For both panel A and B, black = L. rostrata, grey = metE7, error bars = sd, n=4.
In the natural environment the ultimate source of B12 is from prokaryotes since they are the only known B12 producers (34). In separate studies it was shown that B12-dependent growth of L. rostrata and metE7 can be supported by the B12 synthesising bacterium Mesorhizobium loti (17, 35). We therefore compared directly the growth of metE7 and L. rostrata in B12-supplemented (100 ng·l−1) axenic culture and in coculture with M. loti in media lacking a carbon source (TP) (Fig 4A). Even though metE7 grew much more quickly and to a higher density than L. rostrata under axenic, B12-supplemented conditions, it grew less well in coculture with M. loti (Fig. 4B), indicating B12 provision from the bacterium is less effective at supporting the growth of metE7 than of L. rostrata, perhaps simply due to their different B12 requirements, but possibly due to more sophisticated symbiotic interactions.
Experimental evolution in coculture improves B12-use efficiency and resilience to B12 deprivation
Together our data suggest that the newly evolved metE7 line is poorly adapted to coping with B12 deprivation, but we wanted to determine whether the metE7 line could evolve improved tolerance to B12 limiting conditions, so we employed an experimental evolution approach. We designed three distinct conditions, referred to as H, L and C. Condition H (TAP medium with high (1000 ng·l−1) B12) was a continuation of the conditions that had initially generated metE7 (17). Condition L (TAP medium with low (25 ng·l−1) B12) was chosen so that B12 would limit growth. Condition C (coculture with M. loti in TP medium) was a simplification of an environmental microbial community. Eight independent cultures for each condition were established from a single colony and then subcultured once per week over a total period of 10 months. To account for the different growth rates in the three conditions, we applied the following dilution rates of 10,000, 100, and 5 times per week in condition H, L and C respectively (Fig S8). After 10 months under selective conditions all 24 cultures had survived and were then treated with antibiotics to remove the M. loti from condition C and to ensure that there were no other contaminating bacteria. We then subcultured the lines in mixotrophic conditions with TAP + 200 ng·l−1 B12 three times over nine days to ensure they were all acclimated to the same conditions. The behaviours of the algal populations, hereafter referred to as metE7H, metE7L, and metE7C, were then compared alongside the progenitor metE7 line, which had been maintained on TP agar with 1000 ng·l−1 B12 without subculturing.
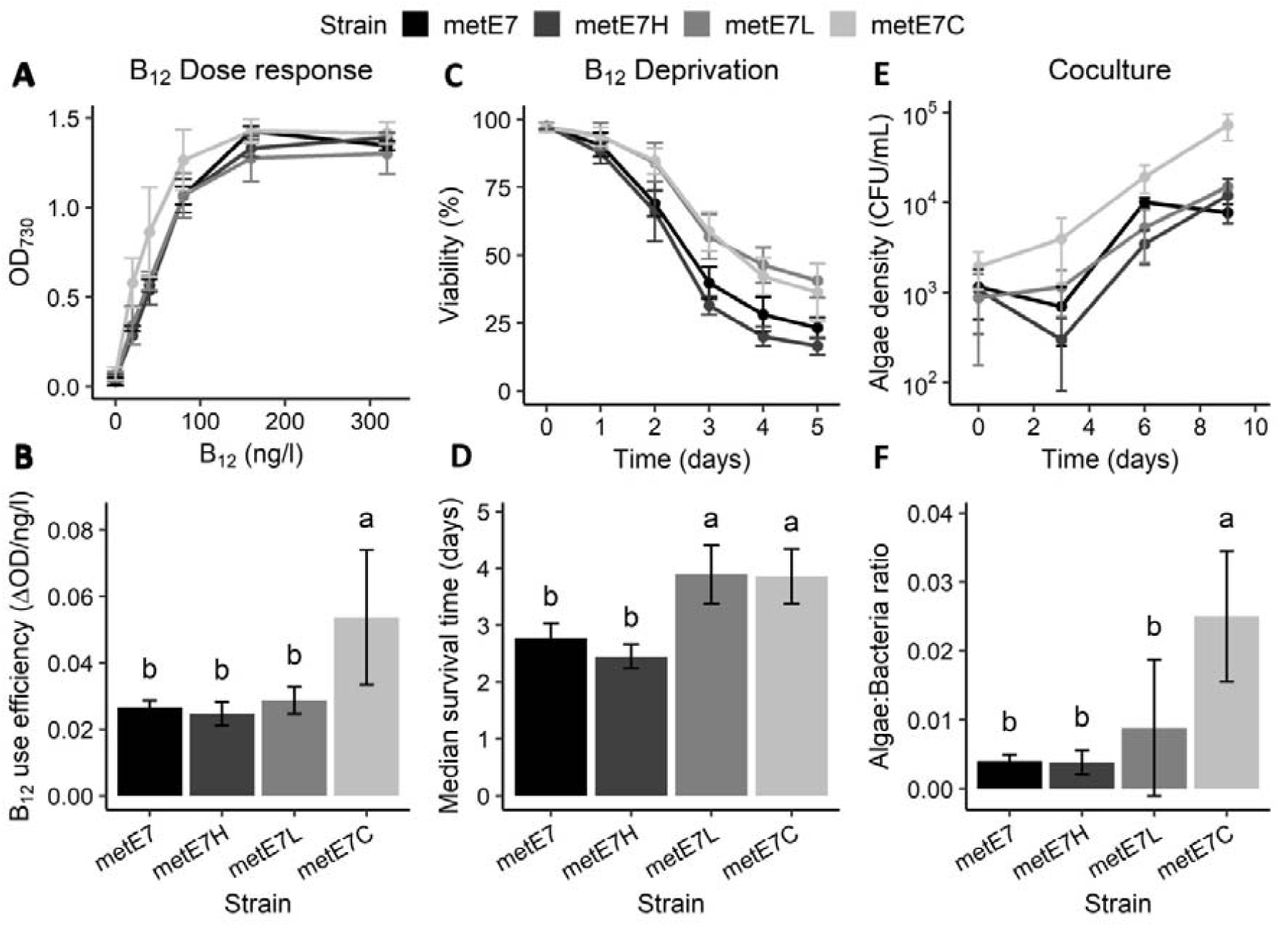
