
Abstract
Myeloproliferative neoplasms (MPNs) are diseases caused by mutations in the haematopoietic stem cell (HSC) compartment. Most MPN patients have a common acquired mutation of Janus kinase 2 (JAK2) gene in HSCs1,2,3,4 that renders this kinase constitutively active, leading to uncontrolled cell expansion. The bone marrow microenvironment might contribute to the clinical outcomes of this common event. We previously showed that bone marrow nestin+ mesenchymal stem cells (MSCs) innervated by sympathetic nerve fibres regulate normal HSCs5,6. Here we demonstrate that abrogation of this regulatory circuit is essential for MPN pathogenesis. Sympathetic nerve fibres, supporting Schwann cells and nestin+ MSCs are consistently reduced in the bone marrow of MPN patients and mice expressing the human JAK2(V617F) mutation in HSCs. Unexpectedly, MSC reduction is not due to differentiation but is caused by bone marrow neural damage and Schwann cell death triggered by interleukin-1β produced by mutant HSCs. In turn, in vivo depletion of nestin+ cells or their production of CXCL12 expanded mutant HSC number and accelerated MPN progression. In contrast, administration of neuroprotective or sympathomimetic drugs prevented mutant HSC expansion. Treatment with β3-adrenergic agonists that restored the sympathetic regulation of nestin+ MSCs5,6 prevented the loss of these cells and blocked MPN progression by indirectly reducing the number of leukaemic stem cells. Our results demonstrate that mutant-HSC-driven niche damage critically contributes to disease manifestation in MPN and identify niche-forming MSCs and their neural regulation as promising therapeutic targets.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Accession codes
Primary accessions
Gene Expression Omnibus
Data deposits
The gene expression data have been deposited in the Gene Expression Omnibus (GEO) databank (http://www.ncbi.nlm.nih.gov/geo) under the accession number GSE55802.
References
James, C. et al. A unique clonal JAK2 mutation leading to constitutive signalling causes polycythaemia vera. Nature 434, 1144–1148 (2005)
Kralovics, R. et al. A gain-of-function mutation of JAK2 in myeloproliferative disorders. N. Engl. J. Med. 352, 1779–1790 (2005)
Levine, R. L. et al. Activating mutation in the tyrosine kinase JAK2 in polycythemia vera, essential thrombocythemia, and myeloid metaplasia with myelofibrosis. Cancer Cell 7, 387–397 (2005)
Baxter, E. J. et al. Acquired mutation of the tyrosine kinase JAK2 in human myeloproliferative disorders. Lancet 365, 1054–1061 (2005)
Méndez-Ferrer, S. et al. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 466, 829–834 (2010)
Méndez-Ferrer, S., Lucas, D., Battista, M. & Frenette, P. S. Haematopoietic stem cell release is regulated by circadian oscillations. Nature 452, 442–447 (2008)
Raaijmakers, M. H. et al. Bone progenitor dysfunction induces myelodysplasia and secondary leukaemia. Nature 464, 852–857 (2010)
Walkley, C. R. et al. A microenvironment-induced myeloproliferative syndrome caused by retinoic acid receptor gamma deficiency. Cell 129, 1097–1110 (2007)
Walkley, C. R., Shea, J. M., Sims, N. A., Purton, L. E. & Orkin, S. H. Rb regulates interactions between hematopoietic stem cells and their bone marrow microenvironment. Cell 129, 1081–1095 (2007)
Colmone, A. et al. Leukemic cells create bone marrow niches that disrupt the behavior of normal hematopoietic progenitor cells. Science 322, 1861–1865 (2008)
Iriuchishima, H. et al. Neovascular niche for human myeloma cells in immunodeficient mouse bone. PLoS ONE 7, e30557 (2012)
Pikman, Y. et al. MPLW515L is a novel somatic activating mutation in myelofibrosis with myeloid metaplasia. PLoS Med. 3, e270 (2006)
