
Abstract
Antibody class-switch DNA recombination (CSR) is initiated by AID-introduced DSBs in the switch (S) regions targeted for recombination, as effected by Ku70/Ku86-mediated NHEJ. Ku-deficient B cells, however, undergo (reduced) CSR through an alternative(A)-NHEJ pathway, which introduces microhomologies in S–S junctions. As microhomology-mediated end-joining requires annealing of single-strand DNA ends, we addressed the contribution of single-strand annealing factors HR Rad52 and translesion DNA polymerase θ to CSR. Compared with their Rad52+/+ counterparts, which display normal CSR, Rad52−/− B cells show increased CSR, fewer intra-Sμ region recombinations, no/minimal microhomologies in S–S junctions, decreased c-Myc/IgH translocations and increased Ku70/Ku86 recruitment to S-region DSB ends. Rad52 competes with Ku70/Ku86 for binding to S-region DSB ends. It also facilitates a Ku-independent DSB repair, which favours intra-S region recombination and mediates, particularly in Ku absence, inter-S–S recombination, as emphasized by the significantly greater CSR reduction in Rad52−/− versus Rad52+/+ B cells on Ku86 knockdown.
Similar content being viewed by others

Introduction
Immunoglobulin (Ig) class-switch DNA recombination (CSR) and somatic hypermutation (SHM) are central to the maturation of the antibody response1,2,3,4. CSR endows antibodies with new biological effector functions by exchanging the gene encoding the Ig heavy chain constant region (CH) with a downstream CH region. By introducing mainly point mutations in Ig V(D)J sequences, SHM provides the structural substrate for antigen-mediated selection of higher-affinity antibody mutants1,2,4. Similar to SHM, CSR requires activation-induced cytidine deaminase (AID)-mediated generation of DNA lesions1,2,4. AID, expressed in activated B cells, deaminates deoxycytosines to yield deoxyuridine:deoxyguanine mispairs2,5. These mispairs trigger DNA repair processes that lead to insertion of double-strand DNA breaks (DSBs) in the upstream (donor) and downstream (acceptor) switch (S) regions (CSR)2,6. Synapse of a S region, such as Sμ, DSB ends with DSB ends of a downstream S region, such as Sγ1, leads to deletion of the intervening DNA which is released as extrachromosomal S circle, and juxtaposition of a VHDJH exon to a downstream CH exon cluster (in the above case Cγ1), thereby completing the CSR process2,4. Other outcomes can occur. Multiple DSBs are introduced into each of the S regions that will be the targets of recombination—Sμ being particularly prone to accumulating many DSBs. DSBs in a given S region can synapse with DSBs within the same S region, thereby yielding intra-S region deletions and non-CSR events. S-region DSB ends can also recombine with the DSB ends in other chromosomes to yield translocations, including c-Myc/IgH translocations7.
Synapsis of DSBs is generally effected by two major DNA repair pathways: non-homologous end-joining (NHEJ) or homologous recombination (HR). Unlike HR, which involves substantial DSB resection yielding overhangs with extensive sequence complementarity8,9, NHEJ synapses DSBs which have blunt/virtually blunt ends10,11. NHEJ entails recruitment of Ku70/Ku86 heterodimer, which, after binding to DSB ends, activates DNA-dependent protein kinase DNA-PKcs. This recruits the XRCC4/XRCC4-like factor/Ligase IV (Lig4) complex to complete the end-joining process10,12,13. Ku70/Ku86-mediated NHEJ plays an important role in recombining an upstream with a downstream S region, leading to CSR2,12,14,15. Substantial CSR, however, occurs in the absence of critical NHEJ components (Ku70/Ku86, XRCC4 or Lig4), suggesting that a Ku-independent or alternative NHEJ (A-NHEJ) DSB synapse also plays a role in CSR12,16,17,18. Although the nature of such a Ku-independent process remains to be defined, the extensive microhomologies at S–S junctions12,16,17,18,19 indicate that the CSR A-NHEJ involves microhomology-mediated end-joining (MMEJ). Microhomology-mediated A-NHEJ depends on moderate resection of DSBs and annealing of complementary single-strand DNA overhangs9,20,21. It provides the junctional mechanism effecting inter-chromosomal translocations12,22 and can be repressed by the NHEJ machinery (Ku70/Ku86, Lig4 or XRCC4)23,24,25,26,27.
In CSR and possibly in c-Myc/IgH inter-chromosomal translocations, A-NHEJ is initiated by the DNA damage sensor Parp1 and an early HR element, the DSB end-processing factor CtIP, which facilitates DSB resection to generate protruding (‘staggered’) ends28,29. These are annealed through stretches of complementarity21,27, which leads to introduction of microhomologies at S–S (and c-Myc-IgH) junctions16,18,29. We contend here that microhomology-mediated A-NHEJ in CSR and c-Myc/IgH translocations also critically relies on another HR factor Rad52, a DNA-binding element that promotes annealing of complementary DSB single-strand ends8,30,31. Rad52 plays a central role in HR DSB repair and is also involved in HR-independent DSB repair32. We previously showed that Rad52 is recruited together with Rad51, another HR factor, to AID-resected DSB protruding ends (Rad51 recruitment to DNA DSBs is dependent on Rad52) in the human IgH locus during antibody diversification33. In addition to Rad52, the translesion DNA polymerase θ (Polθ), which promotes annealing of complementary single DNA strands, may also be involved in Ku-independent CSR. Polθ facilitates MMEJ34,35,36. It bypasses lesions by inserting and extending past mispairs. Polθ also copies an undamaged DNA template efficiently but in an error-prone manner and, as we have shown, plays a significant role in Ig locus SHM37. Finally, Polθ can mediate DSB synapses in chromosomal translocation and is essential for cell survival when HR is impaired35.
Here we addressed the role of single-strand DNA annealing factors Rad52 and Polθ in CSR. Using Rad52−/− and Polθ−/− B cells in vitro together with molecular genetic methods, we found that Rad52 deficiency profoundly altered CSR to all Ig classes, whereas Polθ deficiency did not. We validated the in vitro findings by analysing specific class-switched antibody responses in Rad52−/− and Polθ−/− mice. We adapted chromatin immunoprecipitation (ChIP) and competition assays involving recombinant proteins, to analyse the recruitment of Rad52 (RAD52) and Ku70/Ku86 (KU70/KU86) to CSR-targeted S-region DSB ends. We studied the expression kinetics of Rad52 (RAD52) and Polθ (POLθ), and compared them with that of Ku70/Ku86 (KU70/KU86) in mouse and human B cells on exposure to Aicda (AICDA)/CSR-inducing stimuli. Further, we determined the impact of Rad52 deficiency on c-Myc/IgH translocations associated with CSR in p53−/− B cells—the p53 tumour suppressor is essential for protecting B cells from c-Myc/IgH translocations and p53 deficiency significantly enhances AID-dependent c-Myc/IgH translocations without detectable effect on CSR38. Finally, we enforced Rad52 expression in normal B cells and knocked down Ku86 by specific Ku86 short hairpin RNA (shRNA) in normal and Rad52−/− B cells, to assess the reciprocal contribution of Rad52 and Ku proteins to CSR. Our findings show that Rad52 competes with Ku70/Ku86 for binding to S-region DSB-free ends to modulate CSR by facilitating a microhomology-mediated S-region DSB A-NHEJ synaptic process. This favours intra-S region recombination, but also mediates Ku-independent inter-S–S region DSB recombination.
Results
Rad52−/− B cells increase CSR
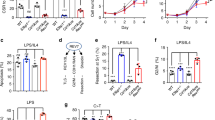
To test the hypothesis that Rad52, a DNA single-strand annealing (SSA) and HR factor, plays a role in CSR, we analysed B cells from Rad52−/− C57BL/6 mice and Rad52+/+ C57BL/6 littermates for their ability to undergo CSR. Previous report on targeted inactivation of Rad52 showed that Rad52 knockout reduced HR and stated that ‘Rad52 is not necessary for CSR39. Data, however, were not presented to support the latter claim, which was putatively based on Sμ-Sɛ recombination only. We stimulated Rad52−/− and Rad52+/+ B cells, as well as Aicda−/− and Aicda+/+ B cells with lipopolysaccharide (LPS; to induce CSR to IgG3), mCD154 or LPS plus interleukin (IL)-4 (IgG1), LPS plus interferon (IFN)-γ (IgG2c) and LPS plus transforming growth factor (TGF)-β, IL-5, IL-4 and anti-δ monoclonal antibody/dex (IgA). After 96 h of culture, the proportion of surface IgG1+, IgG3+, IgG2c+ and IgA+ B cells among Rad52−/− B cells was ∼1.9-, 1.6-, 2.1- and 2.0-folds of those among Rad52+/+ B cells (Fig. 1a). This reflected an effective increase of CSR in Rad52−/− B cells, as these completed the same number of divisions as their Rad52+/+ counterparts in response to different doses of LPS or mCD154 plus IL-4, while yielding a >50% increase in switched IgG1+ B cells (Fig. 1b,c). Increased Rad52−/− B cell CSR was confirmed by detection of recombinant Sμ–Sγ1 or Sμ–Sγ3 DNA (semi-quantitative digestion–circularization PCR; Fig. 1d). Increased Rad52−/− B-cell CSR was not associated with changes in germline intervening-constant heavy chain region (IH–CH) transcripts or AID, Ku70/Ku86 and Polθ levels, as shown by unchanged Iγ1-Cγ1, Iγ3-Cγ3, Aicda, Ku70/Ku86 and Polθ transcripts (real-time quantitative reverse transcriptase–PCR (qRT–PCR)) in Rad52−/− B cells, after a 60 h culture with LPS plus IL-4 or LPS alone (Fig. 1e), as well as unchanged AID, Ku70/Ku86 and Polθ protein levels (immunoblotting) after a 72 h culture with LPS plus IL-4 (Fig. 1f). This contrasted with the increased circle Iγ1-Cμ and Iγ3-Cμ transcripts, and post-recombination Iμ-Cγ1 and Iμ-Cγ3 transcripts, which are generated only after completion of CSR. Thus, intrinsic B-cell Rad52 deficiency increases CSR in the presence of normal levels of germline IH–CH transcripts and Aicda, Ku70/Ku86 and Polθ expression, as well as normal cell division rates.

(a) B cells purified from Rad52+/+ and Rad52−/− C57BL/6 littermates were stimulated with mCD154 or LPS plus IL-4 (for CSR to IgG1), LPS alone (IgG3), LPS plus IFN-γ (IgG2c) or LPS plus TGF-β, IL-4, IL-5 and anti-δ mAb/dex (IgA). Purified Aicda+/+ and Aicda−/− B cells were stimulated with LPS plus IL-4. After 96 h of culture, the cells were analysed for surface IgG1, IgG3, IgG2c or IgA by flow cytometry. (b) Proliferation of Rad52+/+ and Rad52−/− B cells labelled with CFSE and stimulated for 72 h with different amounts of LPS or mCD154 plus IL-4. Progressive left shift of fluorescence intensity indicates B220+ B cell division. (c) Proliferation of Rad52+/+ and Rad52−/− B cells labelled with the cell division tracking fluorochrome CFSE and stimulated by mCD154 plus IL-4 or LPS plus IL-4 for 96 h. CFSE intensity and surface IgG1 expression analysed by flow cytometry. Proportion of surface IgG1+ B cells at each cell division indicated. P-values determined using a paired Student’s t-test. Data are from one representative (left panels of each condition) or mean±s.d. of three independent experiments (right panels of each condition). (d) Recombinant Sμ–Sγ1 or Sμ–Sγ3 DNAs analysed by digestion–circularization PCR (DC-PCR) using serially twofold diluted HindIII digested and T4 DNA ligase-ligated genomic DNA from Rad52+/+ or Rad52−/− B cells after stimulation with LPS or LPS plus IL-4 for 96 h. Gapdh was used as a control for ligation and DNA loading. Data are from one representative of three independent experiments. (e) Rad52+/+ and Rad52−/− B cells were cultured with LPS or LPS plus IL-4 for 60 h. Aicda, Rad52, Ku70, Ku86, Polθ, germline Iγ1-Cγ1 and Iγ3-Cγ3, circle Iγ1-Cμ and Iγ3-Cμ, and post-recombination Iμ-Cγ1 and Iμ-Cγ3 transcripts analysed by real-time qRT–PCR. Each sample was run in triplicate; expression normalized to Cd79b expression and depicted as relative to the expression in Rad52+/+ B cells, set as 1. Data are from three independent experiments involving three pairs of Rad52+/+ and Rad52−/− mice (mean±s.d.). P values determined using a paired Student’s t-test. (f) Expression of AID, Rad52, Ku70, Ku86, Polθ and β-Actin proteins in unstimulated Rad52+/+ and Rad52−/− B cells or Rad52+/+ and Rad52−/− B cells stimulated with LPS plus IL-4 for 72 h were analysed by immunoblotting. Data are from one representative of three independent experiments.
