
Abstract
Programmed cell death is essential for the development and maintenance of the immune system and its responses to exogenous and endogenous stimuli. Studies have demonstrated that in addition to caspase-dependent apoptosis, necrosis dependent on the kinases RIP1 and RIP3 (also called necroptosis) is a major programmed cell-death pathway in development and immunity. These two programmed cell-death pathways may suppress each other, and necroptosis also serves as an alternative when caspase-dependent apoptosis is inhibited or absent. Here we summarize recent advancements that have identified the molecular mechanisms that underlie necroptosis and explore the mechanisms that regulate the interplay between apoptosis and necroptosis.
Similar content being viewed by others

Main
Normal development and function of the immune system require cell death mediated by distinct and highly regulated intracellular signaling pathways1. Programmed cell-death pathways are essential for the elimination of reactive T cells and B cells after massive clonal expansion in response to infection2. They also have a key role in the elimination of autoreactive cells during the education of immune cells in the thymus and bone marrow to establish tolerance to self tissues1. Innate immune responses, such as natural killer (NK) cell–mediated killing of virus-infected or transformed cells, also require programmed cell death3. Defects in programmed cell death are linked to immunodeficiencies, autoimmune diseases and malignant growth1. Apoptosis, characterized by the activation of caspases, DNA fragmentation and membrane blebbing, was once considered the sole form of programmed cell death. In contrast, necrotic cell death, which features swelling of cellular organelles and cytoplasm with subsequent rupture of the plasma membrane and cell lysis, was originally considered a nonspecific mode of cell death. However, growing evidence has unambiguously demonstrated that certain types of necrosis are tightly controlled by orchestrated intrinsic cellular programs and are now called necroptosis4,5,6. Because of the link between necrosis and inflammation and the role of necrosis in pathogenesis of many human diseases, understanding the molecular mechanisms that underlie programmed necrosis, its relationship with apoptotic pathways and the pathophysiological importance of necroptosis remains an important goal of immunological research.
Programmed necrosis (necroptosis)
It has been known for more than two decades that the proinflammatory cytokine tumor necrosis factor (TNF) can induce apoptosis in some cell lines and necrosis in others7. Physiological necrosis in vivo was also observed a decade ago under various physiological settings, including mammalian development and tissue homoeostasis8,9. The understanding of a specific cellular program for necrosis (necroptosis) was initiated by the discovery that the serine-threonine kinase RIP1 (RIPK1) is required for the necrosis of Jurkat human T lymphocyte cells triggered by the death receptor Fas (CD95)10 and the necrosis of mouse embryonic fibroblasts (MEFs) induced by TNF11. RIP1 inhibitors were later developed and proved to be very useful in studies of RIP1-dependent necroptosis6,12. The RIP1 family member RIP3 has been identified as a key mediator of caspase-independent cell death13,14,15. Thus, necroptosis is an intrinsic pathway that may be of importance equal to that of apoptosis.
Mechanisms of necroptosis
TNF is the most extensively studied extracellular signal that leads to necroptosis. Many other signals that trigger necroptosis have also been identified, such as ligation of Fas and the engagement of sensors of innate immunity (pathogen-recognition receptors), including Toll-like receptors (TLRs) and sensors of viral DNA and RNA16,17,18,19. Lipopolysaccharide, a component of the cell walls of Gram-negative bacteria, interacts with TLR4 and induces macrophage necroptosis when caspase-8 activity is inhibited15,20. The Gram-negative bacterium Shigella flexneri triggers necroptosis in neutrophils21 and human monocyte–derived macrophages22. Viral double-stranded RNA, most probably through TLR3, induces necroptotic death of Jurkat T cells and L929 mouse fibroblasts19. Signaling via the T cell antigen receptor (TCR) induces necroptosis in caspase-8-defective T cells23. Many viruses encode caspase inhibitors that function to suppress infection-induced apoptosis of host cells, which leads to more spreading of the virus and less clearance of virus24,25. However, infection by such viruses, which include vaccinia virus, herpes virus and cowpox virus, often brings about necrosis-like cell death13,26. As caspase inhibitor–deficient viral strains can induce apoptotic cell death26, necroptosis seems to be a backup mechanism that acts in response to viral infection when caspases are inhibited. More importantly, murine cytomegalovirus (MCMV) encodes an inhibitor of necroptosis17, which suggests that some viruses have already evolved mechanisms to antagonize necroptosis to alleviate host control over them. Beyond immunology-related stimuli, some genotoxic stressors and other adverse environmental conditions can also induce necroptosis. For example, nitric oxide–mediated nitration of proteins elicits RIP1- and RIP3-mediated necroptosis by targeting the respiratory complex I subunit NDUFB8 (ref. 27), and etoposide-induced cell death involves RIP1 (ref. 28).
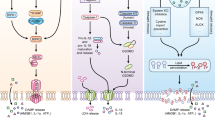
At present, the most informative studies of necroptosis derive from systems that use TNF as a trigger (Fig. 1). After a ligand binds, the cytosolic portion of TNF receptor 1 (TNFR1) recruits multiple proteins that collectively constitute the signal complex called 'complex I'29 (Fig. 1). Complex I provides a platform for the recruitment of downstream kinases and effector proteins to activate the transcription factor NF-κB and mitogen-activated protein kinases30,31. NF-κB is believed to initiate a survival pathway because it mediates the expression of some genes encoding cytoprotective molecules32. Although RIP1 is an important component of complex I that stimulates NF-κB33, TNF can induce NF-κB activation in RIP1-deficient (Ripk1−/−) MEFs and other types of cells34, which suggests that RIP1 is not indispensable for complex I–mediated activation of NF-κB. The internalization of ligand-bound TNFR1 is accompanied by the formation of complex II (also called 'DISC' (cytosolic death-inducing signaling complex); Fig. 1). This complex contains deubiquitinated RIP1, caspase-8 and the adaptors TRADD and FADD29. RIP1 is involved in the activation of caspase-8 and apoptosis30,35; however, caspase-8 can also be activated by deletion of cFLIP, the cellular form of the caspase-8 inhibitor FLIP35; thus, RIP1 may not be indispensable for TNF-induced apoptosis. This is consistent with data showing that Ripk1−/− MEFs still undergo caspase-dependent apoptosis after TNF treatment11.

