
Abstract
Plant pathogens deploy an array of virulence factors to suppress host defense and promote pathogenicity. Numerous strains of Pseudomonas syringae produce the phytotoxin coronatine (COR). A major aspect of COR function is its ability to mimic a bioactive jasmonic acid (JA) conjugate and thus target the JA-receptor COR-insensitive 1 (COI1). Biological activities of COR include stimulation of JA-signaling and consequent suppression of SA-dependent defense through antagonistic crosstalk, antagonism of stomatal closure to allow bacterial entry into the interior of plant leaves, contribution to chlorotic symptoms in infected plants, and suppression of plant cell wall defense through perturbation of secondary metabolism. Here, we review the virulence function of COR, including updates on these established activities as well as more recent findings revealing COI1-independent activity of COR and shedding light on cooperative or redundant defense suppression between COR and type III effector proteins.
Similar content being viewed by others


Introduction
Phytotoxins are microbe-produced secondary metabolites that interfere with and sometimes kill plant cells. They are either directly active or are produced as prototoxins that become activated by plant enzymes (Duke and Dayan 2011; Pruess et al. 1973; Uchytil and Durbin 1980). Pseudomonas syringae pathovars produce a repertoire of virulence effectors that are active inside plant cells, including numerous phytotoxins (Hogenhout et al. 2009). One class of effects mediated by phytotoxins is disruption of amino acid metabolism. For example, phaseolotoxin blocks the production of arginine by inhibiting ornithine transcarboxylase (Ferguson and Johnston 1980). Tabtoxin gets converted in planta to a glutamate analog that inhibits glutamine synthetase thus causing a buildup of ammonia and glutamine deficiency (Turner 1981; Uchytil and Durbin 1980). Other effects of phytotoxins are quite diverse and include perturbation of metabolism of lipids, sugars, and cell walls, synthesis of proteins and nucleic acids, membrane integrity and mitosis (Duke and Dayan 2011; Ferguson and Johnston 1980; Goudet et al. 1999; Pruess et al. 1973; Hoffman 1995; King and Calhoun 2009; Strobel et al. 1996; Thuleau et al. 1988; Walton 2006; Daub et al. 2005; Tanaka 1996). Another effect of phytotoxins is perturbation of hormone signaling. Phytohormones play key roles in a variety of physiologic and cellular processes, including numerous processes related to plant defense that have been extensively reviewed elsewhere (Bari and Jones 2009; Howe and Jander 2008; Katagiri and Tsuda 2010; Pieterse et al. 2009). While some phytotoxins likely perturb hormone signaling indirectly, COR directly engages JA-signal transduction proteins to co-opt hormone signaling.
COR is a polyketide phytotoxin produced by pathovars of P. syringae, including alisalensis, atropurpurea, glycinea, maculicola, morsprunorum, porri, and tomato (Bender et al. 1999; Gross and Loper 2009; Mitchell 1982; Mitchell et al. 1983; Preston 2000; Ullrich et al. 1993; Wiebe and Campbell 1993; Zhao et al. 2000; Cintas 2002) (for these and other bacterial strains discussed, refer to Table 1). Additionally, COR-analogs are produced by Xanthomonas campestris pv. phormiicolai (Tamura et al. 1992; Mitchell 1991). Consistent with infection of a diverse set of host plants by these pathovars and species, COR is a non-host specific toxin that causes diffuse chlorosis in a wide variety of plant species (Rohde et al. 1998; Brooks et al. 2004).
COR biosynthesis and structure
COR is composed of two moieties, the polyketide coronafacic acid (CFA) and coronamic acid (CMA) (Bender et al. 1999; Ichihara et al. 1977; Mitchell 1985; Parry et al. 1993). CMA is derived from L-alloisoleucine, a diastereomer of l-isoleucine activated by the nonribosomal peptide synthetase adenylation domain of CmaA (Rohde et al. 1998; Buell et al. 2003; Worley et al. 2013). CFA is synthesized from a cyclopentenone compound with subsequent modification carried out by genes of the cfa operon (Gross and Loper 2009). Coronafacate ligase, one of nine open reading frames within cfa operon, joins CFA and CMA with an amide linkage to form COR (Bender et al. 1993; Liyanage et al. 1995) (Fig. 1).

Roles of coronatine and type III effectors in modulating defense-related hormone signaling. (1) Roles of coronatine. Coronatine (COR) is composed of two moieties: coronafacic acid (CFA) and coronamic acid (CMA) (Bender et al. 1999). Once COR moves into the plant cell (presumably through diffusion), it activates JA-signaling through mimicking JA-amino acid conjugates such as (+)-7-JA-isoleucine (JA-Ile) shown in the model. COR is able to interact with SCFCOI1 receptor complex with modestly higher affinity than JA-Ile (Sheard et al. 2010; Katsir et al. 2008; Fonseca et al. 2009b). Like JA-Ile, COR serves as ‘molecular glue’ between the receptor complex SCFCOI1 and the negative regulator JAZ protein (Sheard et al. 2010), and triggers the degradation of JAZ through 26S proteosomal-mediated pathway (Chini et al. 2007; Thines et al. 2007). Upon JAZ degradation, positive regulator TFs (e.g. MYC2, bHLH, and MYBs) are released from suppression, and activate JA-responsive genes (Wasternack and Hause 2013). MYC2 also regulates several NAC TFs that suppress SA accumulation through regulating SA-biosynthesis gene ICS1 and SA modifying gene BSMT1. These NAC TFs were also found to be required for stomatal reopening induced by COR (Zheng et al. 2012). In return, SA-activated, cytosolic NPR1 monomers suppress the JA-signaling pathway. COR’s ability to contribute to chlorotic disease symptoms is also mediated through COI1 (Mecey et al. 2011). COR is able to suppress callose deposition through inhibiting an ET-dependent indole glucosinolate pathway where the role of COI1 is unknown (Geng et al. 2012; Millet et al. 2010). Perhaps the CMA moiety of COR mimics the ET precursor ACC, and interferes with ET production. Additionaly, COR perturbs auxin and ABA signaling which could potentially offset the restriction of bacterial growth caused by flg22-induced suppression of auxin signaling (Navarro et al. 2006) or ABA-induced stomatal closure (Melotto et al. 2006), respectively. Whether COI1 is engaged in auxin and/or ABA perturbation is unknown. 2) Roles of type III effectors. AvrB or COR, cooperatively with other T3Es and dependent on COI1, induce expression of an ET responsive factor—RAP2.6 (He et al. 2004). HopZ1a acetylates JAZ proteins, causing them to become destabilized dependent on COI1, and restores virulence to a cor- mutant of Pto DC3000 (Jiang et al. 2013). HopX1 directly destabilizes JAZ proteins without a requirement for COI1, likely via its cysteine protease activity, and restores virulence to a cor- mutant of Pto DC3000. HopX1 shares additional activities with COR, including reopening of stomata, causing plant cells to lose chlorophyll, and induction of chlorosis in susceptible plants (Gimenez-Ibanez et al. 2014). HopM1 affects SA-dependent secretory pathway through interacting with and degrading an ARF-GEF family protein involved in vesicle trafficking called AtMIN7 (Nomura et al. 2006). HopM1 is also functionally redundant with COR in suppressing an SA-independent defense sector of which the mechanism is unknown (Geng et al. 2012). Solid lines indicate established interactions. Question marks indicate unknown mechanisms. Hormone/coronatine/effector-specifc functions are color coded: bold orange lines coronatine-related functions, yellow lines JA-related functions, green lines SA-related functions, blue lines ET-related functions, black lines T3Es-related functions. Hormones are color coded, and indicated by solid circles. Type III effectors are color coded, and indicated by solid stars. Structural similarities between compounds are indicated by same color shading of the respective chemical structures
CFA and CMA are synthesized independently and the operons encoding the COR biosynthetic genes differ between P. syringae pv. tomato (Pto) strain DC3000 and P. syringae pv. glycinea (Pgl) strain 4180 (Worley et al. 2013; Sreedharan et al. 2006). The COR biosynthesis operons are encoded on a 90-kb plasmid in Pgl 4180 (Bender et al. 1993). On the other hand, the COR biosynthetic genes of Pto DC3000 exist within two distinct chromosomal clusters; the CFA operon is separated by ~26 kb of intervening DNA from the CMA biosynthesis genes and the adjacent genes regulating COR expression (Brooks et al. 2004). Biosynthesis of COR, as well as CFA and CMA, is thermo-regulated in Pgl 4180 and several other pathovars of P. syringae (Rohde et al. 1998). Consistent with the symptom development in infected plants, COR production is negligible at 30 °C and reaches maximal level at 18 °C (Bender 1999; Rohde et al. 1998). On the other hand, COR production is not thermo-regulated in Pto DC3000 and the production is much less in vitro (Braun et al. 2008; Weingart et al. 2004). This is due to the difference of a histidine protein kinase CorS between two strains (Braun et al. 2008; Smirnova et al. 2008; Weingart et al. 2004; Ullrich et al. 1995), although the specific mechanism is not yet clear.
COR both structurally and functionally mimics the most active isoleucine conjugate of JA (+)-7-iso-JA-Ile (JA-Ile) (Wasternack and Xie 2010; Fonseca et al. 2009b) (Fig. 1). The functional resemblance between COR and JA-Ile has been widely noted (Chini et al. 2007; Thines et al. 2007; Sheard et al. 2010; Glazebrook 2005; Gimenez-Ibanez and Solano 2013; Haider et al. 2000; Weiler et al. 1994) and is now demonstrated experimentally by solved crystal structures of each molecule in association with a COI1 (COR-insensitive 1) receptor complex (Sheard et al. 2010).
In addition to the proven ability of COR to mimic JA-Ile, similarity between the CMA moiety of COR and 1-aminocyclopropane-1-carboxylic acid (ACC) has been noted (Brooks et al. 2004) (Fig. 1). ACC, the rate-limiting precursor of ethylene (ET) biosynthesis in higher plants, and CMA each contain a cyclopropane ring. Although the individual moieties of COR (CMA and CFA) show very limited activity in plant tissues (Uppalapati et al. 2005), intact COR perturbs ET homeostasis or known outputs of ET-signaling (Kenyon and Turner 1992; Ferguson and Mitchell 1985; Geng et al. 2012; Millet et al. 2010) leading to the hypothesis that COR, through its CMA moiety, perturbs ET biosynthesis or signaling. However, since a direct effect of COR on ET-signaling, for example via mimicry of ACC by the CMA moiety, has not been demonstrated, the possibility that COR modulates ET-signaling indirectly cannot be ruled out.
Suppression of plant defense and promotion of disease symptoms by COR
COR activates JA-signaling by mimicking JA-Ile
COR makes multiple contributions to bacterial virulence, several of which are mediated via its ability to mimic bioactive jasmonates (Fig. 1). Jasmonates (JAs) are lipid-derived plant hormones that regulate a broad range of plant cellular and physiological responses to control plant growth and development, as well as responses to biotic and abiotic stresses (Wasternack and Hause 2013). The final step of converting JA to its active version is carried out by JAR1 (for this and other plant genes discussed, refer to Table 2). JAR1 is a jasmonate:amino acid synthetase that conjugates JA to several amino acids, notably creating bioactive JA-Ile (Staswick et al. 2002; Suza and Staswick 2008). Among biotic stress responses, JA-signaling typically is activated when plants are attacked by necrotrophic pathogens or herbivores (Hopke et al. 1994; Norman et al. 1999; Schenk et al. 2000; Stotz et al. 2000; Karban and Baldwin 1998; Pieterse et al. 2012).
