

Abstract
Animal development and homeostasis depend on precise temporal and spatial intercellular signaling. Components shared between signaling pathways, generally thought to decrease specificity, paradoxically can also provide a solution to pathway coordination. Here we show that the Bone Morphogenetic Protein (BMP) and Wnt signaling pathways share Apcdd1 as a common inhibitor and that Apcdd1 is a taxon-restricted gene with novel domains and signaling functions. Previously, we showed that Apcdd1 inhibits Wnt signaling, here we find that Apcdd1 potently inhibits BMP signaling in body axis formation and neural differentiation in chicken, frog, zebrafish, and humans. Our results from experiments and modeling suggest that Apcdd1 may coordinate the outputs of two signaling pathways central to animal development and human disease.
Significance Statement Apcdd1 is a taxon-restricted gene that inhibits both BMP and Wnt intercellular signaling pathways in multiple organisms including mice, frog, zebrafish, and chicken. It encodes a bi-functional protein with a novel protein domain that can bind to Wnt and BMP receptors and block downstream signaling.
Introduction
Complex multicellular organisms develop through a series of exquisitely choreographed cellular events, such as differentiation and migration (Nusslein-Volhard and Wieschaus, 1980), which must be temporally and spatially coordinated through the synchronized use of signal transduction pathways. Although the specification of cell fate and function by signaling through individual pathways has been well described, the molecular-level intersections of distinct signaling pathways are often unknown.
Less than 20 signaling pathways are used to build and maintain multicellular eukaryotic organisms (Pires-daSilva and Sommer, 2003). Five pathways predominate: Wnt, BMP/Transforming Growth Factor ß (TGF-ß), receptor tyrosine-kinases (RTK), hedgehog and Notch. The Wnt and BMP pathways operate early in development to establish the anterior-posterior and dorsal-ventral axes of the body and generate essential neuronal populations. At later stages, BMP and Wnt are essential for maintaining the integrity and organization of many adult tissues including skin and hair (Lee and Tumbar, 2012). While seminal studies have shown that periodic, sequential and precisely coordinated BMP and Wnt signaling is required during the adult mouse hair cycle (Kandyba et al., 2013; Plikus et al., 2011; Plikus et al., 2008), the molecular nature of pathway coordination and the control of pathway activation dynamics have remained largely unclear.
Signaling specificity and temporal coordination appear to be conflicting requirements: signal specific responses require the regulation of independent components, while temporally coordinating the activation of signaling pathways requires at least some common components. This problem can be resolved by the existence of factors that interact with multiple signaling pathways. Although a few Wnt/BMP pathway coordinators, including Cerberus and Dkk, have previously been identified (Cruciat and Niehrs, 2013; Piccolo et al., 1999), to elucidate the molecular nature of further pathway coordinators, we examined APCDD1, a gene mutated in a human hair and skin condition called Hereditary Hypotrichosis Simplex (Shimomura et al., 2010). First, Apcdd1 has been shown to be induced by the Wnt pathway on tissues that include the skin, retina, central nervous system (CNS) and vasculature (Chen et al., 2015; Daneman et al., 2010; Kuraku et al., 2005; Mazzoni et al., 2017). APCDD1 then inhibits Wnt signaling in both multiple cell types, including the skin, nervous system, and head structures, and multiple species (human, chicken, frog) (Mazzoni et al., 2017; Shimomura et al., 2010). Second, we previously found that Apcdd1 encodes a dimeric transmembrane protein that complexes with the LRP5 Wnt receptor and lowers Wnt signaling output after activation by the Wnt3a ligand (Shimomura et al., 2010). Third, we noted that human Apcdd1 heterozygous dominant mutations show more severe skin and hair phenotypes than mouse homozygous loss-of-function mutations (Mazzoni et al., 2017; Shimomura et al., 2010), suggesting APCDD1 may block an additional signaling pathway. Since Wnt depletion also affects BMP targets in various tissues (Hikasa and Sokol, 2013; Lim and Nusse, 2013; Xu et al., 2011), we have assessed the hypothesis that Apcdd1 is a multi-functional inhibitor of both the BMP and Wnt pathways.
In this paper, we have examined the effect of APCDD1 on BMP signaling during critical stages of cellular patterning during embryonic development. We found that that Apcdd1 is a potent inhibitor of BMP signaling. We find that in addition to its role in modulating WNT signaling, Apcdd1 directly tunes BMP pathway signaling during gastrulation and tissue patterning, and has a critical role in acquisition of neuronal cell fates. APCDD1 blocks BMP signaling by binding to the BmprIa receptor and inhibiting the nuclear localization of Smad1, the main BMP effector. Using frog, chicken and zebrafish embryos, we find that APCDD1 represses the expression of BMP targets and, interferes with major developmental processes including gastrulation, body axis formation, neural specification and axon pathfinding. APCDD1 is also a Wnt/BMP pathway intersection point: single-cell imaging of BMP and Wnt effectors, coupled with mathematical modeling, show that APCDD1 can coordinate Wnt/BMP activation, suggesting a dynamic explanation for Wnt/BMP periodic sequential activation in mouse hair cycle (Plikus et al., 2011). Finally, Apcdd1 is an ancient orphan gene that may have arisen de novo. It is present in few Metazoan phyla in which BMP signaling and Wnt signaling are used for axial specification, and contains two novel protein domains which resemble each other and are unique to APCDD1 proteins.
Results
Apcdd1 inhibits both Wnt and BMP signaling both in vitro, and in vivo during Xenopus embryogenesis
As a first approach to assessing whether Apcdd1 enables the temporal coordination of BMP and Wnt pathway activation, we tested the effect of APCDD1 expression on activation of BMP and Wnt pathways in multiple independent assay systems. First, we measured the levels of phosphorylated Smad1 (pSmad1, activated by BMP signaling) and β-catenin (activated by Wnt signaling) in cultured NIH 3T3 mouse fibroblast cells after BMP and/or Wnt stimulation. Using quantitative immunofluorescence, we found that the levels of pSmad1 and β-catenin were elevated in the nuclei of NIH 3T3 cells by their cognate ligands BMP2 and Wnt3a (Fig. 1A-C). In contrast, pSmad1 and β-catenin were present at low levels in the absence of ligands (data not shown, Fig. 1C). Similarly, the levels of pSmad1 and β-catenin were reduced in cells transfected with APCDD1 compared to untransfected neighboring cells (Fig. 1A-B, quantification in Fig. 1C), suggesting that Apcdd1 represses the output of both the Wnt and BMP pathways.

A-B. The activation of BMP and Wnt pathways was measured at single-cell level, using NIH 3T3 mouse fibroblast cells cultured in the presence of BMP and Wnt ligands. Cells transfected with APCDD1 and a fluorescence reporter (displayed in red) have low levels of both pSmad1 (green in A) and β-catenin (green in B), while neighboring cells that are not transfected have higher levels of both effectors. Nuclei are visualized with DAPI (blue).
C. The immunofluorescence levels of pSmad1 and β-catenin were measured relative to the background, in the presence (0) or absence (1) of one or both ligands (Bmp2, Wnt3A) or of Apcdd1 (n = 40 cells per condition). The outputs of BMP and Wnt signaling are inhibited by Apcdd1 (p < 0.001 in Bonferroni-adjusted 2-way heteroscedastic t-test comparisons).
D. Effect of Xenopus Apcdd1 (XA1) modulation on BMP-induced transcriptional activity in Xenopus animal injections. Apcdd1 depletion increases transcription induced by exogenous BMP2 RNA, while overexpression of apcdd1 RNA reduces transcription from both the BRE reporter induced by exogenous BMP2, and of the Vent promoter induced by endogenous BMP ligands. *** p < 0.001 in Student’s t-test.
E-I. The phenotype of XA1 depletion on the dorsal side is rescued most effectively by simultaneous BMP7 depletion (quantification in Fig. S3E). Embryos injected at the 4-cell stage on the dorsal side were collected at stage 36. The dorso-anterior index (DAI) values (Kao and Elinson, 1988) are: E – DAI5 (normal), F – DAI2, G – DAI2, H – DAI3, I – DAI4.
J. Human APCDD1 specifically reduces the protein level of type I BMP receptor BMPRIA, but not the type II receptor BMPR2 and nodal type I receptor ACVR1B. Beta-galactosidase serves as loading control. Xenopus embryos were injected in the animal poles.
K. APCDD1 interacts directly with BMPRIA, but not with BMPR2, in CHO cells. APCDD1 and BMPRIA coprecipitated in both APCDD1 (center panel) and BMPRIA (right panel) immunoprecipitation. BMPR2 coprecipitated with BMPRIA (right panel), but not with APCDD1 (center panel).
Next, we tested the effect of Apcdd1 on Wnt and BMP pathway signaling using microarray analysis in Apcdd1-depleted Xenopus embryos. To identify Apcdd1 target genes, and further explore its function across developmental pathways, we examined role of Apcdd1 in the early development of the African clawed frog (Xenopus laevis). Xenopus Apccd1 (XA1) is expressed at neurula and tadpole stages, with an expression pattern similar to that of mouse embryos (Jukkola et al., 2004). Early expression is maternal and zygotic, from 8-cell to gastrula stages, including in the dorsal lip (Spemann organizer) (Fig S1). We performed a microarray analysis of XA1 morpholino oligonucleotide (MO)-depleted Xenopus dorsal cells (Fig. S3A, B). Consistent with results from mouse 3T3 cells, many of the genes that were up-regulated >2-fold in Apcdd1-knockdowns were associated with either the BMP pathway (10 out of 26 protein coding genes, Suppl. Table 1) or the Wnt pathway (2 genes). We thus examined whether Xenopus Apcdd1 could regulate BMP signaling either in vivo (Fig. S2) or in in vitro transcription assays (Fig. 1D). We found that MO-depletion of XA1 in Xenopus embryos expands the expression of sizzled, a downstream target activated by BMP signaling (Fig. S2G-J). The elevated expression of sizzled was reduced by the co-injection of a MO-resistant Apcdd1 RNA (Fig. S2K, L). In contrast, MO-depletion of XA1 reduced the expression of Sox2, a negative target of BMP signaling (Fig S2M-S2R). In the transcription assays, we used a BMP Responsive Element (BRE) reporter (Korchynskyi and ten Dijke, 2002) to characterize the extent of BMP2 stimulation in early Xenopus embryos (Fig. 1D). Strikingly, depleting endogenous XA1 protein resulted in a >3-fold increase in BMP2 responsiveness, whereas the overexpression of XA1 repressed the ability of BMP2 to activate either the BRE reporter or the endogenous Vent2 promoter activated by endogenous BMPs (Fig. 1D) (Hata et al., 2000). This finding was also observed in mouse NIH 3T3 cells; cells transfected with human APCDD1 (hAPCDD1) showed a reduced response to BMP4 responsivity (Fig. S3C). We also transfected cells with the mutant form of hAPCDD1 (hAPCDD1-L9R) present in hereditary hypotrichosis and thought to downregulate APCDD1 protein levels by preventing it from reaching the cell membrane (Shimomura et al., 2010). hAPCDD1-L9R was considerably less effective at inhibiting BMP activity in both NIH 3T3 cells (Fig. S3C) and Xenopus embryos (Fig. S3D), similar to previous findings related to Wnt-dependent transcriptional activity (Shimomura et al., 2010).