After ligand binds to the receptor, the intracellular tails of TNFR1 recruit multiple proteins to form the membrane-proximal supramolecular structure complex I. Lys63-linked polyubiquitination (Lys63-Ub) of RIP1 in complex I mediated by cIAP ligases is crucial for the activation of NF-κB and mitogen-activated protein kinases (MAPKs). Deubiquitination of RIP1 by cylindromatosis (CYLD) or inhibition of cIAP proteins promote the conversion of complex I to complex II and inhibits NF-κB activation. Complex II contains RIP1, FADD, caspase-8 and TRADD. Caspase-8 becomes activated in complex II and initiates apoptosis, whereas cFLIPL can prevent activation of caspase-8. In cells with high expression of RIP3, RIP3 enters complex II via interaction with RIP1 after stimulation. The RIP3-containing complex is called complex IIb or the necrosome. In the presence of cFLIPL, caspase-8 is unable to initiate apoptosis but can cleave RIP1 and RIP3 and thus inhibits necroptosis. Depletion of FADD or caspase-8, inhibition of caspase-8 or induction of RIP3 can free RIP1-RIP3 from inhibition and initiate necroptosis of TNF-treated cells.
The idea of necrotic signaling complex, a multiprotein complex that resembles DISC (complex II) in apoptosis, was proposed after RIP3 was found in complex with RIP1 in TNF-induced necroptotic cells. As FADD and caspase-8 are also present in the RIP1-RIP3 complex13,14, the necrotic signaling complex bears a strong resemblance to complex II in composition; hence, some investigators have called it 'complex IIb'5, whereas others have called it the 'necrosome'36 (Fig. 1). Caspase-8 in the necrosome can inactivate RIP1 and RIP3 by direct cleavage of these two kinases; thus, inhibition or deletion of caspase-8 promotes necroptosis23,37,38,39,40,41 (Fig. 1).
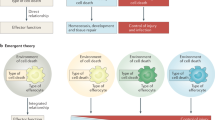
Analysis of cell death induced by the synthetic RNA duplex poly(I:C) or etoposide has identified a 2-megadalton protein complex called the 'ripoptosome'16,28 that contains RIP1, FADD, caspase-8, caspase-10 and cFLIP proteins. Like complex II in death receptor–mediated cell death, the ripoptosome seems to govern the death signaling in cells infected with RNA viruses or exposed to genotoxic stress. In a manner similar to the way that deubiquitinating RIP1 promotes the formation of complex II34, loss of cIAP ubiquitin ligases promotes the spontaneous formation of the ripoptosome16,28. As with complex II, the incorporation of RIP3 into the ripoptosome generates a necrotic signaling complex16,28. Apoptotic and necrotic signaling complexes share several components and the presence of RIP3 determines if the complex type is necroptotic or apoptotic. Although it has not been reported yet, it could be predicted that necroptosis induced by the TCR, TLR4 and other signals requires necrotic complexes containing RIP3. TRIF is an adaptor for TLR3 and TLR4 that interacts with RIP1 through its RIP homotypic interaction motif (RHIM)42,43,44. As RHIM is also present in RIP3, it is possible that the necrotic complex induced by signaling via TLR3 and TLR4 is formed through the interaction of TRIF with RIP1and RIP3. In addition, the RNA sensor RIG-I is known to recruit caspase-8 and RIP1 (ref. 45), so it is possible necroptosis could occur if RIP3 were recruited to the RIG-I complex under certain conditions. On the basis of published work, we propose that different death stimuli are recognized or sensed by specific receptors or sensors on the cell surface or inside cells and the necrotic complexes that form may vary in their composition, but all should contain RIP3. Necroptosis is involved in a variety of physiological and pathological processes, including development and homeostasis of the immune system and the pathogenesis of some inflammatory diseases (Fig. 2).

Different necroptotic stimuli are recognized or sensed by specific receptors or sensors on the cell surface or inside cells. The initiation of the necroptotic response to different stimuli is mediated by different receptor-sensor complexes, although the nature of some of these complexes is unknown at present. The formation of the necrotic signaling complex determines cell fate. Different necrotic complexes are found after ligation of different receptors or under genotoxic stress. The necrosome, which contains TRADD, FADD, caspase-8, RIP1 and RIP3, is formed after the ligation of death receptors; the ripotosome, which has a composition slightly different from that of necrosome, is formed in response to double-stranded RNA or genotoxic stress. In each complex, the release of RIP1-RIP3 from suppression by caspase-8 is required for necroptosis. As TRIF can interact with RIP1 and RIP3 through its RHIM domain, it probably forms a necrotic complex with RIP1 and RIP3 downstream of TLR3-TLR4 signaling. As necroptosis induced by the double-stranded DNA virus MCMV is RIP3 dependent and RIP1 independent, a viral or cellular protein (or proteins) might directly interact with RIP3 to induce necroptosis of host cells. It is clear that the necrotic complexes are heterogenous in their components, but all contain RIP3. The necroptosis induced by the various stimuli participates in a variety of biological processes, including cell death induced by death receptors, pathogens and genotoxic stress, and contributes to T cell population expansion, homeostasis, embryogenesis and the pathogenesis of many inflammation-related diseases. FAF, Fas-associated factor; Anti-, antibody; CD3, invariant signaling protein; CD28, coreceptor; Lck, Zap70 and Fyn, tyrosine kinases; PLC-γ, phospholipase C-γ; LPS, lipopolysaccharide; MD2, TLR4 coreceptor; CD14, lipopolysaccharide receptor; TIRAP, MyD88, TRIF and TRAM, adaptors; dsRNA, double-stranded RNA; IAPs, inhibitors of apoptosis.