Similar to signaling by other plant hormones such as auxin (Dharmasiri et al. 2005) and gibberellic acid (Yamaguchi 2008; Schwechheimer and Willige 2009), JA-signaling results from the proteasome-mediated removal of transcriptional repressors. In the case of JA-signaling, these repressors are called JAZ (containing Jasmonate ZIM domain) proteins (Chini et al. 2007; Thines et al. 2007). When levels of bioactive JA-Ile are low, JAZ proteins are stable and function as transcriptional repressors by physically interacting with a variety of transcription factors (TFs), including MYC2, a basic-helix loop helix TF that activates a significant proportion of JA-induced responses (Lorenzo et al. 2004; Dombrecht et al. 2007). Transcriptional repression by JAZ proteins occurs through recruitment of the general co-repressor TOPLESS (TPL), usually via the adapter protein Novel Interactor of JAZ (NINJA) (Pauwels et al. 2010; Pauwels and Goossens 2011; Shyu et al. 2012; Szemenyei et al. 2008).
The JAZ family in Arabidopsis has 12 members (Chini et al. 2007). In addition to MYC2, JAZ proteins also interact with two other MYC2-related bHLH TFs, which regulate overlapping as well as distinct responses with MYC2, and other bHLH/MYB transcriptional factors, like MYB75 and MYB21, which also regulate JA responses (Cheng et al. 2009; Qi et al. 2011; Song et al. 2011). Additionally, interactome and functional analyses indicate that JAZ proteins likely interact with a wide variety of TFs to regulate development and stress responses (Kazan and Manners 2012; Qi et al. 2011; Seo et al. 2011; Song et al. 2011). For example, Song et al. (2011) found that JAZ1, JAZ8 and JAZ11 interact with MYB21 and MYB24 in both yeast and planta to mediate JA-regulated development processes. Similarly, Seo et al. (2011) found that OsJAZ1 interact with OsbHLH148 to regulate JA-regulated drought stress in rice. Specificity of individual JAZ proteins for diverse transcription factors likely contributes to tuning the JA-response to specific contexts, such as in different cell-types, developmental stages, and stresses, via integration with other signaling pathways.
JA-signaling is activated when JAZ proteins are destabilized by proteasome-mediated degradation. A typical SCF ubiquitin ligase complex consists of an F-box protein in complex with SKP1 and a Cdc53 (Hershko and Ciechanover 1998; Deshaies 1999). In Arabidoposis, the F-box protein COI1 associates with Skp1-like proteins ASK1, ASK2 (Gray et al. 1999; Gray et al. 2001) and Cdc53-like protein AtCUL1 (del Pozo and Estelle 1999) to assemble the SCFCOI1 E3 ubiquitin ligase complex (Devoto et al. 2002; Xu et al. 2002). JA-Ile binding to co-receptor complexes composed of a JAZ protein and COI1 within SCFCOI1 triggers ubiquitination of the JAZ proteins. The resulting 26S proteasome-mediated degradation relieves JAZ-mediated transcriptional repression to activate JA-responsive gene expression (Fonseca et al. 2009a; Katsir et al. 2008). High affinity binding of COI1 complexes to JAZ proteins requires both JA-Ile (or COR) and inositol pentakisphosphate, which interacts with both COI1 and JAZ adjacent to the ligand (Sheard et al. 2010). A recent report indicates that COI1 has a function additional to functioning as a receptor for active JA-conjugates. The vascular pathogen, Verticillium longisporum, requires a COI1 activity that is independent of JA or JA-mimicry to complete its life cycle in Arabidopsis, indicating an unknown function of COI1 during the V. longisporum-plant interaction (Ralhan et al. 2012).
It has been long known that significant overlap exists between COR- and JA-signaling in tomato (Palmer and Bender 1995). Structural and pharmacological studies revealed that COR, as a structural mimic of JA-Ile, binds with high affinity to Arabidopsis COI1 (Sheard et al. 2010). The most active diastereomer for promoting pull down of plant expressed COI1 by JAZ proteins and for promoting anthocyanin accumulation in wild-type and jar1 mutant Arabidopsis seedlings is (+)-7-iso-JA-Ile (Fonseca et al. 2009b). The cyclopentanone ring of COR is a stereoisomer of and demonstrates slightly higher activity than (+)-7-iso-JA-Ile in these assays (Wasternack and Xie 2010; Fonseca et al. 2009b). Also, COR may be resistant to catabolic and epimeric inactivation of JA-Ile (Fonseca et al. 2009b; Koo and Howe 2012). Thus, not only does COR mimic the active JA-Ile conjugate, but it may also function as a hyperactive agonist of JA-signaling.
COR and JA-Ile contact not only COI1, but also the JAZ protein within the COI1-JAZ co-receptor (Sheard et al. 2010), which raises the interesting possibility that COR is biased, relative to JA-Ile, toward specific COI1-JAZ co-receptors. For example, the JA-Ile interacting degron of Arabidopsis JAZ proteins is sequence divergent in JAZ7/8. While this variation renders JAZ8 largely insensitive to JA-Ile, COR retains, albeit at a lower affinity than for other JAZ proteins, the ability to bind and induce degradation of JAZ8 (Shyu et al. 2012). Differences in the ability of COR to target different JAZ proteins could be interpreted in two, non-mutually exclusive ways. Selective targeting of specific JAZs could reflect a “fine-tuning” of transcriptional activation by COR. Alternatively, the inability of COR to target individual JAZs, including alternative C-terminal splice variants with reduced affinity for JA-Ile and COR (Moreno et al. 2013; Chung et al. 2010), could reflect an adaptive, counter-defense strategy of the plant to resist the effect of COR (Chung et al. 2009). In either (or both) case(s), understanding the JAZ-selectivity of COR and JA-Ile is an important area to be explored.
COR suppresses SA-signaling via antagonistic SA-JA crosstalk
Hormone crosstalk is used to fine-tune defense responses against biotic challengers with distinct lifestyles. To exploit these networks to their benefit, numerous plant pathogens produce hormones, hormone mimics, or effectors that stimulate plant production of hormones or modulate hormone signaling. An example of hormone crosstalk relevant to biotic defense occurs between the SA- and JA-dependent signaling pathways (Fig. 1). Generally, the JA/SA balance dictates whether plants mount defense responses tailored to necrotrophic pathogens and herbivores, by favoring JA-signaling, or to biotrophic and hemibiotrophic pathogens, by favoring SA-signaling (Baldwin et al. 1994; Creelman and Mullet 1997; Gimenez-Ibanez and Solano 2013; Kessler and Baldwin 2002; Paschold et al. 2008; Petersen et al. 2000; El Oirdi et al. 2011; Gao et al. 2011; Spoel et al. 2003).
Salicylic acid (SA) is a key phytohormone in plant defense against a variety of biotrophic and hemibiotrophic pathogens, including bacterial strains producing COR (Fig. 1). SA is a monohydroxybenzoic acid that mediates changes in redox potential, probably through S-nitrosylation and thioredoxin activity, when it accumulates in plant cells (Tada et al. 2008). A key protein in SA-signaling is NPR1 (Nonexpresser of PR genes 1) (Cao et al. 1997). The SA-induced redox change leads to the reduction of cytosolic, thiol-linked NPR1 oligomers to monomers that translocate to the nucleus (Cao et al. 1994; Kinkema et al. 2000; Mou et al. 2003). NPR1 monomers activate expression of pathogenesis responsive (PR) genes (Uknes et al. 1992) through interaction with TGA TFs, including TGA2, TGA3, TGA5, TGA6, and TGA7, that bind to activator sequence-1 (as-1) or as-1-like promoter elements (Fan and Dong 2002; Zhou et al. 2000; Zhang et al. 1999; Kim and Delaney 2002; Gimenez-Ibanez and Solano 2013; Despres et al. 2000). Through interaction with TL1-binding factor 1, an HSF-like transcription factor, nuclear-localized NPR1 also activates genes with TL1 promoter elements that support secretion of PR, and perhaps other classes of proteins, through the ER (Pajerowska-Mukhtar et al. 2012; Wang et al. 2005). Additionally, NPR1 induces expression of several WRKY TFs that function as both activators and suppressors of defense (Wang et al. 2006). During its activation by SA, NPR1-phosphorylation facilitates targeting of NPR1 by a Cullin3-based ubiquitin ligase and proteosome-mediated NPR1 turnover is required for full induction of NPR1 target genes (Spoel et al. 2009). One recent report indicated that NPR3 and NPR4, two paralogues of NPR1, are SA-receptors in Arabidopsis that function as adaptors to mediate NPR1 degradation (Fu et al. 2012). A second recent report used equilibrium dialysis ligand binding to show that NPR1 is itself an SA-receptor (Wu et al. 2012). Thus, clearly defining the nature of the SA-receptor(s) remains an important area for further work.
SA plays a central role in regulating plant biotic defenses. In addition to activating defense against biotrophs, for example through inducing expression of defense-promoting secretory genes and antimicrobial PR genes, SA-signaling also has an antagonistic effect on JA-signaling (Fig. 1). Induction of JA-responsive genes is suppressed by SA when SA and MeJA are together exogenously applied to Arabidopsis plants. In SA-deficient NahG plants infected by P. syringae, JA accumulates to 25-fold higher levels and consequently JA-responsive genes are expressed to higher levels (Spoel et al. 2003; Glazebrook et al. 2003). Activation of SA-signaling by P. syringae suppresses JA-signaling and thus renders plants more susceptible to a necrotrophic pathogen (Spoel et al. 2007). NPR1 plays a crucial role in SA-mediated inhibition of JA-dependent signaling, with mechanisms including induced expression of the glutaredoxin GRX480 and the WRKY70 transcription factor as well as destabilization of the ORA59 (OCTADECANOID-RESPONSIVE ARABIDOPSIS AP2/ERF domain protein 59) (Li et al. 2004; Ndamukong et al. 2007; Van der Does et al. 2013; Pre et al. 2008). Also, type II TGA factors are essential for the ability of SA to suppress the ET-signaling contribution to the expression of Arabidopsis genes induced dependent on both JA- and ET-signaling (Zander et al. 2014). Notably, cytosolic, but not nuclear, NPR1 is required for crosstalk, indicating that NPR1 has distinct roles in SA-signaling and suppression of JA-signaling (Spoel et al. 2003). The significance of the suppressive effect of SA on JA-signaling is supported by the observation that it occurs in numerous Arabidopsis accessions treated with SA, MeJA, or both in combination (Koornneef et al. 2008).
The inhibitory crosstalk of SA-signaling toward JA-signaling is mirrored by JA-mediated suppression of SA-signaling (Fig. 1). To exploit this crosstalk, P. syringae produces COR to hijack JA-signaling and suppress SA-mediated defense. Compared to wild-type plants, Pto DC3000 infection of coi1-20 plants elicits elevated levels of SA and PR gene expression, and bacterial growth is suppressed. Bacterial multiplication is recovered in coi1-20 plants expressing NahG (Kloek et al. 2001). Also, cor- (a COR-deficient mutant of Pto DC3000) bacteria induced less JA- and more SA-responsive gene expression, and the reduced growth of cor- strains was restored when the bacteria infected SA-signaling deficient plants (Brooks et al. 2005; Uppalapati et al. 2007; Geng et al. 2012). Thus, studies utilizing both plant and bacterial mutants indicate that COR stimulates JA-signaling to suppress SA-signaling and that SA-signaling is a necessary component of the defense suppressed by COR. A mechanism for suppression of SA-signaling by COR is through MYC2-dependent expression of three NAC TFs that both 1) repress expression of genes controlling SA-biosynthesis, including an isochorismate synthase (SID2), and 2) induce expression of a benzoic/salicylic acid carboxyl methyltransferase (BSMT1) that reduces the pool of free (biologically active) SA via methylation (Zheng et al. 2012).
The ability of COR to inhibit defense signaling in plant cells extends beyond its ability to promote JA-signaling. In addition to causing analogous plant physiological responses as JA, such as inhibiting root elongation, inducing anthocyanin production, and promoting senescence, COR also causes responses not associated with JA, including cell wall thickening and changes to chloroplast structure in tomato plants (Palmer and Bender 1995). Furthermore, exogenous treatment with COR causes hypertrophy and increased amylase activity in potato tuber tissue and also causes anomalous cell growth in tobacco leaves (Kenyon and Turner 1992; Sakai et al. 1979; Sakai 1980; Bender et al. 1999; Feys et al. 1994). More recently, COR was shown to inhibit defensive fortification of cell walls independent of targeting COI1 and suppressing SA-signaling (Geng et al. 2012). Thus, in addition to its well-documented ability to dampen SA-signaling, at least some of the virulence activity of COR is independent of the inhibitory crosstalk between JA- and SA-signaling pathways, and even independent of its ability to target COI1.