Under high levels of B12 (320 ng·l−1) a similar optical density was reached by the progenitor metE7 strain and the metE7H and metE7C populations, whereas metE7L growth was somewhat compromised (Fig. S10A). When grown across a range of B12 concentrations to determine a dose response, the metE7C populations reached a significantly higher optical density at the lower concentrations of 20 and 40 ng·l−1 B12 than the other lines(Fig. 5A). The concentration of B12 required to produce half the maximum growth (EC50) of metE7C was therefore much lower than the progenitor metE7 or metE7H (Fig. S10B) and this was reflected in the higher B12 use efficiency i.e. the maximal increase in yield (OD730) that results from an increase in B12 concentration (Fig. 5B). However, the maximal growth rate of metE7C was significantly lower (Fig. S10C), and it is tempting to conclude that this is a necessary trade-off. We also compared the viability of the experimentally evolved lines during B12 deprivation (Fig. 5C). Fig. 5C shows that although all lines lost viability during B12 deprivation, metE7L and metE7C survived substantially better, with a median survival time more than a day longer (Fig. 5D) than both the progenitor metE7 and metE7H.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Experimental coevolution of metE7 with the bacterium M. loti selects for improved algal growth in coculture, increased B12 use efficiency and better resilience to B12 deprivation. (A) Maximum optical density achieved by mixotrophically-grown cultures of experimentally evolved lines of metE7 grown over a period of 12 days in six different concentrations of B12. (B) B12 use efficiency of evolved lines calculated using a fitted Monod equation and expressed as the maximum rate of increase in OD730 that would result from an increase in B12 concentration. (C) Viability (measured as the percentage of cells capable of forming colonies on B12-replete agar) of mixotrophically-grown cultures of experimentally evolved lines of metE7 over a 5-day period cultured in 40 ng· l−1 B12. (D) Median survival time of evolved lines after dilution of culture to 40 ng· l−1 B12 calculated using a fitted Verhulst equation. (E) Algal cell density of photoautotrophically-grown cocultures of experimentally evolved lines of metE7 with M. loti over a 9-day period. (F) Ratio of algae to bacteria on the final day (day 9) of growth in coculture. metE7H = metE7 evolved in TAP + 1000 ng· l−1 B12 for 10 months, metE7L = metE7 evolved in TAP media + 25 ng· l−1 B12, metE7C = metE7 evolved in Tris minimal medium in coculture with the B12-producing bacterium M. loti. Error bars = 95% confidence interval, n = 7-8, letters above error bars indicate statistical groupings provided by Tukey’s test, which was performed following a significant ANOVA result.