Pardanani, A. D. et al. MPL515 mutations in myeloproliferative and other myeloid disorders: a study of 1182 patients. Blood 108, 3472–3476 (2006)
Klampfl, T. et al. Somatic mutations of calreticulin in myeloproliferative neoplasms. N. Engl. J. Med. 369, 2379–2390 (2013)
Nangalia, J. et al. Somatic CALR mutations in myeloproliferative neoplasms with nonmutated JAK2. N. Engl. J. Med. 369, 2391–2405 (2013)
Shih, A. H., Abdel-Wahab, O., Patel, J. P. & Levine, R. L. The role of mutations in epigenetic regulators in myeloid malignancies. Nature Rev. Cancer 12, 599–612 (2012)
Vannucchi, A. M. et al. Mutations and prognosis in primary myelofibrosis. Leukemia 27, 1861–1869 (2013)
Isern, J. et al. Self-renewing human bone marrow mesenspheres promote hematopoietic stem cell expansion. Cell Rep. 3, 1714–1724 (2013)
Tiedt, R. et al. Ratio of mutant JAK2-V617F to wild-type Jak2 determines the MPD phenotypes in transgenic mice. Blood 111, 3931–3940 (2008)
Kubovcakova, L. et al. Differential effects of hydroxyurea and INC424 on mutant allele burden and myeloproliferative phenotype in a JAK2-V617F polycythemia vera mouse model. Blood 121, 1188–1199 (2013)
Schepers, K. et al. Myeloproliferative neoplasia remodels the endosteal bone marrow niche into a self-reinforcing leukemic niche. Cell Stem Cell 13, 285–299 (2013)
Yamazaki, S. et al. Nonmyelinating Schwann cells maintain hematopoietic stem cell hibernation in the bone marrow niche. Cell 147, 1146–1158 (2011)
Nagasawa, T. et al. Defects of B-cell lymphopoiesis and bone-marrow myelopoiesis in mice lacking the CXC chemokine PBSF/SDF-1. Nature 382, 635–638 (1996)
Tzeng, Y. S. et al. Loss of Cxcl12/Sdf-1 in adult mice decreases the quiescent state of hematopoietic stem/progenitor cells and alters the pattern of hematopoietic regeneration after myelosuppression. Blood 117, 429–439 (2011)
Rameshwar, P., Denny, T. N., Stein, D. & Gascon, P. Monocyte adhesion in patients with bone marrow fibrosis is required for the production of fibrogenic cytokines. Potential role for interleukin-1 and TGF-beta. J. Immunol. 153, 2819–2830 (1994)
Lucas, D. et al. Chemotherapy-induced bone marrow nerve injury impairs hematopoietic regeneration. Nature Med. 19, 695–703 (2013)
Mignone, J. L., Kukekov, V., Chiang, A. S., Steindler, D. & Enikolopov, G. Neural stem and progenitor cells in nestin-GFP transgenic mice. J. Comp. Neurol. 469, 311–324 (2004)
Balordi, F. & Fishell, G. Mosaic removal of hedgehog signaling in the adult SVZ reveals that the residual wild-type stem cells have a limited capacity for self-renewal. J. Neurosci. 27, 14248–14259 (2007)
Sousa, V. H., Miyoshi, G., Hjerling-Leffler, J., Karayannis, T. & Fishell, G. Characterization of Nkx6-2-derived neocortical interneuron lineages. Cereb. Cortex 19 (Suppl. 1). i1–i10 (2009)
Brockschnieder, D., Pechmann, Y., Sonnenberg-Riethmacher, E. & Riethmacher, D. An improved mouse line for Cre-induced cell ablation due to diphtheria toxin A, expressed from the Rosa26 locus. Genesis 44, 322–327 (2006)
Frenette, P. S., Subbarao, S., Mazo, I. B., von Andrian, U. H. & Wagner, D. D. Endothelial selectins and vascular cell adhesion molecule-1 promote hematopoietic progenitor homing to bone marrow. Proc. Natl Acad. Sci. USA 95, 14423–14428 (1998)
Stemple, D. L. & Anderson, D. J. Isolation of a stem cell for neurons and glia from the mammalian neural crest. Cell 71, 973–985 (1992)
Pajtler, K. et al. Production of chick embryo extract for the cultivation of murine neural crest stem cells. J. Vis. Exp. 45, e2380 (2010)
Biernaskie, J. A., McKenzie, I. A., Toma, J. G. & Miller, F. D. Isolation of skin-derived precursors (SKPs) and differentiation and enrichment of their Schwann cell progeny. Nature Protocols 1, 2803–2812 (2007)