Apcdd1-depleted Xenopus embryos showed a strong ventralized phenotype, consistent with the idea that APCDD1 normally plays a critical role in tuning BMP signaling. To test this idea, we used morpholinos to quantitatively reduce BMP signaling. Indeed, we found that experimentally inhibiting the BMP pathway rescues the APCDD1-depletion phenotype in Xenopus embryos. Three BMPs, Bmp2, Bmp4, and Bmp7 each have overlapping expression with Apcdd1 in Xenopus (Plouhinec et al., 2011). We generated MOs directed against these BMPs and found that they were able to rescue the ventralized phenotype of Apcdd1-depleted embryos (Fig. 1E-I, S3E), with the most effective rescue observed after BMP7 depletion (Fig. 1I, S3E). Of these three BMPs, XA1 was also most effective at suppressing the activity of the BMP7 ligand (Fig. S3F). These results confirm that APCDD1 plays a critical role in tuning BMP signaling during development.
Our data show that in addition to its previously known role in inhibiting Wnt signaling, Apcdd1 also tunes BMP signaling. Therefore, to determine if Apcdd1 inhibits Bmp signaling directly, we investigated the molecular mechanism of action through which Apcdd1 blocks BMP signaling (Mizutani and Bier, 2008; Moustakas and Heldin, 2009; Umulis et al., 2009). BMP/Growth and Differentiation Factor (GDF) ligands activate a heterodimeric complex of type I and type II BMP serine/threonine receptors (Bmprs) (Mueller and Nickel, 2012). BMP binding leads to the phosphorylation of Bmpr-regulated Smads (R-Smads; Smad1/5/8), which then act as transcriptional regulators. We found that the over-expression of Xenopus Apcdd1 synergized with chordin, a canonical BMP inhibitor, and dramatically reduced signaling through a constitutively active form of the type IA Bmpr (BmprIa, also known as ALK3) (Fig. S3G). Misexpression of Apcdd1 blocked the phosphorylation of endogenous Smad1 in Xenopus animal cap cells (Sasai et al., 1995), with similar effectiveness to chordin (Fig. S3H), together suggesting that Apcdd1 may block BMP signaling function by directly interacting with BmprIa. The interaction with Apcdd1 and BmprIa may be specific. We co-expressed hAPCDD1 together with BMPRIA (Alk3), activin receptor type IB (ACVR1B, ALK4) or BMPRII in Xenopus embryos (Fig. 1J). Of these three receptors, only the levels of BMPRIA was significantly reduced in the presence of hAPCDD1 (compare ALK3 expression levels in lane 1 and 2, Fig. 1J).
Finally, we performed immunoprecipitation studies to investigate whether there is a direct interaction between Apcdd1 and BmprIa (Fig. 1K). Supporting this hypothesis, BMPRIA-myc immunoprecipitated both APCDD1-Flag and BMPRII-HA in CHO cells (Myc-IP, Fig. 1K). The interaction between APCDD1 and BMPRIA may be specific: while APCDD1-Flag immunoprecipitated BMPRIA-myc, it did not immunoprecipitate BMPRII-HA (FLAG-IP, Fig. 1K). Interestingly, while most APCDD1 protein is monomeric (input, Fig. 1K), the immunoprecipitated form is dimeric (Fig. 1K), suggesting that APCDD1 interacts with BMPRIA as a dimer. Finally, we visualized the BMPRIA-APCDD1 interaction by transfecting CHO cells with both BMPRIA and an inducible form of APCDD1. After APCDD1 induction, BMPRIA appeared to be displaced from the membrane (Fig. S3I) to the perinuclear region (Fig. S3I-L), suggesting that APCDD1 interferes with both the stability and membrane localization of BMPRIA.
Apcdd1 is a BMP antagonist in zebrafish
To determine whether the role of Apcdd1 is conserved between species and at different developmental stages, we examined the function of apcdd1-like (apcdd1l) during zebrafish embryogenesis. RT-PCR and whole-mount RNA in situ hybridization revealed that apcdd1l transcript is maternally deposited and is initially found in all cells in the blastula (Fig. S4A). Expression levels subsequently decline during early gastrulation but then increase specifically in dorsal tissues near the end of gastrulation.
Injection of translation-blocking morpholino to knockdown both maternal and zygotic function (Fig. 2A) caused a unique and highly reproducible phenotype characterized by dynamic changes in dorsoventral patterning (Fig. 2). Surprisingly, apcdd1l morphants initially appeared dorsalized: expression domains of early organizer genes such as chd, gsc and dkk-1 showed marked expansion by 6 hours post-fertilization (hpf; early gastrulation), whereas the domain of ventrolateral marker vent showed corresponding reduction at 6 hpf (Fig. 2B, Fig. S4B-D). Because Apcdd1 also acts as a Wnt-antagonist in diverse vertebrate species (Shimomura et al., 2010), the dorsalized phenotype observed here likely reflects disruption in the role of Apcdd1l restricting maternal Wnt8, a key determinant of the zebrafish organizer (Lu et al., 2011). Evidence of dorsalization persisted until 10 hpf (the end of gastrulation), as shown by expanded domains of brachyury/ta in the notochord (Fig. S2E) and sox19b and otx1b in the neurectoderm (Fig. 2C, S2F), and reduced expression of tp63 in the ventral/epidermal ectoderm (Fig. S4H). Remarkably, the degree of dorsalization showed a dramatic reversal by 12 hpf (6 somites stage) as revealed by nearly normal patterns of these same regional markers (Fig. 2D, and data not shown). Moreover, by 24 hpf embryos took on a mildly ventralized phenotype in the trunk and tail region as shown by expanded domains of gata3 and gata1 in ventral skin ionocytes and hematopoietic tissue, respectively (Hartnett et al., 2010; Hsiao et al., 2007; Lin et al., 2006; Soza-Ried et al., 2010) (Fig. 2G, H).

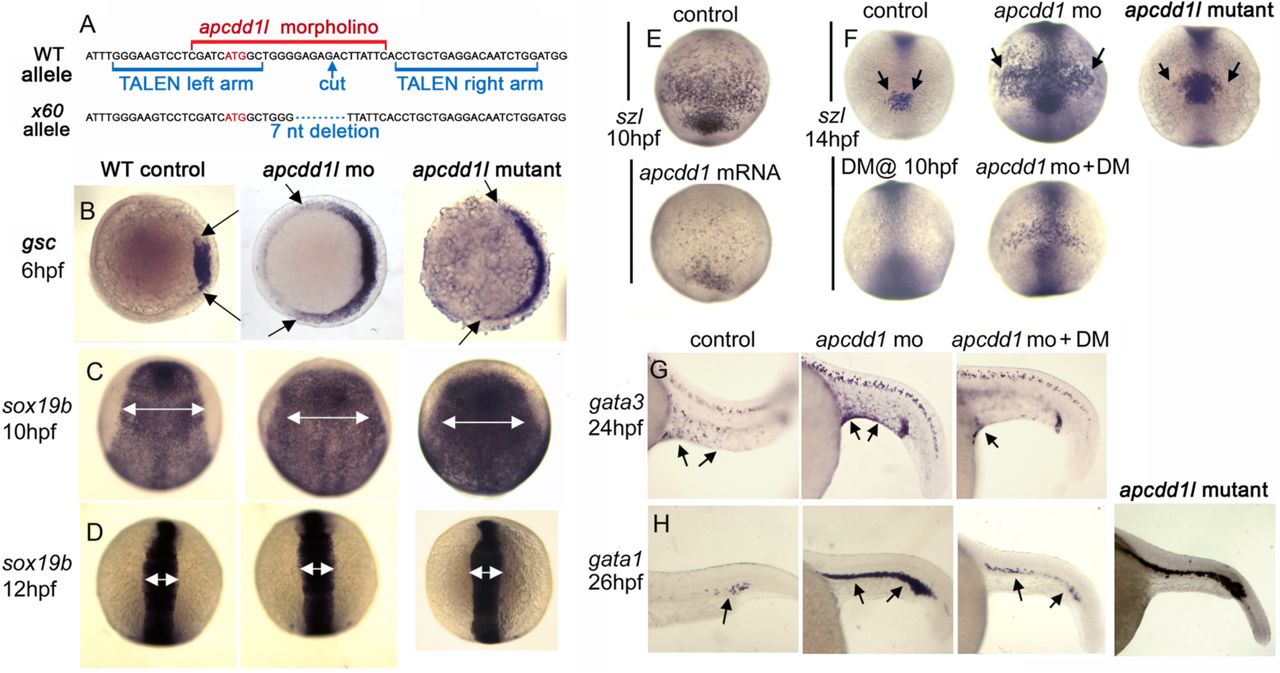
A. Partial sequence of exon 1 surrounding the initiation codon (red font) showing the binding sites for apcdd1l-mo (red font) and TALEN left and right arms (blue font) used to induce double strand breaks. The induced x60 mutant allele has a 7-nucleotide deletion, leading to a frame shift followed by 18 premature stops in exon 1 (a presumptive null).
B. Expression of the organizer gene goosecoid (gsc) at 6 hpf (early gastrula stage). Knockdown of apcdd1l, or loss of apcdd1l in MZmutants, disinhibits maternal Wnt, resulting in expansion of the organizer. Animal pole views with dorsal to the right.
C, D. Expression of neural plate marker sox19b at the end of gastrulation (C) and at the 6-somite stage (D). The early expansion of gsc in apcdd1-morpants and MZmutants results in a dorsalized phenotype that persists through the end of gastrulation, but dorsalization is rapidly reversed by early somitogenesis stages. Dorsal views with anterior to the top.
E. Expression of the Bmp feedback inhibitor sizzled (szl) in ventral ectoderm (anterior up) at the end of gastrulation. Expression of szl marks cells experiencing active Bmp signaling. Injection of apcdd1l mRNA into wild-type embryos at the one-cell stage results in downregulation of szl by the end of gastrulation.
F. Expression of szl in ventral ectoderm (anterior up) at the 10-somite stage. apcdd1l-morphants and MZmutants exhibit an expanded ventral domain of szl, indicating that Bmp signaling is now higher than normal. Treatment of embryos from 10 hpf with 75 µM dorsomorphin (DM) abolishes szl expression in control embryos and partially reverses expansion of szl expression in apcdd1l-morphants.
G, H. Expression of gata3 in ventral epidermis ionocytes (arrows) at 24 hpf (G) and gata1 in blood progenitors (arrows) at 27 hpf (H). Ventral ionocytes and blood progenitors are expanded in apcdd1l-morphants, indicating ventralization of caudal structures. MZapcdd1l mutants also show an expanded domain of gata1. Treatment of apcdd1l-morphants with 75 µM DM from the end of gastrulation partially reverses ventralization. Images show lateral views with anterior to the left.