To elucidate which factors contributed to the improved survival, we performed a multi-parameter physiological analysis (Fig. S11). 16 parameters were measured across the 32 metE7 populations and the dataset visualised in three ways. Fig S11A presents the data as a heatmap with the most similar populations, which generally were those exposed to the same evolution conditions, clustered together to form a phylogenetic tree. Fig. S11B displays the first two components of a principal component analysis of the data, which confirmed that the experimental evolution populations tended to form separate clusters. Fig. S11C is a correlation matrix of the parameters to reveal those pairs that are most positively or negatively correlated with one another. A more definitive statistical approach was then used to determine the most important parameters for predicting survival time during B12 deprivation: Using stepwise minimisation of the Bayesian information criterion of the full linear model the 15 other parameters were reduced to just three. So, it was concluded that higher B12 use efficiency, lower ROS levels and lower maximal growth rate were sufficient to explain longer survival time under B12 deprivation of the metE7 populations.
Comparison of the growth of the evolved lines when cocultured with M. loti showed, perhaps unsurprisingly, that the metE7C lines grew better than the others (Fig 5E), and at the end of the growth period had a significantly higher number of algae supported per bacterium (Fig 5F). This algal:bacterial ratio was also optimally predicted by three parameters: higher algal B12 use efficiency and lower algal maximal growth rate, as for survival time, but also lower algal B12 uptake capacity. Together these results indicate that experimental evolution in coculture not only improves growth in coculture but also increases B12 use-efficiency and survival during B12 deprivation.
Discussion
In this study we exploited a novel model system for the evolution of vitamin B12 dependence by analysing the physiological and metabolic responses to B12 deprivation of an artificially evolved B12-dependent mutant of C. reinhardtii. Our analyses demonstrate that B12 deprivation has important consequences for C1 metabolism: we observed a significant increase in the transcript abundance of C1 cycle enzymes in both the wild type and metE7 strain, and a decrease in the methylation index (SAM:SAH ratio) in metE7 only. Moreover, B12 deprivation of metE7 causes a decrease in chlorophyll, protein and amino acids, and an increase in starch, lipids and saturated fatty acids, characteristic of limitation responses to macronutrients such as nitrogen. The rapid loss of viability seen under B12 deprivation can be averted if the metE7 cells are also limited for nitrogen, suggesting that it is not the lack of B12 per se that causes cell death, but an inability to respond appropriately. Together this suggests a newly evolved B12 auxotroph would be poorly adapted to surviving in the natural environment where a B12 supply is not guaranteed. However, we found that metE7 can be supported for several months by a B12-producing bacterium, and experimental evolution under these conditions caused improved B12 use efficiency and resilience to B12 deprivation.
B12 deprivation of metE7 decreased the SAM:SAH ratio 10-fold, similar to what was reported in a recent B12 deprivation study of the diatom T. pseudonana (13). As SAH is a competitive inhibitor of methyltransferases(36), this decrease would likely lead to general hypomethylation in metE7. The epigenetic marks methyldeoxyadenosine and methylcytosine are similarly abundant in C. reinhardtii and appear to mark active genes and repeat-rich regions respectively, so the consequences of hypomethylation are unclear (37, 38). The reduced abundance of B12-bound METH under B12 deprivation would hinder methionine synthesis and could cause the observed reduction in protein abundance (Fig. 2B). However, methionine levels increased between 12 and 24h of B12 deprivation (Fig S1B), suggesting a reduction in its use, proteolysis, or increased synthesis due to higher METH expression or via alternative pathways such as the S-methylmethionine cycle, as documented in plants (39).
METE transcript abundance showed a much higher dynamic range than METH during B12 deprivation and add-back (Fig. S2A), which is reflected by the higher diurnal range of METE observed in global transcriptomics and proteomics datasets (40). However, on average METE is around 60-fold more abundant than METH in C. reinhardtii (40). This may be due to a lower maximal catalytic rate of METE, as has been observed in E. coli (41), or due to its role in the flagella, which contain METE but not METH (42). Under B12 deprivation conditions the activity of METH would be compromised, yet in both metE7 and the ancestral strains it was upregulated. This is more similar to the B12 dependent algae T. pseudonana and Tisochrysis lutea, which also upregulate METH on B12 deprivation (12, 43), than the B12 independent P. tricornutum, which decreases METH expression (44). However, in both T. pseudonana and P. tricornutum B12 deprivation substantially upregulates C1 cycle enzymes including homologs of METM, MTHFR and SAH1 (12), reflecting our findings and those of Helliwell et al. (2014) (45). Under sulfur and nitrogen deprivation conditions these C1 cycle genes are downregulated, suggesting that their upregulation during B12 deprivation is not a general response to nutrient stress, but a nutrient-specific one, as indeed is the case for T. lutea (43, 46, 47).