Rad52−/− B cells reduce S–S junctional microhomologies
Through NHEJ, which synapses blunt or virtually blunt DSB ends, B cells complete CSR, leading to S–S junctions with no or minimal overlapping sequences (microhomologies)12. In B cells deficient in Ku70/Ku86 or other key NHEJ factors, such as XRCC4 and Lig4, CSR is reduced but not abolished, with the residual switched B cells displaying increased frequency and length of microhomologies at S–S junctions12,16,17,18,19. As we hypothesized, such S–S junctional microhomologies would be introduced by annealing of DSB single-strand DNA overhangs, a function characteristic of Rad52 (refs 23, 27). If our hypothesis were correct, then the increased CSR in Rad52−/− B cells should result in decreased frequency and length of S–S junctional microhomologies. We analysed Sμ–Sγ1 junctions from Rad52−/− and Rad52+/+ B cells stimulated with LPS plus IL-4 for 96 h, as well as Sμ–Sα junctions ex vivo from B220+PNAhi germinal centre (GC) B cells in Peyer’s patches. As expected, the majority of Sμ–Sγ1 and Sμ–Sα junctions in Rad52+/+ B cells displayed microhomologies, with >73% of Sμ–Sγ1 and 77% of Sμ–Sα junctional microhomologies consisting of up to 14 nucleotides, and >36% of Sμ–Sγ1 and 46% of Sμ–Sα junctions with microhomologies of 4 nucleotides or more. The frequency and length of Sμ–Sγ1 and Sμ–Sα junctional microhomologies were significantly reduced in Rad52−/− B cells (Supplementary Figs 1 and 2, and Fig. 2). In these B cells, 57% of Sμ–Sγ1 and 60% of Sμ–Sα junctions contained no microhomology, and only 3.3% of Sμ–Sγ1 and none of Sμ–Sα junctions contained microhomologies of 4 nucleotides or more. Overall, the average microhomologous sequence decreased from 2.87 (±0.66) nucleotides (Sμ–Sγ1) and 3.70 (±0.74) nucleotides (Sμ–Sα) in Rad52+/+ B cells to 0.67 (±0.17) (Sμ–Sγ1, P<0.0001, paired Student’s t-test) and 0.63 (±0.21) (Sμ–Sα, P<0.0001, paired Student’s t-test) nucleotide in Rad52−/− B cells. The dramatic increase in S–S junctions without microhomologies and reduced length of S–S junctional microhomologies in Rad52−/− B cells suggested that the greater number of CSR events in these B cells reflected the activity of NHEJ, which relies on Ku70/Ku86 and introduces no or minimal microhomologies.

Histograms depict percentages of Sμ–Sγ1 junction sequences with indicated numbers of nucleotide overlaps in Rad52+/+ (n=30) and Rad52−/− B cells (n=30) stimulated with LPS plus IL-4 for 96 h (as in Supplementary Fig. 1), and Sμ–Sα junction sequences with indicated numbers of nucleotide overlaps (microhomologies) in Rad52+/+ (n=30) and Rad52−/− B cells (n=30) from the Peyer’s patches of three pairs of Rad52+/+ and Rad52−/− C57BL/6 littermates (as in Supplementary Fig. 2). 0 indicates no microhomology. The average length of nucleotide overlap and the numbers of sequences analysed (n) are indicated. P-values determined using a paired Student’s t-test.
Polθ−/− B cells display normal CSR and S–S microhomologies
The error-prone translesion DNA Polθ, which, as we have shown, plays a role in SHM37, can anneal and end-join DNA single-strand ends, leading to the introduction of microhomologies34,35,36, as in the CSR S–S region synaptic process40. To determine whether Polθ is actually involved in CSR, we stimulated Polθ−/− and Polθ+/+ B cells with LPS plus IL-4, LPS only, LPS plus IFN-γ and LPS plus TGF-β, IL-5, IL-4 and anti-δ monoclonal antibody/dex, to induce CSR to IgG1, IgG3, IgG2c and IgA, respectively. After 96 h of culture, the proportions of class-switched IgG1+, IgG3+, IgG2c+ and IgA+ Polθ−/− B cells were comparable to those of Polθ+/+ B cells, as were B-cell divisions and the proportion of class-switched cells/round of cell division (Fig. 3a,b). The normal CSR in Polθ−/− B cells was supported by the normal levels of Aicda expression, circle Iγ1-Cμ and Iγ3-Cμ transcripts, post-recombination Iμ-Cγ1 and Iμ-Cγ3 transcripts, as well as germline Iγ1-Cγ1 and Iγ3-Cγ3 transcripts (that is, comparable to Polθ+/+ B cells) (Fig. 3c). In addition, Polθ−/− B cells were comparable to Polθ+/+ B cells in Sμ–Sγ1 junction frequency of microhomologies and length (P=0.09, paired Student’s t-test) (Supplementary Fig. 3). Finally, that Polθ−/− B cells functioned normally in CSR was confirmed by analyses in vivo showing comparable levels of serum IgM, IgG1 and IgA, as well as IgG1+ B220+PNAhi GC B cells in the spleens from Polθ−/− and Polθ+/+ littermates that had been injected with NP16-CGG (Fig. 3d,e). Thus, the DNA single-strand annealing Polθ does not apparently play a role in CSR synapsing of S–S region DSBs.

(a) Polθ+/+ and Polθ−/− B cells were stimulated with LPS plus IL-4 (for CSR to IgG1), LPS alone (IgG3), LPS plus IFN-γ (IgG2c) or LPS plus TGF-β, IL-4, IL-5 and anti-δ mAb/dex (IgA). After a 96 h of culture, the B cells were analysed for surface IgG1, IgG3, IgG2c or IgA by flow cytometry. (b) Proliferation of Polθ+/+ and Polθ−/− B cells labelled with CFSE and stimulated by LPS plus IL-4 for 96 h. CFSE intensity and surface IgG1 expression analysed by flow cytometry. Proportion of surface IgG1+ B cells at each cell division indicated. P-values determined using a paired Student’s t-test. Data are from one representative of three independent experiments. (c) Polθ+/+ and Polθ−/− B cells cultured with LPS or LPS plus IL-4 for 60 h. Expression of Polθ, Aicda, germline Iγ1-Cγ1 and Iγ3-Cγ3, circle Iγ1-Cμ and Iγ3-Cμ, and post-recombination Iμ-Cγ1 and Iμ-Cγ3 transcripts analysed by qRT–PCR and normalized to Gapdh transcript, depicted as relative to the expression of each transcript in Polθ+/+ B cells, set as 1. Each sample was run in triplicate. Data are from three independent experiments involving three pairs of Polθ+/+ and Polθ−/− mice (mean±s.d.). P-values determined using a paired Student’s t-test. (d,e) Polθ+/+ and Polθ−/− littermates were injected with NP16-CGG and killed 10 days later. (d) Titres of circulating IgM, IgG1 and IgA analysed by enzyme-linked immunosorbent assay (ELISA), expressed as μgeq ml−1. P-values determined using a paired Student’s t-test. Data are from three independent experiments (mean±s.d.). (e) Surface IgG1 expression in spleen B220+PNAhi GC B cells analysed by flow cytometry. Data are from one representative of three independent experiments.
Rad52 deficiency increases class-switched antibody responses
To define the impact of Rad52 deficiency on CSR in vivo, we analysed the class-switched antibody response to 4-Hydroxy-3-nitrophenyl acetyl hapten (NP) in Rad52−/− mice and Rad52+/+ littermates after injection of NP-conjugated chicken gamma globulin (NP16-CGG), which preferentially induces T-dependent NP-specific IgG1 antibodies. Rad52−/− mice showed significantly higher titres of NP32-binding IgG1 and (high affinity) NP4-binding IgG1, as well as total IgG1 and IgA than their Rad52+/+ counterparts, in the presence of normal IgM levels (Fig. 4a). In addition, similar Rad52−/− mice showed significantly higher titres of total IgE than their Rad52+/+ littermates after injection of ovalbumin (OVA; Fig. 4b). In Rad52−/− mice, high IgG1 and IgA, and NP-binding IgG1 titres were associated with a >75% increase of IgG1+ B220+PNAhi GC B cells in the spleen (Fig. 4c) and a 100% increase in IgA+ B220+PNAhi GC B cells in Peyer’s patches. In Rad52−/− mice, the spleen size, and number and size of Peyer’s patches were comparable to those in their Rad52+/− and Rad52+/+ counterparts. Consistent with a previous report39, the number of B (B220+) and T (CD3+) cells, and the proportions of CD4+ and CD8+ T cells in Rad52−/− mice were comparable to those in their Rad52+/+ littermates. In addition, the proportion of B220+PNAhi GC B cells and B220low CD138+ plasma cells in the spleen were also comparable in Rad52−/− and Rad52+/+ mice; Rad52−/− mice showed normal B-cell viability (Fig. 4d), proliferation and normal cell cycle in total and GC B cells, as measured by bromodeoxyuridine (BrdU) incorporation and 7-aminoactinomycin D (7-AAD) staining (Fig. 4e). Thus, Rad52 deficiency leads to significantly increased class-switched antibody response without affecting B-cell proliferation or survival.

(a) Rad52+/+ and Rad52−/− littermates were injected with NP16-CGG and killed 10 days later. Titres of circulating total IgM, IgG1 and IgA, NP32-binding and (high affinity) NP4-binding IgG1 analysed by enzyme-linked immunosorbent assay (ELISA), expressed as μgeq ml−1 or number of dilutions needed to reach 50% of saturation binding (relative units, RU). (b) Rad52+/+ and Rad52−/− littermates were injected twice with OVA at day 0 and day 7. Titres of circulating total IgE analysed by ELISA at day 7 (before the second OVA injection) and day 13, expressed as ngeq ml−1. Each symbol represents an individual mouse, n=5 or 6 pairs of mice. P-values determined using a paired Student’s t-test. (c) Surface IgG1 expression in spleen B220+PNAhi GC B cells or IgA expression in Peyer’s patch B220+PNAhi GC B cells of NP16-CGG-injected Rad52+/+ and Rad52−/− littermates analysed by flow cytometry. (d) Flow cytometry analysis of spleen cells from NP16-CGG-injected Rad52+/+ and Rad52−/− littermates for the proportion of: B220+ B cells and CD3+ T cells, CD4+ and CD8+ T cells, B220+PNAhi GC B cells, B220loCD138+ plasma cells and viable (7-AAD–) B220+ B cells. (e) Rad52+/+ and Rad52−/− littermates were injected with NP16-CGG. Ten days after the injection, the mice were injected i.p. with BrdU twice within a 16 h interval and killed 4 h after the last injection. Left panels: proliferating B cells (BrdU-stained B220+ B cells) analysed by flow cytometry; middle and right panels: in vivo cell cycle analysis (quadrant corresponding to the G0/G1, S and G2/M phase of the cell cycle) of B220+ B cells and B220+PNAhi GC B cells. Data are from one representative of three independent experiments.