COR induces stomatal reopening
A critical first step in the disease cycle of epiphytic phytopathogenic bacteria is the ability to enter the intracellular spaces of plant tissues. Stomata are natural openings and a key portal exploited for bacterial invasion. Guard cells not only regulate gas exchange and water transpiration, but stomatal closure is also an important strategy for plants to prevent the ingress of pathogens, such as P. syringae. Melotto et al. (2006) discovered that P. syrinage on leaf surfaces congregate at stomata and that stomatal closure triggered by recognition of Pathogen Associated Molecular Pattern (PAMPs) is a plant counter-defense strategy to prevent bacterial entry (Melotto et al. 2006). At least two hormone signaling pathways, SA and abscisic acid (ABA), are critical to PAMP-triggered stomatal closure. Stomatal closure was not observed in SA-deficient nahG transgenic plants and SA-biosynthetic mutant sid2 (also known as eds16) plants (Wildermuth et al. 2001). Extensive studies have shown that ABA is required for stomatal closure when plants are under abiotic stress (Cummins et al. 1971; Fan et al. 2004; Mustilli et al. 2002; Tardieu and Davies 1992). PAMP-induced stomatal closure also was not observed in ost1 kinase mutant and ABA-deficient aba3 mutant plants (Leon-Kloosterziel et al. 1996; Melotto et al. 2006; Mustilli et al. 2002).
COR exploits a role for JA-signaling in PAMP-induced stomatal closure. Similar to treatment with purified PAMPs, Pto DC3000 causes stomata to close. But unlike PAMPs, the bacteria quickly reverse the closure, thus allowing for bacterial invasion into the apoplast. The ability of Pto DC3000 to overcome the PAMP-induced stomatal defense is dependent on COR; a cor- strain fails to reopen closed stomata. Furthermore, COR inhibits ABA-induced stomatal closure in a COI1-dependent manner (Melotto et al. 2006). Thus, COR is critical for the ability of P. syringae to overcome PAMP-induced stomatal closure by a mechanism that acts either on or downstream of both SA- and ABA-dependent processes. The same NAC TFs through which COR suppresses SA accumulation also contribute to the ability of COR to overcome ABA-induced stomatal closure and to reopen stomata during P. syringae infection (Zheng et al. 2012).
COR promotes chlorotic disease symptoms in infected plants
Mutant strains of P. syringae unable to produce COR elicit reduced disease symptoms including little or no chlorosis (Feys et al. 1994; Bender et al. 1987; Bender et al. 1999; Kloek et al. 2001; Brooks et al. 2004; Brooks et al. 2005; Block et al. 2005; Mittal and Davis 1995). Treatment of tomato leaves with exogenous COR induced shrunken and descended chloroplasts located near the bottom of the palisade mesophyll cells (Uppalapati et al. 2005). Possibly related to this observation, COR or MeJA repress expression of a large number of genes involved in chloroplast metabolism, including genes encoding chlorophyll a/b binding proteins and thylakoid luminal proteins (Palmer and Bender 1995; Uppalapati et al. 2005; Attaran et al. 2014). Despite these long-standing observations, the molecular basis of how COR contributes to chlorosis is just beginning to be understood.
In Arabidopsis, COR alone induces anthocyanin accumulation (Bent et al. 1992; Feys et al. 1994). However, in the context of an infection by Pto DC3000, COR contributes to chlorotic disease symtpoms. Screening for Arabidopsis mutant plants that do not display chlorosis after infection by Pto DC3000 identified a “no chlorosis” mutant (Mecey et al. 2011). Unlike in wild-type plants, the chlorophyll levels in the mutant are relatively unchanged after infection. The mutation causes an amino acid substitution in the nuclear-encoded, chloroplast-localized Staygreen/Non-Yellowing/Mendel’s I locus (SGR) protein. SGR is associated with chlorophyll degradation. Mutation of SGR causes a stable, non-yellowing phenotype during senescence of leaves (Ren et al. 2007). Exogenously applied COR and Pto DC3000 induce SGR expression in a COI1-dependent manner and, conversely, cor- bacteria induce low levels of SGR compared to the wild-type bacteria. Thus, activating expression of SGR by targeting of COI1 (and thus likely by mimicking JA-Ile) plays a critical role in the contribution of COR to the induction of chlorotic disease symptoms by Pto DC3000 (Mecey et al. 2011).
Wangdi et al. 2010 used virus-induced gene silencing to identify several genes with altered COR (ALC) responses following exogenous application of COR. Silencing of ALC1 in N. benthamiana and tomato resulted in a COR-induced necrotic phenotype that occurs without visible chlorosis. In addition to the lack of COR-induced chlorotic symptoms, Pto DC3000 infection of tomato with silenced ALC1 or Arabidopsis with a mutation of the ALC1 homolog (THF1) induced accelerated, coalescing necrotic lesions without apparent chlorosis (Wangdi et al. 2010). ALC1/THF1 is localized in the chloroplast and ALC1 is destabilized by COR in N. benthamiana leaves (Wangdi et al. 2010; Wang et al. 2004). Since the ability of COR to destabilize ALC1 and to cause necrotic lesions without chlorosis depends on COI1, this activity likely results from its mimicry of JA (Wangdi et al. 2010). Thus, ALC1 links activation of COI1 by COR to both chlorotic and necrotic disease symptoms. SGR-mediated chlorophyll breakdown promotes the production of defense-promoting reactive oxygen (Mur et al. 2010) and ALC1/THF1 is speculated to play a role in maintenance of reactive oxygen homeostasis (Wangdi et al. 2010). Thus, the effects of COR highlight the importance of chloroplast physiology, including chloroplast-derived reactive oxygen, during P. syringae infection.
COR disrupts defense-associated secondary metabolism and cell wall reinforcement
Secondary metabolites are not necessary for plant growth and development in pristine growth conditions, but provide important and sometimes essential functions when plants growing in natural conditions are subjected to biotic or abiotic stresses. JA-signaling plays a crucial role in regulating plant secondary metabolites in both a COI1-dependent and a COI1-independent manner (Devoto et al. 2005). Similarly, COR regulates primary and secondary metabolism during P. syringae infection of Arabidopsis, including the induction of genes involved in tryptophan synthesis, anthocyanin synthesis, and methionine-derived glucosinolates (Thilmony et al. 2006). Treatment of Arabidopsis with purified COR also induces the expression of genes involved in glucosinolate and phenylpropanoid metabolism (Attaran et al. 2014). Indolic compounds constitute one branch of the phenylpropanoid pathway. One fate of indole rings is as intermediates in the synthesis of tryptophan that in turn can serve as a precursor to secondary metabolites involved in plant defense, such as benzoxazinoids, indole glucosinolates (IGs) and the phytoalexin camalexin (Ahmad et al. 2011; Bednarek et al. 2009; Frey et al. 1997). Of these myriad potential effects of COR, perturbation of IGs metabolism has of late come into focus as a potentially critical means by which COR suppresses host defense.
IGs are a class of thioglucosides that have been well documented to play a role in the resistance to chewing insects (Bednarek et al. 2009; Clay et al. 2009; Halkier and Gershenzon 2006; Kim and Jander 2007). More recent studies have demonstrated that tryptophan-derived IGs also play a significant role in defense responses of living tissue against microbes (Bednarek et al. 2009; Clay et al. 2009). Callose, a glucan polymer, is deposited as part of cell wall appositions, which are physical barriers formed at pathogen infection sites. The deposition of callose induced by the PAMP flg22 is well studied in Arabidopsis (Clay et al. 2009; Kim and Mackey 2008). Both ET-signaling and IGs are required for PAMP-induced callose deposition in the leaves of liquid-grown Arabidopsis seedlings (Clay et al. 2009). MYB51, a TF involved in the regulation of IGs biosynthesis, is induced dependent on ET-signaling and is required for the response. A role for IGs in PAMP-induced callose deposition was demonstrated by the lack of callose in mutant seedlings deficient in IGs biosynthesis. 4-methoxyindol-3-ylmethylglucosinolate (4MI3G), an IG candidate found by metabolic profiling, rescued callose deposition in seedlings unable to produce 4MI3G. Furthermore, an unknown hydrolysis product(s) from degradation of 4MI3G by PEN2, an atypical myrosinase (Bednarek et al. 2009; Lipka et al. 2005), are also required for PAMP-induced callose deposition (Clay et al. 2009). Similarly, IGs metabolism and 4MI3G are important for broad-spectrum, penetration-stage resistance of plants to biotrophic fungal pathogens (Bednarek et al. 2009), perhaps also through regulation of cell wall-associated defense. It was recently shown that COR suppresses IGs metabolism and PEN2-dependent callose deposition during P. syringae infection of Arabidopsis (Geng et al. 2012).
It is an interesting paradox that COR inhibits the production of specific IGs, including 4MI3G and presumably downstream products necessary for callose deposition, while it more generally stimulates the expression of genes that promote the production of IGs. The mechanism by which COR perturbs the production of specific IGs is unknown, but may be through perturbation of additional plant hormone signaling pathways. A first possibility is that COR generally perturbs metabolism of indole-containing compounds. Reduced production of IGs and increased expression of genes involved in tryptophan metabolism could relate to changes in the synthesis of the phytohormone auxin. Uppalapati et al. 2005 demonstrated that exogenous application of COR induces auxin-related gene expression in tomato, indicating that COR might promote bacterial virulence by perturbing auxin signaling (Uppalapati et al. 2005; Robert-Seilaniantz et al. 2011; Thilmony et al. 2006; Kazan and Manners 2009). In addition to offsetting the flg22-induced suppression of auxin signaling (Navarro et al. 2006), another consequence of COR promoting auxin production may be to limit indole availability for IGs production.
A second possibility is that COR affects IGs metabolism through perturbation of ET-signaling, which plays a key role in IGs metabolism (Fig. 1). COR causes accumulation of ACC, increased ACC-synthase (ACS) activity, and increased ET production in Bean and Nicotiana tabacum plant leaves (Ferguson and Mitchell 1985; Kenyon and Turner 1992). Whether COR directly or indirectly stimulates ET production is unclear, but considering the CMA moiety of COR is a structural mimic of ACC (Brooks et al. 2004); the effect of COR on production of ET from methionine might be direct. The stimulation of ET production by COR is counter-intuitive relative to its ability to suppress ET-dependent responses, i.e., IGs metabolism and callose deposition. However, this apparent contradiction may result from COR-mediated inhibition of ET production disrupting feedback regulation and thus ultimately leading to mis-timed and/or mis-regulated ET production. Hypotheses for how COR might initially inhibit ET production include interaction with ACS enzymes as a non-released substrate analog or with ACC oxidase enzymes as a competitive inhibitor of ACC.
The role of ET-signaling in IGs metabolism and callose deposition differs between Arabidopsis tissues and growth conditions. Both ET-signaling and IGs are required for PAMP-induced callose deposition in the roots of liquid-grown Arabidopsis seedling. Further experiments with seedling roots indicated that both ET-dependent and ET-independent mechanisms contribute to PAMP-induced expression of MYB51 and that COR, dependent on COI1, inhibits callose deposition and suppresses MYB51 expression in both an ET-signaling dependent and an ET-signaling independent manner (Millet et al. 2010). In the leaves of liquid-grown Arabidopsis seedlings, MYB51 expression, IGs synthesis, and callose deposition were each dependent on ET-signaling (Clay et al. 2009). In the leaves of soil-grown plants, COR promoted bacterial multiplication and inhibited IGs metabolism and callose deposition (Geng et al. 2012). Surprisingly, COR was able to suppress callose deposition in the leaves of coi1-16 mutant plants, indicating a COI1-indpendent defense suppressing activity of COR. Collectively, these studies indicate the existence of complicated mechanisms of PAMP-induced callose deposition in different plant tissues and growth conditions and point to the potential for multiple activities of COR suppressing these pathways.