RIP3 and RIP1 are members of the same kinase family, and each has an N-terminal kinase domain46. Unlike RIP1, RIP3 does not contain a death domain. RIP1 and RIP3 interact through their C-terminal RHIM domain. Deletion of either RIP1 or RIP3 abolishes TNF- or Fas-induced necroptosis10,11,13,14,15, which indicates the requirement for these kinases in death receptor–mediated necroptosis. Moreover, their interaction is also crucial, as mutation of sequence encoding the RHIM domain impairs their function in necroptosis13,14,15. Necrostatin-1, an inhibitor of the kinase activity of RIP1, blocks the interaction between RIP1 and RIP3, which indicates that the kinase activity of RIP1 is indispensable for assembly of the RIP1-RIP3 complex13,14. It is not clear whether the kinase activity of RIP3 is required for its interaction with RIP1, but phosphorylation of both RIP1 and RIP3 is considered to be a driving force for assembly of the RIP1-RIP3 complex13. Ser161 in RIP1 and Ser199 in RIP3 have been identified as phosphorylation sites14,36, but mutagenesis studies have shown that inhibition of phosphorylation at these sites has little influence on TNF-induced necroptosis5, which suggests the presence of other functional phosphorylation sites. Although dimerization of RIP1 and RIP3 is commonly described in the formation of the necrotic complexes, it is unclear exactly how many RIP1 and RIP3 molecules are in these complexes. As dimerization of RIP1 can induce necroptosis6 and necroptosis requires RIP3 (ref. 15), it is possible that the necrotic complex contains more than one RIP1-RIP3 dimer.
Although both RIP1 and RIP3 are required for necroptosis initiated by many different triggers, including death receptors and pathogen-recognition receptors, necroptosis can occur in the absence of RIP1. MCMV infection can induce RIP3-dependent necroptosis in the absence of RIP1 (ref. 17), although the sensor for MCMV-induced necroptosis is unknown at present (Fig. 2). The cytosolic receptor DAI could be a candidate here, as it can sense double-stranded DNA under certain conditions47 and contains a RHIM domain. Recruitment of RIP1 and RIP3 by DAI has been reported, and this process can be inhibited by the RHIM-containing MCMV protein M45 (refs. 48,49), which indicates a RHIM-dependent interaction. DAI-induced activation of NF-κB requires both RIP1 and RIP3 (ref. 47), but cell death induced by DAI overexpression, like MCMV-induced death17, requires only RIP3 (J.H., C.-Q.Z. and D.-W.Z., unpublished data). It is possible that the DAI-RIP3 interaction induces necroptosis, whereas the DAI-RIP1-RIP3 complex activates NF-κB. A cell-based study has also supported the notion that RIP3 can trigger necroptosis without RIP1, as RIP3 overexpression leads to necroptosis of Ripk1−/− cells15. In contrast, RIP1-mediated necroptosis requires RIP3, as RIP1 overexpression leads to necroptosis of wild-type MEFs but leads to apoptosis of RIP3-deficient (Ripk3−/−) MEFs15. Thus, RIP3 is indispensable for necroptosis, whereas RIP1 participates only in certain stimulus-induced cell necroptosis.
Mitochondrial reactive oxygen species (ROS) are crucial for the necrotic response of L929 cells to TNF50. Activation of RIP3 may link TNFR1 signaling to mitochondrial bioenergetics and subsequent overproduction of ROS. RIP3 physically interacts with and activates several metabolic enzymes, which can enhance glycogenolysis, glycolysis and glutaminolysis and thus contribute to the respiratory burst15. ROS production by the plasma-membrane NADPH oxidase NOX1 can also contribute to TNF-induced necrotic cell death in some experimental scenarios51. However, it is noteworthy that TNF-induced necroptosis in HT-29 colon adenocarcinoma cells is not ROS dependent14, which indicates the existence of other operative executing mechanisms.
Proteins of the Bcl-2 family regulate necroptosis, as overexpression of Bcl-XL inhibits TNF-induced necroptosis52. Bcl-XL most probably protects mitochondria from ROS-mediated damage, but whether RIP1-RIP3 has functional linkage to Bcl-2 family members remains unknown. The NF-κB modulator NEMO (IKKγ) may have an NF-κB-independent function that links RIP1-RIP3 to aberrant mitochondrial function53. Also, a RIP1-dependent signal is involved in the inhibition of adenine nucleotide translocase54, which may be responsible for the increase in the mitochondrial transmembrane potential observed in necroptotic cells. Other mitochondrial events, such as the release of apoptosis-inducing factor, may participate in necroptosis55, but whether calpain activation, a process required for that release56,57,58, is involved in necroptosis is not yet clear.
Depletion of ATP has been proposed to promote necrosis, as overactivation of the poly(ADP-ribose) polymerase PARP-1, an ATP-consuming process, has been noted in TNF-induced necroptosis59. However, another study has shown that treatment with TNF does not lead to lower intracellular ATP concentrations in L929 cells52, which indicates that the decrease in ATP is dispensable for necroptosis. TNF-induced necroptosis of L929 cells is associated with a greater total lysosomal volume and more enzymatic activities60,61, and the degree of lysosomal response correlates with necroptosis62,63. Lysosomal membrane permeabilization, which allows the release of lysosomal enzymes or other contents into the cytoplasm and has been linked to the initiation of apoptosis64, probably participates in necroptosis65,66,67,68,69,70,71.
So far, many cellular events have been reported to act downstream of the necrotic signaling complex to execute necrosis, but their involvement in necroptosis seems to be dependent on the cell type and stimulus. Therefore, further investigation is required for complete understanding of the relative importance of these intracellular mediators.
Interplay between apoptosis and necroptosis
In some cell lines and primary cells, inhibition of caspases blocks apoptosis, but the cells then die of necroptosis. Inhibition of caspase-8 is mainly responsible for this phenomenon10,72. Studies have shown that FADD and caspase-8, the core proteins of the apoptosis signaling complexes (complex II and the ripoptosome), act together to suppress necroptosis during development, as well as in adult tissues or cells. Deletion of Fadd or Casp8 causes mice to die in utero at mid-gestation, with defects in cardiovascular and blood cell formation. It is clear now that the death of caspase-8-deficient (Casp8−/−) and FADD-deficient (Fadd−/−) mice results from the loss of suppression of RIP1-RIP3–dependent necroptosis, as crossing FADD-deficient or caspase-8-deficient mice onto a Ripk1−/− or Ripk3−/− background rescues mice from embryonic death37,38,39. Casp8−/−Ripk3−/− double-mutant embryos have functional heart and vascular systems and are able to mature into fertile adults37,39. The embryogenesis of Fadd−/−Ripk1−/− embryos is normal, but the mice die shortly after birth because of the absence of RIP1 (ref. 38). Casp8−/−Ripk3−/− mice seem to be immunocompetent with a full complement of myeloid and lymphoid cell types. Depletion of RIP3 completely rescues caspase-8-null T cells from antigen receptor–induced necroptosis23 and restores the proliferation of T cells bearing an interfering FADD mutant (FADDdd)41. RIP1 deficiency restores the proliferation of FADD-null T cells but not that of FADD-null B cells, which suggests that FADD might inhibit necroptosis only in certain cell types38. Nevertheless, the suppressive effect of the caspase-8–FADD complex on necroptosis seems to be important for the normal development and homeostasis of the immune system.