Rad52 competes with Ku70/Ku86 for binding to S-region DSB ends
The increased CSR in Rad52−/− B cells in vitro and in vivo suggested that Rad52 is recruited to S-region DSB-ends where it would compete with Ku70/Ku86 to modulate CSR. To demonstrate that Rad52 and Ku70/Ku86 indeed bind to CSR-targeted S regions, we performed ChIP assays with anti-Rad52 antibody and anti-Ku70/Ku86 monoclonal antibodies. These showed that similar to Ku70/Ku86, Rad52 was specifically recruited to Sμ and Sγ1 regions, not Sγ3, in B cells stimulated by LPS plus IL-4 (undergoing CSR to IgG1), and to Sμ and Sγ3, not Sγ1, in B cells stimulated by LPS (CSR to IgG3). Rad52 and Ku70/Ku86 could be readily detected on Sμ, and Sγ1 or Sγ3, but not Cμ region, in Aicda+/+ B cells stimulated with LPS plus IL-4 or LPS alone, but failed to associate with such S regions in similarly activated Aicda−/− B cells, showing that similar to Ku70/Ku86, recruitment of Rad52 to CSR-targeted S regions was dependent on S region AID processing (Fig. 5a,b). To analyse the impact of Rad52 on Ku70/Ku86 recruitment to CSR-targeted S regions, we performed ChIP assays using anti-Ku70/Ku86 monoclonal antibody in Rad52+/+ and Rad52−/− B cells stimulated with LPS plus IL-4 or LPS. We complemented this approach with in situ DNA end-labelling by biotin-16-dUTP (bio-dUTP)41 followed by ChIP with anti-Ku70/86 monoclonal antibody or anti-Rad52 antibody and capture of biotin-labelled DNA fragments with streptavidin magnetic beads. This approach, which allows for detection of Ku70/Ku86 or Rad52 on DSB ends, showed that Rad52 indeed bound to the free ends of DSBs in the CSR-targeted S regions, but not to Cμ region or S regions not involved in CSR. In the absence of Rad52, Ku70/Ku86 bound to CSR-targeted Sγ1 and Sγ3 DSB free ends, up to 7.6-fold more than in the presence of Rad52 (Fig. 5c,d).

(a) Ku70/Ku86 and Rad52 are recruited to CSR-targeted S region DNA in an AID-dependent manner in B cells undergoing CSR. Aicda+/+ and Aicda−/− B cells were stimulated with LPS or LPS plus IL-4 for 60 h. Cross-linked chromatin was precipitated using rabbit anti-Rad52 antibody or mouse anti-Ku70/Ku86 mAb. Precipitated Sμ, Sγ1 and Sγ3 DNA quantified by real-time quantitative PCR (qPCR); amounts relative to those in Aicda+/+ B cells, set as 1. Each precipitated DNA sample was run as triplicate in qPCR; the average of each triplicate was used as data point for that individual sample. Data are from three independent experiments involving three pairs of Aicda+/+ and Aicda−/− mice (mean±s.d.). P-values determined using a paired Student’s t-test. (b) Precipitated Sμ and Cμ DNA from Aicda+/+ and Aicda−/− B cells detected by PCR. Data are one representative of three independent experiments. (c) Rad52+/+ and Rad52−/− B cells were stimulated with LPS or LPS plus IL-4 for 60 h. Chromatin was cross-linked and precipitated using a mouse anti-Ku70/Ku86 mAb. The precipitated Sμ, Sγ1 and Sγ3 DNAs quantified by real-time qPCR; amounts in Rad52−/− B cells are as relative to those in Rad52+/+ B cells, set as 1. Each precipitated DNA sample was run as triplicate in qPCR; the average of each triplicate was used as data point for that individual sample. Data are from three independent experiments involving three pairs of Rad52+/+ and Rad52−/− mice (mean±s.d.). P-values determined using a paired Student’s t-test. (d) Rad52+/+ and Rad52−/− B cells were stimulated with LPS or LPS plus IL-4 for 60 h. DNA ends were labeled in situ with bio-dUTP using TdT. Chromatin was cross-linked and precipitated using mouse anti-Ku70/Ku86 mAb (left panel) or rabbit anti-Rad52 antibody (right panel). After resuspension, DNA with broken ends was pulled down with streptavidin magnetic beads. Precipitated Sμ, Sγ1, Sγ3 and Cμ DNAs quantified by real-time qPCR (this approach allowed for detection of Ku70/Ku86 or Rad52 bound to DSB-free ends); amounts of precipitated DNA are relative to those in Rad52+/+ B cells, set as 1 (left panel), or relative to respective input DNA (right panel). Each precipitated DNA sample was run as triplicate in qPCR; the average of each triplicate was used as data point for that individual sample. Data are from three independent experiments involving three pairs of Rad52+/+ and Rad52−/− mice (mean±s.d.). P-values determined using a paired Student’s t-test. (e,f) EMSA using a biotin-labelled double-stranded Sμ DNA probe and (e) recombinant human RAD52 and KU70/KU86 (constant amount) proteins at different ratios (1:1, 2:1 and 4:1), or (f) nuclear extracts (10 μg protein) from Rad52+/+ and Rad52−/− B cells (stimulated with LPS plus IL-4 for 60 h) incubated with mouse anti-Ku70/Ku86 mAb or irrelevant mouse IgG (control). The formation of protein–DNA complexes was shifted by anti-Ku70/Ku86 mAb. Numbers below gel image indicate relative density of protein–DNA complex bands normalized with the density of free probe. Shown is one representative gel of three independent experiments.
To prove that Rad52 competes with Ku70/Ku86 for binding to S-region DSB ends, we incubated recombinant human RAD52 and/or KU70/KU86 proteins with an 84 bp biotin-labelled double-strand Sμ probe and then submitted the reactants to electrophoretic mobility shift assays (EMSAs). Incubation of KU70/KU86 or RAD52 with biotin-Sμ DNA probe alone gave rise to a KU70/KU86 or RAD52 protein–Sμ DNA complex (Fig. 5e). Incubation of increasing amounts of recombinant human RAD52 protein with the same amount of KU70/KU86 led to increasing binding of RAD52 and decreasing binding of KU70/KU86 to biotin-Sμ DNA probe, indicating that RAD52 competed with KU70/KU86 for binding to Sμ DSB ends. Further, incubation of cell extracts from Rad52+/+ B cells stimulated with LPS plus IL-4 with the biotin-Sμ DNA probe gave rise to protein–DNA complexes containing Ku70/Ku86, as determined by supershift with an anti-Ku70/Ku86 monoclonal antibody (Fig. 5f). Such DNA–Ku70/Ku86 complexes were increased by about twofold in Rad52−/− B cells—relative density of the protein–DNA complex bands changed from 1.27 to 2.49. Thus, Rad52 competes with Ku70/Ku86 for binding to CSR-targeted S-region DSB ends.
Rad52−/− B cells reduce c-Myc/IgH translocations
Not only does CSR introduce DSBs in S regions but it can also instigate translocations between S regions on mouse chromosome 12 and exon 1 of the c-Myc gene on chromosome 15 (ref. 12). In addition to decreasing CSR, deficiency of critical NHEJ elements, such as XRCC4, Ku70 or combined Ku70 and Lig4, results in greatly increased c-Myc/IgH translocations, as mediated by A-NHEJ16,17,25,42. Such translocations are characterized by microhomologies at c-Myc-IgH junctions, the expression of a DSB end-joining process initiated by annealing of DSB single-strand DNA overhangs. We reasoned that in Rad52−/− B cells, increased CSR, a reflection of the enhanced recruitment of Ku70/Ku86 to S-region DSBs and increased NHEJ activity, would be associated with a decreased frequency of c-Myc/IgH translocations, possibly including only limited microhomologies, as a result of lack of Rad52 single-strand DNA annealing activity. To prove that Rad52 deficiency has a negative impact on c-Myc/IgH translocations, we bred p53−/− mice with Rad52+/– mice to generate p53−/−Rad52+/+ and p53−/−Rad52−/− mice—p53 deficiency significantly enhances AID-dependent c-Myc/IgH translocations but has no detectable effect on CSR38. We stimulated p53−/−Rad52+/+ and p53−/−Rad52−/− B cells with LPS plus IL-4. After 96 h of culture, we detected c-Myc/IgH translocations by long-range PCR and confirmed their identity by Southern blot hybridization (using c-Myc and IgH probes) and sequencing. In p53−/−Rad52−/− B cells, c-Myc/IgH translocations were induced at a frequency (1.2 × 10−7 translocation per cell) about threefold lower than in p53−/−Rad52+/+ B cells (3.2 × 10−7 translocation per cell, P=0.011, paired Student’s t-test) and comprised a lower number of microhomologies at c-Myc–IgH junctions (P=0.0014, paired Student’s t-test; Fig. 6). Thus, the greatly reduced frequency of c-Myc/IgH translocations in p53−/−Rad52−/− B cells undergoing CSR supports a role of Rad52 in the single-strand DNA annealing process in inter-chromosomal translocations.

B cells from p53−/−Rad52+/+ and p53−/−Rad52−/− mice were stimulated with LPS plus IL-4 for 96 h before genomic DNA isolation. (a) c-Myc/IgH translocations were identified by amplifying DNA using long-range nested PCR involving primers specific to the IgH and c-Myc locus, and verified by Southern blot hybridization with an IgH or c-Myc-specific probe. Each PCR assay was performed using template DNA from 106 cells. Twenty-five amplicons from p53−/−Rad52+/+ B cells and 25 amplicons from p53−/−Rad52−/− B cells are shown. The frequencies of c-Myc/IgH translocations per cell are indicated below the gel images. PCR amplification products that can be detected by both IgH and c-Myc DNA probes were from c-Myc/IgH translocations: 8 of 25 and 3 of 25 amplicons from p53−/−Rad52+/+ and p53−/−Rad52−/− B cells, respectively, contained PCR amplification products from c-Myc/IgH translocations. (b) Sequences of the c-Myc/IgH translocation junctions. Amplified c-Myc–IgH junctional DNAs were cloned and sequenced. Each sequence is compared with germline c-Myc (above) and IgH (below) sequences. Microhomologies are bold and underlined. Sequences containing microhomologies are in blue. Point mutations are in red. Data are from three pairs of mice.
Rad52−/− B cells reduce intra-S region DNA recombination
AID generates multiple DSBs within the targeted S regions, many of which are rejoined or joined to other DSBs within the same S region12. Each S region consists of highly repetitive DNA motifs. These can give rise to complementary protruding ends in upstream and downstream DSBs that are suitable substrates for DNA annealing by Rad52. Subsequent synapse of such DSB ends leads to intra-S region recombination and deletion of intervening DNA (of variable length). All S regions include characteristic and highly repetitive motifs. Such highly repetitive motifs differ in both nature and frequency in different S regions. It follows that DSB protruding ends from two different S regions, such as Sμ and Sγ1, will encompass sequences of virtually no complementarity, making them poor substrates for Rad52-mediated complementary annealing than protruding ends from DSBs within the same S region. Indeed, Pustell Matrix dot plot analysis of human and mouse Sμ and Sγ1 revealed maximal sequence complementarity within the individual Sμ or Sγ1 region, in particular within their core region, and maximal lack of complementarity between these two S regions, in particular in their core sequences (Fig. 7a). Accordingly, the reduced CSR in Ku70 deficiency, which, as we have hypothesized, would result in increased Rad52 recruitment to S-region DSB-ends, was shown to be associated with significantly increased occurrence of intra-S region deletions, remnants of occurred intra-S region recombinations17.

