





Abstract
Darwin's finches comprise a group of 15 species endemic to the Galápagos (14 species) and Cocos (1 species) Islands in the Pacific Ocean. The group is monophyletic and originated from an ancestral species that reached the Galápagos Archipelago from Central or South America. Descendants of this ancestor on the Archipelago then colonized Cocos Island. In the present study, we used sequences of two mitochondrial (mt) DNA segments (922 bp of the cytochrome b gene and 1,082 bp of the control region), as well as two nuclear markers (830 bp of numt2, consisting of 140 bp of mtDNA control region and 690 bp of flanking nuclear DNA; and 740 bp of numt3, consisting of 420 bp of mt cytochrome b sequence flanked by 320 bp of nuclear DNA) to identify the species group most closely related to the Darwin's finches. To this end, we analyzed the sequences of 28 species representing the main groups (tribes) of the family Fringillidae, as well as 2 outgroup species and 13 species of Darwin's finches. In addition, we used mtDNA cytochrome b sequences of some 180 additional Fringillidae species from the database for phylogeny reconstruction by maximum-parsimony, maximum-likelihood, minimum-evolution, and neighbor-joining methods. The study identifies the grassquit genus Tiaris, and specifically the species Tiaris obscura, as the nearest living relative of Darwin's finches among the species surveyed. Darwin's finches diverged from the Tiaris group shortly after the various extant species of Tiaris diverged from one another. The initial adaptive radiation of the Tiaris group apparently occurred on the Caribbean islands and then spread to Central and South America, from where the ancestors of Darwin's finches departed for the Galápagos Islands approximately 2.3 MYA, at the time of the dramatic climatic changes associated with the closure of the Panamanian isthmus and the onset of Pleistocene glaciation.
Introduction
The designation “Darwin's finches” refers to a group of 15 finch-like species, 14 of which are endemic to the Galápagos Archipelago (the Galápagos finches), while one is confined to Cocos Island in the Pacific Ocean (Lack 1947 ; Grant 1999 ). Gould (1837) , the ornithologist who, with the help of assistants, examined and described the bird skins collected by Charles Darwin during his trip around the world on H.M.S. Beagle, included all of the Galápagos finches available to him in the genus Geospiza. He divided the genus into the subgenera Geospiza, Camarhynchus, Cactornis, and Certhidea, which he assigned to the family Coccothraustinae, the hawfinch-like birds. In current classifications, Darwin's finches fall into five groups: ground finches (Geospiza, including the species assigned by Gould to the subgenus Cactornis); tree finches (Camarhynchus, Cactospiza); the vegetarian finch (Platyspiza); warbler finches (Certhidea); and the Cocos finch (Pinaroloxias). Dissenting classifications are discussed by Lack (1947) and Grant (1999) . On morphological grounds, all of the Darwin's finch species have been thought to be closely related to one another and to resemble finches (i.e., primarily seed-eating birds with cone-shaped bills; e.g., Snodgrass 1903 ; Sushkin 1925, 1929 ; Lowe 1936 ). Doubts were raised only about the Certhidea, which some authors thought was not a finch at all, but rather a warbler (i.e., a small perching bird with a thin, pointed bill). However, as early as the beginning of the last century, a comparative anatomical analysis by Snodgrass (1903) provided evidence for the close relationship of Certhidea to the rest of the Darwin's finches. Recent mitochondrial (mt) DNA sequence (Sato et al. 1999 ) and microsatellite (Petren, Grant, and Grant 1999 ) analyses strongly support the monophyletic status of the whole group of Darwin's finches. This conclusion is consistent with behavioral (Bowman 1983 ), karyotypic (Jo 1983 ), and biochemical (Yang and Patton 1981 ) studies. The phylogenetic relationships among Darwin's finches have been elucidated by analyses of mtDNA sequences (Sato et al. 1999 ; Freeland and Boag 1999a, 1999b ) and microsatellite markers (Petren, Grant, and Grant 1999 ).
The original assignment of the group to the Coccothraustinae has now been abandoned, and the group has been relegated to a separate taxon, Geospizinae or Geospizini. On morphological grounds, the Geospizini have been assigned to the finch family Fringillidae (e.g., Salvin 1876 ; Ridgway 1897 ; Rothschild and Hartert 1899, 1902 ; Snodgrass and Heller 1904 ; Sushkin 1925, 1929 ; Swarth 1931 ; Lowe 1936 ; Sibley and Ahlquist 1990 ) in the order Passeriformes. The taxonomy of the Fringillidae is currently in a state of flux, but the family is commonly divided into two subfamilies, Fringillinae and Emberizinae (e.g., Sibley and Ahlquist 1990 ). The Fringillinae include primarily Old World finches—the tribes Fringillini and Carduelini, as well as the Hawaiian honeycreepers (tribe Drepanidini). The Emberizinae consist primarily of New World taxa, specifically the tribes Emberizini (true buntings and New World sparrows), Parulini (wood-warblers), Thraupini (tanagers and tanager finches), Cardinalini (cardinal-grosbeaks), and Icterini (blackbirds and allies; see, e.g., Klicka, Johnson, and Lanyon 2000 ).