The amount of RIP3 protein expression in various cell lines correlates with their responsiveness to necroptosis induction14. RIP3 overexpression switches the TNF-induced death of NIH3T3 mouse fibroblasts to necroptosis15, which suggests that RIP3 can suppress caspase-dependent apoptosis in some experimental settings. RIP3 has different expression in different cell types, with high protein expression in macrophages and lymphocytes, yet barely detectable expression in retina and cardiac myocytes. Thus, it is likely that necroptosis does not happen in some cell types under normal situations. A switch from apoptosis to necrosis could occur if RIP3 is induced under a given physiological or pathological condition in cells with low or no RIP3 expression. In a model of acute pancreatitis, RIP3 protein is considerably induced in the pancreas and necroptosis emerges14. Induction of RIP3 has also been reported in many other disease-related conditions, including hepatocytes showing steatohepatitis73, photoreceptors after retinal detachment74, and vascular smooth muscle cells that have undergone arterial injury stimulation75. Samples of the terminal ileum from patients with Crohn's disease show high RIP3 expression in Paneth cells76, which suggests association of RIP3 with the pathogenesis of this disease. The RIP3 induction probably promotes the necroptosis observed in these conditions. Atherosclerosis is a chronic inflammatory disease in which macrophage death has an important role77. In the early stage of disease, macrophage death in atherosclerotic plaques is mainly apoptotic, but in the advanced disease stage, necroptosis becomes predominant (J.H., C.-Q.Z. and D.-W.Z., unpublished data). This transition is probably related to RIP3 activity during the course of disease development, which suggests that RIP3 somehow controls the switch from apoptosis to necroptosis in vivo.
Although suppression of necroptosis is important for the normal development of mice, the necroptosis pathway seems to be a backup for apoptosis in the elimination of excess T cells23,37,39. Fas ligation–induced T cell death is essential for T cell homeostasis, as abnormal T cells accumulate with age in Fas-deficient lpr/lpr mice or Fas ligand–deficient gld/gld mice78,79. Deletion of Casp8 or Fadd does not lead to lymphoproliferative disease, although sometimes a modest lymphoproliferation can be observed80,81. Interestingly, mice that lack both Casp8 and Ripk3, as well as those with T cell–specific deletion of Casp8 or with T cells expressing FADDdd on a Ripk3−/− background, develop a progressive disease with a high degree of lymphocyte accumulation, which resembles that of lpr/lpr and gld/gld mice23,37,39,41. Thus, it seems that RIP3 functions to eliminate excess T cells when caspase-8 is absent, or perhaps even together with caspase-8 under specific conditions.
Although RIP3-dependent necroptosis is a backup for caspase-8-mediated apoptosis in Fas ligation–induced T cell death, RIP1-RIP3–induced necroptosis can lead to a defect in antigen receptor–stimulated T cell clonal expansion when RIP3 suppression by caspase-8 or FADD is lifted41,82. FADDdd mice are unable to mount an effective immune response to viral pathogens such as mouse hepatitis virus because of a defect in T cell activation and survival83. RIP3 clearly has a key role in this phenotype, as crossing FADDdd mice onto a Ripk3−/− background rescues mice from the defects in virus-specific T cell infiltration of the infected tissue, cytotoxic T cell activity and clearance of mouse hepatitis virus41. As deletion of Ripk3 restores the expansion capacity of antiviral T cells bearing FADDdd, RIP3-mediated inhibition of T cell population expansion is the cause of the failure of FADDdd-expressing mice to mount an antigen-specific response.
Mechanistic insight about the choice of a cell death pathway is largely unknown. However, studies of TNF-induced cell death have provided some information about the differences between apoptotic and necroptotic signaling complexes. TRADD-deficient cells are resistant to apoptosis and necroptosis induced by TNF84, but TRADD is not detected in complex II formed after ligation of TNFR in the presence of mimetics of the supramolecular activation cluster (chemical agents that block cIAP proteins by mimicking the activity of the supramolecular activation cluster)35 or in the ripoptosome16,28, which suggests that TRADD is an upstream signaling molecule that is dispensable for downstream cell-death processes. As noted above, FADD, RIP1 and caspase-8 are required for the assembly of complex II85. If RIP3 is present, the incorporation of RIP3 into complex II will generate complex IIb or the necrosome5,36 (Fig. 1). The interaction of FLIP long form (FLIPL) with caspase-8 in this complex is essential for necroptotic signaling39. In the presence of FLIPL, caspase-8 lacks enough activity to initiate apoptosis, but its proteolytic ability can block necroptosis efficiently by directly cleaving RIP1 and RIP3 (refs. 86,87). A study has shown that RIP1 and RIP3 are cleaved in OT-I T cells (which express an ovalbumin-specific TCR) after stimulation with ovalbumin peptide or antibody to Fas, as well as in Jurkat T cells stimulated with TNF or TRAIL (a ligand for a cell surface receptor)41. Cleavage of RIP1 and RIP3 prevents the elaboration of a necroptotic response after ligation of the TCR or death receptor. The data showing that deletion of Fadd or Casp8 converts the cell death to necroptosis support the idea that caspase-8 activity negatively regulates necroptosis. However, overexpression of RIP3 may shift the balance of intact RIP3 versus cleaved RIP3 toward the intact form, thus converting apoptosis to necrosis. However, it is not clear how overexpression of RIP3 can suppress TNF-induced activation of caspase-8.