We hypothesized that the rapid shift from a dorsalized to ventralized phenotype resulted from rising levels of BMP activity near the end of gastrulation due to disruption of Apcdd1l-mediated BMP antagonism. In support of this hypothesis, the expression of bmp4 was upregulated by the end of gastrulation in apcdd1l morphants (Fig. S4G). Additionally, expression of sizzled (Salic et al., 1997), a direct feedback inhibitor of BMP signaling (Yabe et al., 2003) began to be upregulated and expand by 8 hpf in apcdd1l morphants, several hours before the shift in dorsal-ventral (DV) patterning. Sizzled expression remained expanded through at least 14 hpf (10 somites stage) (Fig. 2F). In contrast, injection of wild-type apcdd1l mRNA strongly reduced sizzled expression by 10 hpf (Fig. 2E). Finally, treating apcdd1l morphants at 10 hpf with dorsomorphin (DM), a pharmacological inhibitor of BMP signaling (Kwon et al., 2010; Yu et al., 2008), strongly reduced expression of sizzled at 14 hpf (Fig. 2F) and restored normal expression of gata1 and gata3 in ventral tissues (Fig. 2G, H). These data support the hypothesis that zebrafish Apcdd1l is required near the end of gastrulation to limit BMP activity.
To validate the morpholino knockdown phenotype, we used TALENs to target exon 1 near the translation start site and recovered a knockout allele (termed x60) with a 7-nucleotide deletion (Fig. 2A). The mutant allele leads to a frame shift and a series of premature stops, likely a null. Subsequent breeding showed that the apcdd1l mutation is homozygous-viable, enabling the generation of mutant embryos lacking both maternal and zygotic function (MZmutants). MZmutants appeared very similar to apcdd1l morphants, showing a markedly dorsalized phenotype during gastrulation (Fig. 2B, C). Dorsalization was later reversed (Fig. 2D) as MZmutants became mildly ventralized (Fig. 2H). Moreover, the ventral domain of sizzled expression was enlarged at 10 somites stage relative to wild-type embryos (Fig. 2F), though not to quite the extent of apcdd1l morphants. Thus, both mutant and morphant phenotypes support the role of Apcdd1l as a BMP antagonist in zebrafish.
Apcdd1 misexpression blocks BMP-dependent activities in the developing spinal cord
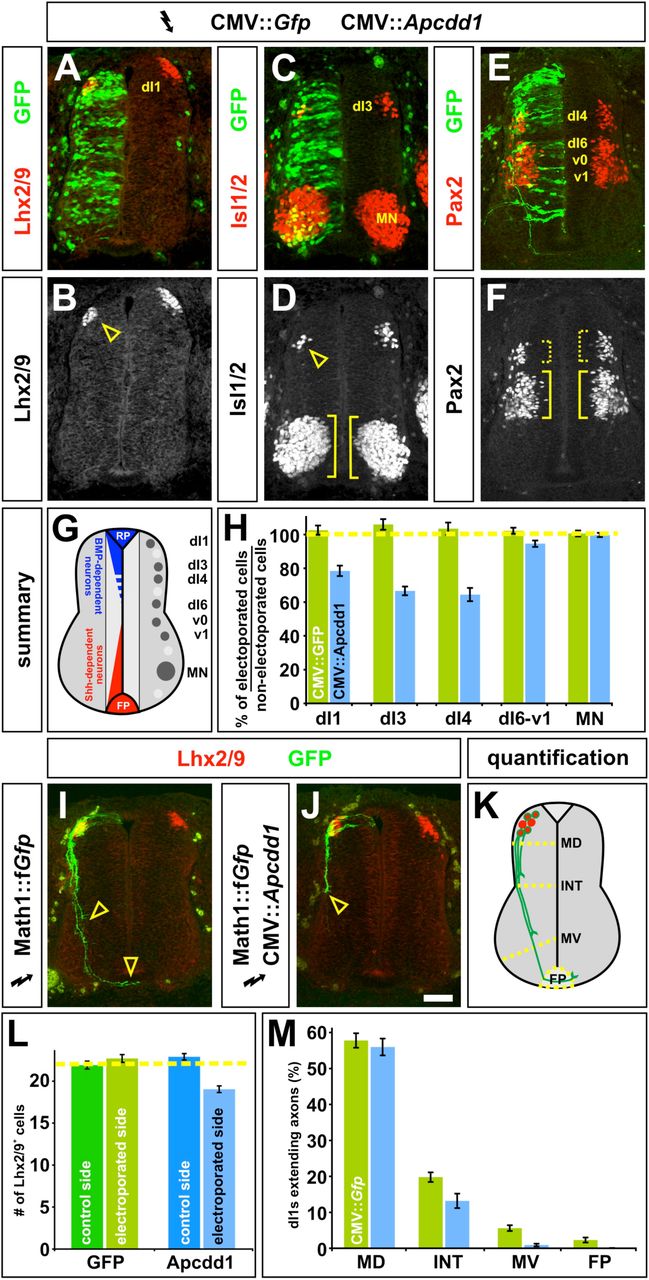
To test the ability of Apcdd1 to inhibit BMP signaling in a heterologous system and in later developmental contexts, we examined the effects of Apcdd1 mis-expression during chick embryogenesis. It has been previously shown that BMP signaling has multiple roles during the development of the vertebrate dorsal spinal cord. First, BMPs and activin act from the roof plate (RP) to direct the fate of dorsal interneurons (dI) 1 to dI3 (Chizhikov and Millen, 2005; Liem et al., 1997) (Fig 3G). The more ventral dorsal neurons form independently of RP signals (Lee et al., 2000; Timmer et al., 2002; Wine-Lee et al., 2004). Blocking BMP signaling using the inhibitory (I) Smad, Smad7, results in the loss of dI1 and dI3 (Hazen et al., 2011). Second, BMP signaling controls the rate of dI1 (commissural) axon growth (Hazen et al., 2012; Phan et al., 2010; Yamauchi et al., 2013). When BMP signaling is blocked by mis-expression of another I-Smad, Smad6, dI1 axon growth stalls (Hazen et al., 2011). While Apcdd1 is expressed in the anterior neural tube, it is only present in blood vessels at spinal levels (Fig. S5), making it unlikely that APCDD1 has an endogenous role controlling BMP signaling in the dorsal spinal cord. Nevertheless, the absence of Apcdd1 expression in the spinal cord provides an opportunity to examine whether APCDD1 has analogous activities to the I-Smads.

A-F, H. Gfp alone (control, H), or Gfp (green) in combination with Apcdd1 (experimental, A-F, H), was ectopically expressed throughout the spinal cord under the control of the CMV enhancer by in ovo electroporation at Hamilton Hamburger (HH) stages 14/15. Embryos were harvested at HH stage 23 and examined for the number of Lhx2/9+ dI1 (commissural) neurons (red, A, B), Isl1/2+ dI3 and motor neurons (MNs) (red, C, D) or Pax2+ dI4 and dI6-v1 neurons (red, E, F).
G. The relative positions of the labeled post-mitotic neurons are shown schematized within the spinal cord. The dorsal most populations of spinal neurons (dI1-dI3) are known to be dependent on BMP signaling, the role of BMP signaling in the more ventral-dorsal populations (dI4-dI6) has remained unresolved.
H. Quantification revealed that the misexpression of Gfp (n=42 sections from 6 embryos) had no significant effect on any of the monitored populations of spinal neurons (dI1, student’s t-test p>0.38 compared to the non-electroporated control side of the same embryo; dI3, p>0.19; dI4, p>0.40; dI6-v1, p>0.35; MN, p>0.49). In contrast, the ectopic expression of both Gfp and Apcdd1 (n=35 sections from 6 embryos) resulted in a 25-35% loss in BMP-dependent dorsal neural populations: dI1 neurons (p<8.1×10-8 different from the GFP+ control electroporation) dI3 neurons (p<1.8×10-9) and dI4 neurons (p<2.3×10-6). However, Apcdd1 had no detectable effect on the numbers of BMP-independent neurons, specifically dI6, vo, v1 interneurons (p>0.12) and MNs (p>0.49).
I. Lhx2/9+ dI1 neurons (red) electroporated with farnesylated (f) Gfp under the control of the Math1 (Atoh1) enhancer (Math1::fGfp) at HH stages 16/17, GFP+ axons (green) are extending into the ventral spinal cord by HH stage 22, with a few axons having reached the floor plate (FP, arrowheads).
J. In contrast, Lhx2/9+ dI1 neurons (red) concomitantly electroporated with both Math1::fGfp (green) and CMV:: Apcdd1 showed over an 80% reduction of axon outgrowth at the same stage, with no axons reaching the FP (arrowhead).
K. The extent of the dI1 axon outgrowth was quantified by determining whether dI1 axons had crossed one of four arbitrary lines in the spinal cord: mid-dorsal (MD), intermediate (INT), mid-ventral (MV) or the FP, as in (Phan et al., 2010).
L. Misexpression of Apcdd1 throughout the spinal cord at HH stage 16/17 has a small but significant effect on the numbers of dI1 neurons. There is a 10 % reduction in dI1 neurons on the Apcdd1 electroporated side (p<2.2×10-9, n=173 sections from 6 embryos) compared to the control Gfp electroporated side (n=178 sections from 5 embryos).
M. By HH stage 22, approximately 55 % of Lhx2/9+ neurons electroporated with gfp (control) or gfp with Apcdd11 (experimental) have extended GFP+ axons. In control embryos (n=178 sections, taken from 5 embryos), 10% of these axons extended to the MV line and 4% reached the FP. In contrast, only 1.6% of Lhx2/9+ neurons in the embryos electroporated with both Gfp and Apcdd1 (n=173 sections, taken from 6 embryos) had extended to axons to the MV line (p<8.5×10-8 similar to GFP controls) with none reaching the FP (p<7.6×10-4). Scale bar: 25 μm.
To test this hypothesis, we ubiquitously mis-expressed Apcdd1 in chicken spinal cords using in ovo electroporation. When Apcdd1 was expressed at high levels immediately before neurogenesis commenced in the dorsal spinal cord (Hamilton Hamburger (HH) stages 10/12), a severe proliferation defect was observed throughout the spinal cord, as previously observed (Shimomura et al., 2010). However, if Apcdd1 1 was mis-expressed at lower levels while neurogenesis was ongoing (HH 14/15), a different phenotype was observed. There was a 25-35% loss of the dI1 (Fig. 3A, B, H), dI3 (Fig. 3C, D, H) and dI4 (Fig. 3E, F, H) neurons, a similar consequence to Smad7 mis-expression (Hazen et al., 2011). The ectopic expression of Apcdd1 had no detectable effect on the BMP-independent classes of spinal neurons, strongly suggesting that Apcdd1 specifically blocks the ability of BMP signaling to induce dorsal cell fates.