It is unclear if the COI1-independent, defense suppressing activities of COR in Arabidopsis are mediated through mimicry of JA-Ile, ET, both or neither. Consistent with the JA-mimicry hypothesis, it has been shown that MeJA induces some genes independent of targeting COI1 (Devoto et al. 2005). COR, through its ability to mimic active JA-conjugates, could similarly alter gene expression independent of targeting COI1. One of the COI1-independent, MeJA-induced genes is an ACC-synthase (Devoto et al. 2005), indicating a possible mechanism for how COR might indirectly affect ET-signaling. An alternative and non-exclusive hypothesis is that the COI1-independent function of COR is also independent of its ability to mimic active JA-conjugates. An intriguing possibility is that the CMA moiety of COR, through mimicry of ACC, directly perturbs ET biosynthesis.
COR and type III effectors cooperate to promote bacterial virulence
Bacterial pathogens deploy a variety of virulence factors, including toxins and type III effectors (T3Es), that work in a ‘multifunctional, cooperative, and redundant’ manner (Dean and Kenny 2009). As a result, bacteria often maintain their overall disease-causing ability even when one of their virulence strategies fails due to mutation or incompatibility on a given host. Consistent with this idea, several recent studies have indicated that the multifunctional COR toxin has functions that overlap with various T3Es in a cooperative or (semi-)redundant manner.
Transcription remodeling by COR and type III effectors
An examination of Arabidopsis gene expression following infiltration with wild-type Pto DC3000, Ptocor-, or Ptocor- hrpS (a COR- and type III secretion system (TTSS)-deficient double mutant of Pto DC3000) revealed overlapping yet still distinct roles of COR and T3Es in reprogramming of the host transcriptome (Thilmony et al. 2006). Both COR and T3Es contribute to the regulation of genes responsive to auxin, ABA, and cytokinin, suggesting that Pto DC3000 utilizes multiple virulence factors to ensure the successful perturbation of the host hormone network. One unique effect of COR, the significant induction of JA- responsive genes, is in accordance with JA-mimicry by COR and earlier work in tomato showing COR from Pto DC3000 induces JA-related gene expression and contributes to virulence (Zhao et al. 2003). Another unique effect of COR likely related to JA-mimicry by COR, is the prominent modulation on genes involved in secondary metabolism consistent with induced expression of Arabidopsis genes involved in secondary metabolism by JAs (Sasaki-Sekimoto et al. 2005; Taki et al. 2005). Interestingly, some of the secondary metabolism-related genes are antagonistically regulated by COR and type III effectors, presumably due to the distinct functions of these virulence factors. Contrary to COR, the prominent function of T3Es was differential regulation of SA-related genes and suppression of basal defense-related genes. Suppression of a few basal defense-related genes by COR was also observed. This study defined effects of COR by comparing wild-type Pto DC3000 to the Ptocor- mutant, both of which deliver the full complement of T3Es. Thus, some effects of COR were likely masked by T3Es and more functional overlap is expected between COR and T3Es than revealed by this study alone. Examples of functional overlap between COR and specific T3Es are described in the following sections.
Perturbation of hormone signaling by COR and T3Es
A variety of T3Es have been demonstrated to perturb hormone signaling pathways also targeted by COR. The T3Es AvrPto and AvrPtoB from Pto DC3000 induce ET production and signaling contributing to cell death in susceptible tomato plants (Cohn and Martin 2005). The ability of these T3Es to stimulate ET production correlates with their ability to induce the expression of two tomato ACC oxidase genes. Thus, in tomato, the virulence activity AvrPto and AvrPtoB might functionally overlap with that of COR. While functional redundancy between COR, AvrPto and AvrPtoB is speculative, the following paragraphs describe examples of overlapping function of COR with three different T3Es that target the JA-signaling pathway. Interestingly, relative to direct targeting of COI1 by COR, these T3Es perturb JA-signaling upstream of or at the COI1-signaling node.
The first example is AvrB, a T3E that targets upstream of COI1 (Fig. 1). He et al. 2004 showed that Pto DC3000, dependent on both COR and T3Es, induces the expression of an Arabidopsis ERF (ethylene responsive factor) gene, RAP2.6. Both a TTSS mutant and a cor- mutant of DC3000 failed to induce RAP2.6. Interestingly, AvrB complemented the ability of the cor- mutant strain to induce RAP2.6, indicating overlapping activities for the T3E and toxin. Further support for this overlap came from the observation that the ability of AvrB to induce RAP2.6 was dependent on COI1 (He et al. 2004). Later work indicated the ability of AvrB to activate JA-signaling is mediated by targeting of MAP kinase 4 (Cui et al. 2010). Thus, AvrB appears to induce JA-response genes by activating JA-signaling upstream of COI1.
The other two effectors activate JA-signaling by targeting JAZ proteins (Fig. 1). HopZ1a from P. syringae pv. syringae strain A2 acetylates JAZ proteins causing them to become destabilized dependent on COI1 (Jiang et al. 2013). HopX1 from P. syringae pv. tabaci (Pta) strain 11528 destabilizes JAZ proteins without a requirement for COI1, likely via its cysteine protease activity that directly cleaves the central Zim domain of the JAZ proteins (Gimenez-Ibanez et al. 2014). Like COR, both HopX1 and HopZ1a can induce expression of JA-response genes, suppress SA-signaling, and restore virulence to cor- mutant Pto DC3000 (Jiang et al. 2013; Gimenez-Ibanez et al. 2014). HopX1 shares additional activities with COR, including reopening of stomata, causing plant cells to lose chlorophyll, and induction of chlorosis in susceptible plants (Gimenez-Ibanez et al. 2014). Since Pta 11528 does not produce COR, HopX1 may serve as an alternative evolutionary strategy to compensate for the lack of COR. It is interesting to consider whether HopZ1a and HopX1 will target all or a specific subset of JAZ proteins in host plants. When tested against a nearly complete set of Arabidopsis JAZ proteins, HopX1 targeted all and HopZ1a targeted a subset. One model is that JAZ-selectivity reflects fine-tuning of the virulence activity of a T3E. Another possibility is that resistant JAZ proteins, such as those derived from alternate splice variants or sequence divergent homologs, are present in co-evolved hosts to help overcome the effects of T3Es. In either case, HopX1, which comes from a tabaci pathovar of P. syringae, might be expected to target only a subset of JAZ proteins from tobacco plants.
Suppression of cell wall defense by COR and T3Es
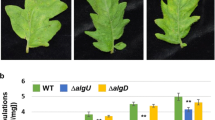
HopM1 is a T3E encoded by a gene located in the conserved effector locus (CEL) of Pto DC3000. HopM1 overcomes SA-dependent host immunity by destabilizing AtMIN7 to disrupt G-protein-mediated endomembrane trafficking as well as perturbing another Arabidopsis target(s) to disrupt an SA-independent pathway supporting Arabidopsis defense (Nomura et al. 2006; Gangadharan et al. 2013; Nomura et al. 2011) (Fig. 1). CEL, which is physically adjacent to the locus containing genes that encode the structural proteins of the type III secretion system apparatus, contains effectors important for the virulence of a variety of bacterial pathogens (Alfano et al. 2000; Badel et al. 2006; DebRoy et al. 2004; Ham et al. 2006; Kvitko et al. 2009). A recent study revealed a functional overlap between COR and HopM1 in suppressing cell wall-associated defense (Geng et al. 2012).
The ability of COR to suppress cell wall-associated defense escaped detection until recently (Geng et al. 2012; Millet et al. 2010) because the effect of COR is masked by T3Es of the CEL. Further obscuring this activity of COR, PtoΔCEL (CEL deletion of Pto DC3000) elicits SA-signaling that overcomes the suppressive effect of COR. The new activity of COR was detected by examining defense responses against a PtoΔCEL cor- (CEL deletion and COR-deficient double mutant strain) in SA-signaling deficient mutant plants (sid2 and npr1). The PtoΔCEL cor- double mutant elicited more callose and grew less than the PtoΔCEL single mutant in SA-signaling mutant plants. Furthermore, those higher levels of callose elicited by PtoΔCEL cor- were suppressed by either exogenous application of COR or expression of hopM1 in the double mutant strain. Thus, COR and HopM1 carry out overlapping roles in suppressing cell wall-associated defense.
Although COR and T3Es of the CEL locus share the ability to suppress cell wall-associated defense, their mode of action differs. In SA-signaling competent plants, HopM1 suppressed the high levels of callose deposition induced by PtoΔCEL cor- while COR could not. Thus, COR and HopM1 distinctly suppress signaling within the plant defense network by functioning in mechanistically distinct manners. The ability of COR to suppress callose deposition and promote bacterial growth in SA-signaling deficient mutants indicates that COR targets an SA-independent sector. Consistent with this idea, Geng et al. 2012 showed that COR perturbs IGs metabolism. Unlike COR that can only do so in SA-signaling deficient mutant plants, HopM1 suppresses callose deposition induced by PtoΔCEL cor- and restores bacterial growth in both wild-type and SA-signaling deficient mutant plants. Thus, HopM1 suppresses both SA-dependent and SA-independent sectors, perhaps through downstream effects on defense-associated vesicle trafficking. The mode of action by which COR and HopM1 each target distinct sectors of a converged portion of the plant defense network remains to be elucidated.
Conclusions and future questions
Plants consistently face environmental stresses, including biotic stresses, due to their sessile lifestyle. The key roles of hormone signaling and secondary metabolic pathways in the ability of plants to deal with these stresses make targeting of them an effective strategy deployed by plant bacterial pathogens to suppress host defense. Here, we reviewed the demonstrated ability of the phytotoxin COR to hijack JA-signaling and thus suppress SA-signaling. We also speculated about targeting of ET-signaling by COR and considered how one, or perhaps both, of these activities contributes to the various virulence activities of COR. The finding that COR promotes virulence independent of targeting COI1 opens a path to study this novel activity of COR separate from the confounding effects of COI1 activation. The perturbation of ET-dependent IGs metabolism provides an attractive system for this effort. In addition to producing hormones and/or hormone mimics, such as COR, bacteria also produce T3Es. COR cooperates with other T3Es to promote bacterial virulence and dampen the induced defense responses. The collaborative targeting of plant hosts by pathogen-produced virulence factors, for example by COR and T3Es that target plant hormone signaling pathways, is an area that, with further exploration, will reveal a better view of the elegant network comprising the plant immune system and how it is collaboratively defeated by pathogen-derived virulence factors.