(a) Each S region consists of highly repetitive motifs, which can facilitate the formation of microhomologies, in particular within the S region core. As the characteristically repetitive sequences are virtually unique to S regions, DSB ends in the same S region are better suited substrates for Rad52-mediated complementary DNA single-strand annealing than those in two different S regions, such as Sμ and Sγ1. Repetitive sequence elements in human and mouse Sμ and Sγ1 that can potentially form microhomologies were identified by Pustell Matrix dot plot using MacVector software and are depicted by small dots. Thick lines indicate the core regions of Sμ and Sγ1. (b) Schematic representation of the detection of intra-S region recombination (deletion) in Sμ region by PCR amplification. DNA-amplified sequences of Sμ region that underwent intra-S region DNA recombination are shorter than those of Sμ in the germline configuration. (c) Rad52+/+ and Rad52−/− B cells were stimulated with LPS plus IL-4 for 96 h. Sμ region DNA was amplified by nested PCR. PCR amplification products were then cloned into the TOPO cloning vector. Sμ region sequences from individual clones amplified by PCR and resolved through a 0.8% agarose gel. PCR amplification products smaller than that amplified from the germline Sμ region DNAs (indicated by arrows) are from Sμ region DNAs that underwent intra-S region recombination, thereby deleting variable lengths of DNA: 14 of 30 Sμ region DNAs in Rad52+/+ B cells and 4 out of 30 Sμ region DNAs in Rad52−/− B cells underwent intra-S region recombination. Data are from three pairs of Rad52+/+ and Rad52−/− C57BL/6 mice.
We hypothesize here that such increased intra-S region deletions are mediated by a DSB synaptic process involving Rad52. If our hypothesis is correct, then the absence of Rad52—which, as shown by the preceding experiments, led to increased recruitment of Ku70/Ku86 to S-region DSB ends and increased CSR—would result in reduction of intra-S region deletions. To test our hypothesis, we set up to assay for deletions within the Sμ region of genomic DNA from Rad52+/+ and Rad52−/− B cells activated for CSR to IgG1—intra-S region deletions occur far more frequently in Sμ than the other (downstream) S regions17—using specific primers flanking both sides of Sμ to amplify this DNA region. Sμ DNAs with internal deletions were detected by visualizing amplification products that were shorter than germline Sμ and then positively identified by DNA sequencing (Fig. 7b,c and Supplementary Fig. 4). This revealed that Rad52−/− B cells displayed a substantially lower frequency of intra-Sμ region deletions as compared with their Rad52+/+ counterparts (4 out of 30, that is, 13.3%, versus 14 out 30, that is, 46.7%, P=0.0006, paired Student’s t-test). Even taking into account a remote possibility that some of the S-region DNA deletions might have been stemmed from PCR amplification or propagation of S region-containing plasmids in bacteria, these findings indicate that the Rad52-mediated DSB synaptic process favours intra-S region over inter-S–S region recombination.
Rad52 and Ku86 reciprocally modulate CSR
To define whether, in addition to mediating intra-S region recombination, Rad52 can also mediate inter-S–S region recombination, we knocked down Ku86 in Rad52−/− and Rad52+/+ B cells using a pGFP-C-Ku86-shLenti lentiviral vector. Expression of a Ku86-specific shRNA reduced Ku86 protein level by 87% (average) (when taking into consideration the 80% lentiviral transduction efficiency in our experiments) without altering Rad52 protein expression (Fig. 8a)—a pGFP-C-scrambled-shLenti lentiviral vector, which did not alter Ku86 expression, was used as a control. The transduced B cells were cultured with LPS and IL-4 for 96 h before analysing CSR to IgG1. Similar to untransduced Rad52−/− B cells, pGFP-C-scrambled-shLenti lentiviral vector-transduced Rad52−/− B cells displayed a (54%) higher level of CSR to IgG1 (P=0.003, paired Student’s t-test) as compared with their Rad52+/+ counterparts (Fig. 1a,b). In similar Rad52−/− B cells, Ku86 knockdown by pGFP-C-Ku86-shLenti lentiviral vector resulted in a nearly tenfold reduction of CSR to IgG1, as compared with the twofold reduction in Rad52+/+ B cells transduced by the pGFP-C-Ku86-shLenti lentiviral vector—knockdown of Ku86 in Rad52+/+ B cells reduced CSR to an extent comparable to that reported for Ku70- or Ku86-deficient B cells18. The less profound CSR reduction by Ku86 knockdown in Rad52+/+ than Rad52−/− B cells reflected the contribution of Rad52 to inter-S–S region recombination in Rad52+/+ B cells—the residual 3.4% CSR in pGFP-C-Ku86-shLenti lentiviral vector-transduced Rad52−/− B cells was (probably) due to the residual (∼12%) functional Ku86 in these B cells (Fig. 8a). Although knockdown of Ku86 in Rad52−/− B cells virtually ablated CSR, knockdown of Ku86 in Rad52+/+ B cells resulted only in partial CSR reduction, thereby pointing at a contribution of Rad52 to inter-S–S region synapses in the absence of Ku86. Thus, Rad52 partially rescues CSR in B cells with a compromised Ku-dependent/NHEJ repair pathway.

Rad52−/− and Rad52+/+ B cells were transduced with pGFP-C-Ku86-shRNALenti lentiviral vector expressing Ku86-specific shRNA and GFP, or pGFP-C-scr-shRNALenti lentiviral vector expressing scrambled shRNA and GFP. Rad52+/+ B cells were activated with LPS for 12 h and transduced with empty pMIG-GFP retroviral vector or pMIG-GFP-Rad52 retroviral vector expressing recombinant Rad52. After lentiviral or retroviral transduction, B cells were cultured for 96 h with LPS plus IL-4. (a) B-cell expression of Ku86, Rad52 and β-Actin proteins analysed by immunoblotting. (b) Proportions of surface IgG1+ B cells among pGFP-C-scr-shRNALenti or pGFP-C-Ku86-shRNALenti lentiviral vector-transduced (B220+GFP+) Rad52−/− and Rad52+/+ B cells were analysed by flow cytometry. Data are from one representative of three independent experiments. (c) Proportions of surface IgG1+ B cells among empty pMIG-GFP or pMIG-GFP-Rad52 retroviral vector-transduced (B220+GFP+) Rad52+/+ B cells were analysed by flow cytometry. Data are from one representative of three independent experiments.
Our experiments have shown that in the absence of Ku86, Rad52 can mediate inter-S–S region synapses. In the presence of Ku70/Ku86, Rad52 would compete with this heterodimer for binding to S-region DSB ends, thereby skewing the S-region DSB synaptic process towards intra-S region recombination to the detriment of inter-S–S region recombination and CSR. To test the hypothesis that overexpression of Rad52 will further reduce recruitment of Ku70/Ku86 to S-region DSB ends and significantly dampen CSR, we transduced Rad52+/+Ku86+/+ (normal) B cells with pMIG-Rad52 or an empty pMIG retroviral vector as control, cultured them with LPS plus IL-4 for 96 h, before analysing CSR. Enforced Rad52 expression in Rad52+/+Ku86+/+ B cells increased Rad52 protein level by 2.4-fold without altering Ku86 protein expression and reduced by >65% CSR to IgG1 (Fig. 8a,c). Thus, Rad52 (in excess) competed with Ku70/Ku86 for binding to S-region DSB ends, thereby skewing the DSB synaptic process towards intra-S region recombination and significantly decreasing CSR.
Switching B cells decrease Rad52 and increase Ku70/Ku86
Given the reciprocal modulation of CSR by Rad52 and Ku70/Ku86, we hypothesized that, on exposure to CSR-inducing stimuli, B cells downregulate Rad52 and upregulate Ku70/Ku86 to reduce intra-S region and facilitate inter-S–S region recombination, thereby ensuring maximal CSR rates. Indeed, in B cells stimulated with increasing amounts of LPS and LPS or mCD154 plus IL-4, which induced Aicda and CSR, Rad52 transcript levels were reduced by 73%–96% (P<0.0001, paired Student’s t-test) after 24–48 h of culture (Fig. 9a). In contrast to Rad52, Ku70 and Ku86 transcripts were increased by two- to threefolds (P=0.00013 or P<0.0001, respectively, paired Student’s t-test) within 24 h and returned to baseline values by 48 h. Polθ followed the upregulation/downregulation kinetics of Ku70/Ku86. The expression of Rad52, Ku70/Ku86, AID and Polθ proteins followed tightly with the kinetic of expression of their corresponding transcripts. Rad52 protein was downregulated on stimulation by LPS plus IL-4, whereas Ku70/Ku86 and Polθ proteins were upregulated. Similar to mouse B cells, human B cells also displayed a reciprocal modulation of RAD52 and POLθ, KU70, KU86 by AICDA/CSR-inducing stimuli (mCD154 plus IL-4 and IL-21), with the exception of a delayed recovery of RAD52 expression (Fig. 9b). Thus, B cells induced to undergo CSR modulate Rad52 (RAD52) and Ku70/Ku86 (KU70/KU86) expression in a reciprocal manner at both transcription and protein levels, thereby skewing the S-region DSB synaptic process towards inter-S–S region recombination.

(a) C57BL/6 B cells were stimulated with different amounts of LPS only, or LPS or mCD154 plus IL-4 for 0, 24, 48 and 72 h. Aicda, Rad52, Polθ and Ku70/Ku86 transcripts were analysed by real-time qRT–PCR. Expression of Aicda normalized to Cd79b expression, expression of Rad52, Ku70/Ku86 and Polθ normalized to Gapdh expression and depicted relative to the expression in unstimulated B cells, set as 1. Expression of AID, Rad52, Ku70, Ku86, Polθ and β-Actin proteins in B cells stimulated with LPS (3.0 μg ml−1) plus IL-4 were analysed by immunoblotting. (b) Human naive B cells were stimulated with different amounts of mCD154 plus human IL-4 and human IL-21 for 0, 24, 48, 72 or 96 h. AICDA, RAD52, KU70/KU86 and POLθ transcripts were analysed by real-time qRT–PCR. Expression normalized to CD79b expression and depicted relative to the expression in unstimulated B cells, set as 1. P-values determined using a paired Student’s t-test. Data are from one representative of three independent experiments.
Discussion
Resolution of DSBs is a highly coordinated process that uses the DNA repair machinery to maintain genomic integrity. DSBs are generally repaired by HR or NHEJ—NHEJ is also referred to as classical-NHEJ. HR accurately repairs staggered post-replicative DSBs using a long, homologous single-strand template in the form of a sister chromatid, whereas NHEJ fuses blunt or virtually blunt DSB ends that lack substantial joining complementarity, to form ‘direct’ junctions10,12. Rad52 and Ku70/Ku86 are critical and early elements in HR and NHEJ DSB, respectively. Both Rad52 and Ku70/Ku86 bind to DSB ends and facilitate end-to-end interactions43. Rad52 binds to and wraps around resected DSBs, thereby efficiently promoting annealing of complementary DNA single strands31, although it can also bind to blunt DSB ends43,44,45. Ku70/Ku86 binds to blunt DSB ends and gives rise to ‘clean’ (that is, free of microhomologies) DSB junctions, although it can also bind, albeit less efficiently, to DSBs with relatively short protruding ends44,46.
Efficient inter-S–S region recombination that leads to CSR requires DSB resolution by NHEJ, which recombines S-region DSBs to form S–S junctions with no or minimal microhomologies10,11,12. Substantial CSR, however, occurs in the absence of core NHEJ components by A-NHEJ, which yields S–S junctions with microhomologies12,16,17,18. In addition to CSR, microhomology-mediated A-NHEJ would also effect oncogenic inter-chromosomal translocations, which are indeed characterized by DSB junctions with significant levels of microhomologies8,25. Here we show that the SSA and HR element Rad52 plays a central role in facilitating the A-NHEJ DSB synaptic process underpinning CSR, as well as t(15;12) c-Myc/IgH translocation. Rad52 functions as a recombination mediator and is involved in maintenance of genomic integrity. It interacts with replication protein A47, another single-strand DNA-binding protein, which plays a role in CSR. Replication protein A associates preferentially with resected single-strand DNA ends created by AID, thereby helping stabilize AID itself48.