Davis, S. & Meltzer, P. S. GEOquery: a bridge between the Gene Expression Omnibus (GEO) and BioConductor. Bioinformatics 23, 1846–1847 (2007)
Johnson, W. E., Li, C. & Rabinovic, A. Adjusting batch effects in microarray expression data using empirical Bayes methods. Biostatistics 8, 118–127 (2007)
Acknowledgements
We thank S. Martín-Salamanca, A. M. Martín, A. B. Ricote, J. M. Ligos, S. Bartlett and the CNIC Comparative Medicine, Genomics and Bioinformatics Units for assistance; members of S.M.-F. and B. Ibáñez groups and M. García-Fernández for data discussion; G. Enikolopov, G. Fishell and D. Riethmacher for providing mice. This work was supported by Fundación CNIC, Spanish Ministry of Economy and Competitiveness (TerCel, Spanish Cell Therapy Network; Plan Nacional SAF-2011-30308) and ConSEPOC-Comunidad de Madrid S2010/BMD-2542 grants to S.M.-F.; Ramón y Cajal Program grants RYC-2009-04703/2011-09726 and Marie Curie grants FP7-PEOPLE-2011-RG-294262/294096 to A.S.-A. and S.M.-F.; Swiss National Science Foundation 310000-120724/1, 32003BB_135712/1 and Swiss Cancer League KLS-02398-02-2009 grants to R.C.S.; A.S.-A. received a Research Fellowship from the European Hematology Association. S.M.-F. is supported in part by an International Early Career Scientist grant of the Howard Hughes Medical Institute.
Author information
Authors and Affiliations
Contributions
L.A. designed and performed most experiments and analyses, prepared figures and wrote the manuscript. A.S.-A. performed in vivo transplantation assays. D.M.-P. performed qPCR, genome-wide expression analyses and together with X.L. provided technical assistance. J.I. performed neonatal cell isolation and culture. X.L. and A.T. performed histological analyses. S.M. provided human samples. Y.-S.T. and D.-M.L. provided Cxcl12-floxed mice. P.L., J.S. and R.C.S. provided samples, designed experiments and human studies. S.M.-F. designed the overall study, supervised the experiments and wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
Extended Data Figure 1 BM MSC reduction in compound mutant mice and recipients of mutant haematopoietic cells is not due to cell differentiation.
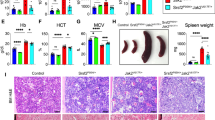
a, b, Blood counts of Nes-gfp;Mx1-cre;JAK2(V617F) and control mice 4-8 weeks after pIpC treatment (a) and Nes-gfp mice 10-12 weeks after transplantation with BM cells from MPN and control mice (b). Note the similar erythrocytosis, neutrophilia and thrombocytosis in compound mutant mice and Nes-gfp mice transplanted with mutant cells. Each dot represents a mouse. c, GFP fluorescence in skull BM of Nes-gfp mice 6–8 weeks after transplantation with MPN and control BM cells (n = 8; scale bar, 100 µm). d, Frequency of CD45−CD31−Ter119−Nes-GFP+ BM cells in MPN (n = 9) and control (n = 7) mice 6–8 weeks after pIpC induction. e, Giemsa-stained fibroblastic colony-forming units (CFU-F) from immunomagnetically-enriched BM CD45−Ter119− cells 30 weeks after transplantation with MPN and control BM cells. f, FACS analyses of BM CD45−CD31−Ter119− cells from MPN and control mice 8 weeks after pIpC treatment (n = 3). The specified MSC markers were used. Mean ± s.e.m. g, BM reticulin staining; MPN mice showed incipient fibrosis (arrow) 6 weeks after pIpC treatment (magnification, ×200). h, i, Lineage-tracing studies of BM nestin+ cells in MPN. h, Experimental design. Nes-creERT2;RCE:loxP mice were transplanted with MPN and control BM cells and fed with tamoxifen diet. Disease development was monitored over 28 weeks. i, Blood counts showing progressive neutrophilia and thrombocytosis in recipients of MPN BM cells (n = 3). j, Femoral haematoxylin and eosin stainings showing abnormal bone formation in recipients of MPN BM cells (magnification, ×100). *P < 0.05, **P < 0.01, ***P < 0.001 (unpaired two-tailed t test).