References
Ahmad S, Veyrat N, Gordon-Weeks R, Zhang Y, Martin J, Smart L, Glauser G, Erb M, Flors V, Frey M, Ton J (2011) Benzoxazinoid metabolites regulate innate immunity against aphids and fungi in maize. Plant Physiol 157(1):317–327. doi:10.1104/pp.111.180224
Alfano JR, Charkowski AO, Deng WL, Badel JL, Petnicki-Ocwieja T, van Dijk K, Collmer A (2000) The Pseudomonas syringae Hrp pathogenicity island has a tripartite mosaic structure composed of a cluster of type III secretion genes bounded by exchangeable effector and conserved effector loci that contribute to parasitic fitness and pathogenicity in plants. Proc Natil Acad Sci USA 97(9):4856–4861
Attaran E, Major IT, Cruz JA, Rosa BA, Koo AJ, Chen J, Kramer DM, He SY, Howe GA (2014) Temporal dynamics of growth and photosynthesis suppression in response to jasmonate signaling. Plant Physiol 165(3):1302–1314. doi:10.1104/pp.114.239004
Badel JL, Shimizu R, Oh HS, Collmer A (2006) A Pseudomonas syringae pv. tomato avrE1/hopM1 mutant is severely reduced in growth and lesion formation in tomato. MPMI 19(2):99–111. doi:10.1094/MPMI-19-0099
Baldwin IT, Schmelz EA, Ohnmeiss TE (1994) Wound-induced changes in root and shoot jasmonic acid pools correlate with induced nicotine synthesis in Nicotiana sylvestris spegazzini and comes. J Chem Ecolo 20(8):2139–2157. doi:10.1007/BF02066250
Bari R, Jones JD (2009) Role of plant hormones in plant defence responses. Plant Mol Biol 69(4):473–488. doi:10.1007/s11103-008-9435-0
Bednarek P, Pislewska-Bednarek M, Svatos A, Schneider B, Doubsky J, Mansurova M, Humphry M, Consonni C, Panstruga R, Sanchez-Vallet A, Molina A, Schulze-Lefert P (2009) A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense. Science 323(5910):101–106. doi:10.1126/science.1163732
Bender CL (1999) Chlorosis-inducing phytotoxins produced by Pseudomonas syringae. Eur J Plant Pathol 105:1–12
Bender CL, Stone HE, Sims JJ, Cooksey DA (1987) Reduced pathogen fitness of Pseudomonas syringae pv. tomato Tn5 mutants defective in coronatine production. Physiol Mol Plant Pathol 30:273–283
Bender CL, Liyanage H, Palmer D, Ullrich M, Young S, Mitchell R (1993) Characterization of the genes controlling the biosynthesis of the polyketide phytotoxin coronatine including conjugation between coronafacic and coronamic acid. Gene 133(1):31–38
Bender CL, Alarcon-Chaidez F, Gross DC (1999) Pseudomonas syringae phytotoxins: mode of action, regulation, and biosynthesis by peptide and polyketide synthetases. MMBR 63(2):266–292
Bent AF, Innes RW, Ecker JR, Staskawicz BJ (1992) Disease development in ethylene-insensitive arabidopsis thaliana infected with virulent and avirulent Pseudomonas and Xanthomonas Pathogens. MPMI 5(5):372–378
Block A, Schmelz E, Jones JB, Klee HJ (2005) Coronatine and salicylic acid: the battle between Arabidopsis and Pseudomonas for phytohormone control. Mol Plant Pathol 6(1):79–83. doi:10.1111/j.1364-3703.2004.00265.x
Braun Y, Smirnova AV, Schenk A, Weingart H, Burau C, Muskhelishvili G, Ullrich MS (2008) Component and protein domain exchange analysis of a thermoresponsive, two-component regulatory system of Pseudomonas syringae. Microbiology 154(Pt 9):2700–2708. doi:10.1099/mic.0.2008/018820-0
Brooks DM, Hernandez-Guzman G, Kloek AP, Alarcon-Chaidez F, Sreedharan A, Rangaswamy V, Penaloza-Vazquez A, Bender CL, Kunkel BN (2004) Identification and characterization of a well-defined series of coronatine biosynthetic mutants of Pseudomonas syringae pv. tomato DC3000. MPMI 17(2):162–174. doi:10.1094/mpmi.2004.17.2.162
Brooks DM, Bender CL, Kunkel BN (2005) The Pseudomonas syringae phytotoxin coronatine promotes virulence by overcoming salicylic acid-dependent defences in Arabidopsis thaliana. Mol Plant Pathol 6(6):629–639. doi:10.1111/j.1364-3703.2005.00311.x
Buell CR, Joardar V, Lindeberg M, Selengut J, Paulsen IT, Gwinn ML, Dodson RJ, Deboy RT, Durkin AS, Kolonay JF, Madupu R, Daugherty S, Brinkac L, Beanan MJ, Haft DH, Nelson WC, Davidsen T, Zafar N, Zhou L, Liu J, Yuan Q, Khouri H, Fedorova N, Tran B, Russell D, Berry K, Utterback T, Van Aken SE, Feldblyum TV, D’Ascenzo M, Deng WL, Ramos AR, Alfano JR, Cartinhour S, Chatterjee AK, Delaney TP, Lazarowitz SG, Martin GB, Schneider DJ, Tang X, Bender CL, White O, Fraser CM, Collmer A (2003) The complete genome sequence of the Arabidopsis and tomato pathogen Pseudomonas syringae pv. tomato DC3000. Proc Natil Acad Sci USA 100(18):10181–10186. doi:10.1073/pnas.1731982100
Cao H, Bowling SA, Gordon AS, Dong X (1994) Characterization of an Arabidopsis mutant that is nonresponsive to inducers of systemic acquired resistance. Plant Cell 6(11):1583–1592
Cao H, Glazebrook J, Clarke JD, Volko S, Dong X (1997) The Arabidopsis NPR1 gene that controls systemic acquired resistance encodes a novel protein containing ankyrin repeats. Cell 88(1):57–63
Cheng H, Song S, Xiao L, Soo HM, Cheng Z, Xie D, Peng J (2009) Gibberellin acts through jasmonate to control the expression of MYB21, MYB24, and MYB57 to promote stamen filament growth in Arabidopsis. PLoS Gen 5(3):e1000440. doi:10.1371/journal.pgen.1000440
Chini A, Fonseca S, Fernandez G, Adie B, Chico JM, Lorenzo O, Garcia-Casado G, Lopez-Vidriero I, Lozano FM, Ponce MR, Micol JL, Solano R (2007) The JAZ family of repressors is the missing link in jasmonate signalling. Nature 448(7154):666–671. doi:10.1038/nature06006
Chung HS, Niu Y, Browse J, Howe GA (2009) Top hits in contemporary JAZ: an update on jasmonate signaling. Phytochemistry 70(13–14):1547–1559. doi:10.1016/j.phytochem.2009.08.022
Chung HS, Cooke TF, Depew CL, Patel LC, Ogawa N, Kobayashi Y, Howe GA (2010) Alternative splicing expands the repertoire of dominant JAZ repressors of jasmonate signaling. Plant J 63(4):613–622. doi:10.1111/j.1365-313X.2010.04265.x
Cintas NAK ST, Bull CT (2002) A new pathovar, Pseudomonas syringae pv. alisalensis pv. nov., proposed for the causal agent of bacterial blight of broccoli and broccoli raab. Plant Dis 86:992–998
Clay NK, Adio AM, Denoux C, Jander G, Ausubel FM (2009) Glucosinolate metabolites required for an Arabidopsis innate immune response. Science 323(5910):95–101. doi:10.1126/science.1164627
Cohn JR, Martin GB (2005) Pseudomonas syringae pv. tomato type III effectors AvrPto and AvrPtoB promote ethylene-dependent cell death in tomato. Plant J 44(1):139–154. doi:10.1111/j.1365-313X.2005.02516.x (for cell and molecular biology)
Creelman RA, Mullet JE (1997) Biosynthesis and action of Jasmonates in plants. Annu Rev Plant Physiol Plant Mol Biol 48:355–381. doi:10.1146/annurev.arplant.48.1.355
Cui H, Wang Y, Xue L, Chu J, Yan C, Fu J, Chen M, Innes RW, Zhou JM (2010) Pseudomonas syringae effector protein AvrB perturbs Arabidopsis hormone signaling by activating MAP kinase 4. Cell Host Microbe 7(2):164–175. doi:10.1016/j.chom.2010.01.009
Cummins WR, Kende H, Raschke K (1971) Specificity and reversibility of the rapid stomatal response to abscisic acid. Planta 99(4):347–351. doi:10.1007/BF00385826
Daub ME, Herrero S, Chung KR (2005) Photoactivated perylenequinone toxins in fungal pathogenesis of plants. FEMS Microbiol Lett 252(2):197–206. doi:10.1016/j.femsle.2005.08.033
Dean P, Kenny B (2009) The effector repertoire of enteropathogenic E. coli: ganging up on the host cell. Curr Opin Microbiol 12(1):101–109. doi:10.1016/j.mib.2008.11.006
DebRoy S, Thilmony R, Kwack YB, Nomura K, He SY (2004) A family of conserved bacterial effectors inhibits salicylic acid-mediated basal immunity and promotes disease necrosis in plants. Proc Natil Acad Sci USA 101(26):9927–9932. doi:10.1073/pnas.0401601101
Delaney TP, Uknes S, Vernooij B, Friedrich L, Weymann K, Negrotto D, Gaffney T, Gut-Rella M, Kessmann H, Ward E, Ryals J (1994) A central role of salicylic acid in plant disease resistance. Science 266(5188):1247–1250
del Pozo JC, Estelle M (1999) The Arabidopsis cullin AtCUL1 is modified by the ubiquitin-related protein RUB1. Proc Natil Acad Sci USA 96(26):15342–15347
Deshaies RJ (1999) SCF and Cullin/Ring H2-based ubiquitin ligases. Annu Rev Plant Physiol Plant Mol Biol 15:435–467. doi:10.1146/annurev.cellbio.15.1.435
Despres C, DeLong C, Glaze S, Liu E, Fobert PR (2000) The Arabidopsis NPR1/NIM1 protein enhances the DNA binding activity of a subgroup of the TGA family of bZIP transcription factors. Plant Cell 12(2):279–290
Devoto A, Nieto-Rostro M, Xie D, Ellis C, Harmston R, Patrick E, Davis J, Sherratt L, Coleman M, Turner JG (2002) COI1 links jasmonate signalling and fertility to the SCF ubiquitin-ligase complex in Arabidopsis. Plant J 32(4):457–466
Devoto A, Ellis C, Magusin A, Chang HS, Chilcott C, Zhu T, Turner JG (2005) Expression profiling reveals COI1 to be a key regulator of genes involved in wound- and methyl jasmonate-induced secondary metabolism, defence, and hormone interactions. Plant Mol Biol 58(4):497–513. doi:10.1007/s11103-005-7306-5
Dharmasiri N, Dharmasiri S, Weijers D, Lechner E, Yamada M, Hobbie L, Ehrismann JS, Jurgens G, Estelle M (2005) Plant development is regulated by a family of auxin receptor F box proteins. Develop Cell 9(1):109–119. doi:10.1016/j.devcel.2005.05.014
Dombrecht B, Xue GP, Sprague SJ, Kirkegaard JA, Ross JJ, Reid JB, Fitt GP, Sewelam N, Schenk PM, Manners JM, Kazan K (2007) MYC2 differentially modulates diverse jasmonate-dependent functions in Arabidopsis. Plant Cell 19(7):2225–2245. doi:10.1105/tpc.106.048017
Duke SO, Dayan FE (2011) Modes of action of microbially-produced phytotoxins. Toxins 3(8):1038–1064. doi:10.3390/toxins3081038
El Oirdi M, El Rahman TA, Rigano L, El Hadrami A, Rodriguez MC, Daayf F, Vojnov A, Bouarab K (2011) Botrytis cinerea manipulates the antagonistic effects between immune pathways to promote disease development in tomato. Plant 23(6):2405–2421. doi:10.1105/tpc.111.083394
Fan W, Dong X (2002) In vivo interaction between NPR1 and transcription factor TGA2 leads to salicylic acid-mediated gene activation in Arabidopsis. Plant Cell 14(6):1377–1389
Fan LM, Zhao Z, Assmann SM (2004) Guard cells: a dynamic signaling model. Curr Opin Plant Biol 7(5):537–546. doi:10.1016/j.pbi.2004.07.009
Ferguson AR, Johnston JS (1980) Phaseolotoxin: chlorosis, ornithine accumulation and inhibition of ornithine carbamoyltransferase in different plants. Physiol Plant Pathol 16(2):269–275