Extensive resection of DSBs yielding long overhangs (200 nt to 2–4 kb)8,9 leads to HR, whereas less extensive DSB resection (generally >30 but <200 nt)20,32 promotes what is referred to as SSA pathway, which is mediated by Rad52 but does not require a sister chromatid template49,50. More limited (<30 nt) DSB resection facilitates single-strand DNA annealing leading to MMEJ9,20,48, suggesting that the distinction between SSA and MMEJ is based on the length of the complementary protruding ends involved in the synaptic process. The requirement for Rad52 in SSA but not MMEJ has been claimed to be another criterion to distinguish SSA from MMEJ21. Data in fission yeast, however, suggest an extensive overlap between SSA and MMEJ, as effected by Rad22, the fission yeast homologue of Rad52 (refs 23, 27). Other reports have attributed contradictory roles to Rad52 in MMEJ. Rad52 has been shown to suppress MMEJ in budding yeast, possibly by promoting Rad51 filament formation and thereby HR51,52, but it has also been shown to be required for MMEJ in both budding and fission yeast22,23,53. DSB junctional microhomologies in yku70/80Δrad52Δ budding yeast cells were thought to indicate an independence of MMEJ from Rad52. Such microhomologies, however, were likely to be introduced by Rad59, a Rad52 homologue that anneals single-strand DNAs with short homologous sequences23,27,45,54.
By virtue of its low-fidelity translesion DNA synthesis, Polθ plays an important role, as we have shown, in SHM37. Polθ−/− B cells were comparable to their Polθ+/+ counterparts in switching to IgG1, IgG3, IgG2c and IgA, as well as in frequency and extent of Sμ–Sγ1 junctional microhomologies. These findings confirm and extend a previous observation40, and further suggest that Polθ does not play a role in Ku-independent (microhomology-mediated) A-NHEJ of CSR. This is in spite of the ability of Polθ to promote annealing of resected DSBs and facilitate MMEJ32,34,35,36. The failure of Polθ deficiency to alter CSR would not necessarily rule out other contributions by Polθ to this process, such as insertion of mutations into inter-S–S junction surrounding areas, perhaps consistent with the upregulation of Polθ in induced B cells, as we have previously shown and confirmed here1,37. Such an upregulation of Polθ may be part of an overall ‘early’ repair response, which includes primarily Ku70/Ku86, to the DNA damage associated with CSR and probably SHM. The apparent lack of an obvious role of Polθ in CSR might also reflect a functional redundancy of Polθ in this process, as other DNA polymerases, such as human Polλ and Polβ, have been shown to be interchangeable in mediating DNA single-strand annealing and elongation55—Polλ in conjunction with Polδ also plays a role in Saccharomyces cerevisiae MMEJ52. Redundancy in A-NHEJ would not be limited to Polθ, as deficiency of Lig1 or Lig3 (the DNA ligases involved in A-NHEJ) alone did not affect CSR, possibly due to the functional interchangeability of these two ligases in the late stage of CSR56.
Unlike Polθ, Rad52 plays an important role in CSR. Similar to Ku70/Ku86, Rad52 was recruited to CSR-targeted S regions after AID processing. Increased CSR in the absence of Rad52 entailed enhanced recruitment of Ku70/Ku86 to S-region DSB ends, decreased intra-Sμ DSB recombinations, reduced frequency and length of microhomologies in S–S junctions, as well as decreased inter-chromosomal c-Myc/IgH translocations. All of these features are opposite to those in (NHEJ-deficient) Ku70−/− B cells, which showed increased intra-S region DSB recombination, increased frequency and length of microhomologies in S–S junctions, as well as increased inter-chromosomal c-Myc/IgH translocations16,17,18, and point at Rad52 as mediator of A-NHEJ in CSR (Supplementary Table 1)17,18. Thus, our findings argue for a role of Rad52 in a Ku-independent DSB synaptic process that gives rise to S–S junctions. These contain microhomologies, which are more abundant than those, if any, of Ku-dependent S–S synapses16,17,18.
Recruitment of Rad52 or Ku70/Ku86 to DSB ends would lead to different synaptic processes43. Rad52 preferentially synapsed DSB ends within the same S region, as emphasized by the greatly decreased intra-Sμ region deletions, which are marks of intra-S region DNA recombination or ‘rejoining’, in Rad52−/− B cells. Conversely, intra-S region deletions were found to be greatly increased in B cells lacking Ku70 or Ku70 and Lig4 (ref. 17), as well as in B cells lacking 53BP1, a DNA-damage response protein that favours long-range S–S recombination (leading to CSR), but prevents short-range rejoining of intra-S region DSBs, possibly because of its protection of DNA ends from resection57. Rad52 recruitment to CSR-targeted Sγ was as efficient as recruitment to Sμ region. However, because of the distinctive, albeit characteristically repetitive, sequences of the IgH locus S regions, DSB protruding ends in two different S regions, such as Sμ and Sγ1, are less suitable substrates for Rad52-mediated annealing than DSB protruding ends in the same S region. Thus, although (Ku-dependent) NHEJ skewed the DSB repair process towards inter-S–S region recombination, thereby increasing CSR rates, Rad52-dependent annealing of DNA single strands favoured DSB synapsing within the same S region, which led to intra-S region recombinations, thereby lowering CSR rates. The latter process was further emphasized by the dampening of CSR on enforced expression of Rad52.
We show here that, similar to Ku70/Ku86, Rad52 bound to the DSB free ends of CSR-targeted S regions, but did not bind to S regions not involved in CSR or Cμ region. By binding to S-region DSB free ends, Rad52 prevented Ku70/Ku86 from accessing such DSB ends, as shown by the specific ChIP assays involving anti-Ku70/Ku86 monoclonal antibody and Rad52−/− B cells, the specific competition assays using recombinant Rad52 and Ku70/Ku86 proteins, as well as the supershift EMSAs involving anti-Ku70/Ku86 monoclonal antibody and nuclear extracts from Rad52−/− B cells. The competition of Rad52 with Ku70/Ku86 for binding to S-region DSB ends was further emphasized by the decreased CSR on enforced expression of Rad52 in (normal) B cells. Such CSR decrease was more pronounced than that resulting from knocking down Ku86 in normal B cells. In these B cells, Ku86 was present in residual amounts but competed with lower amounts of Rad52 than in B cells in which Rad52 was enforced expressed. The virtual ablation of (the otherwise much increased) CSR by Ku86 knockdown in Rad52−/− B cells not only further emphasized the critical role of the Ku70/Ku86 heterodimer in the inter-S–S region DSB synaptic process but also revealed a contribution of Rad52 to inter-S–S region recombination and CSR.
In B cells induced to undergo CSR, execution of the DSB synaptic process towards inter-S–S region recombination would be further facilitated by the downregulation of Rad52 (RAD52) with concomitant upregulation of Ku70/Ku86 (KU70/KU86) together with Aicda (AICDA) and Polθ (POLθ) transcripts at early time points (24–48 h). The expression of Rad52, Ku70/Ku86, AID and Polθ proteins followed tightly the kinetic expression of their corresponding transcripts. This was particularly the case for Rad52, which is a short-lived protein58, in contrast to the relatively stable Ku70/Ku86. In switching B cells, Rad52 (RAD52)/Rad52 downregulation was profound but not complete, possibly allowing for some residual intra-S recombination. After 48–72 h, the return of Ku70/Ku86 (KU70/KU86) transcripts and Ku70/Ku86 proteins to a lower expression levels would be important to maintain a Rad52:Ku70/Ku86 ratio that prevents CSR dysregulation. Thus, the modulatory changes in Rad52 and Ku70/Ku86 are likely to be the result of CSR induction rather than a consequence of cell proliferation.
As we showed here, Rad52 deficiency did not alter B cell cycle or B-cell division and increased CSR independently of cell division. Because of its use of sister chromatids as templates to mediate faithful repair, HR is generally restricted to S and G2 phases. NHEJ and MMEJ generally operate in G0–G18, although both can function throughout the whole cell cycle8. In yeast, Rad52 is generally recruited to DSBs and readily form foci in response to DNA damage in S and G2/M, when a sister chromatid template becomes available to guide the repair process59. Rad52, however, can also accumulate at DNA damage sites immediately after insertion of DSBs outside the S and G2/M phases60,61, thereby promoting annealing of single-strand DNA and leading to (micro)homology-mediated end-joining in the absence of a sister chromatid template. Furthermore, HR has been suggested to contribute to S-region DSB repair and CSR, in addition to NHEJ/A-NHEJ48,62. Indeed, DSBs not processed by NHEJ or A-NHEJ in G0–G1 would be further resected in S-G2/M and repaired by HR using the second (intact) IgH allele as a template48. Thus, given the role of Rad52 in both A-NHEJ and HR, the contribution of Rad52 to S region DSB repair would extend throughout the whole cell cycle. Rad52-facilitated microhomology-mediated A-NHEJ may complement NHEJ in the presence of high DSB rates, such as those in hypermutating/switching GC B cells, thereby functioning as a salvage pathway for DSBs that are not repaired by NHEJ.
Collectively, our findings show that Rad52 competes with Ku for binding to S-region DSB free ends, where it facilitates a DSB synaptic process, which favours intra-S region recombination, and also mediates, in particular in the absence of a functional NHEJ pathway, inter-S–S region recombinations (Supplementary Fig. 5).
Methods
Mice
Rad52−/− mice were generated by Dr Albert Pastink (Leiden University, Leiden, The Netherlands) by replacing exon 3 of the Rad52 gene with positive selection marker neomycin, as driven by the phosphoglycerate kinase promoter, and an upstream mouse sequence functioning as a transcription terminator39. These mice were backcrossed to C57/BL6 mice for more than six generations. No full-length or truncated Rad52 protein was produced from the disrupted allele. Rad52−/− mice were viable and fertile, and showed no gross abnormalities39. p53−/− (B6.129S2-Trp53tm1Tyj/J) mice were purchased from Jackson Laboratory (Bar Harbor, Maine). p53−/−Rad52+/+ and p53−/−Rad52–/– mice were generated in our facilities by cross-breeding p53−/− with Rad52+/− mice. Aicda−/− mice (C57BL/6 background)63 were obtained from Dr Tasuku Honjo (Kyoto University, Kyoto, Japan). Polθ−/− mice (C57BL/6 background) were generated by Dr John Schimenti (Cornell University, Ithaca, NY) by placing an in‐frame stop codon into exon 1 and replacing exons 2–5 with a neomycin resistance (neo) gene37. All mice were housed in pathogen-free conditions. Both male and female mice aged 8–12 weeks were used for the experiments. The Institutional Animal Care and Use Committee of the University of Texas Health Science Center at San Antonio approved all animal protocols.
NP16-CGG and OVA immunization and antibody titration
Rad52+/+, Rad52−/−, Polθ+/+ and Polθ−/− mice (8–12 weeks of age) were injected intraperitoneally (i.p.) with 100 μg of NP16-CGG (average 16 molecules of 4-hydroxy-3-nitrophenyl acetyl coupled to 1 molecule of chicken γ-globulin; Biosearch Technologies) in 100 μl of alum (Imject Alum, Thermo Fisher Scientific). Serum was collected 10 days later for titration of circulating total and NP-binding IgM and IgG1 using enzyme-linked immunosorbent assays, as we described64,65,66. To analyse the impact of Rad52 deficiency in IgE response, Rad52+/+ and Rad52−/− mice were injected i.p. with 20 μg of OVA in 100 μl of alum and ‘boost’ injected 7 days later with 20 μg of OVA in PBS. Serum was collected before or 5 days after the ‘boost’ injection for titration of circulating total IgE.