Although Darwin's finches have become a textbook example of adaptive radiation (Lack 1947 ; Grant 1999 ), the question of their origin has remained open. It is generally assumed that their ancestors arrived in the Galápagos Archipelago from Central or South America, where species related to them have been identified. Several species have been proposed by different authors to be the closest living relatives of Darwin's finches. They include the bananaquit (Coereba flaveola) (Harris 1972 ), the blue-black grassquit (Volatinia jacarina) (Steadman 1982 ), the blackfaced grassquit (Tiaris bicolor) (Baptista and Trail 1988 ), and the St. Lucia black finch (Melanospiza richardsoni) (Bowman 1983 ; Trail and Baptista 1989 ). The proposals have been based on morphological and behavioral similarities between Darwin's finches and the particular South/Central American candidate species. No agreement has, however, been reached as to which of these candidates is the nearest extant relative of the geospizines.
The aim of the present study was to shed light on the origin of Darwin's finches by using molecular markers, specifically the control region (cr) and the cytochrome b (cytb) gene of the mtDNA. During the study, various portions of the mtDNA genome were found to be transferred and integrated into the nuclear genome. These so-called nuclear mitochondrial, or numt, DNA sequences (Tsuzuki et al. 1983 ; Lopez et al. 1994 ; Quinn 1997 ), too, were used as phylogenetic markers. To preclude the possibility that none of the proposed candidate species, but rather a species of a different tribe, is the actual nearest relative of the Geospizinae, we sampled and tested a wide range of representatives of the various Fringillidae tribes. Resolving the phylogeny of the Fringillidae was, however, not the purpose of the study; rather, the goal was to narrow down the group of the possible candidates from which the ancestors of the Darwin's finches may have originated so that the information thus obtained could be used to make inferences about the age, place of origin, and appearance of the ancestors.
Materials and Methods
Birds
Blood samples (10–20 μl from wing veins) were taken from mist-netted specimens collected by H.T. on St. Lucia and in Ecuador (table 1 ). Each bird was identified with the help of local ornithologists, photographed, bled, marked, and released. Government permission could be obtained for this procedure only; voucher specimens could therefore not be collected. Photographs of the birds and other documentation are, however, available for inspection. Additional specimens of Tiaris canora, Coryphospingus cucullatus, and Poospiza hispaniolensis were obtained from a dealer (Zoo Züpke, Wesel-Bislich, Germany). The nine specimens of T. obscura were sampled from three localities, 50–70 km apart, near Santos Domingos, Pichincha Province, Ecuador. Blood was stored in an AS-Buffer (Qiagen Blood Kit, Qiagen). This kit was also used for DNA extraction. The Darwin's finch data set was that described in an earlier publication (Sato et al. 1999 ).
Polymerase Chain Reaction, Cloning, and Sequencing
Short polymerase chain reaction (PCR) conditions were as follows: one cycle of denaturation for 30 s at 94°C, annealing for 15 s at the annealing temperature, and extension for 7 min at 72°C, followed by 34 cycles of denaturation for 15 s at 94°C, annealing for 15 s at the annealing temperature, and extension for 1 min at 72°C, and a final extension for 7 min at 72°C. In each reaction, 2 μl of genomic DNA, 0.2 mM of each of the four deoxyribonucleotides, 0.5 μM of each of the sense and antisense primers, 2.5 U of Taq polymerase (Amersham Pharmacia Biotech), and 0.4 U Pfu DNA polymerase (Stratagene) were added to 10 μl of 5 × PCR buffer. Hot-start PCR was carried out using HotWax 3.5 mM Mg2+ beads (Invitrogen). The DNA was amplified in the GeneAmp PCR System 9700 (PE Applied Biosystems) or the PTC-200 Thermal Controller (Biozym Diagnostik, Hess. Oldendorf, Germany).
Long PCR conditions involving the Expand Long Template PCR System (Roche Diagnostics) were as follows: denaturation for 2 min at 92°C, followed by 10 cycles of 10 s at 92°C, 30 s at 64°C, and 25 min at 68°C, and then by 20 cycles of 10 s at 92°C, 30 s at 64°C, and 25 min at 68°C (20-s cycle elongation for each successive cycle), followed by 7 min at 68°C. The DNA was amplified in the GeneAmp PCR system 9700. The annealing temperature was 56°C for primer pair CB1/CB2, 58°C for M1/H1261, M1/RF12, NB77/NB74, and RR55/RR54, and 54°C for RR51/RR52. The list of PCR primers appears in table 2 .