Ferguson IB, Mitchell RE (1985) Stimulation of ethylene production in bean leaf discs by the pseudomonad phytotoxin coronatine. Plant Physiol 77(4):969–973
Feys B, Benedetti CE, Penfold CN, Turner JG (1994) Arabidopsis mutants selected for resistance to the phytotoxin coronatine are male sterile, insensitive to methyl jasmonate, and resistant to a bacterial pathogen. Plant Cell 6(5):751–759. doi:10.1105/tpc.6.5.751
Fonseca S, Chico JM, Solano R (2009a) The jasmonate pathway: the ligand, the receptor and the core signalling module. Curr Opin Plant Biol 12(5):539–547. doi:10.1016/j.pbi.2009.07.013
Fonseca S, Chini A, Hamberg M, Adie B, Porzel A, Kramell R, Miersch O, Wasternack C, Solano R (2009b) (+)-7-iso-Jasmonoyl-l-isoleucine is the endogenous bioactive jasmonate. Nat Chem Biol 5(5):344–350. doi:10.1038/nchembio.161
Frey M, Chomet P, Glawischnig E, Stettner C, Grun S, Winklmair A, Eisenreich W, Bacher A, Meeley RB, Briggs SP, Simcox K, Gierl A (1997) Analysis of a chemical plant defense mechanism in grasses. Science 277(5326):696–699
Fu ZQ, Yan S, Saleh A, Wang W, Ruble J, Oka N, Mohan R, Spoel SH, Tada Y, Zheng N, Dong X (2012) NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 486(7402):228–232. doi:10.1038/nature11162
Gaffney T, Friedrich L, Vernooij B, Negrotto D, Nye G, Uknes S, Ward E, Kessmann H, Ryals J (1993) Requirement of salicylic acid for the induction of systemic acquired resistance. Science 261(5122):754–756
Gangadharan A, Sreerekha MV, Whitehill J, Ham JH, Mackey D (2013) The Pseudomonas syringae pv. tomato type III effector HopM1 suppresses arabidopsis defenses independent of suppressing salicylic acid signaling and of targeting AtMIN7. PLoS ONE 8(12):e82032. doi:10.1371/journal.pone.0082032
Gao QM, Venugopal S, Navarre D, Kachroo A (2011) Low oleic acid-derived repression of jasmonic acid-inducible defense responses requires the WRKY50 and WRKY51 proteins. Plant Physiol 155(1):464–476. doi:10.1104/pp.110.166876
Geng X, Cheng J, Gangadharan A, Mackey D (2012) The coronatine toxin of Pseudomonas syringae is a multifunctional suppressor of Arabidopsis defense. Plant Cell 24(11):4763–4774. doi:10.1105/tpc.112.105312
Gimenez-Ibanez S, Solano R (2013) Nuclear jasmonate and salicylate signaling and crosstalk in defense against pathogens. Frontier Plant Sci 4:72. doi:10.3389/fpls.2013.00072
Gimenez-Ibanez S, Boter M, Fernandez-Barbero G, Chini A, Rathjen JP, Solano R (2014) The bacterial effector HopX1 targets JAZ transcriptional repressors to activate jasmonate signaling and promote infection in Arabidopsis. PLoS Biol 12(2):e1001792. doi:10.1371/journal.pbio.1001792
Glazebrook J (2005) Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Ann Rev Phytopathol 43:205–227. doi:10.1146/annurev.phyto.43.040204.135923
Glazebrook J, Chen W, Estes B, Chang HS, Nawrath C, Metraux JP, Zhu T, Katagiri F (2003) Topology of the network integrating salicylate and jasmonate signal transduction derived from global expression phenotyping. Plant J 34(2):217–228
Goudet C, Benitah JP, Milat ML, Sentenac H, Thibaud JB (1999) Cluster organization and pore structure of ion channels formed by beticolin 3, a nonpeptidic fungal toxin. Biophy J 77(6):3052–3059. doi:10.1016/S0006-3495(99)77136-5
Gray WM, del Pozo JC, Walker L, Hobbie L, Risseeuw E, Banks T, Crosby WL, Yang M, Ma H, Estelle M (1999) Identification of an SCF ubiquitin-ligase complex required for auxin response in Arabidopsis thaliana. Genes Dev 13(13):1678–1691
Gray WM, Kepinski S, Rouse D, Leyser O, Estelle M (2001) Auxin regulates SCF(TIR1)-dependent degradation of AUX/IAA proteins. Nature 414(6861):271–276
Gross H, Loper JE (2009) Genomics of secondary metabolite production by Pseudomonas spp. Nat Prod Rep 26(11):1408–1446. doi:10.1039/b817075b
Haider G, von Schrader T, Fusslein M, Blechert S, Kutchan TM (2000) Structure-activity relationships of synthetic analogs of jasmonic acid and coronatine on induction of benzo[c]phenanthridine alkaloid accumulation in Eschscholzia californica cell cultures. Biol Chem 381(8):741–748. doi:10.1515/BC.2000.094
Halkier BA, Gershenzon J (2006) Biology and biochemistry of glucosinolates. Ann Rev Plant Biol 57:303–333. doi:10.1146/annurev.arplant.57.032905.105228
Ham JH, Majerczak DR, Arroyo-Rodriguez AS, Mackey DM, Coplin DL (2006) WtsE, an AvrE-family effector protein from Pantoea stewartii subsp. stewartii, causes disease-associated cell death in corn and requires a chaperone protein for stability. MPMI 19(10):1092–1102
He P, Chintamanani S, Chen Z, Zhu L, Kunkel BN, Alfano JR, Tang X, Zhou JM (2004) Activation of a COI1-dependent pathway in Arabidopsis by Pseudomonas syringae type III effectors and coronatine. Plant J 37(4):589–602
Hershko A, Ciechanover A (1998) The ubiquitin system. Ann Rev Biochem 67:425–479. doi:10.1146/annurev.biochem.67.1.425
Hoffman JCV KC (1995) Post-translational tubulin modifications in spermatogeneous cells of the pteridophyte Ceratopteris richardii. Protoplasma 186(3–4):169–182
Hogenhout SA, Van der Hoorn RA, Terauchi R, Kamoun S (2009) Emerging concepts in effector biology of plant-associated organisms. MPMI 22(2):115–122. doi:10.1094/MPMI-22-2-0115
Hopke J, Donath J, Blechert S, Boland W (1994) Herbivore-induced volatiles: the emission of acyclic homoterpenes from leaves of Phaseolus lunatus and zea mays can be triggered by a beta-glucosidase and jasmonic acid. FEBS Lett 352(2):146–150
Howe GA, Jander G (2008) Plant immunity to insect herbivores. Ann Rev Plant Biol 59:41–66. doi:10.1146/annurev.arplant.59.032607.092825
Ichihara A, Shiraishi K, Sato H, Sakamura S, Nishiyama K, Sakai R, Furusaki A, Matsumoto T (1977) The Structure of coronatine. J Am Chem Soc 99(2):636–637
Jiang S, Yao J, Ma KW, Zhou H, Song J, He SY, Ma W (2013) Bacterial effector activates jasmonate signaling by directly targeting JAZ transcriptional repressors. PLoS Pathog 9(10):e1003715. doi:10.1371/journal.ppat.1003715
Karban R, Baldwin IT (1998) Induced responses to herbivory. Trends Ecol Evol 13(2):83
Katagiri F, Tsuda K (2010) Understanding the plant immune system. MPMI 23(12):1531–1536. doi:10.1094/MPMI-04-10-0099
Katsir L, Schilmiller AL, Staswick PE, He SY, Howe GA (2008) COI1 is a critical component of a receptor for jasmonate and the bacterial virulence factor coronatine. Proc Natil Acad Sci USA 105(19):7100–7105. doi:10.1073/pnas.0802332105
Kazan K, Manners JM (2009) Linking development to defense: auxin in plant-pathogen interactions. Trends Plant Sci 14(7):373–382. doi:10.1016/j.tplants.2009.04.005
Kazan K, Manners JM (2012) JAZ repressors and the orchestration of phytohormone crosstalk. Trends Plant Sci 17(1):22–31. doi:10.1016/j.tplants.2011.10.006
Kenyon JS, Turner JG (1992) The stimulation of ethylene synthesis in nicotiana tabacum leaves by the phytotoxin coronatine. Plant Physiol 100(1):219–224
Kessler A, Baldwin IT (2002) Plant responses to insect herbivory: the emerging molecular analysis. Ann Rev Plant Biol 53:299–328. doi:10.1146/annurev.arplant.53.100301.135207
Kim HS, Delaney TP (2002) Over-expression of TGA5, which encodes a bZIP transcription factor that interacts with NIM1/NPR1, confers SAR-independent resistance in Arabidopsis thaliana to Peronospora parasitica. Plant J 32(2):151–163
Kim JH, Jander G (2007) Myzus persicae (green peach aphid) feeding on Arabidopsis induces the formation of a deterrent indole glucosinolate. Plant J 49(6):1008–1019. doi:10.1111/j.1365-313X.2006.03019.x
Kim M, Mackey D (2008) Measuring cell-wall-based defenses and their effect on bacterial growth in Arabidopsis. Met Mol Biol 415:443–452
King RR, Calhoun LA (2009) The thaxtomin phytotoxins: sources, synthesis, biosynthesis, biotransformation and biological activity. Phytochemistry 70(7):833–841. doi:10.1016/j.phytochem.2009.04.013
Kinkema M, Fan W, Dong X (2000) Nuclear localization of NPR1 is required for activation of PR gene expression. Plant Cell 12(12):2339–2350
Kloek AP, Verbsky ML, Sharma SB, Schoelz JE, Vogel J, Klessig DF, Kunkel BN (2001) Resistance to Pseudomonas syringae conferred by an Arabidopsis thaliana coronatine-insensitive (coi1) mutation occurs through two distinct mechanisms. Plant J 26(5):509–522
Koo AJ, Howe GA (2012) Catabolism and deactivation of the lipid-derived hormone jasmonoyl-isoleucine. Frontiers Plant Sci 3:19. doi:10.3389/fpls.2012.00019
Koornneef A, Leon-Reyes A, Ritsema T, Verhage A, Den Otter FC, Van Loon LC, Pieterse CM (2008) Kinetics of salicylate-mediated suppression of jasmonate signaling reveal a role for redox modulation. Plant Physiol 147(3):1358–1368. doi:10.1104/pp.108.121392
Kvitko BH, Park DH, Velasquez AC, Wei CF, Russell AB, Martin GB, Schneider DJ, Collmer A (2009) Deletions in the repertoire of Pseudomonas syringae pv. tomato DC3000 type III secretion effector genes reveal functional overlap among effectors. PLoS Pathog 5(4):e1000388. doi:10.1371/journal.ppat.1000388
Leon-Kloosterziel KM, Gil MA, Ruijs GJ, Jacobsen SE, Olszewski NE, Schwartz SH, Zeevaart JA, Koornneef M (1996) Isolation and characterization of abscisic acid-deficient Arabidopsis mutants at two new loci. Plant J 10(4):655–661
Li J, Wang XQ, Watson MB, Assmann SM (2000) Regulation of abscisic acid-induced stomatal closure and anion channels by guard cell AAPK kinase. Science 287(5451):300–303
Li J, Brader G, Palva ET (2004) The WRKY70 transcription factor: a node of convergence for jasmonate-mediated and salicylate-mediated signals in plant defense. Plant Cell 16(2):319–331. doi:10.1105/tpc.016980
Lipka V, Dittgen J, Bednarek P, Bhat R, Wiermer M, Stein M, Landtag J, Brandt W, Rosahl S, Scheel D, Llorente F, Molina A, Parker J, Somerville S, Schulze-Lefert P (2005) Pre- and postinvasion defenses both contribute to nonhost resistance in Arabidopsis. Science 310(5751):1180–1183
Liyanage H, Penfold C, Turner J, Bender CL (1995) Sequence, expression and transcriptional analysis of the coronafacate ligase-encoding gene required for coronatine biosynthesis by Pseudomonas syringae. Gene 153(1):17–23
Lorenzo O, Chico JM, Sanchez-Serrano JJ, Solano R (2004) JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell 16(7):1938–1950. doi:10.1105/tpc.022319
Mecey C, Hauck P, Trapp M, Pumplin N, Plovanich A, Yao J, He SY (2011) A critical role of STAYGREEN/Mendel’s I locus in controlling disease symptom development during Pseudomonas syringae pv tomato infection of Arabidopsis. Plant Physiol 157(4):1965–1974. doi:10.1104/pp.111.181826