B and T cells, cell cycle and proliferation
B cells (B220+), CD4+ and CD8+ T cells, dead B cells, GC (B220+PNAhi) B cells and plasma cells (B220loCD138+) were identified and analysed by flow cytometry64,65,67 using a FACSCalibur or LSR-II flow cytometer (BD Biosciences). Data analysis was performed using FlowJo software (Tree Star). Single-cell suspensions were prepared from spleens of Rad52+/+ and Rad52−/− mice, and stained with PE-anti-B220 monoclonal antibody (clone RA3-6B2, eBioscience, 0.05 μg ml−1), fluorescein isothiocyanate (FITC)-anti-CD3 monoclonal antibody (clone 17A2, BioLegend, 0.1 μg ml−1), FITC-anti-CD4 (clone GK1.5, BioLegend, 0.1 μg ml−1) monoclonal antibody and/or allophycocyanin (APC)-anti-CD8 monoclonal antibody (53-6.7, BD Biosciences, 0.1 μg ml−1), Alexa Fluor 647-peanut agglutinin (PNA; Invitrogen) and biotin-anti-CD138 monoclonal antibody (clone 281-2, BD Biosciences, 0.05 μg ml−1) followed by FITC-streptavidin (11-4317-87, eBioscience, 0.5 μg ml−1) or PE-streptavidin (12-4317-87, eBioscience, 0.1 μg ml−1), PE-anti-IgM monoclonal antibody (clone AF6-78, BD Biosciences, 0.1 μg ml−1), FITC-anti-IgG1 monoclonal antibody (clone A85-1, BD Biosciences, 0.1 μg ml−1), FITC-anti-IgG3 monoclonal antibody (R40-82, BD Biosciences, 0.1 μg ml−1), FITC-anti-IgA monoclonal antibody (clone C10-3, BD Biosciences, 0.1 μg ml−1) and biotin-rat-anti-mouse IgG2a/c monoclonal antibody (clone R19-15, BD Biosciences, 0.1 μg ml−1), followed by APC-streptavidin (550874, eBioscience, 0.5 μg ml−1) and 7-AAD (BD Biosciences, 0.25 μg per 1 × 106 cells). For analysis of B-cell proliferation and cell cycle in vivo, mice were immunized with NP16-CGG. Ten days after NP16-CGG immunization, the mice were injected i.p. with BrdU (1 mg in 200 μl PBS) twice within a 16 h interval and killed 4 h after the last injection. Spleen cells stained with PE-anti-B220 monoclonal antibody and APC-anti-BrdU monoclonal antibody (APC BrdU Flow Kit, BD Biosciences) to detect intracellular BrdU were analysed by flow cytometry. To analyse cell cycle, B220+ B cells or B220+PNAhi GC B cells were stained for surface B220 and PNA with PE-anti-B220 monoclonal antibody (clone RA3-6B2, eBioscience, 0.05 μg ml−1) and Alexa Fluor 647-labelled PNA (Invitrogen), fixed/permeabilized and stained with APC-anti-BrdU monoclonal antibody and 7-AAD (APC BrdU Flow Kit; BD Biosciences), and then analysed by flow cytometry. B-cell division was analysed by carboxyfluorescein succinimidyl ester (CFSE) dilution using the CellTrace CFSE Cell Proliferation Kit (Invitrogen). B cells were incubated for 5 min at 37 °C in 3 ml PBS with 2.5 μM CFSE at a density of 1 × 107 cells per ml and then washed in fetal bovine serum (FBS)–RPMI. Cells were then cultured in the presence of LPS plus IL-4 or membrane fragments of baculovirus-infected Sf21 insect cells expressing mouse CD154 (ref. 68) (mCD154, 1 or 2 U ml−1) plus IL-4 for 72 h, before being stained with PE-anti-B220 monoclonal antibody, 7-AAD and APC-anti-IgG1 monoclonal antibody (clone X56, BD Biosciences, 0.1 μg ml−1), and analysed by flow cytometry.
CSR analysis in vitro and in vivo
B cells isolated from red blood cell-depleted mouse splenocytes were purified by negative selection of cells expressing CD43, CD4, CD8, CD11b, CD49b, CD90.2, Gr-1 or Ter-119 using the EasySep Mouse B-cell Isolation kit (StemCell Technologies). B cells were resuspended in RPMI 1640 medium with 10% FBS, 50 mM β-mercaptoethanol and 1 × antibiotic–antimycotic mixture (15240-062; Invitrogen) (FBS–RPMI) at 37 °C in 48-well plates and stimulated with the following reagents: LPS (3 μg ml−1) from Escherichia coli (055:B5; Sigma-Aldrich) for CSR to IgG3; LPS (3 μg ml−1) or mCD154 (1 U ml−1) plus IL-4 (5 ng ml−1; R&D Systems) for CSR to IgG1; LPS (3 μg ml−1) plus TGF-β (2 ng ml−1; R&D Systems), IL-4 (5 ng ml−1) and IL-5 (3 ng ml−1) (R&D Systems), and anti-IgD monoclonal antibody/dextran (anti-δ monoclonal antibody/dex) (Fina Biosolutions) for CSR to IgA; LPS (3 μg ml−1) plus IFN-γ (50 ng ml−1) (R&D Systems) for CSR to IgG2a/c. After 96 h, B cells were stained with FITC-labelled rat monoclonal antibody to mouse IgG1 (clone A85-1, 0.1 μg ml−1), mouse IgG2a/c (clone R19-15, 0.1 μg ml−1), mouse IgG3 (clone R40-82, 0.1 μg ml−1), mouse IgA (clone C10-3, 0.1 μg ml−1) or PE-labelled rat monoclonal antibody to mouse B220 (clone RA3-6B2, 0.05 μg ml−1; all from BD Biosciences). Stained cells were analysed by flow cytometry. To analyse in vivo CSR, spleen or Peyer’s patch B cells were isolated from Rad52+/+ and Rad52−/− mice 10 days after immunization with NP16-CGG, and were stained with PE-labelled anti-B220 monoclonal antibody or Alexa Fluor 647-labelled PNA, as well as FITC-labelled anti-IgG1 (clone A85-1, 0.1 μg ml−1) or FITC-labelled anti-IgA (clone C10-3, 0.1 μg ml−1) monoclonal antibodies (BD Biosciences), and then analysed by flow cytometry.
Human B cells and stimulation
Human naive B cells (>95% pure) were purified by negative selection, using the EasySep Human Naive B Cell Enrichment Kit (19254; StemCell Technologies), from healthy donor peripheral blood mononuclear cells, following the manufacturer’s instructions. Naive B cells were then cultured in FBS–RPMI and stimulated with mCD154 (1, 2 or 4 U ml−1), recombinant human IL-4 (20 ng ml−1; R&D Systems) and recombinant human IL-21 (50 ng ml−1; R&D Systems) for 24, 48, 72 and 96 h. RNA was extracted from stimulated and unstimulated B cells and used for qRT–PCR analysis.
qRT–PCR analysis of transcripts
Total RNA was extracted from 2–5 × 106 B cells using the RNeasy Mini Kit (Qiagen). Residual DNA was removed using genomic DNA eliminator columns (Qiagen). cDNA was synthesized from 1 to 2 μg total RNA with the SuperScript III First-Strand Synthesis System (Invitrogen) using oligo-dT primer. The expression of mouse germline Iγ1-Cγ1 and Iγ3-Cγ3, circle Iγ1-Cμ and Iγ3-Cμ, post-recombination Iμ-Cγ1 and Iμ-Cγ3, Aicda, Rad52, Ku70, Ku86, Polθ, Cd79b and Gapdh transcripts, as well as human AICDA, RAD52, POLθ, KU70, KU86 and CD79b transcripts, was measured by real-time qRT–PCR with appropriate primers3,64,65 (Supplementary Table 2). A Bio-Rad MyiQ Real-Time PCR Detection System (Bio-Rad Laboratories) was used to measure SYBR Green (IQ SYBR Green Supermix, Bio-Rad Laboratories) incorporation with the following protocol: 95 °C for 15 s, 40 cycles of 94 °C for 10 s, 60 °C for 30 s, 72 °C for 30 s. Data acquisition was performed during this 72 °C extension step. Melting curve analysis was performed from 72 to 95 °C. The change in cycling threshold (ΔΔCt) method was used to analyse levels of transcripts and data were normalized to the level of Cd79b/CD79b, which encodes the BCR Igβ chain constitutively expressed in B cells, or Gapdh.
Immunoblotting
B cells were lysed in Laemmli buffer. Cell extracts containing equal amounts of protein (20 μg) were fractionated through SDS–PAGE (10%). The fractionated proteins were transferred onto polyvinylidene difluoride membranes (Bio-Rad) overnight (30 V/90 mA) at 4 °C. After blocking and overnight incubation at 4 °C with anti-AID antibody (H-80, Santa Cruz, 0.2 μg ml−1), anti-Ku70 antibody (C-19, Santa Cruz, 0.2 μg ml−1), anti-Ku86 monoclonal antibody (clone 111, Thermo Fisher Scientific, 0.2 μg ml−1), anti-Rad52 antibody (H-300, Santa Cruz Biotechnology, 0.4 μg ml−1), anti-Polθ antibody (AAS04492C, Antibody Verify, 1 μg ml−1) or anti-β-Actin monoclonal antibody (2F1-1, BioLegend, 0.1 μg ml−1), the membranes were incubated with horseradish peroxidase (HRP)-conjugated secondary antibodies. After washing with TBS–Tween 20 (0.05%), bound HRP-conjugated antibodies were detected using Western Lightning Plus-ECL reagents (PerkinElmer Life and Analytical Sciences).
Semi-quantitative digestion–circularization PCR
B-cell genomic DNA was digested with HindIII before being ligated with T4 DNA ligase to generate circularized DNA65. Sμ–Sγ1 and Sμ–Sγ3 HindIII fragments were amplified by nested PCR using Phusion High-Fidelity DNA polymerase (New England Biolabs) with reported primers69. The completion of HindIII digestions and ligations was verified by PCR with a primer set specific for Gapdh. The first round PCR was carried out for 25 cycles at 94 °C for 30 s, 58 °C for 30 s, 72 °C for 35 s. Product (1 μl) of the first-round PCR was used as template for the second round of PCR for 30 cycles at 94 °C for 30 s, 60 °C for 30 s, 72 °C for 30 s.
DNA S–S junctional sequences
Genomic DNA was prepared from B cells cultured for 96 h with LPS and rmIL-4, or B220+PNAhi B cells from Peyer’s patches. Sμ–Sγ1 or Sμ–Sα DNA was amplified by two sequential rounds of specific PCR using Phusion high-fidelity DNA polymerase (New England Biolabs) and nested oligonucleotide primers41,67 (Supplementary Table 2). The first and second rounds of PCR were performed at 94 °C for 45 s, 55 °C for 45 s, 72 °C for 4 min (30 cycles). PCR products were purified using a QIAquick PCR purification kit (Qiagen) and cloned into the pCR-Blunt II-TOPO vector (Invitrogen) for sequencing. Sequence alignment was performed by comparing the sequences of PCR products with Sμ and Sγ1 or Sα genomic sequences using National Center for Biotechnology Information BLAST (www.ncbi.nih.gov/BLAST).
ChIP assays
ChIP assays were performed as described64,65,70. B cells (1 × 107) were treated with formaldehyde (1% v/v) for 10 min at 25 °C to cross-link chromatin. After quenching with 100 mM of glycine (pH 8.0) and washing with cold PBS containing protease inhibitors (Roche), B cells were suspended in lysis buffer (20 mM Tris-HCl, 200 mM NaCl, 2 mM EDTA, 0.1% w/v SDS and protease inhibitors pH 8.0). Chromatin was sonicated to yield DNA fragments (∼200 to 1,000 bp in length), pre-cleared with Pierce Protein A beads (Thermo Fisher Scientific) and then incubated with rabbit anti-Rad52 antibody (H-300; Santa Cruz Biotechnology, 5 μg ml−1), anti-Ku70/86 monoclonal antibody (MA1-21818, Thermo Fisher Scientific, 5 μg ml−1), or control rabbit or mouse IgG with irrelevant specificities at 4 °C overnight. Immune complexes were precipitated by Protein A agarose beads, washed and eluted (50 mM Tris-HCl, 0.5% SDS, 200 mM NaCl, 100 μg ml−1 proteinase K pH 8.0), followed by incubation at 65 °C for 4 h. DNA was purified using a QIAquick PCR purification kit (Qiagen) and used as a template for analysis of Sμ, Sγ1 and Sγ3 by quantitative PCR using specific primers70 (Supplementary Table 2).