We next examined whether the presence of Apcdd1 affected dI1 axon growth by mis-expressing Apcdd1 at HH stages 16/17, as dI1 axons extend axons away from the dorsal midline (Fig. 3K). The Apcdd1+ GFP+ dI1 axons displayed significant defects in axon extension (Fig. 3J) compared to GFP+ control axons (Fig. 3I). We quantified axon outgrowth by determining whether dI1 axons had crossed one of four arbitrarily drawn lines in the spinal cord: mid-dorsal (MD), intermediate (INT), mid-ventral (MV) spinal cord, or the FP (Fig. 3K). By HH 22, similar numbers of control and experimental dI1s had extended axons (Fig 3M). 10% of control GFP+ axons then projected into the ventral spinal cord with 4% reaching the FP. In striking contrast, only 1.6% of Apcdd1+ dI1 neurons extended axons to the MV line and none reached the FP, a phenotype similar to that observed after the mis-expression of Smad6 (Hazen et al., 2011).
Apcdd1 is expressed in hair follicles
Finally, we characterized the putative role of Apcdd1 as a dual BMP/Wnt inhibitor in mammals. We previously reported the localization of APCDD1 in the dermal papilla (DP), matrix (Mx), hair shaft cortex (HSCx), and cuticle (HSCu) of the human hair follicle (Shimomura et al., 2010). Here, we examined the expression of Apcdd1 in the skin and hair of adult mice. We found that Apcdd1 is expressed in the mature keratinocytes of the inner root sheet (Fig. 4A-C), consistent with its role in downregulating both Wnt signaling (Shimomura et al., 2010) and BMP signaling. While previous experiments in mouse revealed coordinated and periodic asynchronous BMP and Wnt signaling activation in mouse hair follicles ((Kandyba et al., 2013; Plikus et al., 2011; Plikus et al., 2008), the molecular nature of the coordination of temporal dynamics remained elusive. Since Apcdd1 is expressed at the right time and place in the hair follicles, we set out to determine if it may control such dynamics. Thus, to gain quantitative insight into the manner in which BMP and Wnt pathways are coordinated in the hair follicle, we developed a model with coupled differential equations, whose variables represent experimentally measurable BMP and Wnt signaling components, including Apcdd1. As in similar genetic circuits (Suel et al., 2007), equations represent known interactions between circuit components and use kinetic parameters inferred from our experiments. First, the model reproduces our data and predicts the activation of two BMP and Wnt pathway outputs, pSmad1 and β-catenin (Fig. S6), as a function of BMP and Wnt inputs (Kandyba et al., 2013; Miyoshi et al., 2012; Song et al., 2014). Second, we modelled cases relevant to hair follicle dynamics (Fig. 4D), where BMP and Wnt ligand inputs are constant or periodic, and predict that interesting temporal dynamics may emerge if ligands are periodic, as happens in in vivo in hair and skin (Kandyba et al., 2013; Plikus et al., 2011; Plikus et al., 2008). When BMP and Wnt oscillate out of phase, as in vivo in mouse hair follicles (Plikus et al., 2011), the presence of Apcdd1 should shorten the time when BMP effector pSmad1 is at high levels (Fig. 4D).

A. Immunofluorescence staining of adult mouse hair follicle cryosections detects expression of Apcdd1 (upper row) and respectively pSmad1/5 (lower row) in relation to hair follicle matrix marker P-cadherin and proliferation marker Ki67. The two sections are 20 microns apart. Nuclei are visualized with DAPI.
B. In mouse hair follicles paraffin sections, activated β-catenin (brown) is present in the hair matrix abutting the dermal papilla (DP) and in the outer root sheet (ORS). Scale bar: 50 μm.
C. Schematic of immunodetection results. Activated β-catenin (brown), an effector of Wnt signaling, is localized along the outer root sheath (ORS) and hair bulb, surrounding the dermal papilla (DP) that produces the growth signals during anagen. Nuclear pSMAD-1/5 (yellow), an effector of BMP signaling, is located in the distal portion of the hair matrix, where keratinocytes begin to differentiate into the layers of the inner root sheath (IRS). Apcdd1 expression (green) is localized to the mature keratinocytes of the IRS, consistent with its function to downregulate both Wnt and BMP signaling.
D. Mathematical model of the hair follicle temporal activation dynamics of the BMP and Wnt pathways (pSmad1 in blue, β-catenin in red). The BMP and Wnt ligand inputs are periodic, have the same period and are out of phase by a quarter of a period. Cases shown: (i) no Apcdd1 is present, as in null mutants; (ii) mutant Apcdd1, which is a very weak inhibitor (Shimomura et al., 2010); (iii) wild type Apcdd1, which inhibits Wnt signaling (Shimomura et al., 2010) and BMP signaling; (iv) wild type Apcdd11, and added moderate repression of Wnt signaling by Bmp signaling; (v) wild type Apcdd1, added moderate repression of Wnt signaling by Bmp signaling, and Apcdd1 is induced by Wnt signaling (Fig. 5). The absence of APCDD1 allows higher levels of pSmad1; wild type Apcdd1 depresses pSmad1 levels.
E. Apcdd1 controls the duration of the phases of the hair follicle cycle. Summary of Apcdd1 activity: First, Apcdd1 expression is induced by Wnt signaling and also by a currently unidentified factor X, which maintains Apcdd1 after Wnt signaling ceases. Second, Apcdd1 represses both Bmp signaling and Wnt signaling (which also is repressed by Bmp signaling). Bmp and Wnt signaling together control the duration of hair follicle phases anagen (when hair grows) and telogen (when hair falls out). Third, mutations inactivating Wnt signaling, or Bmp signaling, or Apcdd1, result in changed durations of anagen and telogen.
Apcdd1 has an evolutionarily novel protein domain
To understand the origin of Apcdd1 and find whether it has protein domains, we mapped its evolutionary emergence and analyzed its predicted protein structural features. First, we found Apcdd1 is present in only 10 of the 35 animal phyla (Fig. 5A). Apcdd1 is expanded in 2 phyla (Chordates and Arthropods) and is syntenic within, but not between, phyla (data not shown). While Apcdd1 appears in Metazoans but not in fungi or plants (Fig. 5A), we did find a 205 amino-acid protein resembling human APCDD1 in a few bacterial species. Apcdd1 is similar to orthologous genes in several other genomes, but to no other genes, suggesting Apcdd1 is a taxon-restricted gene that may have arisen at the origin of Metazoa. Interestingly, Apcdd1 was maintained in a few metazoan phyla where it acquired useful functions in Wnt and BMP signaling, which in these phyla control axial specification during embryonic development (Angerer et al., 2000; Darras et al., 2018; Guder et al., 2006; Henry et al., 2010; Henry et al., 2008; Hogvall et al., 2014; Kuo and Weisblat, 2011; Lambert et al., 2016; Lowe et al., 2006; Marlow et al., 2014; Molina et al., 2011; Pang et al., 2011; Pang et al., 2010; Park and Priess, 2003; Plickert et al., 2006; Rentzsch et al., 2007; Schenkelaars et al., 2017; Scimone et al., 2016; Suzuki et al., 1999; Treffkorn and Mayer, 2013; Wikramanayake et al., 1998; Windsor and Leys, 2010) and was lost in phyla that do not utilize Bmp for axial specification have lost APCDD1 (Fig. 5A, Supp. Table 2). Second, we examined the secondary structure content (Yachdav et al., 2014) of 441 Apcdd1 protein sequences (Fig. S7). We found low disorder, and 45% secondary structure elements (16 % α-helix, 29% β-strand). The β-strand content of Apcdd1 proteins is higher than that of ancient proteins (Schaefer et al., 2010) but similar to that of new proteins (Abrusan, 2013). By aligning multiple Apcdd1 sequences, we found that Apcdd1 proteins have two ∼210 amino acid novel domains that resemble each other, are present only in Apcdd1 proteins in different species, and likely arose by internal duplication (Fig. 5B). The Pfam database (El-Gebali et al., 2019) has also recently annotated this domain as Apcddc. Since the signal peptide and the transmembrane domain are not present in all Apcdd1 proteins, Apcdd1 subcellular localization may vary between species. Thus Apcdd1 is a taxon-restricted gene that encodes a protein with a novel protein domain and inhibits BMP and Wnt signaling in organisms that use BMP and Wnt for embryonic axial specification.

A. Apcdd1 is present in 9 of 35 Metazoan phyla. Species divergence tree in animal phyla having at least one fully sequenced genome. TimeTree.org was used to generate the phylogeny by specifying Metazoa (group) and phylum (rank), and displaying time (MYA) (Kumar et al., 2017). Solid circles denote nodes that mapped directly to the NCBI taxonomy; open circles denote nameless nodes that were not listed as higher-level taxa in the Taxonomy browser. The presence of the Apcdd1 gene in at least one species of the phylum is indicated by a black box, while the absence of Apcdd1 is indicated by a white box. The most parsimonious explanation suggests an early Metazoan origin of the Apcdd1 gene, followed by repeated gene loss in multiple phyla.
B. Apcdd1 protein sequence divergence tree. 30 protein sequences similar to human Apcdd1 were used to build a sequence divergence tree. Consistent with many phylogenomics studies, the topology of the gene tree is similar but not identical to that of a species tree (Edwards et al., 2007). The presence of the Apcddc domains is indicated by the pink and green boxes. APCDD1 has two novel Apcddc domains (positions in human Apcdd1 protein: pink, amino acids 51-282; green, amino-acids 284-473) that resemble each other and may be an internal duplication. Some Apcdd1 proteins are shorter and have only one domain (Hemichordate phylum).
Discussion
Here we report that Apcdd1 is a potent inhibitor of BMP signaling. We show that Apcdd1 blocks BMP signaling by binding to the type I BMP receptor, BmprIa, inhibiting the nuclear localization of pSmad1, the primary BMP effector. Overexpression of Apcdd1 represses the expression of BMP targets, and in Xenopus, chicken, and zebrafish embryos, interferes with major developmental processes including gastrulation, body axis formation, neural specification, and axon pathfinding. We previously showed that Apcdd1 also blocks canonical Wnt signaling by binding Wnt ligands as well as the LRP5 coreceptor. Thus, Apcdd1 provides a nexus for co-inhibition of both Bmp and Wnt signaling pathways. Protein expression and mathematical modeling of Apcdd1 inhibition of BMP and Wnt in hair follicles are consistent with the observed Wnt/BMP periodic sequential activation in mouse hair cycle (Lim and Nusse; Plikus et al., 2011) and with the complementary agent-based computational modeling that accompanied those experiments (Kandyba et al., 2013; Plikus et al., 2011; Plikus et al., 2008). Thus, Apcdd1 controls the duration of the phases of the hair follicle cycle by coordinating the timing of Wnt and BMP signaling activity (Fig. 5). These findings have a number of functional and evolutionary implications, as discussed below.