Melotto M, Underwood W, Koczan J, Nomura K, He SY (2006) Plant stomata function in innate immunity against bacterial invasion. Cell 126(5):969–980. doi:10.1016/j.cell.2006.06.054
Millet YA, Danna CH, Clay NK, Songnuan W, Simon MD, Werck-Reichhart D, Ausubel FM (2010) Innate immune responses activated in Arabidopsis roots by microbe-associated molecular patterns. Plant Cell 22(3):973–990. doi:10.1105/tpc.109.069658
Mitchell RE (1982) Coronatine production by some phytopathogenic pseudomonads. Physiol Plant Pathol 20(1):83–89
Mitchell RE (1985) Coronatine biosynthesis: incorporation of l-[U-14C]isoleucine and l-[U-14C] threonine into the 1-amido-1-carboxy-2-ethylcyclopropyl moiety. Phytochemistry 24(2):247–249
Mitchell R (1991) Coronatine analogues produced by xanthomonas campestris pv. phormiicola. Phytochemistry 30(12):3917–3920
Mitchell RE, Hale CN, Shanks JC (1983) Production of different pathogenic symptoms and different toxins by strains of Pseudomonas syringae pv. tomato not distinguishable by gel-immunodiffusion assay. Physiol Plant Pathol 23(3):315–322
Mittal S, Davis KR (1995) Role of the phytotoxin coronatine in the infection of Arabidopsis thaliana by Pseudomonas syringae pv. tomato. MPMI 8(1):165–171
Moreno JE, Shyu C, Campos ML, Patel LC, Chung HS, Yao J, He SY, Howe GA (2013) Negative feedback control of jasmonate signaling by an alternative splice variant of JAZ10. Plant Physio 162(2):1006–1017. doi:10.1104/pp.113.218164
Mou Z, Fan W, Dong X (2003) Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 113(7):935–944
Mur LA, Aubry S, Mondhe M, Kingston-Smith A, Gallagher J, Timms-Taravella E, James C, Papp I, Hortensteiner S, Thomas H, Ougham H (2010) Accumulation of chlorophyll catabolites photosensitizes the hypersensitive response elicited by Pseudomonas syringae in Arabidopsis. New Phytol 188(1):161–174. doi:10.1111/j.1469-8137.2010.03377.x
Mustilli AC, Merlot S, Vavasseur A, Fenzi F, Giraudat J (2002) Arabidopsis OST1 protein kinase mediates the regulation of stomatal aperture by abscisic acid and acts upstream of reactive oxygen species production. Plant Cell 14(12):3089–3099
Navarro L, Dunoyer P, Jay F, Arnold B, Dharmasiri N, Estelle M, Voinnet O, Jones JD (2006) A plant miRNA contributes to antibacterial resistance by repressing auxin signaling. Science 312(5772):436–439. doi:10.1126/science.1126088
Ndamukong I, Abdallat AA, Thurow C, Fode B, Zander M, Weigel R, Gatz C (2007) SA-inducible Arabidopsis glutaredoxin interacts with TGA factors and suppresses JA-responsive PDF1.2 transcription. Plant J 50(1):128–139. doi:10.1111/j.1365-313X.2007.03039.x
Nomura K, Debroy S, Lee YH, Pumplin N, Jones J, He SY (2006) A bacterial virulence protein suppresses host innate immunity to cause plant disease. Science 313(5784):220–223. doi:10.1126/science.1129523
Nomura K, Mecey C, Lee YN, Imboden LA, Chang JH, He SY (2011) Effector-triggered immunity blocks pathogen degradation of an immunity-associated vesicle traffic regulator in Arabidopsis. Proc Natil Acad Sci USA 108(26):10774–10779. doi:10.1073/pnas.1103338108
Norman C, Vidal S, Palva ET (1999) Oligogalacturonide-mediated induction of a gene involved in jasmonic acid synthesis in response to the cell-wall-degrading enzymes of the plant pathogen Erwinia carotovora. MPMI 12(7):640–644. doi:10.1094/MPMI.1999.12.7.640
Pajerowska-Mukhtar KM, Wang W, Tada Y, Oka N, Tucker CL, Fonseca JP, Dong X (2012) The HSF-like transcription factor TBF1 is a major molecular switch for plant growth-to-defense transition. Curr Biol 22(2):103–112
Palmer DA, Bender CL (1995) Ultrastructure of tomato leaf tissue treated with the Pseudomonas phytotoxin coronatine and comparison with methyl jasmonate. MPMI 8(5):683–692
Parry R, Mhaskar SV, Lin M, Walker AE, Mafoti R (1993) Investigations of the biosynthesis of the Phytotoxin coronatine. Can J Chem 72:86–99
Paschold A, Bonaventure G, Kant MR, Baldwin IT (2008) Jasmonate perception regulates jasmonate biosynthesis and JA-Ile metabolism: the case of COI1 in Nicotiana attenuata. Plant Cell Physiol 49(8):1165–1175. doi:10.1093/pcp/pcn091
Pauwels L, Goossens A (2011) The JAZ proteins: a crucial interface in the jasmonate signaling cascade. Plant Cell 23(9):3089–3100. doi:10.1105/tpc.111.089300
Pauwels L, Barbero GF, Geerinck J, Tilleman S, Grunewald W, Perez AC, Chico JM, Bossche RV, Sewell J, Gil E, Garcia-Casado G, Witters E, Inze D, Long JA, De Jaeger G, Solano R, Goossens A (2010) NINJA connects the co-repressor TOPLESS to jasmonate signalling. Nature 464(7289):788–791. doi:10.1038/nature08854
Petersen M, Brodersen P, Naested H, Andreasson E, Lindhart U, Johansen B, Nielsen HB, Lacy M, Austin MJ, Parker JE, Sharma SB, Klessig DF, Martienssen R, Mattsson O, Jensen AB, Mundy J (2000) Arabidopsis map kinase 4 negatively regulates systemic acquired resistance. Cell 103(7):1111–1120
Pieterse CM, Leon-Reyes A, Van der Ent S, Van Wees SC (2009) Networking by small-molecule hormones in plant immunity. Nat Chem Biol 5(5):308–316. doi:10.1038/nchembio.164
Pieterse CM, Van der Does D, Zamioudis C, Leon-Reyes A, Van Wees SC (2012) Hormonal modulation of plant immunity. Ann Rev Cell Dev Biol 28:489–521. doi:10.1146/annurev-cellbio-092910-154055
Pre M, Atallah M, Champion A, De Vos M, Pieterse CM, Memelink J (2008) The AP2/ERF domain transcription factor ORA59 integrates jasmonic acid and ethylene signals in plant defense. Plant Physiol 147(3):1347–1357. doi:10.1104/pp.108.117523
Preston GM (2000) Pseudomonas syringae pv. tomato: the right pathogen, of the right plant, at the right time. Mol Pl Pathol 1(5):263–275
Pruess DL, Scannell JP, Ax HA, Kellett M, Weiss F (1973) Antimetabolites produced by microorganisms. VII. L-(N5-phosphono)methionine-S-sulfoximinyl-L-alanyl-l-alanine. J Antibiot 26(5):261–266
Qi T, Song S, Ren Q, Wu D, Huang H, Chen Y, Fan M, Peng W, Ren C, Xie D (2011) The Jasmonate-ZIM-domain proteins interact with the WD-Repeat/bHLH/MYB complexes to regulate Jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell 23(5):1795–1814. doi:10.1105/tpc.111.083261
Ralhan A, Schottle S, Thurow C, Iven T, Feussner I, Polle A, Gatz C (2012) The vascular pathogen verticillium longisporum requires a jasmonic acid-independent COI1 function in roots to elicit disease symptoms in Arabidopsis shoots. Plant Physiol 159(3):1192–1203. doi:10.1104/pp.112.198598
Ren G, An K, Liao Y, Zhou X, Cao Y, Zhao H, Ge X, Kuai B (2007) Identification of a novel chloroplast protein AtNYE1 regulating chlorophyll degradation during leaf senescence in Arabidopsis. Plant Physiol 144(3):1429–1441. doi:10.1104/pp.107.100172
Robert-Seilaniantz A, Grant M, Jones JD (2011) Hormone crosstalk in plant disease and defense: more than just jasmonate-salicylate antagonism. Ann Rev Phytopathol 49:317–343. doi:10.1146/annurev-phyto-073009-114447
Rohde BH, Pohlack B, Ullrich MS (1998) Occurrence of thermoregulation of genes involved in coronatine biosynthesis among various Pseudomonas syringae strains. J Basic Microbiol 38(1):41–50
Sakai R (1980) Comparison of physiological activities between coronatine and Indole-3-acetic acid to some plant tissues. Annals Phytopathol Soc Japan 46:499–503
Sakai R, Nishiyama K, Ichihara A, Shiraishi K, Sakamura S (1979) Studies on the mechanism of physiological activity of coronatine effect of coronatine on cell wall extensibility and expansion of potato tuber tissue. Annals of the Phytopathological Society of Japan 45:645–653
Sasaki-Sekimoto Y, Taki N, Obayashi T, Aono M, Matsumoto F, Sakurai N, Suzuki H, Hirai MY, Noji M, Saito K, Masuda T, Takamiya K, Shibata D, Ohta H (2005) Coordinated activation of metabolic pathways for antioxidants and defence compounds by jasmonates and their roles in stress tolerance in Arabidopsis. Plant J 44(4):653–668. doi:10.1111/j.1365-313X.2005.02560.x
Schenk PM, Kazan K, Wilson I, Anderson JP, Richmond T, Somerville SC, Manners JM (2000) Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc Natil Acad Sci USA 97(21):11655–11660. doi:10.1073/pnas.97.21.11655
Schroeder JI, Allen GJ, Hugouvieux V, Kwak JM, Waner D (2001) Guard cell signal transduction. Ann Rev Plant Physiol Plant Mol Biol 52:627–658. doi:10.1146/annurev.arplant.52.1.627
Schwechheimer C, Willige BC (2009) Shedding light on gibberellic acid signalling. Curr Opinion Plant Biol 12(1):57–62. doi:10.1016/j.pbi.2008.09.004
Seo JS, Joo J, Kim MJ, Kim YK, Nahm BH, Song SI, Cheong JJ, Lee JS, Kim JK, Choi YD (2011) OsbHLH148, a basic helix-loop-helix protein, interacts with OsJAZ proteins in a jasmonate signaling pathway leading to drought tolerance in rice. Plant J 65(6):907–921. doi:10.1111/j.1365-313X.2010.04477.x
Sheard LB, Tan X, Mao H, Withers J, Ben-Nissan G, Hinds TR, Kobayashi Y, Hsu FF, Sharon M, Browse J, He SY, Rizo J, Howe GA, Zheng N (2010) Jasmonate perception by inositol-phosphate-potentiated COI1-JAZ co-receptor. Nature 468(7322):400–405. doi:10.1038/nature09430
Shyu C, Figueroa P, Depew CL, Cooke TF, Sheard LB, Moreno JE, Katsir L, Zheng N, Browse J, Howe GA (2012) JAZ8 lacks a canonical degron and has an EAR motif that mediates transcriptional repression of jasmonate responses in Arabidopsis. Plant Cell 24(2):536–550. doi:10.1105/tpc.111.093005
Smirnova AV, Braun Y, Ullrich MS (2008) Site-directed mutagenesis of the temperature-sensing histidine protein kinase CorS from Pseudomonas syringae. FEMS Microbiol Let 283(2):231–238. doi:10.1111/j.1574-6968.2008.01179.x
Song S, Qi T, Huang H, Ren Q, Wu D, Chang C, Peng W, Liu Y, Peng J, Xie D (2011) The Jasmonate-ZIM domain proteins interact with the R2R3-MYB transcription factors MYB21 and MYB24 to affect Jasmonate-regulated stamen development in Arabidopsis. Plant Cell 23(3):1000–1013. doi:10.1105/tpc.111.083089
Spoel SH, Koornneef A, Claessens SM, Korzelius JP, Van Pelt JA, Mueller MJ, Buchala AJ, Metraux JP, Brown R, Kazan K, Van Loon LC, Dong X, Pieterse CM (2003) NPR1 modulates cross-talk between salicylate- and jasmonate-dependent defense pathways through a novel function in the cytosol. Plant Cell 15(3):760–770
Spoel SH, Johnson JS, Dong X (2007) Regulation of tradeoffs between plant defenses against pathogens with different lifestyles. Proc Natil Acad Sci USA 104(47):18842–18847. doi:10.1073/pnas.0708139104