In situ DNA end-labelling with biotin
DNA DSB ends were labelled with biotin41. Rad52+/+ and Rad52−/− B cells were stimulated with LPS or LPS plus IL-4 for 60 h. Live B cells were separated through a Ficoll gradient, followed by fixation, permeabilization and in situ DNA end-labelling with bio-dUTP using TdT (Fig. 6d). Chromatin was pulled down with rabbit anti-Rad52 antibody (H-300, Santa Cruz Biotechnology, 5 μg ml−1), anti-Ku70/Ku86 monoclonal antibody (MA1-21818, Thermo Fisher Scientific, 5 μg ml−1) or control rabbit or mouse IgG with irrelevant specificity. Genomic DNA was prepared using a QIAquick PCR purification kit (Qiagen). Biotin-labelled DNA fragments were captured by streptavidin magnetic beads (Promega) before being analysed by quantitative PCR.
Electrophoretic mobility shift assays
An ‘asymmetrical’ biotin-labelled double-strand Sμ DNA probe was generated by PCR using a synthetic oligonucleotide Sμ1F, which consisted of part of the mouse Sμ region sequence 5′-CATTAATCTAGGTTGAATAGAGCTAAACTCTACTGCCTACACTGGACTGTTCTGAGCTGAGATGAGCTGGGGTG-3′ as template, and amplified with forward Sμ3F-Kpn (5′-AATTCGGTACCAACTTCATTAATCTAGGTTGAATAG-3′) primer, which included a KpnI site, and a Sμ3R (5′-CACCCCAGCTCATCTCAGC-3′) primer. Bio-dUTP was added to the PCR reaction. The PCR product was treated with T4 DNA polymerase in the presence of dNTP and digested by KpnI to generate a 4-nt 3′-single-strand overhang at one end of the oligonucleotide, the other end being blunt. The biotin-labelled probe was purified with the QIAquick Gel Extraction Kit (Qiagen) and used for EMSA reactions (5 fmol per reaction). EMSA assays were performed using the LightShift Chemiluminescent EMSA Kit (Thermo Fisher Scientific) according to the manufacturer’s instructions. For the Rad52–Ku70/Ku86 competition experiments, purified recombinant human RAD52 (Axxora) or KU70/KU86 (Cell Sciences) proteins were used alone (0.32 pmol), or mixed at a 1:1 ratio (0.32 pmol RAD52 and 0.32 pmol KU70/KU86), 2:1 (0.64 pmol RAD52 and 0.32 pmol KU70/KU86) or 4:1 (1.28 pmol RAD52 and 0.32 pmol KU70/KU86). In supershift experiments, nuclear extracts (10 μg protein) from Rad52+/+ and Rad52−/− B cells stimulated with LPS plus IL-4 for 60 h were pre-incubated for 30 min at room temperature with anti-Ku70/Ku86 monoclonal antibody (MA1-21818, Thermo Fisher Scientific, 1 μg per reaction) or irrelevant mouse IgG before being incubated with the biotin-labelled Sμ probe. Electrophoresis of the DNA–protein complexes was carried out on a 5% non-denaturing polyacrylamide gel with 0.5 × TBE. The samples were then transferred to Hybond-N+ membranes with a semi-dry transfer followed by ultraviolet cross-linking. Detection was performed using the Chemiluminescent Nucleic Acid Detection Module (Thermo Fisher Scientific) according to the manufacturer’s instructions.
c-Myc/IgH translocations
Genomic DNA was isolated from p53−/−Rad52–/– and p53−/−Rad52+/+ B cells stimulated with LPS plus IL-4 for 96 h. Nested PCRs for translocations were performed on genomic DNA from 1 × 106 cells with GoTaq Long PCR Master Mix (Promega) using IgH forward and c-Myc reverse primers (Supplementary Table 2). PCR conditions were as follows: 95 °C for 2 min followed by 25 cycles (95 °C, 10 s; 62 °C, 45 s; and 65 °C, 6 min) for both the first and second rounds. Amplified DNA were fractionated through 1.0% agarose, blotted onto Hybond-N+ membranes and hybridized to biotin labelled c-Myc-specific oligonucleotide probe 5′-GACGCCACTGCACCAGAGACCCTGCAGCGATTCAG-3′ or IgH-specific oligonucleotide probe 5′-CCTGGTATACAGGACGAAACTGCAGCAG-3′ and detected using Streptavidin-HRP. To analyse the c-Myc–IgH junctional sequences, PCR products were treated with T4 DNA polymerase in the presence of dNTP, cloned into the pCR-Blunt II-TOPO vector (Invitrogen) and sequenced. Sequence alignment was performed by comparing the sequences of PCR products with germline c-Myc and IgH genomic sequences using National Center for Biotechnology Information BLAST.
Analysis of intra-Sμ region recombination
To analyse intra-Sμ region DNA recombination, Sμ region DNA was amplified by nested PCR using mouse Sμ-specific primers (Supplementary Table 2) and GoTaq Long PCR Master Mix (Promega) from genomic DNA isolated from Rad52+/+ and Rad52−/− B cells stimulated with LPS plus IL-4 for 96 h. Amplified DNA was treated with T4 DNA polymerase in the presence of dNTP and cloned into the pCR-Blunt II-TOPO vector (Invitrogen). Sμ region DNA of individual clones were re-amplified using the Sμ-specific primers (as above) and GoTaq Long PCR Master Mix. Amplified DNA was fractionated through 0.8% agarose gel. The intra-Sμ recombined DNAs, that is, Sμ region DNA with internal deletions, were detected by comparing the length of the amplified DNA with that of the respective germline Sμ region and positively identified by sequencing.
Rad52 retroviral construct and enforced expression
Rad52 cDNA was amplified from mouse B cells using the appropriate primers (Supplementary Table 2) and cloned into the pMIG retroviral expression vector (green fluorescent protein (GFP) translation initiated by the internal ribosome entry sites (IRES) in transduced B cells). To generate the retrovirus, pMIG vector encoding GFP or pMIG-Rad52 construct encoding GFP and Rad52, together with the pCL-Eco retrovirus-packaging vector (Imgenex), were used to transfect HEK293T cells by a Ca++ phosphate-mediated transfection procedure (ProFection Mammalian Transfection System, Promega). Viral supernatants were collected and used to transduce spleen B cells from C57BL/6 mice, as we reported64,65, after a 12 h LPS activation. Transduced B cells were then stimulated with LPS plus IL-4 for 96 h before analysis of GFP+ and IgG1+ B cells by flow cytometry64,65—dead (7-AAD+) cells were excluded from analysis. Expression of Rad52, Ku86 and β-Actin proteins in B cells transduced with empty or Rad52-expression pMIG retroviral construct were analysed by immunoblotting66.
Ku86 knockdown using lentiviral shRNA construct
The Ku86-specific shRNA lentiviral construct pGFP-C-Ku86-shRNALenti (TL502435) and non-effective 29-mer scrambled shRNA lentiviral construct pGFP-C-scr-shLenti (TR30021) were obtained from Origene Technologies. To generate the lentivirus, pGFP-C-shLenti vector and packaging vectors were used to cotransfect HEK293T cells according to the manufacturer’s instructions (Lenti-vpak Lentiviral Packaging Kit, Origene). Viral supernatants were collected and used to transduce Rad52+/+ and Rad52−/− B cells. Transduced B cells were then stimulated with LPS plus IL-4 for 96 h before analysing GFP+ and IgG1+ B cells by flow cytometry—dead (7-AAD+) cells were excluded from analysis. Expressions of Rad52, Ku86 and β-Actin proteins in the transduced B cells were analysed by immunoblotting.
Statistical analysis
Statistical analysis was performed using Excel (Microsoft), to determine P-values by paired and unpaired Student’s t-tests; and P-values <0.05 were considered significant. Differences in the number and length of microhomologies in Sμ–Sγ1 and Sμ–Sα junctions between stimulated Rad52+/+ and Rad52−/−, or Polθ+/+ and Polθ−/− B cells were analysed using the paired Student’s t-test.
Data availability
The data that support the findings of this study are available within the article and its Supplementary Information file or from the corresponding authors upon reasonable request.
Additional information
How to cite this article: Zan, H. et al. Rad52 competes with Ku70/Ku86 for binding to S-region DSB ends to modulate antibody class-switch DNA recombination. Nat. Commun. 8, 14244 doi: 10.1038/ncomms14244 (2017).
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Casali, P., Pal, Z., Xu, Z. & Zan, H. DNA repair in antibody somatic hypermutation. Trends Immunol. 27, 313–321 (2006).
Xu, Z., Zan, H., Pone, E. J., Mai, T. & Casali, P. Immunoglobulin class switching: induction, targeting and beyond. Nat. Rev. Immunol. 12, 517–531 (2012).
Li, G., Zan, H., Xu, Z. & Casali, P. Epigenetics of the antibody response. Trends Immunol. 34, 460–470 (2013).
Casali, P. in: Lewin’s Genes XII (eds Krebbs, J.et al.) 397–439 (Jones & Bartlett, 2017).
Zan, H. & Casali, P. Regulation of Aicda expression and AID activity. Autoimmunity 46, 83–101 (2013).
Liu, M. & Schatz, D. G. Balancing AID and DNA repair during somatic hypermutation. Trends Immunol. 30, 173–181 (2009).
Unniraman, S. & Schatz, D. G. AID and IgH switch region-Myc chromosomal translocations. DNA Repair 5, 1259–1264 (2006).
Symington, L. S. & Gautier, J. Double-strand break end resection and repair pathway choice. Annu. Rev. Genet. 45, 247–271 (2011).
Truong, L. N. et al. Microhomology-mediated end joining and homologous recombination share the initial end resection step to repair DNA double-strand breaks in mammalian cells. Proc. Natl Acad. Sci. USA 110, 7720–7725 (2013).
Lieber, M. R. The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu. Rev. Biochem. 79, 181–211 (2010).
Pannunzio, N. R., Li, S., Watanabe, G. & Lieber, M. R. Non-homologous end joining often uses microhomology: implications for alternative end joining. DNA Repair 17, 74–80 (2014).
Alt, F. W., Zhang, Y., Meng, F. L., Guo, C. & Schwer, B. Mechanisms of programmed DNA lesions and genomic instability in the immune system. Cell 152, 417–429 (2013).
Deriano, L. & Roth, D. B. Modernizing the nonhomologous end-joining repertoire: alternative and classical NHEJ share the stage. Annu. Rev. Genet. 47, 433–455 (2013).
Manis, J. P. et al. Ku70 is required for late B cell development and immunoglobulin heavy chain class switching. J. Exp. Med. 187, 2081–2089 (1998).
Casellas, R. et al. Ku80 is required for immunoglobulin isotype switching. EMBO J. 17, 2404–2411 (1998).
Yan, C. T. et al. IgH class switching and translocations use a robust non-classical end-joining pathway. Nature 449, 478–482 (2007).
Boboila, C. et al. Alternative end-joining catalyzes robust IgH locus deletions and translocations in the combined absence of ligase 4 and Ku70. Proc. Natl Acad. Sci. USA 107, 3034–3039 (2010).
Boboila, C. et al. Alternative end-joining catalyzes class switch recombination in the absence of both Ku70 and DNA ligase 4. J. Exp. Med. 207, 417–427 (2010).
Han, L. & Yu, K. Altered kinetics of nonhomologous end joining and class switch recombination in ligase IV-deficient B cells. J. Exp. Med. 205, 2745–2753 (2008).
McVey, M. & Lee, S. E. MMEJ repair of double-strand breaks (director’s cut): deleted sequences and alternative endings. Trends Genet. 24, 529–538 (2008).