Extended Data Figure 2 Treatment with the JAK1/2 inhibitor ruxolitinib reduces haematopoietic cell expansion in MPN but it does not rescue BM MSCs.
Nes-gfp mice were transplanted with BM cells from pIpC-induced Mx1-cre;JAK2(V617F) mice and treated with ruxolitinib (n = 4) or vehicle (n = 6) over 8 weeks, starting 2 weeks after transplantation. a, Peripheral blood counts. b, Frequency of immunophenotypically-defined BM CD45−CD31−Ter119− cells. Mean ± s.e.m. *P < 0.05 (unpaired two-tailed t test).
Extended Data Figure 3 Depletion of nestin+ cells or their Cxcl12 production accelerates MPN.
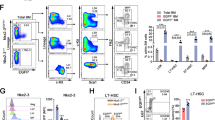
a, Experimental design for in vivo nestin+ cell depletion. Nes-creERT2;iDTA and control iDTA mice were transplanted with BM cells from pIpC-treated Mx1-cre;JAK2(V617F) and control mice and treated with tamoxifen (n = 3). b–e, Unlike Schwann cells, MSCs are reduced in Nes-creERT2;iDTA BM. b, c, Frequency of BM CD45−CD31−Ter119−CD90+ cells (b) and fibroblastic colony-forming units (CFU-F, c) in BM CD45−Ter119− cells from Nes-creERT2;iDTA and control mice after 4-week tamoxifen treatment. d, Expression of glial fibrillary acidic protein (Gfap) mRNA (normalized to Gapdh) in the BM of mice in a. e, Quantification of Gfap immunostaining of femoral BM from mice in b, c. f–k, Early mutant HSC expansion and mobilisation correlates with reduced Cxcl12 expression in BM nestin+ cells. f, g, BM lin−sca-1+c-kit+ (f) CD34−Flt3− long-term (LT) and CD34+Flt3− short-term (ST) HSCs, and CD34−Flt3− multipotent progenitors (MPP) and (g) frequency of haematopoietic colony-forming units in culture (CFU-C) from BM nucleated cells, spleen or peripheral blood from Nes-Gfp;Mx1-cre;JAK2(V617F) (n = 6) and control (n = 11) mice 6 weeks after pIpC treatment. h, Appearance (inset; scale bar, 1 cm) and haematoxylin and eosin stainings of spleens (magnification, ×100). i–k, Cxcl12 protein (i) and mRNA levels (j, k) in BM extracellular fluid (i), nucleated cells (j) and stromal Nes-GFP+ cells (k) of Mx1-cre;JAK2(V617F) and control mice 6 weeks after pIpC treatment (i, j) or Nes-gfp mice 10 weeks after transplantation with mutant BM (n = 4) or control cells (n = 7) (k). l, Circulating platelets in Nes-creERT2;Cxcl12fl/fl (n = 11) and control Cxcl12fl/fl littermates (n = 14) 18 weeks after transplantation with BM cells from MPN mice and 12 weeks after tamoxifen treatment. Mean ± s.e.m. *P < 0.05, **P < 0.01 (unpaired two-tailed t test).
Extended Data Figure 4 Neuroglial damage in MPN-affected BM stroma.
a–d, Expression of neuroglial gene sets in MPN BM Nes-GFP+ cells. a–c, Selected transcript expression in fragments per kilobase of exon per million fragments mapped (FPKM) of mesenchymal genes (a), HSC-niche related genes (b) and neural-related genes (c). RNA-Seq in CD45−CD31−Ter119−GFP+ cells isolated from compound MPN and control mice 6 weeks after pIpC treatment (samples pooled from 3 mice). d, Gene set enrichment analyses (GSEA) of RNA-Seq data. e, f, Enrichment plot of coordinated changes in neuroactive ligand-receptor interactions (e) and the transcriptome of neurons, astrocytes and oligodendrocytes (f). g, h, BM Nes-GFP+ cells are different from sympathetic nerve fibres and mature Schwann cells. Immunostaining of tyrosine hydroxylase (TH) to visualize sympathetic nerve fibres (g) and glial fibrillary acidic protein (GFAP) for mature Schwann cells (h) in Nes-gfp BM. Note the close association of Nes-GFP+ (green) cells to distinctive TH+ or GFAP+ (red) cells. i, BM biopsies from (left and middle) control or (right) MPN patients immunostained with (left) secondary Ab as negative control or (middle and right) anti-TH antibodies. Nuclei were counterstained with DAPI (blue). Scale bars, 75 µm (g), 50 µm (h), 100 µm (i).
