
Bees react to a perfume reminiscent of a distant food source by revisiting the site.
Abstract
The honeybee, Apis mellifera, navigates rapidly and accurately to food sources that are often kilometres away1,2. They achieve this by learning visual cues, such as the location and colour of nectar-bearing flowers1,2,3, and chemical cues, such as the scent and the taste of the nectar1,2. Here we train bees to visit differently scented sugar feeders placed at specific outdoor locations and find that they can be induced to visit the same locations simply by having the corresponding scent blown into the hive, even when the destinations no longer have the food or carry the scent. A familiar nectar scent can trigger specific memories of a route and therefore expedite navigation to the food source.
Similar content being viewed by others

Main
Von Frisch1 observed that a floral scent on a forager bee returning to the hive from a natural food source can induce other experienced foragers to revisit that source without communicating with each other by waggle dancing4. This and similar observations2,5,6,7 raise the question of what memories are triggered by scent. We now investigate this in an experiment designed to ensure that the bees do not just home in on a familiar scent1,2,5,6,7 and are not recruited by waggle dances.
We trained individually marked honeybees to visit two sugar feeders positioned at two different locations, each 50 m from the hive; the distance between the two locations was 30 m. Each feeder carried a different scent, and feeders were offered alternately, switching every half hour. In the first experiment, scent one (offered at location one) was rose, and scent two (offered at location two) was lemon. After two days of training, the bees were tested periodically.
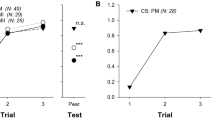
In the tests, the feeders were replaced by two empty, unscented jars. Scents one and two were blown into the hive, in turn, for 8 min each, by using a small fan attached to the hive entrance. During each scent-blowing interval, we noted the number of marked bees arriving at each test feeder. We also noted which feeder was visited first, the number of circlings (flights around the feeder), the number of landings on the feeder and the total number of visits (sum of all circlings and landings, including first visits). Tests were performed four times for each scent.
The trained bees that emerged from the hive upon injection of a scent showed a significant preference for the location that had carried the scent during training (Fig. 1). Injection of rose scent into the hive caused the trained bees to fly predominantly to location one. When lemon scent was blown into the hive, the overwhelming majority of trained bees visited location two. We repeated the experiment with different pairs of scents: rose and almond (experiment two), and lemon and almond (experiment three). Each time we obtained similar results (Fig. 1). These findings indicate that each injected scent triggers visual memories of a specific location that the trained bees had previously visited.

Three pairs of scent combinations were tested: rose and lemon, rose and almond, and almond and lemon. The proportion of first visits, circlings, landings and total visits to the two test feeders are shown. Colours indicate preferred feeder location: yellow, rose; green, lemon; orange, almond. The number of individually marked bees that visited both test feeders, the number of choices (n) and the P-values (χ2-test for choice frequencies) are shown at the bottom.
Our test feeders were unscented, so the bees could not have found the correct site simply by homing in on the familiar scent, which is a well-known foraging strategy in bees1,2,8. As the test feeders were empty, the arriving bees were not rewarded and so would not have danced upon return to the hive to recruit nestmates1,4. Bees arriving at the correct location must therefore have relied on previously acquired navigational memories evoked by the injected scent.
Our findings reveal that associative recall is a mechanism used by honeybees, in addition to the dance language1,4, to navigate successfully and repeatedly to an attractive food source. The taste and scent of nectar samples distributed by foragers on return to the hive1 could trigger recall of specific visual and route memories associated with the food site in experienced recruits. Recalling factors such as the distance and direction of the food source, the landmarks en route and at the destination, and the flower's colour and shape would speed up flight to the food site, enhancing the foraging efficiency of the colony.
References
von Frisch, K. The Dance Language and Orientation of Bees (Harvard Univ. Press, London, 1993).
Gould, J. L. in Insect Learning (eds Papaj, D. R. & Lewis, A. C.) 18–50 (Chapman & Hall, New York, 1993).
Wehner, R. in Handbook of Sensory Physiology (ed. Autrum, H.) 288–616 (Springer, Berlin, 1981).
Dyer, F. C. Annu. Rev. Entomol. 47, 917–949 (2002).
Ribbands, C. R. Proc. R. Entomol. Soc. A 29, 141–144 (1954).
Free, J. B. Nature 222, 778 (1969).
Jakobsen, H. B., Kristjansson, K., Rohde, B., Terkildsen, M. & Olsen, C. E. J. Chem. Ecol. 21, 1635–1648 (1995).
Tautz, J. & Sandeman, D. C. J. Comp. Physiol. A 189, 293–300 (2003).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
About this article
Cite this article
Reinhard, J., Srinivasan, M. & Zhang, S. Scent-triggered navigation in honeybees. Nature 427, 411 (2004). https://doi.org/10.1038/427411a
Issue Date:
DOI: https://doi.org/10.1038/427411a
This article is cited by
-
How India Changed My Ideas About Honey Bees
Journal of the Indian Institute of Science (2023)
-
Varieties of visual navigation in insects
Animal Cognition (2023)
-
Evaluation of efficacy of pinene compounds as mosquitocidal agent against Aedes aegypti Linn. (Diptera: culicidae)
International Journal of Tropical Insect Science (2022)
-
Indirect plant defenses: volatile organic compounds and extrafloral nectar
Arthropod-Plant Interactions (2021)
-
Australian stingless bees detect odours left at food sources by nestmates, conspecifics and honey bees
Insectes Sociaux (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