Cloning and sequencing were carried out using methods described elsewhere (Sato et al. 1998 ). The sequencing reactions were processed by the LI-COR Long Read IR DNA sequencer (MWG Biotech) and the Automated Laser Fluorescent (A.L.F.) sequencer (Amersham Pharmacia Biotech). Each clone was sequenced in both directions, and at least two independent clones were sequenced from each individual.
Phylogenetic Reconstruction
Alignments were made for the cr using the CLUSTAL X program (Thompson et al. 1997 ) with default gap penalty settings. Phylogenetic reconstructions were undertaken both including highly variable segments of the cr (particularly the 3′-terminal 200 bp of the 1,208-bp alignment including insertions/deletions) and excluding such segments. The cytb, numt2, and numt3 sequences, which contained few indels, were aligned by eye. The aligned sequences were subjected to phylogenetic reconstructions using the maximum-parsimony (MP; Swofford 1998 ), maximum-likelihood (ML; Felsenstein 1993 ), neighbor-joining (NJ; Saitou and Nei 1987 ), and minimum-evolution (ME; Rzhetsky and Nei 1992 ) methods. The MP, ML, and ME analyses were carried out using the PAUP program (Swofford 1998 ). The different methods were chosen to minimize the influence of possible confounding factors (rate variation, compositional biases). Evidence of saturation, indicated by a decrease in the transition/transversion (Ts/Tv) ratio, facilitated the identification of the less reliable parts of the phylogenetic reconstructions. For the MP analyses, heuristic search methods of the PAUP program were applied. A starting tree was obtained by the stepwise random addition of sequences with one tree held per addition. Ten replications of the addition procedure were performed. Optimization was performed by branch-swapping using tree bisection and reconnection. Either all sites or transversions only were used in reconstructions, with equal weights given to each substitution. Trees were also constructed in which either third-position transversions (cytb) or all transversions (cr) were given a fivefold weighting over transitions. The degree of weighting was determined from the biases found in the Ts : Tv ratio for the species examined and was similar to that used by Burns (1997) . To construct the ME trees, the PAUP heuristic search was used to optimize an NJ tree based on LogDet distances. LogDet is known to reduce the influence of compositional biases (Lockhart et al. 1994 ). Ten replications using the random addition of taxa with tree bisection and reconnection as the optimization method were used to obtain the final ME trees. Prior to ML phylogenetic reconstructions, the ME LogDet tree was used to estimate the Ts/Tv ratio, the rate ratio κ, and the γ-distribution shape parameter α. The estimated rate variations and empirically observed nucleotide frequencies were used to reconstruct phylogenies on the basis of the Hasegawa-Kishino-Yano model (Hasegawa, Kishino, and Yano 1985 ) with γ-distribution by the ML method, giving two substitution types and four rate categories. Starting branch lengths were obtained using the Rogers-Swofford approximation method. A heuristic approach was based on nearest-neighbor interchange for branch swapping to optimize the ML tree. The MEGA program (Kumar, Tamura, and Nei 1993 ) was used to draw NJ trees. Both Kimura's (1980) two-parameter method and the Tamura-Nei method (Tamura and Nei 1993 ) with a γ shape parameter taken from the ME calculation were used to estimate distances.
Strategy and Procedures
Since there is general agreement that the geospizines are finch-like birds, members of the family Fringillidae, we focused our search for their nearest living relative on this family and used representatives of Tyrannidae (Capsiempis flaveola, Elaenia martinica) as an outgroup. Of the Fringillidae, we sampled the tribes (sensuSibley and Monroe 1990 ) represented in the Neotropical region (Thraupini, Emberizini, Parulini, Icterini, Cardinalini, and Carduelini) to identify the tribe to which the Darwin's finches could be assigned. We then sampled this tribe for the possible candidate genus and species. The sampling included the four species proposed by earlier investigators as being the closest relatives of Darwin's finches. Altogether, we tested 30 species (one to eight individuals per species), not counting Darwin's finches, whose phylogenetic relationships we described earlier (Sato et al. 1999 ).