Extended Data Figure 5 Contribution of IL-1β to early MPN pathogenesis.
a, Increased BM IL-1β levels early in MPN. Multiplex ELISA analysis of pro-inflammatory (IL-1β, TNF-α, IFN-γ, IL-6), regulatory (IL-17) and anti-inflammatory (IL-10) mediators in plasma (control, n = 16; MPN, n = 13) and BM extracellular fluid samples (control, n = 11; MPN, n = 6; only IL-1β and TNF-α were detectable) from control and Mx1-cre;JAK2(V617F) mice 4–8 weeks after pIpC treatment. b–f, Nes-gfp mice were transplanted with BM cells from MPN (n = 6) and control (n = 3) mice and analysed 2 weeks later. b–d, mRNA expression of IL-1β (b) and its activating enzyme caspase-1 (Casp1, c), and BM frequencies of lin−sca-1+c-kit+ haematopoietic progenitors and CD11b+Ly-6G(1A8)− monocytes (d). e, f, mRNA expression of IL-1 receptor (IL1r) (e) and its antagonist (IL1ra) (f) in BM CD45−CD31−Ter119−Nes-GFP+/− cells. g, Number of circulating platelets in WT mice transplanted with MPN and control BM cells and treated over 18 weeks with IL1ra, starting 2 weeks after transplant. h, Frequency of BM CD45−CD31−Ter119−CD90+ cells in mice in g. i, j, qPCR analyses of IL-1β (i) and Casp1 (j) mRNA expression in haematopoietic progenitors and monocytes isolated from the BM of mice in g. Gapdh was used as housekeeping gene (n = 5). Mean ± s.e.m. *P < 0.05, **P < 0.01, ***P < 0.001 (unpaired two-tailed t test).
Extended Data Figure 6 Compensatory treatment with β3-adrenergic agonist BRL37344 blocks MPN progression.
a, Expression of β2- and β3-adrenergic receptors in immunomagnetically-enriched CD45+ and CD45− BM cells from control and pIpC-induced Mx1-cre;JAK2(V617F) mice (n = 3). b-c, Blood counts of WT mice (b) 4 weeks after transplantation with MPN or control BM cells, before (control, n = 11; MPN, n = 13) and (c) 4-12 weeks after chronic BRL37344 (2 mg kg−1) or vehicle treatment (i.p., twice a day; n = 5). CD11b+ monocytes and granulocytes, B220+ B cells and CD3+ T cells were determined by flow cytometry. d, Blood counts of WT mice transplanted with control BM cells and treated with BRL37344 (n = 6) or vehicle (n = 4) as described above. Mean ± s.e.m. *P < 0.05, **P < 0.01, ***P < 0.001 (unpaired two-tailed t test).
Extended Data Figure 7 β3-adrenergic agonists or neuroprotective drugs prevent neuroglial gene induction in nestin+ cells and indirectly affect mutant haematopoietic cells.
a, Inhibition of mutant HSC expansion by β3-adrenergic agonists or neuroprotective drugs is HSC-niche dependent. Immunomagnetically-enriched CD45+ haematopoietic cells were obtained from the BM of Nes-gfp;Mx1-cre;JAK2(V617F) mice and control littermates 16 weeks after pIpC treatment (n = 3). BRL37344 (BRL) and 4-methylcatechol (4MC) were added in vitro at the indicated concentrations and the frequency of colony-forming units in culture (CFU-C) was scored after one week in culture. b, Compensatory treatment with the human β3-adrenergic agonist Mirabegron delays MPN in mice. Blood counts of WT mice 8 weeks after transplantation with MPN or control (n = 5) BM cells. Mirabegron (2 mg kg−1, n = 8) or vehicle (n = 7) treatment (i.p., twice a day) was administered the last two weeks. c, BM Nes-GFP+ cells from MPN mice treated with BRL37344 do not express neuroglial genes; mRNA expression of the Schwann cell markers myelin basic protein (Mbp), myelin-associated oligodendrocyte basic protein (Mobp) and pleckstrin homology domain containing, family B (evectins) member 1 (Pleckhb1) in BM CD45−CD31−Ter119−GFP+ cells sorted from Nes-gfp mice treated over 8 weeks with BRL37344 or vehicle, starting 4 weeks after transplantation with MPN or control BM cells (n = 4). Mean ± s.e.m. *P < 0.05, **P < 0.01, ***P < 0.001 (unpaired two-tailed t test).