Sfeir, A. & Symington, L. S. Microhomology-mediated end joining: a back-up survival mechanism or dedicated pathway? Trends Biochem. Sci. 40, 701–714 (2015).
Villarreal, D. D. et al. Microhomology directs diverse DNA break repair pathways and chromosomal translocations. PLoS Genet. 8, e1003026 (2012).
Decottignies, A. Microhomology-mediated end joining in fission yeast is repressed by pku70 and relies on genes involved in homologous recombination. Genetics 176, 1403–1415 (2007).
Bennardo, N., Cheng, A., Huang, N. & Stark, J. M. Alternative-NHEJ is a mechanistically distinct pathway of mammalian chromosome break repair. PLoS Genet. 4, e1000110 (2008).
Simsek, D. & Jasin, M. Alternative end-joining is suppressed by the canonical NHEJ component Xrcc4-ligase IV during chromosomal translocation formation. Nat. Struct. Mol. Biol. 17, 410–416 (2010).
Zhang, Y. & Jasin, M. An essential role for CtIP in chromosomal translocation formation through an alternative end-joining pathway. Nat. Struct. Mol. Biol. 18, 80–84 (2011).
Decottignies, A. Alternative end-joining mechanisms: a historical perspective. Front. Genet. 4, 48 (2013).
Robert, I., Dantzer, F. & Reina-San-Martin, B. Parp1 facilitates alternative NHEJ, whereas Parp2 suppresses IgH/c-Myc translocations during immunoglobulin class switch recombination. J. Exp. Med. 206, 1047–1056 (2009).
Lee-Theilen, M., Matthews, A. J., Kelly, D., Zheng, S. & Chaudhuri, J. CtIP promotes microhomology-mediated alternative end joining during class-switch recombination. Nat. Struct. Mol. Biol. 18, 75–79 (2011).
West, S. C. Molecular views of recombination proteins and their control. Nat. Rev. Mol. Cell Biol. 4, 435–445 (2003).
Grimme, J. M. et al. Human Rad52 binds and wraps single-stranded DNA and mediates annealing via two hRad52-ssDNA complexes. Nucleic Acids Res. 38, 2917–2930 (2010).
Ceccaldi, R., Rondinelli, B. & D’Andrea, A. D. Repair pathway choices and consequences at the double-strand break. Trends Cell Biol. 26, 52–64 (2016).
Zan, H., Wu, X., Komori, A., Holloman, W. K. & Casali, P. AID-dependent generation of resected double-strand DNA breaks and recruitment of Rad52/Rad51 in somatic hypermutation. Immunity 18, 727–738 (2003).
Ceccaldi, R. et al. Homologous-recombination-deficient tumours are dependent on Poltheta-mediated repair. Nature 518, 258–262 (2015).
Mateos-Gomez, P. A. et al. Mammalian polymerase theta promotes alternative NHEJ and suppresses recombination. Nature 518, 254–257 (2015).
Kent, T., Chandramouly, G., McDevitt, S. M., Ozdemir, A. Y. & Pomerantz, R. T. Mechanism of microhomology-mediated end-joining promoted by human DNA polymerase theta. Nat. Struct. Mol. Biol. 22, 230–237 (2015).
Zan, H. et al. The translesion DNA polymerase theta plays a dominant role in immunoglobulin gene somatic hypermutation. EMBO J. 24, 3757–3769 (2005).
Ramiro, A. R. et al. Role of genomic instability and p53 in AID-induced c-myc-Igh translocations. Nature 440, 105–109 (2006).
Rijkers, T. et al. Targeted inactivation of mouse RAD52 reduces homologous recombination but not resistance to ionizing radiation. Mol. Cell Biol. 18, 6423–6429 (1998).
Yousefzadeh, M. J. et al. Mechanism of suppression of chromosomal instability by DNA polymerase POLQ. PLoS Genet. 10, e1004654 (2014).
Zan, H. et al. Endonuclease G plays a role in immunoglobulin class switch DNA recombination by introducing double-strand breaks in switch regions. Mol. Immunol. 48, 610–622 (2011).
Wang, J. H. et al. Mechanisms promoting translocations in editing and switching peripheral B cells. Nature 460, 231–236 (2009).
Van Dyck, E., Stasiak, A. Z., Stasiak, A. & West, S. C. Binding of double-strand breaks in DNA by human Rad52 protein. Nature 398, 728–731 (1999).
Ristic, D., Modesti, M., Kanaar, R. & Wyman, C. Rad52 and Ku bind to different DNA structures produced early in double-strand break repair. Nucleic Acids Res. 31, 5229–5237 (2003).
Wu, Y., Siino, J. S., Sugiyama, T. & Kowalczykowski, S. C. The DNA binding preference of RAD52 and RAD59 proteins: implications for RAD52 and RAD59 protein function in homologous recombination. J. Biol. Chem. 281, 40001–40009 (2006).
Downs, J. A. & Jackson, S. P. A means to a DNA end: the many roles of Ku. Nat. Rev. Mol. Cell Biol. 5, 367–378 (2004).
Sugiyama, T. & Kantake, N. Dynamic regulatory interactions of rad51, rad52, and replication protein-a in recombination intermediates. J. Mol. Biol. 390, 45–55 (2009).
Yamane, A. et al. RPA accumulation during class switch recombination represents 5′-3′ DNA-end resection during the S-G2/M phase of the cell cycle. Cell Rep. 3, 138–147 (2013).
Rothenberg, E., Grimme, J. M., Spies, M. & Ha, T. Human Rad52-mediated homology search and annealing occurs by continuous interactions between overlapping nucleoprotein complexes. Proc. Natl Acad. Sci. USA 105, 20274–20279 (2008).
Honda, M., Okuno, Y., Yoo, J., Ha, T. & Spies, M. Tyrosine phosphorylation enhances RAD52-mediated annealing by modulating its DNA binding. EMBO J. 30, 3368–3382 (2011).
Deng, S. K., Gibb, B., de Almeida, M. J., Greene, E. C. & Symington, L. S. RPA antagonizes microhomology-mediated repair of DNA double-strand breaks. Nat. Struct. Mol. Biol. 21, 405–412 (2014).
Meyer, D., Fu, B. X. & Heyer, W. D. DNA polymerases delta and lambda cooperate in repairing double-strand breaks by microhomology-mediated end-joining in Saccharomyces cerevisiae. Proc. Natl Acad. Sci. USA 112, E6907–E6916 (2015).
Daley, J. M. & Wilson, T. E. Rejoining of DNA double-strand breaks as a function of overhang length. Mol. Cell Biol. 25, 896–906 (2005).
Feng, Q. et al. Rad52 and Rad59 exhibit both overlapping and distinct functions. DNA Repair 6, 27–37 (2007).
Crespan, E., Czabany, T., Maga, G., Hubscher, U. & Microhomology-mediated, D. N. A. strand annealing and elongation by human DNA polymerases lambda and beta on normal and repetitive DNA sequences. Nucleic Acids Res. 40, 5577–5590 (2012).
Masani, S., Han, L., Meek, K. & Yu, K. Redundant function of DNA ligase 1 and 3 in alternative end-joining during immunoglobulin class switch recombination. Proc. Natl Acad. Sci. USA 113, 1261–1266 (2016).
Bothmer, A. et al. 53BP1 regulates DNA resection and the choice between classical and alternative end joining during class switch recombination. J. Exp. Med. 207, 855–865 (2010).
Sacher, M., Pfander, B., Hoege, C. & Jentsch, S. Control of Rad52 recombination activity by double-strand break-induced SUMO modification. Nat. Cell Biol. 8, 1284–1290 (2006).
Barlow, J. H. & Rothstein, R. Rad52 recruitment is DNA replication independent and regulated by Cdc28 and the Mec1 kinase. EMBO J. 28, 1121–1130 (2009).
Alabert, C., Bianco, J. N. & Pasero, P. Differential regulation of homologous recombination at DNA breaks and replication forks by the Mrc1 branch of the S-phase checkpoint. EMBO J. 28, 1131–1141 (2009).
Koike, M., Yutoku, Y. & Koike, A. The C-terminal region of Rad52 is essential for Rad52 nuclear and nucleolar localization, and accumulation at DNA damage sites immediately after irradiation. Biochem. Biophys. Res. Commun. 435, 260–266 (2013).
Buerstedde, J. M., Lowndes, N. & Schatz, D. G. Induction of homologous recombination between sequence repeats by the activation induced cytidine deaminase (AID) protein. eLife 3, e03110 (2014).
Muramatsu, M. et al. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell 102, 553–563 (2000).
Park, S.-R. et al. HoxC4 binds to the promoter of the cytidine deaminase AID gene to induce AID expression, class-switch DNA recombination and somatic hypermutation. Nat. Immunol. 10, 540–550 (2009).
Zan, H. et al. Rev1 recruits Ung to switch regions and enhances dU glycosylation for immunoglobulin class switch DNA recombination. Cell Rep. 2, 1220–1232 (2012).
White, C. A. et al. Histone deacetylase inhibitors upregulate B cell microRNAs that silence AID and Blimp-1 expression for epigenetic modulation of antibody and autoantibody responses. J. Immunol. 193, 5933–5950 (2014).
Zan, H. & Casali, P. AID- and Ung-dependent generation of staggered double-strand DNA breaks in immunoglobulin class switch DNA recombination: a post-cleavage role for AID. Mol. Immunol. 46, 45–61 (2008).
Pone, E. J. et al. BCR-signalling synergizes with TLR-signalling for induction of AID and immunoglobulin class-switching through the non-canonical NF-kappaB pathway. Nat. Commun. 3, 767 (2012).
Wang, L., Wuerffel, R., Feldman, S., Khamlichi, A. A. & Kenter, A. L. S region sequence, RNA polymerase II, and histone modifications create chromatin accessibility during class switch recombination. J. Exp. Med. 206, 1817–1830 (2009).
Li, G. et al. Combinatorial H3K9acS10ph histone modification in IgH locus S regions targets 14-3-3 adaptors and AID to specify antibody class-switch DNA recombination. Cell Rep. 5, 702–714 (2013).
Acknowledgements
We thank Dr Albert Pastink (Leiden University, Leiden, The Netherlands) for the Rad52−/− mice and Dr Tasuku Honjo (Kyoto University, Kyoto, Japan) for the Aicda−/− mice. We are grateful to Meghan Guzman for her contribution to EMSA experiments, Helia N. Sanchez for help with some in vivo experiments and Dr Jingsong Zhang for his help with some in vitro experiments. We thank Ken L. Hayama and Crystal LaFleur for technical help. This work was supported by NIH grants AI 105813, AI 079705 and the Alliance for Lupus Research Target Identification in Lupus Grant ALR 295955 (P.C.), and the Arthritis National Research Foundation research grant (H.Z.).
Author information
Authors and Affiliations
Contributions
H.Z. designed and performed experiments, analysed data, supervised the work and wrote the manuscript. C.T., Z.Q., T.S., J.R.T. and J.A.G. performed experiments. P.C. planned the study, designed the experiments, analysed the data, supervised the work and wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary Figures and Supplementary Tables (PDF 13591 kb)
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zan, H., Tat, C., Qiu, Z. et al. Rad52 competes with Ku70/Ku86 for binding to S-region DSB ends to modulate antibody class-switch DNA recombination. Nat Commun 8, 14244 (2017). https://doi.org/10.1038/ncomms14244
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/ncomms14244
This article is cited by
-
SHLD1 is dispensable for 53BP1-dependent V(D)J recombination but critical for productive class switch recombination
Nature Communications (2022)
-
Rad52 mediates class-switch DNA recombination to IgD
Nature Communications (2022)
-
Rad52’s DNA annealing activity drives template switching associated with restarted DNA replication
Nature Communications (2022)
-
B cell-intrinsic epigenetic modulation of antibody responses by dietary fiber-derived short-chain fatty acids
Nature Communications (2020)
-
Repair of G1 induced DNA double-strand breaks in S-G2/M by alternative NHEJ
Nature Communications (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