We isolated DNA from blood samples, amplified it by PCR, and either sequenced the amplification products directly or cloned and then sequenced them. We focused on four DNA segments: mt cytb (922 bp, sites 102–1023; primer pair CB1/CB2), mt cr (1,082–1,157 bp, sites 125–1260; primer pair M1/H1261), numt2 (830 bp, with the initial 140 bp corresponding to sites 125–264 from the cr, followed by 690 bp of flanking nuclear DNA sequence; primer pair M1/RF12), and numt3 (740 bp, with the initial 420 bp corresponding to sites 382–801 from cytb, followed by 320 bp of flanking nuclear DNA sequence; primer pair NB77/NB74). (The known organization of the numts is diagramed in fig. 1 .) To distinguish the numt from the mtDNA sequences, we used the long PCR method (primer pair RR32/RR33 and, for reamplification in some cases, RR42/RR43) to amplify nearly the entire length (∼16 kb) of the mtDNA (fig. 2 ) and then amplified the cr and cytb segments from the product of the first amplification by nested PCR with specific primers (RR51/RR52 for one end and RR55/RR54 for the other end of the cr, and CB1/CB2 for the cytb). Comparison of the sequences established the identity of the cytb and cr sequences obtained by short PCR for all of the species tested. The four DNA segments were chosen to sample slowly (numt), intermediately (cytb), and rapidly (cr) evolving regions, and coding (cytb) and noncoding (cr, numt) regions, as well as gene (cytb) and pseudogene (numt) regions, of the genome. The sequences obtained from each of the four DNA segments were aligned and examined for evidence of base composition bias, sequence saturation, and rate variation.
Results
Evolutionary Dynamics
To answer the questions posed by the present study, we determined and evaluated the sequences of four genomic segments, two nuclear (numt2 and numt3) and two mitochondrial (cr and cytb). It can be expected that the tempo and mode of the evolution of these segments differ, and these differences must be taken into account when using the sequences in phylogenetic reconstructions. To define and characterize the differences, we examined the relative frequencies of transitions and transversions by pairwise comparisons of the individual sequences, and in one case (cytb) also codon positional effects. The comparisons were of two kinds: in one case, we used the entire set of sequences; in the other, we focused on sequences derived from T. obscura, T. canora, T. bicolor, M. richardsoni, and Loxigilla noctis. The latter group was used to enable us to compare the dynamics of all four segments, two of which (the numts) were present in only some of the species surveyed. The reason for choosing these five particular species will become apparent later. Here, we show the results of the second comparison (fig. 3 ) and refer to the first only where it provides additional information. The results of the comparisons enable us to draw three main conclusions.
First, in all segments, transition rates exceeded transversion rates, 10-fold in the case of mtDNA and two–threefold in the case of nuclear DNA. The maxima of per-site transitions in figure 3 were approximately 6% for both cr and cytb and 2.7% and 4.3% for numt2 and numt3, respectively. Similarly, the maxima of per-site transversions were close to 2% for cr and cytb and 1.3% and 1.9% for numt2 and numt3, respectively. The most reliable estimates were obtained from comparisons within the Darwin's finch group, in which the use of a large number of pairwise comparisons reduced stochastic effects and lower divergences reduced saturation effects, particularly in the case of mtDNA.
Second, substitutional saturation occurred in the two mitochondrial segments. The Ts : Tv ratio fell from 10:1 in comparisons between Darwin's finches to 5:1 in comparisons between Darwin's finches and the Tiaris/Melanospiza/Loxigilla species, and ultimately approached 1:1 in intrathraupine comparisons. There was no evidence of substitutional saturation in the two numt segments. Even in the comparison of distant thraupine taxa, the Ts : Tv ratio of 3:1 to 2:1 appears to hold.
Third, evolutionary rates differed between the segments: in comparisons involving the same taxa, the mtDNA segments diverged at two–fourfold higher rates than the nuclear segments. However, the excess depended strongly on the type of mtDNA sites used in the comparisons. For example, the third-position sites of the cytb gene and certain sites of the cr segment evolved more rapidly than the remainder of the sites. There was also some indication of a slight difference in the evolutionary rates of the two numts: the number of per-site substitutions in numt3 was approximately 1.5 times as high as that in numt2.
Substitutional saturation at the two mitochondrial segments presents a problem if these segments are to be used to resolve basal levels of phylogenetic divergence. Although a correction method can be applied based, for example, on weighting transversions (e.g., Burns 1997 ), the phylogenetic signal is inevitably weakened and lost at some sites. By contrast, the two numt segments, which show little or no evidence of saturation even when used in basal phylogenetic comparisons, can be regarded as resilient phylogenetic markers. Their resilience can be attributed to three factors: a slow evolutionary rate, absence of an extreme transitional bias, and absence of a strong nucleotide composition bias. The two mtDNA segments are affected by these three factors to different extents. The third-position cytb sites and the rapidly evolving cr sites are the most strongly affected; the effect on the remaining sites is less pronounced. Because of the different rates and saturation points, the mtDNA segments can be expected to be best suited for resolving terminal nodes of phylogenetic reconstructions, whereas the numt segments should be more reliable for resolving deeper divergences.