Effect of necroptosis on inflammation
Necrosis is recognized as a cause for inflammation because the release of intracellular materials (including SAP130, heat-shock proteins, histones, high-mobility group protein B1, RNA, DNA and so on) can trigger inflammatory reactions. However, not all examples of necrosis are proinflammatory88. Sometimes necrotic cells can inhibit inflammatory reactions89. It has been shown that macrophages can engulf necroptotic L929 cells without producing inflammatory cytokines90, which suggests an unexpected complexity in the immune response to necrotic cells. Nevertheless, necroptosis triggers inflammation, as shown by the intestinal inflammation caused by RIP3-dependent cell death in mice with deletion of Fadd40 or Casp8 (ref. 76) specific to intestinal epithelial cells (IECs) and by liver injury resulting from concanavalin A–induced, RIP1-RIP3–dependent necroptosis in mice bearing hepatocyte-specific deletion of Casp8 (ref. 91). Mice with IEC-specific deletion of Fadd or Casp8 spontaneously develop inflammatory lesions in the intestine in conjunction with loss of Paneth cells. RIP3-dependent necroptosis is believed to have an essential role in the chronic intestinal inflammation in these mice, as deletion of Ripk3 prevents the development of pathological changes in the small intestine and colons of mice with IEC-specific deletion of Fadd. It is notable that the triggers of RIP3-dependent necroptosis of FADD-deficient IECs in the small and large bowel seem to be different, as bacteria-induced production of TNF and other cytokines drives colitis in the colon but has no effect on Paneth cell loss and enteritis in mice with IEC-specific deletion of Fadd. As high RIP3 expression is found in Paneth cells of patients with Crohn's disease76, RIP3-mediated necroptosis may have a role in the pathogenesis of this disease.
Necroptosis has been characterized in animal models of acute pancreatitis, ischemic injury and neurodegeneration6,14,17,92. Its occurrence should stimulate the immune system to elicit inflammatory responses, and the inflammation could have an important role in the pathogenesis of these diseases. RIP3-dependent necroptosis can occur during infectious disease, as pathogen-recognition receptors can elicit necroptosis. Viruses such as vaccinia virus elicit RIP3-dependent necroptosis of host cells. The virus-induced necroptosis may have a positive role in limiting viral replication13 and enhancing the inflammation that facilitates the clearance of virus from the inflamed sites. In addition, a role for necroptosis in the phenomenon of immunosurveillance in tumor development is possible and awaits investigation. Overall, necroptosis is an endogenous trigger of inflammation that either positively or negatively affects an organism's health.
Perspectives
The multiple lines of evidence reviewed here illustrate that necroptosis is an important, intrinsic cellular death program. It has a role in the development and maintenance of the mature immune system. A major challenge ahead is the development of approaches that distinguish necroptosis from other forms of cell death in vivo. Although gene-deficient mice and necroptosis-inhibiting necrostatins can be used in animal studies, the method for detecting necrosis in human samples is limited to electron microscopy, which cannot distinguish necroptotic cells from other necrotic cells. For better understanding of necroptosis in human diseases, biomarkers must be identified that distinguish necroptosis from apoptosis. This last will be crucial for advancing the knowledge of how these pathways interact in many human diseases.
Tissue-specific deletion of Casp8 or the component of complex II FADD has been reported. The suppression of RIP3-dependent necroptosis by caspase-8 must be taken into consideration for reinterpretation of the data obtained in such studies (for example, is the phenotype due to necroptosis?). Analysis of the survival of Casp8−/− T cells rescued by RIP3 depletion serves as an example in which RIP3-mediated necroptosis may account for the death of Casp8−/− T cells23. Similarly, the phenotypes that develop after specific deletion of Casp8 in B cells93,94, the liver95,96, endothelial cells96 or epidermal keratinocytes97,98 may require reevaluation, as a wide range of inflammatory abnormalities arise after disruption of caspase-8 in which RIP1-RIP3–dependent necroptosis may determine an underlying mechanism.
The normal development of mice and the immune system on the Casp8−/−Ripk3−/− background raises the question of the importance of the two death pathways mediated by caspase-8 and RIP3. It is clear that Fas-mediated cell death depends heavily on the two pathways, whereas these pathways are dispensable or can be compromised in certain biological processes, as indicated by the fact that Ripk3−/− mouse expressing FADDdd can mount an effective immune response to mouse hepatitis virus. Ripk3−/− mice are viable without apparent defects99, which suggests that necroptosis may not be essential under normal conditions when caspase-8-dependent apoptotic pathway is intact. However, necroptosis has an important role in the pathogenesis of diseases such as inflammation-related pancreatitis and atherosclerosis and viral infection, as well as ischemic injury and neurodegeneration6,14,17,92. It is very likely that necroptosis begins to predominate during the development of many diseases, as higher RIP3 expression is observed in many disease models14,73,74,75. Given the fact that caspase-8 is normal in most organisms, the precise mechanisms that underlie the switch between apoptosis and necroptosis could be a 'hot spot' in this field.
Having been recognized for more than 20 years, necroptosis is now a major topic in the field of cell-death research. Studies of necroptosis have brought new mechanistic insight into cell death–related biological processes. Because of the potential involvement of necroptosis in many human diseases, we speculate that the investigation of necroptosis will provide new targets for clinical applications of cytotoxic and cytoprotective therapies and will lead to better treatment of infectious and inflammatory diseases.
References
Feig, C. & Peter, M.E. How apoptosis got the immune system in shape. Eur. J. Immunol. 37 (suppl. 1), S61–S70 (2007).
Opferman, J.T. & Korsmeyer, S.J. Apoptosis in the development and maintenance of the immune system. Nat. Immunol. 4, 410–415 (2003).
Chowdhury, D. & Lieberman, J. Death by a thousand cuts: granzyme pathways of programmed cell death. Annu. Rev. Immunol. 26, 389–420 (2008).
Vandenabeele, P., Galluzzi, L., Vanden Berghe, T. & Kroemer, G. Molecular mechanisms of necroptosis: an ordered cellular explosion. Nat. Rev. Mol. Cell Biol. 11, 700–714 (2010).
Christofferson, D.E. & Yuan, J. Necroptosis as an alternative form of programmed cell death. Curr. Opin. Cell Biol. 22, 263–268 (2010).
Degterev, A. et al. Chemical inhibitor of nonapoptotic cell death with therapeutic potential for ischemic brain injury. Nat. Chem. Biol. 1, 112–119 (2005).
Laster, S.M., Wood, J.G. & Gooding, L.R. Tumor necrosis factor can induce both apoptic and necrotic forms of cell lysis. J. Immunol. 141, 2629–2634 (1988).
Roach, H.I. & Clarke, N.M. Physiological cell death of chondrocytes in vivo is not confined to apoptosis. New observations on the mammalian growth plate. J. Bone Joint Surg. Br. 82, 601–613 (2000).
Barkla, D.H. & Gibson, P.R. The fate of epithelial cells in the human large intestine. Pathology 31, 230–238 (1999).