Only a few dual pathway inhibitors are known, and one (Sizzled) does not appear to be a bona fide dual Wnt/BMP inhibitor. While the ability to block both BMP and Wnt is a relatively novel feature of Apcdd1, other dual function inhibitors have been described, such as Cerberus, DAN, Gremlin, Coco,Wise/USAG-1 and Sclerostin, which are members of the CAN family of secreted peptides (Cruciat and Niehrs, 2013; Mulloy and Rider, 2015) that block BMP signaling via direct ligand-binding. In Xenopus, Cerberus also binds Wnt ligand, although this function is not conserved in mammals or birds. Of the CAN family, only Wise/USAG-1 and Sclerostin retain the ability to block both Wnt and BMP. Also, of note are members of the secreted Frizzled (sFRP) family of secreted peptides, which are related to the Wnt receptor Frizzled. Most sFRPs block Wnt signaling by direct ligand-binding (Cruciat and Niehrs, 2013). However, Sizzled represents a divergent sFRP member that blocks Bmp signaling. Sizzled, initially described as a Wnt antagonist (Salic et al., 1997), does not block Wnt but instead blocks BMP by stabilizing the Bmp antagonist Chordin (Bijakowski et al., 2012; Bradley et al., 2000; Bu et al., 2017; Lee et al., 2006; Muraoka et al., 2006; Yabe et al., 2003). Although Sizzled can no longer be considered a dual function inhibitor, its evolutionary derivation from sFRPs reflects an ancient relationship between BMP and Wnt regulatory mechanisms.
In contrast to the CAN and sFRP families, Apcdd1 stands alone as a new gene: unlike other inhibitors, Apcdd1 is not related to BMP or Wnt receptors or to any other gene, and has a highly conserved dual function as an inhibitor of both Wnt and BMP signaling. Apcdd1 is a taxon-restricted gene present in a subset of Metazoan phyla, and contains a novel protein domain not found in other proteins, which leaves open the possibility that this gene arose not by copying other genes, but de novo or from long non-coding RNA (Carvunis et al., 2012; Tautz and Domazet-Loso, 2011). A similar example of a new gene arising by non-copying mechanisms at the origin of Metazoans and having important functions is the novel protein-coding gene Percc1 (Oz-Levi et al., 2019). Apcdd1 appears to have evolved at roughly the same time as the BMP and Wnt signaling pathways, near the beginning of Metazoan evolution. Subsequently Apcdd1 has been lost in a number of metazoan phyla. However, it is noteworthy that the Apcdd1 has been maintained in phyla in which both BMP and Wnt play key roles in establishing primary embryonic axes, whereas phyla do not utilize BMP for axial specification have lost Apcdd1 (Fig. 5A, Table S2). While this correlation warrants further investigation, our analysis of Apcdd1 sequence and function illuminates how a taxon-restricted gene has been co-opted to coordinate the functional output of two major intercellular signaling pathways.
Materials and Methods
Xenopus laevis microarrays
4-cell stage embryos were injected marginally in each of the dorsal blastomeres with 10ng anti-apcdd1 MO or control MO. Apcdd1-depleted embryos were also labeled on the ventral side with Nile Blue crystals, because some of the dorsally-depleted embryos displayed a delay in dorsal lip formation. At stage 10.5, when the dorsal lip is clearly marked, the vitelline membrane was removed with tweezers and the dorsal half sectioned with a hair knife(Vonica et al., 2011). 10 embryos were prepared for each condition (control and depletion) in three different experiments. RNA from each batch was purified by Trizol (Ambion) extraction and used for cRNA synthesis with the Gen Chip 3’IVT Express Kit (Affymetrix). Microarray (Xenopus laevis 2.0 Affymetrix Array, GPL10756) hybridization and data collection was performed by the Rockefeller University Genomics Center, and the data was analyzed using GeneSpring software. Complete microarray data was deposited in GEO.
Xenopus gene expression studies
Xenopus Apcdd1 cDNA was injected at the one-cell stage at a concentration of 150 ng/μl.
To evaluate the effect of Apcdd1 on protein levels of TGFβ receptors, Xenopus embryos were injected in the animal poles of all blastomeres at-4 cell stage with RNAs for alk3-myc (100pg), bmprII-HA (200 pg), alk4-HA (250 pg), and LacZ (1 ng) as loading control, with or without apcdd1-FLAG (2 ng). Animal caps were collected at 4 cell stage in 1% NP40 buffer, deglycosylated, and run on NuPage 4-12% gels. Blots were incubated with the same antibodies as above, and with anti-beta-Gal chicken antibodies (1:5000), and visualized with ECL.
To evaluate the effect of Apcdd1 on endogenous C-terminal phosphorylation of Smad1, 1ng apcdd1 RNA was injected in the animal pole of all blastomeres at the 4-cell stage. Embryos injected with chordin RNA (1 ng) served as controls for the inhibition of BMP-dependent Smad1 phosphorylation. Animal caps were dissected at stage 11 and lysed in the presence of protein phosphatase inhibitor cocktails (Sigma). Western blots were stained with rabbit anti-Smad1 antibody (1:1000, Cell Signaling) and rabbit anti-pSmad1 (C-terminal) antibody (1:2000, from E. Laufer, Columbia U.).
Expression plasmids
p64T Xenopus CA-ALK3 (A. Suzuki, Hiroshima U.), pCS2+ Xenopus ALK4-HA (A.V), pCS2-Xnr1 (C. Wright, Vanderbilt U.), pCS2 BMP7 (A. Brivanlou, Rockefeller U.), pSP64T BMP4 and p64T BMP2 (G. Thomsen, SUNY Stony Brook), pCS2 (Xenopus) BMPRII-HA (M. Asashima, Tokyo U.), pcDNA3 (human) ALK3-Myc (E. Laufer, Columbia U. Coll. Phys. & Surg.), pSP35T Chordin (E. Amaya, U. Manchester), pGEM5Zf(-) Wnt8 (R. Harland, U.C. Berkeley), pCS2 apcdd1 (Xenopus) (A.V.), pBS APCDD1, APCDD1 FLAG, APCDD1 L9R, and pCXN2.1 APCDD1 FLAG (Y. Shimomura, Niigata U.).
Embryo manipulation, RNA in vitro transcription, and RNA in situ hybridization
Xenopus embryos were obtained by in vitro fertilization and cultured in standard solution (MMR). RNAs and MOs were injected on the dorsal side (bilaterally) or animal side (both ventral blastomeres for BMP transcription, all 4 cells for phospho Smad1 Western blot). For rescue of anterior structures, apcdd1 MO (20ng) were injected alone or together with 1 ng Bmp2, Bmp4, Bmp7, admp MO in the marginal zone of both dorsal blstomeres at the 4-cell stage and collected at stage 34. At the specified stages, according to Nieuwkoop and Faber, embryos were fixed for in situ hybridization, or extracts prepared for transcription or protein gels. RNA for injection and in situ hybridization was prepared with mMessage Machine with SP6, T7 or T3 RNA polymerases (Ambion), and T3 or T7 polymerase (Roche), for in situ probes with digoxigenin or FITC labeling mix (Roche), respectively. Morpholino oligonucleotides against Xenopus BMPs were: admp MO, 5’ – GGTCCATCTCATCAAGCTGCAGCTC-3’(Reversade and De Robertis, 2005), Bmp2 MO, 5′-GATCCCAGCGACC ATTGTCAACCTG-3′; Bmp4 MO, 5′-CAGCATTCGGTTACCAGGAATCATG-3′; Bmp7 MO, 5′-TTACTGTCAAAGCATTCATTTTGTC-3′(Reversade et al., 2005), and were co-injected with Apcdd1 MO at 1 ng /dorsal blastomere.
Transcription assays in embryos and cells
Xenopus embryos: transcription assays for the Wnt pathway were done as in (Shimomura et al., 2010), using the TOP-FLASH reporter. For BMP-dependent transcription in Apcdd1-depleted embryos, MO against Apcdd1 were injected in the animal poles at 2-cell stage (20 ng in each of 4 injections), followed by injection of BRE reporter (50 pg) and Bmp2 RNA (50 pg) in the animal poles of ventral blastomeres at 4-cell stage. For inhibition of Bmp activity by overexpressed Apcdd1, BRE reporter and Bmp2 RNA were coinjected with RNA of wild-type Apcdd1 or L9R mutant (Shimomura et al., 2010) Apcdd1 (1.5ng) in the animal poles of both ventral blastomeres at 4-cell stage. For constitutive Alk3 transcription, embryos were collected at stage 10.5, four embryos/assay, in triplicate, and each experiment was repeated a minimum of 3 times. Figures show a typical transcription assay for each experiment.
Cells were cultured in 24-well dishes, transfected with DNA (200 ng Bmp4 expression vector, 100 ng BRE reporter, 10 ng TK-Renilla control plasmid, 50 or 150 ng wild-type and L9R mutant APCDD1), and collected after 36 h. Luciferase activity was measured with the Dual Luciferase kit (Promega), in triplicates, and the experiment was repeated three times. The figure shows a typical experiment.
Luciferase transcription assays, in Xenopus and NIH 3T3 cells, for the Wnt pathway were carried out as in (Shimomura et al., 2010), using the TOP-FLASH reporter, while transcription assays for the BMP pathway used the BRE reporter activated by Bmp2 (Hata et al., 2000).
Cell culture immunoprecipitation analysis
CHO cells and NIH 3T3 cells were grown in standard growth medium and transfected with Fugene (Roche) or Lipofectamine 2000 (Life Sciences).
Plasmids expressing human Alk3 Myc (500ng), Bmrp2 HA (1µg), and human Apcdd1 FLAG (500pg) were transfected in CHO cells grown in 6-well plates. Cells were collected after 36 hours, lysed in 1% NP-40 buffer with added 1% Triton, 10% glycerol, 1mM EDTA, and protease inhibitors (Roche), and immunoprecipitated with either anti-FLAG (M2, Sigma), or anti-Myc beads (Millipore). After washes, immunoprecipitated receptors were deglycosylated with PNGase F (New England Biolabs), before resuspending in LDS sample buffer and running on NuPAGE 4-12% Bis Tris precast gels, using MOPS buffer (Invitrogen). Vector-transfected cells served as control. Proteins were transferred to PVDF membranes, incubated with mouse anti-HA (1:1000, Covance), rabbit anti-Myc (1:1000, Millipore), and rabbit anti-mouse Apcdd1 (1:5000, from Dritan Agalliu, UC Irvine), then with HRP-labeled secondary antibodies before staining with ECL (Amersham).
Immunofluorescence for BMP and Wnt signaling effectors in the presence of Apcdd1
NIH 3T3 were cultured in 8 well chamber slides with cover (LabTek II) transfected with empty vector or Apcdd1-eGFP. Cells were starved overnight in Optimem (Gibco) before addition of growth medium with or without recombinant signaling molecules (20 ng/ml human BMP2, 100 ng/ml Wnt3a, separately and together, R&D) for 2 h before fixation. Primary antibodies were mouse monoclonal anti-β-catenin (clone 5H10) and anti-Phospho-Smad1 (C-terminal) as above, and secondary antibodies were anti-mouse Alexa 647 and anti-rabbit Alexa 594 anti-rabbit antibodies (ThermoFisher).