Chlorosis is a common symptom of nutrient deficiency in C. reinhardtii, evident in nitrogen, sulfur, iron, and zinc limiting conditions and so it is not surprising that B12 deprivation of metE7 caused a substantial decline in total chlorophyll (Fig. 2B) (48–50). The decrease in total protein content occurred more slowly and was less substantial (50% reduction over four days) than reported under nitrogen and sulphur deprivation (80% reduction within one day) (51). During nitrogen and iron starvation in C. reinhardtii membrane lipids decrease drastically concomitant with the increase in TAGs (52, 53). This is very much like what we observed for metE7 under B12 deprivation, although here the level of free fatty acids and polar lipids decreased by a roughly similar amount to the increase in TAGs indicating there is little to no de novo fatty acid synthesis. In addition, B12 deprivation causes similar shifts in fatty acid composition to nitrogen and iron deprivation, most notably a substantial increase in palmitic acid (16:0) and decrease in polyunsaturated 16:4 fatty acid (53, 54). Despite these similarities, B12 deprivation may elicit an increase in TAGs by a different pathway due to disrupted C1 metabolism, as has been observed in several organisms (55–57). This is thought to be due to a reduction in the methylation potential limiting membrane lipid synthesis and hence diverting more lipids towards TAGs (57, 58). Therefore, B12 deprivation could provide a complementary approach to other nutrient deprivation experiments in improving our understanding of lipid metabolism in C. reinhardtii and other algae.
From an evolutionary perspective, the prevalence of vitamin B12 dependence among algae appears somewhat at odds with the severe fitness penalties that would be incurred given limiting dissolved B12 concentrations, particularly when the fitness benefit in replete B12 is marginal (17). However, relative to optimal axenic laboratory conditions in which the metE7 line evolved, in the environment multiple nutrients may colimit growth perhaps even eliciting responses that mitigate against B12 deprivation, as we observed here, and B12-producing bacteria may not simply co-occur with algae but also actively engage in mutualistic interactions (1, 35, 59, 60). Furthermore, our evidence suggests that selection under coculture conditions led to the newly evolved B12 auxotroph developing increased B12 use efficiency and becoming better adapted to tolerating B12 limitation, which could make this line more robust to the unreliable B12 supply in the natural environment. However, these improvements appeared to come at the expense of maximal growth rate in B12 replete conditions (Fig. S10C), which is not unexpected in light of previous experimental evolution studies in C. reinhardtii (61). As one of the conserved responses of C. reinhardtii upon detecting depletion of various nutrients is to decrease cell division, it is possible that slower growth might even be selected for under B12 deprivation. Indeed, a low growth rate was found to be a significant predictor of greater survival time under B12 deprivation, alongside low ROS levels and high B12 use efficiency.
The fact that metE7 survived a 10-month period either with limited artificial supplementation of B12 or by relying completely on bacterial B12 provision, does suggest that even a newly evolved and poorly adapted B12 auxotroph would have ample opportunity to adapt further. What adaptations are likely to improve growth and survival under B12 deprivation are not altogether clear, but it is not unreasonable to assume that exaptation of existing nutrient limitation responses would play a major role. B12 dependence is certainly a risky evolutionary strategy, and one which may have ended in extinction countless times, but our work suggests that even the simplest of symbioses with B12-producing bacteria may be sufficient to ensure the survival and drive the continued evolution of B12-dependent algae.
Competing interests
The authors declare no competing interests.
Author contributions
F.B., P.M. and A.G.S designed the research; F.B., D.L.S. and A.H. performed the research; N.S., D.L.S. and M.P.D. contributed new reagents or analytic tools; F.B. and D.L.S. analysed the data; F.B., A.G.S., K.E.H. and P.M. wrote the paper with input from all authors.
Acknowledgements
This work was supported by the BBSRC Doctoral Training Partnership BB/M011194/1 (FB, AGS), BBSRC BB/M018180/1 (PM, AGS), BB/I013164/1 (KEH & AGS), Leverhulme Trust RPG 2017-077 (MPD, AGS), Natural Environment Research Council NE/R015449/1 (KEH).
Parsed Citations