A comparison of GC composition did not reveal any major differences between the cytb sequences used. The third position showed a tendency toward avoidance of GT, which is a characteristic of all bird cytb sequences (Moore and DeFilippis 1997 ). The first-, second-, and third-position biases in all sequences were close to the averages of 0.057, 0.222, and 0.502, respectively. Plots of the number of transitions against the number of transversions in third positions obtained by pairwise comparisons indicated a tendency toward saturation of transitions in comparisons between the most divergent sequences within tribes. Nevertheless, NJ trees drawn using transversions only and those drawn using transversions and transitions did not differ significantly in their topologies.
Phylogenetic Analysis of Fringillidae
To determine the position of the Darwin's finches within the Fringillidae, the sequences of the four genomic segments (cytb, cr, numt2, and numt3) were analyzed by the four methods of phylogenetic reconstruction (MP, ML, ME, and NJ). Because of the absence of the numts in some of the taxa, these taxa had to be excluded from the analysis using these two segments. The cytb data set consisted of 44 nearly full length sequences, not counting identical sequences and the 41 sequences obtained earlier from 13 Darwin's finch species (Sato et al. 1999 ). To make the set more amenable to analysis by the MP and ML methods, its size was reduced to 35 by randomly choosing one sequence per species (or species group in the cases of Darwin's finches). The reduced set consisted of 28 Fringillidae, 2 Tyrannidae, and 5 Darwin's finch sequences, representing the ground, tree, Cocos, vegetarian, and warbler finches (fig. 4 ). Comparisons of NJ trees drawn for the full and reduced sets did not reveal any bias introduced by the selection of sequences. Application of the various methods of phylogenetic reconstruction to the cytb data set yielded trees in which the major clades shown in figure 4 were recovered consistently. The mutual relationships of the clades varied, however, depending on the method and the parameters (e.g., the distance measure) used. The clades themselves were relatively robust, but the statistical support for their branching order was not significant. The assortment of the species into clades only partially agrees with the tribal classification of the Fringillidae by Sibley and coworkers (Sibley and Ahlquist 1990 ; Sibley and Monroe 1990 ) or the classification systems of other investigators. The two major deviations from these systems are the consistent separation of Euphonia musica from the Thraupini and of Sturnella bellicosa from the Icterini. The interpretation of these anomalies and of our reconstructions in the context of data reported by other authors is provided in the Discussion section. Here, we focus on one of the two main goals of the study—determining the position of the geospizines within the Fringillidae.
In the various phylogenetic reconstructions, Darwin's finches consistently clustered with five Thraupini species: T. obscura, T. bicolor, T. canora, M. richardsoni, and L. noctis. These are, then, the five living species most closely related to Darwin's finches among the species we surveyed; we refer to them as the “Tiaris group.” We place their observed mutual affinity within the framework of other studies below (see Discussion), but we note here that the clustering is consistent with some similarities in morphology (Baptista and Trail 1988 ; Webster and Webster 1999 ) and vocalization (Bowman 1983 ; Baptista and Trail 1988 ; Grant, Grant, and Petren 2000 ) between the two groups. The statistical support for the geospizine-Tiaris group clade is very high (close to 100% in terms of bootstrap values). The clade is further supported by the consistency of the grouping observed in the applications of the four methods of phylogenetic reconstruction and its recovery in the analysis of all four genomic segments (see below), as well as by the existence of several shared derived substitutions and insertions/deletions in the various segments. Less consistently, the clade was joined in most of the trees by Coereba flaveola and Volatinia jacarina. These two species, together with the Tiaris group, include all four candidates for the closest relative of Darwin's finches proposed on morphological grounds (Harris 1972 ; Steadman 1982 ; Baptista and Trail 1988 ; Trail and Baptista 1989 ).
The cr data set comprised sequences of 21 Fringillidae and 13 Darwin's finch (Sato et al. 1999 ) species; the sequences of some of the more distantly related species were difficult to align with the geospizine sequences and were therefore excluded from the analysis. To expedite the analysis, the same five representative species of the main Darwin's finch groups were used as in the cytb analysis. In all cases, based on the use of the four tree-drawing programs, the Tiaris group came out as the closest of the Fringillidae clades to the Darwin's finches, followed by C. flaveola and V. jacarina. This clustering was observed regardless of whether we used the entire cr sequence or a sequence from which ∼200 bp of the most variable parts were excluded. Similarly, the inclusion of all 13 species of Darwin's finches in the analysis or the exclusion of the nonthraupine sequences from it had no effect on the sister group relationship of the Tiaris species to Darwin's finches.