The images were acquired with a confocal Zeiss LSM 510 confocal microscope. Images were acquired sequentially for each channel such that no image region was saturated or below detection The images were pseudocolored in Image J as follows: Apcdd1 in red, pSmad1 or β-catenin in green, DAPI (to visualize the nuclei) in blue. Channels were split in Image J and the level of pSmad1, β-Catenin, and DAPI were measured in the nucleus, whose contours were determined using DAPI. GFP levels were measured in the whole cell, using the β-catenin peri-membrane signal to determine the outline of the cell. For each of the eight groups of cells (two groups: transfected or not-transfected cells; for each group, there are 4 subgroups: no ligand, BMP2, Wnt3A, BMP2 + Wnt3A), at least 40 cells were imaged. For every cell, the background was determined by using the same mask to measure the signal in an empty area of the slide. The adjusted signal was reported as Sadjusted = (Scell -Sbackground)/Sbackground. Within the figures (Fig. 1A-B), individual channels were merged and displayed as follows: DAPI (blue), fluorescent protein reporter of Apcdd1 (red), pSmad1 (green, Fig. 1A), β-catenin (green, Fig. 1B). The images were pseudocolored in Image J as follows: APCDD1 in red, pSmad1 or β-catenin in green, DAPI (to visualize the nuclei) in blue.
Immunofluorescence for the effect of inducible expression of Apcdd1 on Alk3 expression in NIH 3T3 cells
The T-Rex system (Invitrogen) was used for tetracycline-dependent expression of human Apcdd1. NIH3T3 cells cultured in 8 well slides were transfected with 50 ng ALK3-Myc, 50 ng pcDNA 4/TO apcdd1, and 100 ng 6/TR (expressing the TET repressor) plasmids, with empty vectors, ALK3-Myc plasmid alone, and no-tetracycline wells as controls. Slides were fixed at time 0, and 1, 2, 4, 8, 12 h after tetracycline addition, stained with mouse anti-Myc (1:200) and rabbit anti-Apcdd1, secondary anti-mouse Alexa 488, anti-rabbit Alexa 594 (Life Technologies) and subjected to confocal microscopy.
Morpholino injections
To knockdown apcdd1 in zebrafish, embryos were co-injected with 10 ng/nl apcdd1 translation blocking morpholino (5’GAATAAGTCTCTCCCCAGCCATGAT3’) along with 5 ng/nl p53-MO, in a total volume of 1 nl/embryo. p53-MO was included to block non-specific cell death, though cell death was negligible even without p53-MO.
Pharmacological inhibition of Bmp
Dorsomorphin (Sigma, P5499) stock was dissolved in DMSO at a concentration of 10 mM. It was subsequently diluted to a concentration of 75 μM in embryo medium and added to zebrafish embryos at 10 hpf until fixation.
Apcdd1 probe synthesis
After extraction of total RNA, cDNA was synthesized using oligo dT primer using Invitrogen kit. Zebrafish apcdd1l cDNA was PCR amplified and cloned into pCS2+. The probe was subsequently synthesized using sense strand as a template.
Chicken immunohistochemistry
Antibody staining and RNA in situ hybridization histochemistry were performed on 15 μm transverse sections from Hamburger and Hamilton (HH) stage 14-24 (Hamburger and Hamilton, 1992) chicken spinal cords as previously described (Augsburger et al., 1999). Fluorescence images were collected on a Carl Zeiss LSM510 confocal and Axioplan 2 microscope. Images were processed using Adobe Photoshop CS4.
An RNA probe for in situ hybridization against the 3’ UTR of the chicken APCDD1 gene was prepared using the following primers: forward, 5’-GAG ATT AAC CCT CAC TAA AGG GAT GCT GCC TCA AAA ACA GAT G -3’, reverse 5’-CAG CCT TGA GGC CTT TAC TG -3’. The underlined region denotes a T3 polymerase site embedded in the primer sequence. The target sequence was amplified from HH stage 16/20/24 chicken spinal cord cDNA by PCR. Qiaquick (Qiagen) purified products were used in an in vitro transcription reaction using the Roche DIG RNA labeling kit.
Antibodies were used to detect several proteins: rabbit anti Lhx2/9 (pan Lh2a/b), 1:1000 (Liem et al., 1997); rabbit Islet1/2 (K5), 1:2000 (Tsuchida et al., 1994), rabbit anti-Pax2, 1:250 (Invitrogen); mouse anti-GFP at 1:1000 (3E6, Invitrogen). Species-appropriate Cyanine-3 and Fluorescein conjugated secondary antibodies were used (Jackson ImmunoResearch Laboratories).
Expression constructs and in ovo electroporation
Fertile White Leghorn eggs (McIntyre Poultry) were incubated to Hamilton and Hamburger (HH) stages 14/15 to examine cell fate specification phenotypes or stages 16/17 to axon outgrowth phenotypes. The following expression constructs were electroporated in ovo into the developing neural tube as described (Briscoe et al., 2000): CMV::gfp (pCIG, 0.5μg/μl), Math1::farnesylated (f) gfp (0.2μg/μl-0.5μg/μl), CMV::APCDD1 (0.2μg/μl) and the resulting eggs incubated until HH stages 22/23. Cell fate and dI1 axon phenotypes were quantified as described (Hazen et al., 2011; Phan et al., 2010), respectively. All statistical analyses were performed using a two-tailed Student’s t-test.
Statistical analysis
Using Excel, several parameters of the distributions of the adjusted fluorescence signals were computed for each condition (mean, median, mode, standard deviation, coefficient of variation, standard error of the mean, skew of the distribution, kurtosis). Since the distributions were essentially normal, the different conditions were compared using Student’s two-tailed t-test, adjusting the p-values for multiple comparisons using the Bonferroni correction.
Hair follicle immunochemistry and immunofluorescence
Swiss-Webster mice were selected to have a clear view of the hair matrix without melanin pigments. Mice were sacrificed at P31, during mid-anagen, and hair follicles examined were in Anagen V-VI, according to previous criteria (Muller-Rover et al., 2001). Dorsal skins of the mice were prepared for frozen sections and paraffin-embedded sections. Frozen sections 8 µm thick were post-fixed with 4% paraformaldehyde, blocked in 2% fish-skin gelatin, and incubated with primary antibodies to APCDD1 (1:1000, gift of Dr. Dritan Agalliu), pSMAD-1/5 (1:250, gift of Drs. Thomas Jessell and Ed Laufer), P-cadherin (1:500, Invitrogen 13-2000Z) and Ki67 (1:250, Santa Cruz sc-7846) at 4°C overnight, and counterstained with secondary antibodies. Paraffin-embedded sections were rehydrated in graded ethanol, subject to sodium citrate antigen retrieval, peroxidase blocking with 3% hydrogen peroxide, and incubated with primary antibody to β-catenin (1:100, BD Transduction 610153) at 4°C overnight, followed by a HRP-conjugated anti-mouse secondary antibody (Vector Labs, MP-7500), and developed with ImmPACT NovaRED kit (Vector Labs, SK-4805) according to manufacturer’s instructions. Immunofluorescence results of frozen sections were viewed on a Carl Zeiss LSR Confocal Microscope, and all bright-field images of paraffin-stained sections were obtained a Zeiss Axioplan2 Microscope.
Mathematical modeling of signaling pathway activation. Short description of the mathematical model
The model applies to the dynamics of BMP and Wnt signaling within hair follicles. In adult hair follicles, BMP and Wnt signaling are coordinated, are activated with the same period length, and are out of phase (Kandyba et al., 2013; Plikus et al.; Plikus et al., 2008). The coordination of BMP and Wnt activation dynamics in adult mouse hair follicles has previously been modeled with an agent-based model based on cellular states and not on the proteins involved in the process (Kandyba et al., 2013; Plikus et al.; Plikus et al., 2008). Our model of the same process in hair follicles takes a complementary approach, since it is a numerical model based on coupled differential equations used to evaluate protein level dynamics. “It assumes Michaelis-Menten kinetics, following a framework that is commonly used in the context of signaling and metabolic pathways (see, for example, Sections 1.5 and 7.2 of (Keener and Sneyd, 2009), and (Suel et al., 2007)).”
Both BMP and Wnt are treated as model inputs, denoted by f and g (respectively) in the model equations, and measured in arbitrary units. We consider four cases according to whether the inputs are constant or time-dependent, periodic functions:
The two inputs BMP and Wnt are both constant and continuous.
The two inputs BMP and Wnt are both periodic (either in-phase or out-of-phase by 1/4 of the period).
BMP is constant, Wnt is periodic.
BMP is periodic, Wnt is constant.
In Fig. 4D, we show configuration II, in which BMP and Wnt ligand inputs are both periodic and out-of-phase by 1/4 of the period.
We make the following notations: y1=APCDD1, y2=BmpR-total, y3=BmpR-activated, y4=pSmad1, y5=WntR-activated, y6=β-catenin, f=Bmp, g=Wnt (f and g are inputs), c=WntR-total. We are considering the possibility that the BMP pathway may inhibit the Wnt pathway.
We plot pSmad1 (in blue) and β-catenin (in red) against time. In Fig. 4D, the plots correspond to the following 5 cases:
Case i, no inhibition coming from APCDD1, as in null mutants;
Case ii, low inhibition coming from APCDD1, as in mutant APCDD1;
Case iii, full inhibition coming from APCDD1, as in wild type APCDD1;
Case iv, full inhibition coming from APCDD1, as in wild type APCDD1; additionally, there is repression of the Wnt pathway by the BMP pathway.
Case v, full inhibition coming from APCDD1, as in wild type APCDD1; additionally, there is repression of the Wnt pathway by the BMP pathway; in this case, APCDD1 is also induced by Wnt signaling.
Here are the parameters that we use:
η= Gaussian noise with standard deviation 2 ×10-4 and η’= Gaussian noise with standard deviation 2×10-5,
basic levels c=0.035, c1=0.016, c2=0.0054, c3=0.0001, c4=0.004,
parameters a1, a2, a3 are coefficients for various activating terms as follows: a1=6, a2=5, a3=4,
parameters b and b1 are coefficients for various inhibiting terms as follows: b=0.1 and b1=0.0003 for low inhibition (case ii), b1=0.0008 for full inhibition (cases iii, iv,v).
For Figure 4D, the equations for the Case i are:
 The equations for the Figure 4D Cases ii, iii are:
The equations for the Figure 4D Cases ii, iii are:
 The equations for the Figure 4D Case iv are the same as in the previous case, except for the last line, which reads:
The equations for the Figure 4D Case iv are the same as in the previous case, except for the last line, which reads:
 The equations for the Figure 4D Case v are the same as in the previous case, except for the first line, which reads:
The equations for the Figure 4D Case v are the same as in the previous case, except for the first line, which reads:
 We note that we studied many more formal cases which are not shown here.