Holler, N. et al. Fas triggers an alternative, caspase-8-independent cell death pathway using the kinase RIP as effector molecule. Nat. Immunol. 1, 489–495 (2000).
Lin, Y. et al. Tumor necrosis factor-induced nonapoptotic cell death requires receptor-interacting protein-mediated cellular reactive oxygen species accumulation. J. Biol. Chem. 279, 10822–10828 (2004).
Hitomi, J. et al. Identification of a molecular signaling network that regulates a cellular necrotic cell death pathway. Cell 135, 1311–1323 (2008).
Cho, Y.S. et al. Phosphorylation-driven assembly of the RIP1–RIP3 complex regulates programmed necrosis and virus-induced inflammation. Cell 137, 1112–1123 (2009).
He, S. et al. Receptor interacting protein kinase-3 determines cellular necrotic response to TNF-α. Cell 137, 1100–1111 (2009).
Zhang, D.W. et al. RIP3, an energy metabolism regulator that switches TNF-induced cell death from apoptosis to necrosis. Science 325, 332–336 (2009).
Feoktistova, M. et al. cIAPs block ripoptosome formation, a RIP1/caspase-8 containing Intracellular cell death complex differentially regulated by cFLIP isoforms. Mol. Cell 43, 449–463 (2011).
Upton, J.W., Kaiser, W.J. & Mocarski, E.S. Virus inhibition of RIP3-dependent necrosis. Cell Host Microbe 7, 302–313 (2010).
Martinon, F., Gaide, O., Petrilli, V., Mayor, A. & Tschopp, J. NALP inflammasomes: a central role in innate immunity. Semin. Immunopathol. 29, 213–229 (2007).
Kalai, M. et al. Tipping the balance between necrosis and apoptosis in human and murine cells treated with interferon and dsRNA. Cell Death Differ. 9, 981–994 (2002).
Ma, Y., Temkin, V., Liu, H. & Pope, R.M. NF-κB protects macrophages from lipopolysaccharide-induced cell death: the role of caspase 8 and receptor-interacting protein. J. Biol. Chem. 280, 41827–41834 (2005).
François, M., Le Cabec, V., Dupont, M.A., Sansonetti, P.J. & Maridonneau-Parini, I. Induction of necrosis in human neutrophils by Shigella flexneri requires type III secretion, IpaB and IpaC invasins, and actin polymerization. Infect. Immun. 68, 1289–1296 (2000).
Koterski, J.F., Nahvi, M., Venkatesan, M.M. & Haimovich, B. Virulent Shigella flexneri causes damage to mitochondria and triggers necrosis in infected human monocyte-derived macrophages. Infect. Immun. 73, 504–513 (2005).
Ch'en, I.L., Tsau, J.S., Molkentin, J.D., Komatsu, M. & Hedrick, S.M. Mechanisms of necroptosis in T cells. J. Exp. Med. 208, 633–641 (2011).
Galluzzi, L. et al. Viral strategies for the evasion of immunogenic cell death. J. Intern. Med. 267, 526–542 (2010).
Lamkanfi, M. & Dixit, V.M. Manipulation of host cell death pathways during microbial infections. Cell Host Microbe 8, 44–54 (2010).
Ray, C.A. & Pickup, D.J. The mode of death of pig kidney cells infected with cowpox virus is governed by the expression of the crmA gene. Virology 217, 384–391 (1996).
Davis, C.W. et al. Nitration of the mitochondrial complex I subunit NDUFB8 elicits RIP1- and RIP3-mediated necrosis. Free Radic. Biol. Med. 48, 306–317 (2010).
Tenev, T. et al. The ripoptosome, a signaling platform that assembles in response to genotoxic stress and loss of IAPs. Mol. Cell 43, 432–448 (2011).
Micheau, O. & Tschopp, J. Induction of TNF receptor I-mediated apoptosis via two sequential signaling complexes. Cell 114, 181–190 (2003).
Bertrand, M.J. et al. cIAP1 and cIAP2 facilitate cancer cell survival by functioning as E3 ligases that promote RIP1 ubiquitination. Mol. Cell 30, 689–700 (2008).
Häcker, H. & Karin, M. Regulation and function of IKK and IKK-related kinases. Sci. STKE 2006, re13 (2006).
Van Antwerp, D.J., Martin, S.J., Kafri, T., Green, D.R. & Verma, I.M. Suppression of TNF-α-induced apoptosis by NF-κB. Science 274, 787–789 (1996).
Ea, C.K., Deng, L., Xia, Z.P., Pineda, G. & Chen, Z.J. Activation of IKK by TNFα requires site-specific ubiquitination of RIP1 and polyubiquitin binding by NEMO. Mol. Cell 22, 245–257 (2006).
Wong, W.W. et al. RIPK1 is not essential for TNFR1-induced activation of NF-κB. Cell Death Differ. 17, 482–487 (2010).
Wang, L., Du, F. & Wang, X. TNF-α induces two distinct caspase-8 activation pathways. Cell 133, 693–703 (2008).
Vandenabeele, P., Declercq, W., Van Herreweghe, F. & Vanden Berghe, T. The role of the kinases RIP1 and RIP3 in TNF-induced necrosis. Sci. Signal. 3, re4 (2010).
Kaiser, W.J. et al. RIP3 mediates the embryonic lethality of caspase-8-deficient mice. Nature 471, 368–372 (2011).
Zhang, H. et al. Functional complementation between FADD and RIP1 in embryos and lymphocytes. Nature 471, 373–376 (2011).
Oberst, A. et al. Catalytic activity of the caspase-8-FLIPL complex inhibits RIPK3-dependent necrosis. Nature 471, 363–367 (2011).
Welz, P.S. et al. FADD prevents RIP3-mediated epithelial cell necrosis and chronic intestinal inflammation. Nature 477, 330–334 (2011).
Lu, J.V. et al. Complementary roles of Fas-associated death domain (FADD) and receptor interacting protein kinase-3 (RIPK3) in T-cell homeostasis and antiviral immunity. Proc. Natl. Acad. Sci. USA 108, 15312–15317 (2011).
Cusson-Hermance, N., Khurana, S., Lee, T.H., Fitzgerald, K.A. & Kelliher, M.A. Rip1 mediates the Trif-dependent toll-like receptor 3- and 4-induced NF-κB activation but does not contribute to interferon regulatory factor 3 activation. J. Biol. Chem. 280, 36560–36566 (2005).