The numt2 segment was found to be present in all of the Thraupini (with the exception of E. musica) and Cardinalini species tested; it was found to be absent in the Parulini, Icterini, Emberizini, and Carduelini species, as well as in the two species of Tyrannidae. Some of the positive species appear to possess more than one copy of the segment, since in at least one species (T. canora), three numt2 sequences were obtained from a single individual, and since in certain other species, relatively large genetic distances separate the numt2 “alleles.” However, Southern blot analysis using genomic DNA from T. bicolor and numt2-specific probes revealed the presence of only one strongly hybridizing band (not shown). This result could mean that the numt2 segment is tandemly duplicated in some of the species. Exhaustive exploration of PCR conditions and the use of secondary nested primers failed to amplify the segment. The species-specific clustering of the multiple sequences suggests that the duplications occurred independently in different species and therefore do not greatly influence the interpretation of the phylogenetic analysis. A total of 47 unique numt2 sequences were obtained, 16 from Darwin's finches, 28 from Thraupini, and 3 from Cardinalini. The application of the four methods of phylogenetic analysis to these sequences yielded trees in which, again, the Tiaris species formed a sister group to Darwin's finches. The bootstrap support for the geospizine-Tiaris group cluster was very high (99% in most of the trees).
Finally, the numt3 segment was also found only in Thraupini and Cardinalini, and not in the remaining taxa tested. In contrast to numt2, however, only some Thraupini (including all species of Darwin's finches) seem to possess the segment (fig. 4 ). In none of the phylogenetic trees based on the cytb, cr, or numt2 sequences do the six numt3-negative Thraupini species (Diglossa humeralis, Catamenia inornata, Sicalis flaveola, P. hispaniolensis, V. jacarina, and Rhamphocelus carbo) cluster together. Moreover, in terms of numt3 distribution among Thraupini and Cardinalini, the former appear to be paraphyletic. Possible explanations of this observation include lineage sorting during the adaptive radiation of Thraupini and Cardinalini and secondary loss of numt3 in some of the taxa. Alternatively, the numt3 distribution might be indicative of as-yet-unrecognized phylogenetic affinities within Thraupini and Cardinalini. In addition to the clustering of Darwin's finches with the Tiaris group in phylogenetic trees, as seen with the other three genomic segments, the numt3 segment also provides other evidence for the existence of this clade. At the border between the cytb-derived and the nuclear sequences of numt3, there is an 11-bp deletion which is present only in some of the Thraupini. This deletion is a shared derived character for the Darwin's finch species and the Tiaris group, with the exception of M. richardsoni. The absence of the deletion in M. richardsoni may be indicative of allelic segregation of both forms of numt3 in the Tiaris group ancestor.
The numt markers we used here have many useful phylogenetic properties but are not ideal. Problems concern the patchiness of the numt distribution and the presumed orthology of the sequences. Nevertheless, the apparent consistency between numt-based and the mtDNA-based phylogenies (at least in the reliable recent divergences) suggests that a strong phylogenetic signal is shared by these markers. This would rule out scenarios of numt evolution involving horizontal transfer or introgression between species. As discussed above, explanations for the distribution of numts can be found in recent tandem duplications, failed priming, or secondary loss. Any duplications that may have occurred are probably recent and insufficient to scramble the phylogenetic signals of speciation on the phylogeny of the numts.
Nearest Relative
To determine which of the extant species in our collection was most closely related to the Darwin's finches, we focused on the Tiaris group, which all earlier tests identified as the sister group of the geospizines. Here again, all four genomic segments were used in genealogical reconstruction by the MP and ML methods, which are highly sensitive for resolving phylogenies of close relatives (Hillis, Huelsenbeck, and Swofford 1994 ). Examples of the trees obtained are given in figures 5–7 .
In all tests, without exception, the dull-colored grassquit, T. obscura, was identified as a sister species of the Darwin's finch clade. Particularly good resolution was obtained with the cr data set. Using C. flaveola as an outgroup, 1,140 sites of this region were aligned and subjected to MP and ML analyses. Both tree-drawing methods identified T. obscura as the closest of the sample species to the geospizines. The MP method produced two most-parsimonious trees which differed only in the arrangement of the Darwin's finch species. Bootstrap support for the grouping of T. obscura with Darwin's finches ranged from 52% to 77%. The support for the same grouping for trees obtained using the other three genomic segments ranged from 44% to 63%. Weighting did not affect the terminal clade topologies of cytb and cr trees but did improve the bootstrap support for some of the nodes in the cr trees. In the alignments of cytb, cr, and numt3 sequences, seven, five, and three shared derived substitutions, respectively, were found in T. obscura and all of the species of Darwin's finches. We conclude, therefore, that T. obscura, rather than any of the other species proposed earlier, is the closest relative of Darwin's finches among the species we surveyed.