Extended Data Figure 8 Incipient MPN signs are improved by BRL37344 or 4-methylcatechol.
a, b, Haematological parameters of WT mice 8 weeks after transplantation with BM cells from pIpC-treated control and Mx1-cre;JAK2(V617F) mice. The latter received the neuroprotective drug 4-methylcatechol (10 µg kg−1, once daily), BRL37344 (2 mg kg−1) or vehicle injections (twice a day) for the last 4 weeks (i.p.; n = 5). MPN progression was monitored in peripheral blood and mice were killed when they showed only incipient symptoms. a, Blood counts, spleen size, weight and nucleated cell number, BM nucleated cells (limbs and sternum), CD11b+ myeloid, B220+ B-lymphoid and CD3+ T cells. b, BM lin−sca-1+c-kit+ (LSK) haematopoietic progenitors, Ter119+CD71− mature erythroblasts and CD41+/CD42+ megakaryocyte progenitors. Mean ± s.e.m. *P < 0.05, **P < 0.01, ***P < 0.001 (unpaired two-tailed t test).
Extended Data Figure 9 Long-term BRL37344 treatment inhibits JAK2(V617F)+ HSC expansion but it does not compromise normal haematopoiesis.
a, b, Haematopoietic progenitors quantitated by FACS (a) or colony-forming units (CFU-C) (b) in BM and blood of WT mice 12-16 weeks after transplantation with MPN (a) and WT BM cells (b), 16 weeks after BRL37344 (2 mg kg−1) or vehicle injections (twice a day, i.p.; n = 3). Mean ± s.e.m. *P < 0.05 (unpaired two-tailed t test). c, BRL37344 treatment does not change normal HSC BM number. CD45.2 WT mice were transplanted with BM cells from congenic CD45.1 WT mice, together with limiting numbers of BM cells from CD45.2 control mice treated with vehicle/BRL37344 over 4 weeks, starting 11 weeks after the transplant (n = 5). The frequency of mice that failed reconstitution 16 weeks after transplantation is plotted against the number of tested cells. HSC frequencies are indicated; n.s., non-significant; Pearson chi-squared t test.
Extended Data Figure 10 Differentially expressed genes in BM CD45−CD31−Ter119− GFP+ cells from Nes-gfp;Mx1-cre;JAK2(V617F) mice.
a, b, Top most upregulated (a) and downregulated (b) genes in RNaseq analyses of BM CD45−CD31−Ter119− GFP+ cells from Nes-gfp;Mx1-cre;JAK2(V617F) mice (red) versus control mice (blue). Plots represent differentially expressed genes in fragments per kilobase of exon per million fragments mapped (FPKM).
Rights and permissions
About this article

Cite this article
Arranz, L., Sánchez-Aguilera, A., Martín-Pérez, D. et al. Neuropathy of haematopoietic stem cell niche is essential for myeloproliferative neoplasms. Nature 512, 78–81 (2014). https://doi.org/10.1038/nature13383
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature13383
This article is cited by
-
Profound sympathetic neuropathy in the bone marrow of patients with acute myeloid leukemia
Leukemia (2024)
-
Biological drivers of clinical phenotype in myelofibrosis
Leukemia (2023)
-
The roles of bone remodeling in normal hematopoiesis and age-related hematological malignancies
Bone Research (2023)
-
Extracellular vesicle-mediated remodeling of the bone marrow microenvironment in myeloid malignancies
International Journal of Hematology (2023)
-
A mysterious triangle of blood, bones, and nerves
Journal of Bone and Mineral Metabolism (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