Kaiser, W.J. & Offermann, M.K. Apoptosis induced by the toll-like receptor adaptor TRIF is dependent on its receptor interacting protein homotypic interaction motif. J. Immunol. 174, 4942–4952 (2005).
Meylan, E. et al. RIP1 is an essential mediator of Toll-like receptor 3-induced NF-κB activation. Nat. Immunol. 5, 503–507 (2004).
Rajput, A. et al. RIG-I RNA helicase activation of IRF3 transcription factor is negatively regulated by caspase-8-mediated cleavage of the RIP1 protein. Immunity 34, 340–351 (2011).
Zhang, D., Lin, J. & Han, J. Receptor-interacting protein (RIP) kinase family. Cell. Mol. Immunol. 7, 243–249 (2010).
Takaoka, A. et al. DAI (DLM-1/ZBP1) is a cytosolic DNA sensor and an activator of innate immune response. Nature 448, 501–505 (2007).
Rebsamen, M. et al. DAI/ZBP1 recruits RIP1 and RIP3 through RIP homotypic interaction motifs to activate NF-κB. EMBO Rep. 10, 916–922 (2009).
Kaiser, W.J., Upton, J.W. & Mocarski, E.S. Receptor-interacting protein homotypic interaction motif-dependent control of NF-κB activation via the DNA-dependent activator of IFN regulatory factors. J. Immunol. 181, 6427–6434 (2008).
Schulze-Osthoff, K. et al. Cytotoxic activity of tumor necrosis factor is mediated by early damage of mitochondrial functions. Evidence for the involvement of mitochondrial radical generation. J. Biol. Chem. 267, 5317–5323 (1992).
Kim, Y.S., Morgan, M.J., Choksi, S. & Liu, Z.G. TNF-induced activation of the Nox1 NADPH oxidase and its role in the induction of necrotic cell death. Mol. Cell 26, 675–687 (2007).
Ono, K. et al. Metaxin deficiency alters mitochondrial membrane permeability and leads to resistance to TNF-induced cell killing. Protein Cell 1, 161–173 (2010).
Irrinki, K.M. et al. Requirement of FADD, NEMO, and BAX/BAK for aberrant mitochondrial function in tumor necrosis factor α-induced necrosis. Mol. Cell. Biol. 31, 3745–3758 (2011).
Temkin, V., Huang, Q., Liu, H., Osada, H. & Pope, R.M. Inhibition of ADP/ATP exchange in receptor-interacting protein-mediated necrosis. Mol. Cell. Biol. 26, 2215–2225 (2006).
Xu, X. et al. The role of PARP activation in glutamate-induced necroptosis in HT-22 cells. Brain Res. 1343, 206–212 (2010).
Yu, S.W. et al. Mediation of poly(ADP-ribose) polymerase-1-dependent cell death by apoptosis-inducing factor. Science 297, 259–263 (2002).
Cao, G. et al. Critical role of calpain I in mitochondrial release of apoptosis-inducing factor in ischemic neuronal injury. J. Neurosci. 27, 9278–9293 (2007).
Moubarak, R.S. et al. Sequential activation of poly(ADP-ribose) polymerase 1, calpains, and Bax is essential in apoptosis-inducing factor-mediated programmed necrosis. Mol. Cell. Biol. 27, 4844–4862 (2007).
Los, M. et al. Activation and caspase-mediated inhibition of PARP: a molecular switch between fibroblast necrosis and apoptosis in death receptor signaling. Mol. Biol. Cell 13, 978–988 (2002).
Ono, K., Kim, S.O. & Han, J. Susceptibility of lysosomes to rupture is a determinant for plasma membrane disruption in tumor necrosis factor α-induced cell death. Mol. Cell. Biol. 23, 665–676 (2003).
Schneider-Brachert, W. et al. Compartmentalization of TNF receptor 1 signaling: internalized TNF receptosomes as death signaling vesicles. Immunity 21, 415–428 (2004).
Ono, K., Wang, X. & Han, J. Resistance to tumor necrosis factor-induced cell death mediated by PMCA4 deficiency. Mol. Cell. Biol. 21, 8276–8288 (2001).
Liddil, J.D., Dorr, R.T. & Scuderi, P. Association of lysosomal activity with sensitivity and resistance to tumor necrosis factor in murine L929 cells. Cancer Res. 49, 2722–2728 (1989).
Boya, P. & Kroemer, G. Lysosomal membrane permeabilization in cell death. Oncogene 27, 6434–6451 (2008).
Suffys, P. et al. Tumour-necrosis-factor-mediated cytotoxicity is correlated with phospholipase-A2 activity, but not with arachidonic acid release per se. Eur. J. Biochem. 195, 465–475 (1991).
Kim, W.H., Choi, C.H., Kang, S.K., Kwon, C.H. & Kim, Y.K. Ceramide induces non-apoptotic cell death in human glioma cells. Neurochem. Res. 30, 969–979 (2005).
Yamashima, T. & Oikawa, S. The role of lysosomal rupture in neuronal death. Prog. Neurobiol. 89, 343–358 (2009).
Kirkegaard, T. et al. Hsp70 stabilizes lysosomes and reverts Niemann-Pick disease-associated lysosomal pathology. Nature 463, 549–553 (2010).
Nylandsted, J. et al. Heat shock protein 70 promotes cell survival by inhibiting lysosomal membrane permeabilization. J. Exp. Med. 200, 425–435 (2004).
Tang, D. et al. Nuclear heat shock protein 72 as a negative regulator of oxidative stress (hydrogen peroxide)-induced HMGB1 cytoplasmic translocation and release. J. Immunol. 178, 7376–7384 (2007).
Doulias, P.T. et al. Involvement of heat shock protein-70 in the mechanism of hydrogen peroxide-induced DNA damage: the role of lysosomes and iron. Free Radic. Biol. Med. 42, 567–577 (2007).
Vercammen, D. et al. Inhibition of caspases increases the sensitivity of L929 cells to necrosis mediated by tumor necrosis factor. J. Exp. Med. 187, 1477–1485 (1998).
Zorde-Khvalevsky, E. et al. Toll-like receptor 3 signaling attenuates liver regeneration. Hepatology 50, 198–206 (2009).