Age Estimate
Since no fossil record of the Thraupini is available which could be used to calibrate the molecular clock of the cytb segment, we used the calibrations of Shields and Wilson (1987) and Irwin, Kocher, and Wilson (1991) . The Shields and Wilson (1987) calibration is based on mtDNA restriction fragment length polymorphism in geese and assumes a sequence divergence of 2% per million years. The Irwin, Kocher, and Wilson (1991) calibration is based on the fossil record of ungulate mammals and assumes a rate of 0.5% divergence in third-position transversions per million years. Both rates have been widely used in studies of many vertebrate taxa, including thraupine birds (Burns 1997 ). The average genetic distance between T. obscura and Darwin's finches is 4.6% ± 0.6% for all cytb sites and 0.6% ± 0.4% for cytb third-position transversions. Applying the above rates to these values, we estimate 2.3 ± 0.3 Myr and 1.2 ± 0.8 Myr divergence times of T. obscura and Darwin's finches from their common ancestor, respectively. The latter value is based on a small number of substitutions which may lead to an underestimation of the divergence time. The former value is fairly close to the estimate of 2.8 Myr reached by Grant (1994) after the recalibration of the molecular clock was applied to the allozyme data of Yang and Patton (1981) . In principle, cytb-based age estimates can be tested using other loci. Unfortunately, no calibration of evolutionary rates is available for the numts, and calibrations based on the cr for geese are unsuited to the cr of finches (Marshall and Baker 1997 ).
Discussion
Phylogenetic Relationships: Comparisons with Other Data Sets
During the preparation of this manuscript, several papers were published which described cytb sequences of various Fringillidae and allied taxa. These include 10 species of Drepanidini by Feldman et al. (1997) ; 14 species of Piranga and other Thraupini by Burns (1997, 1998) ; an assortment of Fringillini and Emberizini species by Groth (1998) ; Fringilla and Carduelis by Marshall and Baker (1998, 1999) ; 25 recognized species of Icterus by Omland, Lanyon, and Fritz (1999) and other Icterini by Lanyon and Omland (1999) ; and six species of Anairetes (family Tyrannidae) by Roy, Torres-Mura, and Hertel (1999) . We used these sequences, if available in the database, together with our own to draw phylogenetic trees, which contain almost 200 species and which, because of their size, we do not show. Most recently, Klicka, Johnson, and Lanyon (2000) reported cytb and NADH dehydrogenase 2 (ND2) mtDNA sequences of 35 Fringillidae, 12 of which belong to the same genera as some of the 43 species used in the present study. Unfortunately, these sequences were not available at the time of submission of this manuscript in any of the public databases.
In the trees drawn from the different sets of data, a series of clades emerges, only some of which are supported by high bootstrap values and only some of which correspond to the groups (tribes sensuSibley and Ahlquist 1990 ) of the traditional classifications. In the tree in figure 4 , one such clade is composed of the genera Sporophila and Oryzoborus, which have long been known to be closely related. Their merger has been favored by some taxonomists (Olson 1981 ; Wetmore, Pasquier, and Olson 1984 ) but not by others (e.g., Webster and Webster 1999 ); it is supported by the molecular data in figure 4 . The Catamenia/Sicalis/Poospiza clade is part of the South American grassland group erected by Webster and Webster (1999) on osteological grounds. Similarly, the clade composed of Volatinia, Coereba, Loxigilla, Tiaris, Melanospiza, and Darwin's finches is part of Webster and Webster's (1999) seed-eaters group, united by the sharing of several skeletal features (more on this clade later). An affinity of the Icterini to the Emberizini (fig. 4 ) is also seen in the mtDNA cytb and ND2 trees of Klicka, Johnson, and Lanyon (2000) and in trees drawn from the entire collection of Fringillidae mtDNA cytb sequences (not shown). The closeness of the Cardinalini to some of the Thraupini (fig. 4 ) is also indicated by the DNA-DNA hybridization data of Bledsoe (1988) and by mtDNA cytb trees based on all of the available Fringillidae sequences (not shown).
The two major departures from the traditional classifications are the separation of Euphonia from the Thraupini and of Sturnella from Icterini. Euphonia has traditionally been classified as a tanager, largely on the basis of superficial resemblances such as bright plumage coloration and a frugivorous diet. At the same time, however, it has always appeared to be an atypical tanager because of several morphological and behavioral idiosyncrasies (e.g., Ridgway 1901 ). In the mtDNA analysis of the Thraupini by Burns (1997) , Euphonia appeared consistently outside of the group, as it also does in the tree in figure 4 . Burns (1997) , as well as Klicka, Johnson, and Lanyon (2000) , found Euphonia to associate with Chlorophonia, and in both our study and that of Klicka, Johnson, and Lanyon (2000) , Euphonia clusters with the Carduelini. Morphological similarities of Euphonia to cardueline finches have been pointed out by Dickey and Van Rossem (1938) . Burns (1997) proposes that Euphonia is either a derived cardueline group of species or a basal Fringillidae clade. Klicka, Johnson, and Lanyon (2000) suggest that it may represent “goldfinches” secondarily adapted to a fruit diet.