Spoel SH, Mou Z, Tada Y, Spivey NW, Genschik P, Dong X (2009) Proteasome-mediated turnover of the transcription coactivator NPR1 plays dual roles in regulating plant immunity. Cell 137(5):860–872. doi:10.1016/j.cell.2009.03.038
Sreedharan A, Penaloza-Vazquez A, Kunkel BN, Bender CL (2006) CorR regulates multiple components of virulence in Pseudomonas syringae pv. tomato DC3000. MPMI 19(7):768–779. doi:10.1094/mpmi-19-0768
Staswick PE, Tiryaki I, Rowe ML (2002) Jasmonate response locus JAR1 and several related Arabidopsis genes encode enzymes of the firefly luciferase superfamily that show activity on jasmonic, salicylic, and indole-3-acetic acids in an assay for adenylation. Plant Cell 14(6):1405–1415
Stotz HU, Pittendrigh BR, Kroymann J, Weniger K, Fritsche J, Bauke A, Mitchell-Olds T (2000) Induced plant defense responses against chewing insects. Ethylene signaling reduces resistance of Arabidopsis against Egyptian cotton worm but not diamondback moth. Plant Physiol 124(3):1007–1018
Strobel G, Yang X, Sears J, Kramer R, Sidhu RS, Hess WM (1996) Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana. Microbiology 142(Pt 2):435–440
Suza WP, Staswick PE (2008) The role of JAR1 in Jasmonoyl-L: -isoleucine production during Arabidopsis wound response. Planta 227(6):1221–1232. doi:10.1007/s00425-008-0694-4
Szemenyei H, Hannon M, Long JA (2008) TOPLESS mediates auxin-dependent transcriptional repression during Arabidopsis embryogenesis. Science 319(5868):1384–1386. doi:10.1126/science.1151461
Tada Y, Spoel SH, Pajerowska-Mukhtar K, Mou Z, Song J, Wang C, Zuo J, Dong X (2008) Plant immunity requires conformational changes [corrected] of NPR1 via S-nitrosylation and thioredoxins. Science 321(5891):952–956. doi:10.1126/science.1156970
Taki N, Sasaki-Sekimoto Y, Obayashi T, Kikuta A, Kobayashi K, Ainai T, Yagi K, Sakurai N, Suzuki H, Masuda T, Takamiya K, Shibata D, Kobayashi Y, Ohta H (2005) 12-oxo-phytodienoic acid triggers expression of a distinct set of genes and plays a role in wound-induced gene expression in Arabidopsis. Plant Physiol 139(3):1268–1283. doi:10.1104/pp.105.067058
Tamura K, Takikawa Y, Tsuyumu S, Goto M, Watanabe M (1992) Coronatine Production by Xanthomonas campestris pv. phormiicola. Ann Phytopathol Soc Japan 58:276–281
Tanaka THK, Watanabe M, Abbas HK (1996) Isolation, purification and identification of 2, 5-anhydro-d-glucitol as a phytotoxin from Fusarium solani. J Nat Toxin 5(3):317–330
Tardieu F, Davies WJ (1992) Stomatal response to abscisic Acid is a function of current plant water status. Plant Physiol 98(2):540–545
Thilmony R, Underwood W, He SY (2006) Genome-wide transcriptional analysis of the Arabidopsis thaliana interaction with the plant pathogen Pseudomonas syringae pv. tomato DC3000 and the human pathogen Escherichia coli O157:H7. Plant J 46(1):34–53. doi:10.1111/j.1365-313X.2006.02725.x
Thines B, Katsir L, Melotto M, Niu Y, Mandaokar A, Liu G, Nomura K, He SY, Howe GA, Browse J (2007) JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signalling. Nature 448(7154):661–665. doi:10.1038/nature05960
Thuleau P, Graziana A, Rossignol M, Kauss H, Auriol P, Ranjeva R (1988) Binding of the phytotoxin zinniol stimulates the entry of calcium into plant protoplasts. Proc Natil Acad Sci USA 85(16):5932–5935
Turner JG (1981) Tabtoxin, produced by Pseudomonas tabaci, decreases Nicotiana tabacum glutamine synthetase in vivo and causes accumulation of ammonia. Physiol Plant Pathol 19(1):57–67
Uchytil TF, Durbin RD (1980) Hydrolysis of tabtoxins by plant and bacterial enzymes. Experientia 36:301–302
Uknes S, Mauch-Mani B, Moyer M, Potter S, Williams S, Dincher S, Chandler D, Slusarenko A, Ward E, Ryals J (1992) Acquired resistance in Arabidopsis. Plant Cell 4(6):645–656
Ullrich M, Bereswill S, Volksch B, Fritsche W, Geider K (1993) Molecular characterization of field isolates of Pseudomonas syringae pv. glycinea differing in coronatine production. J General Microbiol 139(8):1927–1937
Ullrich M, Penaloza-Vazquez A, Bailey AM, Bender CL (1995) A modified two-component regulatory system is involved in temperature-dependent biosynthesis of the Pseudomonas syringae phytotoxin coronatine. J Bacteriol 177(21):6160–6169
Uppalapati SR, Ayoubi P, Weng H, Palmer DA, Mitchell RE, Jones W, Bender CL (2005) The phytotoxin coronatine and methyl jasmonate impact multiple phytohormone pathways in tomato. Plant J 42(2):201–217. doi:10.1111/j.1365-313X.2005.02366.x
Uppalapati SR, Ishiga Y, Wangdi T, Kunkel BN, Anand A, Mysore KS, Bender CL (2007) The phytotoxin coronatine contributes to pathogen fitness and is required for suppression of salicylic acid accumulation in tomato inoculated with Pseudomonas syringae pv. tomato DC3000. MPMI 20(8):955–965. doi:10.1094/MPMI-20-8-0955
Van der Does D, Leon-Reyes A, Koornneef A, Van Verk MC, Rodenburg N, Pauwels L, Goossens A, Korbes AP, Memelink J, Ritsema T, Van Wees SC, Pieterse CM (2013) Salicylic acid suppresses jasmonic acid signaling downstream of SCFCOI1-JAZ by targeting GCC promoter motifs via transcription factor ORA59. Plant Cell 25(2):744–761. doi:10.1105/tpc.112.108548
Walton JD (2006) HC-toxin. Phytochemistry 67(14):1406–1413. doi:10.1016/j.phytochem.2006.05.033
Wang Q, Sullivan RW, Kight A, Henry RL, Huang J, Jones AM, Korth KL (2004) Deletion of the chloroplast-localized Thylakoid formation1 gene product in Arabidopsis leads to deficient thylakoid formation and variegated leaves. Plant Physiol 136(3):3594–3604. doi:10.1104/pp.104.049841
Wang D, Weaver ND, Kesarwani M, Dong X (2005) Induction of protein secretory pathway is required for systemic acquired resistance. Science 308(5724):1036–1040. doi:10.1126/science.1108791
Wang D, Amornsiripanitch N, Dong X (2006) A genomic approach to identify regulatory nodes in the transcriptional network of systemic acquired resistance in plants. PLoS Pathogen 2(11):e123. doi:10.1371/journal.ppat.0020123
Wangdi T, Uppalapati SR, Nagaraj S, Ryu CM, Bender CL, Mysore KS (2010) A virus-induced gene silencing screen identifies a role for Thylakoid Formation1 in Pseudomonas syringae pv tomato symptom development in tomato and Arabidopsis. Plant Physiol 152(1):281–292. doi:10.1104/pp.109.148106
Wasternack C, Hause B (2013) Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the, 2007 review in Annals of Botany. Annals Bot 111(6):1021–1058. doi:10.1093/aob/mct067
Wasternack C, Xie D (2010) The genuine ligand of a jasmonic acid receptor: improved analysis of jasmonates is now required. Plant Signal Behav 5(4):337–340
Weiler EW, Kutchan TM, Gorba T, Brodschelm W, Niesel U, Bublitz F (1994) The Pseudomonas phytotoxin coronatine mimics octadecanoid signalling molecules of higher plants. FEBS Let 345(1):9–13
Weingart H, Stubner S, Schenk A, Ullrich MS (2004) Impact of temperature on in planta expression of genes involved in synthesis of the Pseudomonas syringae phytotoxin coronatine. MPMI 17(10):1095–1102. doi:10.1094/MPMI.2004.17.10.1095
Wiebe WL, Campbell RN (1993) Characterization of Pseudomonas syringae pv. maculicola and comparison with P.s. tomato. Plant Dis 77:414–419
Wildermuth MC, Dewdney J, Wu G, Ausubel FM (2001) Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 414(6863):562–565. doi:10.1038/35107108
Worley JN, Russell AB, Wexler AG, Bronstein PA, Kvitko BH, Krasnoff SB, Munkvold KR, Swingle B, Gibson DM, Collmer A (2013) Pseudomonas syringae pv. tomato DC3000 CmaL (PSPTO4723), a DUF1330 family member, is needed to produce L-allo-isoleucine, a precursor for the phytotoxin coronatine. J Bacteriol 195(2):287–296. doi:10.1128/jb.01352-12
Wu Y, Zhang D, Chu JY, Boyle P, Wang Y, Brindle ID, De Luca V, Despres C (2012) The Arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid. Cell Rep 1(6):639–647. doi:10.1016/j.celrep.2012.05.008
Xu L, Liu F, Lechner E, Genschik P, Crosby WL, Ma H, Peng W, Huang D, Xie D (2002) The SCF(COI1) ubiquitin-ligase complexes are required for jasmonate response in Arabidopsis. Plant Cell 14(8):1919–1935
Yamaguchi S (2008) Gibberellin metabolism and its regulation. Ann Rev Plant Biol 59:225–251. doi:10.1146/annurev.arplant.59.032607.092804
Zander M, Thurow C, Gatz C (2014) TGA transcription factors activate the salicylic acid-suppressible branch of the ethylene-induced defense program by regulating ORA59 expression. Plant Physiol 165(4):1671–1683. doi:10.1104/pp.114.243360
Zhang Y, Fan W, Kinkema M, Li X, Dong X (1999) Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene. Proc Natil Acad Sci USA 96(11):6523–6528
Zhao Y, Damicone JP, Demezas DH, Rangaswamy V, Bender CL (2000) Bacterial leaf spot of leafy crucifers in Oklahoma caused by Pseudomonas syringae pv. maculicola. Plant Dis 84(9):1015–1020
Zhao Y, Thilmony R, Bender CL, Schaller A, He SY, Howe GA (2003) Virulence systems of Pseudomonas syringae pv. tomato promote bacterial speck disease in tomato by targeting the jasmonate signaling pathway. Plant J 36(4):485–499
Zheng XY, Spivey NW, Zeng W, Liu PP, Fu ZQ, Klessig DF, He SY, Dong X (2012) Coronatine promotes Pseudomonas syringae virulence in plants by activating a signaling cascade that inhibits salicylic acid accumulation. Cell Host Microbe 11(6):587–596. doi:10.1016/j.chom.2012.04.014
Zhou JM, Trifa Y, Silva H, Pontier D, Lam E, Shah J, Klessig DF (2000) NPR1 differentially interacts with members of the TGA/OBF family of transcription factors that bind an element of the PR-1 gene required for induction by salicylic acid. MPMI 13(2):191–202. doi:10.1094/MPMI.2000.13.2.191
Acknowledgments
We would like to thank our reviewers for critical suggestions and comments. We apologize to those authors whose works we could not cite because of space limitations. This work was supported by the National Science Foundation (MCB-0718882), the USDA (NIFA 2008-35319-04506), and the Ohio Agricultural Research and Development Center of The Ohio State University to DM, the Korean RDA Next-Generation BioGreen Program (SSAC, grant # PJ009088) to MGK and DM, and NRF-2012R1A1A1011351, Republic of Korea to MGK.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article

Cite this article
Geng, X., Jin, L., Shimada, M. et al. The phytotoxin coronatine is a multifunctional component of the virulence armament of Pseudomonas syringae . Planta 240, 1149–1165 (2014). https://doi.org/10.1007/s00425-014-2151-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-014-2151-x