We note that we studied many more formal cases which are not shown here.
In Fig. S6, similarly, we plot pSmad1 (in blue) and β-Catenin (in red) against time. The plots correspond to the following 2 cases:
In Fig. S6, Case A (left column in Fig. S6A) b1=b2=b=0, no inhibition coming from APCDD1, with mutual activation between the pathways;
In Fig. S6, Case B (right column in Fig. S6A) APCDD1 is produced by the Wnt branch and also added exogenously, full inhibition from APCDD1, with mutual activation between the pathways.
We note that we studied many more formal cases which are not shown here.
Here are the parameters we use:
basic level a0=10-3 and η= Gaussian noise with standard deviation 2·10-4
basic level a’0=10-4 and η’= Gaussian noise with standard deviation 2 ·10-5
basic level a’’0=8·10-4 and η’’= Gaussian noise with standard deviation 10-4
parameters K and a1 up to a6 are coefficients for various activating terms as follows: a1=6, a21=4, a25=7, a26=1, a3=0.015, a4=0.0053, a5=0.6, a6=0.2, b=0.1, K=0.035
the parameters b1 and b2 are coefficients for inhibition terms (b1, b2) = (0.0006, 0.0003).
The equations for In Fig. S6, Case A are:
 The equations for In Fig. S6, Case B are:
The equations for In Fig. S6, Case B are:

Apcdd1 synteny, detection of conserved domains, and gene divergence analyses
For synteny analyses, within genomes with Apcdd1 annotated, we looked at the nearest three upstream gene neighbors and three downstream gene neighbors using the UCSC Genome Browser and NCBI. To determine whether conserved domains exist within Apcdd1 proteins, we performed BLASTP queries (Altschul et al., 1990) using human APCDD1 with an E-value of 10-20. During alignment, we noted the presence of a novel domain, which was further analyzed with HHrepID (Biegert and Soding, 2008). Consensus Apcdd1 domain sequences for mammals were obtained by aligning the mammalian sequences with CLUSTALW in MEGA 5 (Tamura et al., 2011) followed by WebLogo 3 analysis (Crooks et al., 2004) to obtain the consensus sequence. In some proteins, the domain was found to be duplicated. Further BLASTP searches with the domain 1 and domain 2 mammalian consensus sequences retrieved six bacterial sequences, including Stigmatella, Methanobacterium, and Spirosoma. We note that this domain has also been recently annotated by the Pfam (El-Gebali et al., 2019) database curators, who named it Apcddc. The Pfam Apcddc domain boundaries correspond to the boundaries we had previously identified. Moreover, using the Apcddc domain to query BLASTP and HMMER (Eddy, 1998) retrieves only Apcdd1 and Apcdd1-like genes, indicating this domain occurs only in Apcdd1 genes. Furthermore, a phylogenetic tree of all collected sequences was constructed using MEGA 5.21 (Tamura et al., 2011) with alignment of the protein sequences with CLUSTALW using standard parameters (gap opening penalty 10, extension 0.1). A Neighbor-Joining phylogenetic tree was constructed using the Maximum Composite Likelihood Model with pairwise deletion and bootstrapped 5000 times. The initial tree was used to distinguish Apcdd1 sequences (closest to human APCDD1) from Apcdd1-like sequences.
To detect remote similarity, Apcdd1 sequences were selected for further analysis and a subsequent alignment. A BLASTP query against the NCBI non-redundant database was used to retrieve Apcdd1 protein sequences that are similar to human APCDD1 with an E-value of 10-3, allowing 200,000 as the maximal number of hits. After removing isoforms, 441 sequences remained. Low-quality alignments were removed, including a small number of bacterial sequences.
Sequence alignments of the APCDD1 sequences were performed in 30 chordate species for the protein sequences most similar to human APCDD1. MUSCLE (Edgar, 2004) was used with ClustalW sequence weighting and UPGMB clustering. PartitionFinder2 (Lanfear et al., 2017) was used to identify the optimal partition scheme and evolutionary models. Using these results, RAxML (Stamatakis, 2014) was used to generate a maximum likelihood gene tree using the PROTGAMMAWAG substitution model and 1000 bootstraps. FigTree was used to visualize the gene tree (Fig. S6B).
Apcdd1 protein sequence and secondary structure analysis
To analyze the content of secondary structure elements within Apcdd1 proteins, we used bioinformatics algorithm collections PredictProtein (www.predictprotein.org) and HMMER (hmmer.org) (Eddy, 1998; Yachdav et al., 2014). Within 441 Apcdd1 proteins from different Metazoan species, we scored sequence features and predicted secondary structural properties according to over 25 criteria. These criteria include disorder, content of α-helices and β-strands, number of binding sites to other proteins or to DNA, subcellular localization, propensity to aggregate, propensity to form prions, presence of transmembrane domains, presence of signal peptides, presence of Pfam domains, amino acid composition.
Author contributions
A.V., V.L., and A.M.C. conceived of the project. A.V. performed biochemistry, frog, and cell culture experiments. K.P. and S.J.B. performed the chicken experiments. N.B., J.G., and B.R. performed zebrafish experiments. V.L. performed cell imaging experiments. V.L. and L.I. did the mathematical modeling. E.W. and G.M.D. performed hair follicle immunohistochemistry. A.H.O.L., V.L., A.K., J.W.C. and J.A.W. performed phylogenetic and protein domain analyses, and statistical analyses. V.L., S.J.B., J.A.W., B.R., A.V., and A.M.C. wrote the paper. All authors read and edited the manuscript.
Supplementary Data
Data S1. Supplementary Table 1 with array genes is presented separately as an Excel file.
The presence of the Apcdd1 gene correlates with the involvement of Bmp signaling in axial specification. The involvement of Wnt activity and Bmp activity is listed in Metazoan phyla.

A-T. RNA in situ hybridization for Xenopus Apcdd1. Maternal expression at 8 cell stage (A, B) is localized in the animal pole and is frequently unevenly distributed. In late blastulas (stage 9, D-F), the expression is animal and marginal (arrows in F, blastocoel in outlined in F). At the start of gastrulation (stage 10, G-I) Apcdd1 RNA is present in animal and marginal cells, including the dorsal lip with the Spemann organizer (arrow in H, I, blastocoel is outlined in I). In neurula embryos (stage 18, K-N) and tailbud (stage 22), expression is limited to diencephalon and midbrain precursors (J, K, M, N, Q), and the tailbud (K, M, O, P, Q). Arrowhead in the transversal section (R) indicates ependymal precursors. In tadpoles (stage 30, S) expression closely mirrors that in mouse embryos. D – diencephalon; M – midbrain; Ov – otic vesicle; T – tailbud; A – anus. Bracket in S indicates the branchial arches. For control in situ hybridization (C: 8-cell stage; T: tadpole), the RNA probe used was LacZ.
The scale bar in A is 0.3 mm (all embryos are shown at the same scale)

In situ hybridization of morpholino-depleted (A1MO), or Apcdd1 overexpressing embryos. Embryos were injected at the 4-cell stage dorsally (A-C, double in situ hybridization for chordin and LacZ RNA as injection marker, in red), or 1 cell stage (D-R). Control embryos were injected solely with LacZ RNA (1 ng).
A-F. Spemann organizer genes: chordin is not affected by Apcdd1 depletion (B) (n=18), but inhibited by Apcdd1 RNA (A1 RNA) overexpression (n=23), consistent with the inhibition of the maternal, dorsal Wnt pathway; gsc is unaffected by either treatment (n= 15 and 12, respectively, E, F).
G-L. The ventral marker and BMP target sizzled is expanded by Apcdd1 depletion (n=23, I, J), and restored by expression of a MO-resistant Apcdd1 RNA (n=21, K, L).
M-R. The early neural marker and negative BMP target sox2 is inhibited by Apcdd1 depletion (n=27, O, P, arrows in O indicate a hole in the ventral expression pattern), and partially restored by expression of Apcdd1 RNA (n=17, Q, R).

A. Microarray detection of genes dependent on Xenopus laevis Apcdd1 (XA1) in Xenopus embryos. Apcdd1 protein was depleted with morpholino oligonucleotides in the dorsal part of the embryo injected at the 4-cell stage, and dorsal tissue containing the Spemann organizer was recovered at stage 10. Extracted RNA was used to identify changes in gene expression versus control embryos in Affymetrix microarrays.
B. Protein coding genes with more than 2-fold variation in XA1-depleted embryos. Genes shown were verified by quantitative PCR. Red: BMP-regulated genes; black: Wnt-regulated genes.
C. Effect of human APCDD1 (hA1) on BMP activity induced by recombinant BMP4 in NIH3T3 cells. The L9R mutant form of APCDD1 is significantly less active.
D. Apcdd1 inhibits both the Wnt and BMP pathways induced by exogenous Wnt8 and Bmp2 RNA in Xenopus. The APCDD1-L9R mutant has a considerably reduced effect on both pathways.
E. Quantification of XA1 depletion and rescue experiment (Fig. 1 E-I) results in a percentage of embryos with a specific dorso-anterior index (DAI) (Kao and Elinson, 1988). Numbers on top indicate the average DAI for the batch of embryos.
F. Induction of BMP signaling by different BMP ligands is differentially suppressed by XA1.
G. The combined action of chordin and Xenopus Apcdd1 is sufficient to suppress BMP-dependent reporter gene activation by constitutively active form of the type I BMP receptor (caBmprIa).
H. Xenopus Apcdd1 is sufficient to reduce Smad1 C-terminal phosphorylation induced by endogenous BMPs in Xenopus animal caps. Chordin is a positive control for BMP inhibition.
I-L. Co-expression of type I BMP receptor BMPRIA and tetracyclin-inducible APCDD1 expression in CHO cells causes displacement of BMPRIA after 12 h.
Statistical significance: * p < 0.05; ** p < 0.01; *** p < 0.001, Student’s t test.