Trichonas, G. et al. Receptor interacting protein kinases mediate retinal detachment-induced photoreceptor necrosis and compensate for inhibition of apoptosis. Proc. Natl. Acad. Sci. USA 107, 21695–21700 (2010).
Li, Q. et al. Receptor interacting protein 3 suppresses vascular smooth muscle cell growth by inhibition of the phosphoinositide 3-kinase-Akt axis. J. Biol. Chem. 285, 9535–9544 (2010).
Günther, C. et al. Caspase-8 regulates TNF-α-induced epithelial necroptosis and terminal ileitis. Nature 477, 335–339 (2011).
Moore, K.J. & Tabas, I. Macrophages in the pathogenesis of atherosclerosis. Cell 145, 341–355 (2011).
Nagata, S. & Suda, T. Fas and Fas ligand: lpr and gld mutations. Immunol. Today 16, 39–43 (1995).
Watanabe-Fukunaga, R., Brannan, C.I., Copeland, N.G., Jenkins, N.A. & Nagata, S. Lymphoproliferation disorder in mice explained by defects in Fas antigen that mediates apoptosis. Nature 356, 314–317 (1992).
Hedrick, S.M., Ch'en, I.L. & Alves, B.N. Intertwined pathways of programmed cell death in immunity. Immunol. Rev. 236, 41–53 (2010).
Salmena, L. & Hakem, R. Caspase-8 deficiency in T cells leads to a lethal lymphoinfiltrative immune disorder. J. Exp. Med. 202, 727–732 (2005).
Bell, B.D. et al. FADD and caspase-8 control the outcome of autophagic signaling in proliferating T cells. Proc. Natl. Acad. Sci. USA 105, 16677–16682 (2008).
Beisner, D.R., Chu, I.H., Arechiga, A.F., Hedrick, S.M. & Walsh, C.M. The requirements for Fas-associated death domain signaling in mature T cell activation and survival. J. Immunol. 171, 247–256 (2003).
Ermolaeva, M.A. et al. Function of TRADD in tumor necrosis factor receptor 1 signaling and in TRIF-dependent inflammatory responses. Nat. Immunol. 9, 1037–1046 (2008).
Vucic, D., Dixit, V.M. & Wertz, I.E. Ubiquitylation in apoptosis: a post-translational modification at the edge of life and death. Nat. Rev. Mol. Cell Biol. 12, 439–452 (2011).
Feng, S. et al. Cleavage of RIP3 inactivates its caspase-independent apoptosis pathway by removal of kinase domain. Cell. Signal. 19, 2056–2067 (2007).
Lin, Y., Devin, A., Rodriguez, Y. & Liu, Z.G. Cleavage of the death domain kinase RIP by caspase-8 prompts TNF-induced apoptosis. Genes Dev. 13, 2514–2526 (1999).
Casares, N. et al. Caspase-dependent immunogenicity of doxorubicin-induced tumor cell death. J. Exp. Med. 202, 1691–1701 (2005).
Hirt, U.A. & Leist, M. Rapid, noninflammatory and PS-dependent phagocytic clearance of necrotic cells. Cell Death Differ. 10, 1156–1164 (2003).
Brouckaert, G. et al. Phagocytosis of necrotic cells by macrophages is phosphatidylserine dependent and does not induce inflammatory cytokine production. Mol. Biol. Cell 15, 1089–1100 (2004).
Liedtke, C. et al. Absence of Caspase-8 protects from inflammation-related hepatocarcinogenesis in mice but triggers non-apoptotic liver injury. Gastroenterology published online, doi:10.1053/j.gastro.2011.08.037 (28 August 2011).
Artal-Sanz, M. & Tavernarakis, N. Proteolytic mechanisms in necrotic cell death and neurodegeneration. FEBS Lett. 579, 3287–3296 (2005).
Imtiyaz, H.Z. et al. The Fas-associated death domain protein is required in apoptosis and TLR-induced proliferative responses in B cells. J. Immunol. 176, 6852–6861 (2006).
Beisner, D.R., Ch'en, I.L., Kolla, R.V., Hoffmann, A. & Hedrick, S.M. Cutting edge: innate immunity conferred by B cells is regulated by caspase-8. J. Immunol. 175, 3469–3473 (2005).
Ben Moshe, T. et al. Role of caspase-8 in hepatocyte response to infection and injury in mice. Hepatology 45, 1014–1024 (2007).
Kang, T.B. et al. Caspase-8 serves both apoptotic and nonapoptotic roles. J. Immunol. 173, 2976–2984 (2004).
Kovalenko, A. et al. Caspase-8 deficiency in epidermal keratinocytes triggers an inflammatory skin disease. J. Exp. Med. 206, 2161–2177 (2009).
Li, C. et al. Development of atopic dermatitis-like skin disease from the chronic loss of epidermal caspase-8. Proc. Natl. Acad. Sci. USA 107, 22249–22254 (2010).
Newton, K., Sun, X. & Dixit, V.M. Kinase RIP3 is dispensable for normal NF-κBs, signaling by the B-cell and T-cell receptors, tumor necrosis factor receptor 1, and Toll-like receptors 2 and 4. Mol. Cell. Biol. 24, 1464–1469 (2004).
Acknowledgements
We thank R. Ulevitch and R. Cook for critical reading of the manuscript. Supported by the National Natural Science Foundation of China (30830092, 91029304, 30921005 and 81061160512; and 973 program 2009CB522200), the Sino-Swiss International Collaboration (2009DFA32760) and the Science Planning Program of Fujian province (2009J1010).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
About this article
Cite this article
Han, J., Zhong, CQ. & Zhang, DW. Programmed necrosis: backup to and competitor with apoptosis in the immune system. Nat Immunol 12, 1143–1149 (2011). https://doi.org/10.1038/ni.2159
Published:
Issue Date:
DOI: https://doi.org/10.1038/ni.2159
This article is cited by
-
Saracatinib inhibits necroptosis and ameliorates psoriatic inflammation by targeting MLKL
Cell Death & Disease (2024)
-
Different types of cell death and their shift in shaping disease
Cell Death Discovery (2023)
-
Standard of care drugs do not modulate activity of senescent primary human lung fibroblasts
Scientific Reports (2023)
-
Repurposing of Ibrutinib and Quizartinib as potent inhibitors of necroptosis
Communications Biology (2023)
-
Necroptosis in the developing brain: role in neurodevelopmental disorders
Metabolic Brain Disease (2023)