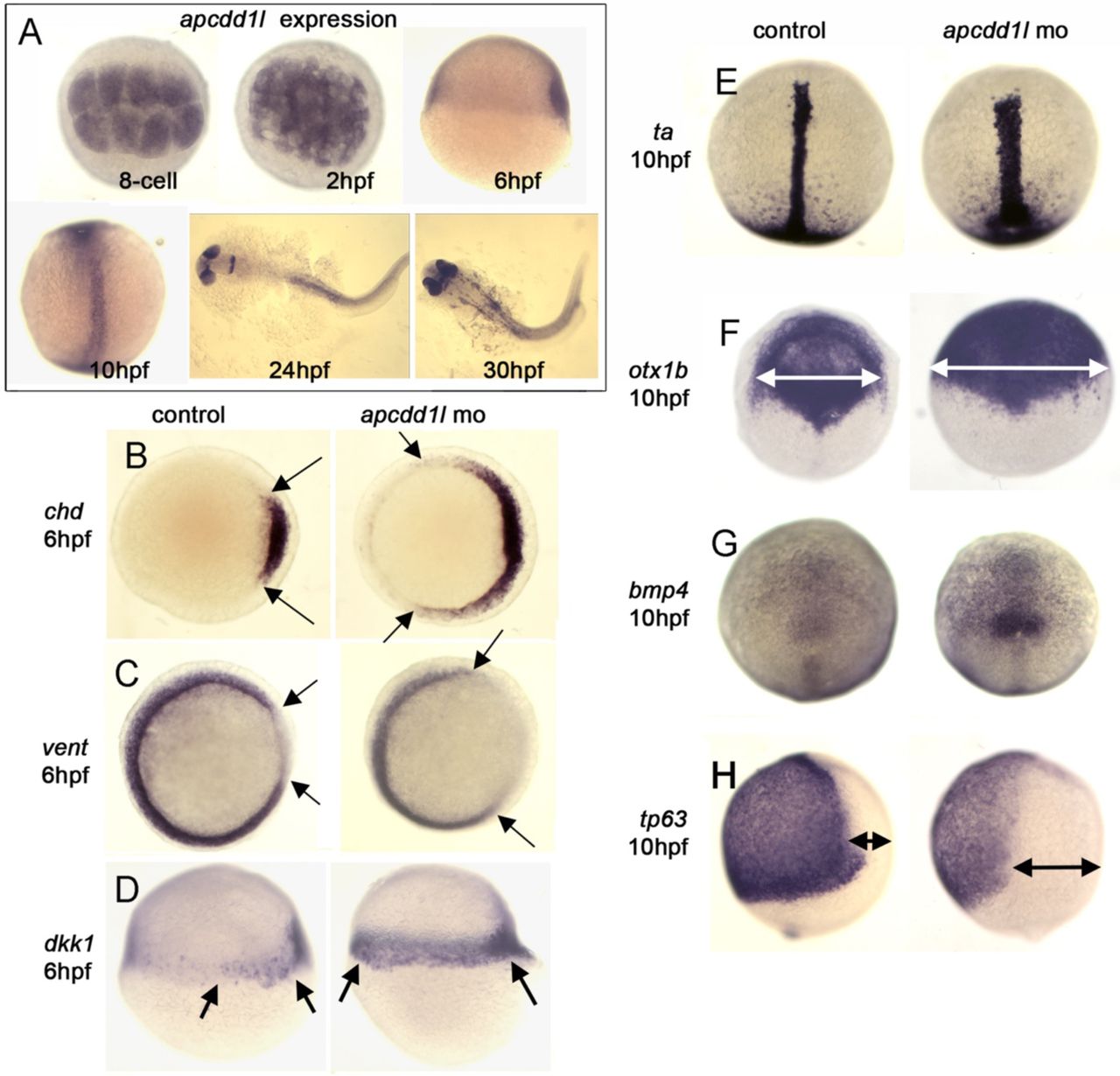
A. The presence of maternal apcdd1l mRNA, confirmed by RT-PCR (not shown), is seen in all cells in the early blastula (animal pole views). By early gastrulation (6 hpf), apcdd1l is only weakly expressed with preferential staining in dorsal tissues (lateral view, dorsal to the right). By the end of gastrulation (10 hpf), apcdd1l is expressed throughout to the dorsal axis (dorsal view and lateral view, anterior up). At 24 and 30 hpf, apcdd1l is expressed in the eyes, restricted regions in the brain, and dorsal tissues in the trunk and tail.
B-H. Domains of expression of various markers of gastrula stage embryos. Expression of chd in the organizer and vent in ventrolateral mesoderm (B, C, animal pole view) and dkk1 in the organizer (D, lateral view). The organizer is expanded laterally at 6 hpf in apcdd1l morphants, and there is a corresponding contraction of ventrolateral tissue. At 10 hpf, expression of brachyury-a (ta) in the notochord (E) and otx1 in the anterior neural plate (F) (dorsal views, anterior up) are expanded in apcdd1l morphants, whereas expression of ventral marker tp63 in epidermal ectoderm (H, lateral view) is contracted. At this time, expression of bmp4 at the ventral midline (G, dorsal view) begins to upregulate and expand in apcdd1l morphants, presaging subsequent ventralization.

A-C. In situ hybridization experiments for Apcdd1 on transverse sections of the spinal cord taken from HH stages 18 (A), 22 (B) and 26 (C) chicken embryos. Apcdd1 is expressed in the presumptive epidermis, as well as blood vessels within the spinal cord and a population of cells surrounding the spinal cord that have a distribution similar to migrating neural crest cells. (D, E) There is no alteration of Apdcc1 expression in the spinal cord after electroporation with either dominant negative (dn, D) or constitutively active (ca, E) BmprIb.
Scale bar: A: 50 μm, B-E: 100 μm.

Mathematical model of the temporal activation dynamics of the BMP and Wnt signaling pathways in NIH 3T3 cells. Cells were cultured in the presence of BMP and /or Wnt (as shown in Fig. 1A-C). We modeled the levels of BMP pathway effector pSmad1 (blue) and Wnt pathway effector β-Catenin (red) against time. Each row shows one case, and each case has two subcases: no APCDD1 is present (left), Apcdd1 is present (right).
Top row: when BMP ligand input and Wnt ligand input are both constant, results are consistent with observations (Fig. 1A-C).
Middle row: if BMP is constant and Wnt is periodic, the model predicts both pathway effectors become periodic.
Bottom row (middle row): if BMP and Wnt are both periodic and out of phase by ¼ of a period, the model predicts a complex periodic dynamics of effectors. In all three cases, the presence of Apcdd1 (right column) lowers the levels of both pSmad1 and β-Catenin compared to when it is absent (left column).
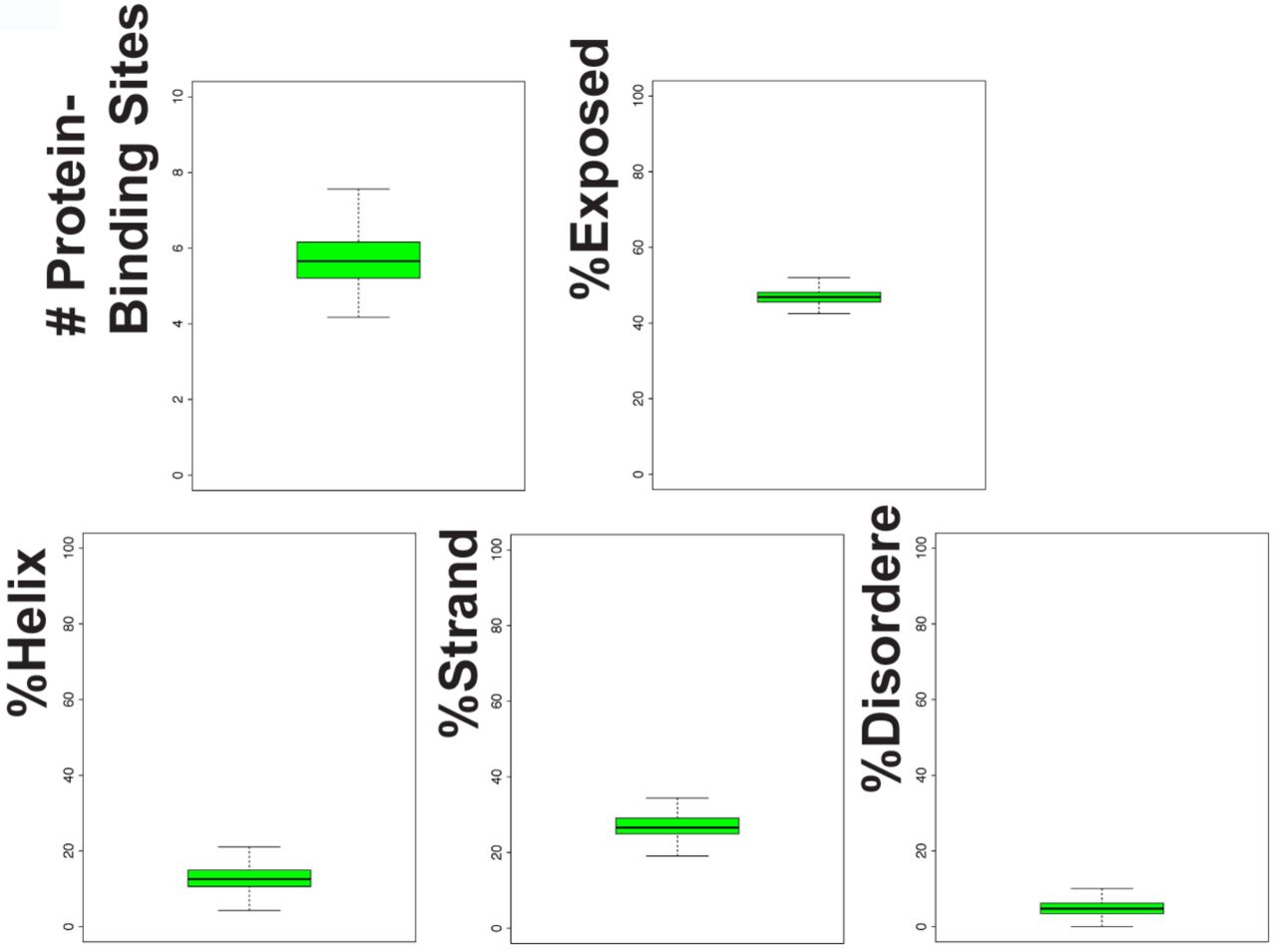
Secondary structure analysis of 441 Apcdd1 proteins shows the presence of alpha-helices, beta-strands, and protein-binding sites. Apcdd1 proteins have low disorder and low exposure to water.
Acknowledgments
V.L. and A.M.C. acknowledge funding support from NIH (grants: P30AR044535 to David Bickers, R01AR052579 to A.M.C). V.L. thanks Marc W. Kirschner and acknowledges funding support from National Institutes of Health (NIH grants R01 HD073104 and R01 HD091846, to M.W.K.). V.L. and L.I. acknowledge partial funding support from the Principal’s Interdisciplinary Fund of the University of Aberdeen and the National Science Foundation (grant number PHY11-25915 to U.C.S.B., K.I.T.P.). Funding support was provided to A.H.O.L. by the NIH National Child Health and Development Institute (K12 HD052896) and by a Boston Children’s Hospital Career Development Fellowship. A.V. was supported by NIH grant R03 HD057334 and is grateful to Dr. Jean Gautier at the Institute of Cancer Genetics, Columbia University in New York City, for access to the lab Xenopus facility. Zebrafish experiments were supported by NIH-NIDCD grant R01-DC03806. S.J.B. and K.P. acknowledge funding support from NIH/NIHDS (R01: NS085097) and from the UCLA Broad Stem Cell Research Center. V.L. thanks Wenzhe Ma and Harry Burgess for insightful comments. V.L. and A.M.C. thank Yutaka Shimomura for his kindness and inspiring style.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}