The genus Sturnella has traditionally been considered an Icterini. In the tree in figure 4 , however, the single Sturnella species tested assumes an outgroup position to the rest of the Emberizinae, albeit with low bootstrap support. In the study by Klicka, Johnson, and Lanyon (2000) , Sturnella associates with Icteria, a Parulini. In the tree based on all of the available Fringillidae cytb sequences and containing >40 Icterini species, Sturnella is not part of the Icterini clade (not shown). Its position in the Fringillidae system remains unresolved.
Position of Darwin's Finches Within Fringillidae
The molecular data indicate that the Fringillidae tribes as defined, for example, by Sibley and Ahlquist (1990) will have to be revised. In particular, some of the genera traditionally included in the Thraupini will have to be taken out of this tribe and reassigned to other positions. Nevertheless, there remain a large number of taxa that appear to form a monophyletic group and to which the designation Thraupini could be restricted. Darwin's finches are unambiguously part of this redefined group. Specifically, they cluster with the genera Tiaris, Melanospiza, and Loxigilla, and this clade receives very high bootstrap resampling support in all the trees analyzed (e.g., the one in fig. 4 ). Somewhat less support is provided for an extended cluster that includes Coereba, and still less support is provided for a group broadened by the inclusion of Volatinia (figs. 4–7 ). The position of Darwin's finches remains unchanged regardless of the method applied to draw the trees, the sequences used (mitochondrial or nuclear), and the taxa included in the trees. It is also indicated by trees based on all the available Fringillidae cytb sequences (not shown). Since the latter collection consists of sequences from some 200 species sampled extensively over the entire range of the Fringillidae, it can scarcely be objected that the indicated position of Darwin's finches is the result of inadequate or biased taxon representation.
The close relationship of Darwin's finches to the Tiaris group of the Thraupini has been favored for some time on morphological grounds, although only two of the four candidates proposed to be the closest living relatives of the geospizines come from this group (T. bicolor and M. richardsoni; see Bowman 1983 ; Baptista and Trail 1988 ; Trail and Baptista 1989 ). Numerous similarities between the geospizines and the Tiaris finches have been noted by different authors (reviewed in Lack 1947 ; Grant 1999 ). The most striking resemblance of Darwin's finches to Tiaris finches has been reported in recent osteological comparisons by Webster and Webster (1999) . These authors examined 39 characters in a detrended correspondence (DCA) ordination and found that Volatinia, Melanospiza, and Tiaris were within the range of variation observed in the four genera of Darwin's finches for 28 characters. In nine characters, Volatinia was outside and Tiaris, as well as Melanospiza, within this range. At least one Tiaris species was always within the range of Darwin's finches in all 39 characters. Webster and Webster (1999) conclude that on the basis of the suite of osteological characters used, Volatinia is the least likely and Tiaris the most likely ancestor of Darwin's finches. It should be noted that some taxonomists (e.g., Ridgway 1901 ) assign M. richardsoni to the genus Tiaris and that in most trees based on molecular data (both mtDNA cytb and nuclear numt2 and numt3), the genus Tiaris is polyphyletic, with M. richardsoni consistently positioning itself among the various Tiaris species (figs. 4–7 ).
The Ancestor of Darwin's Finches
Our data identify the dull-colored grassquit, T. obscura, as the nearest living relative of the Darwin's finches among the species we studied (figs. 4–7 ). Tiaris obscura was originally classified as Sporophila obscura (Paynter 1970 ), but was later moved to Tiaris by several authors (Steadman 1982 ; Clark 1986 ; Ridgely and Tudor 1989 ; Kaiser 1992 ; Bates 1997 ). Its reclassification is strongly supported by the examination of skeletal anatomy (Webster and Webster 1999 ) and by the molecular data of the present study. The species is distributed in relatively narrow strips of land in Venezuela, Colombia, western Ecuador, western and southern Peru, western Bolivia, and northwestern Argentina (Ridgely and Tudor 1989 ). Its habitat ranges from humid forest edges, through scrub, to open woodland and farmlands, and from lowlands to altitudes of 2,000 m in the Andes. It feeds on seeds, either singly or in pairs, and seasonally in small flocks. Like other Tiaris species and all Darwin's finches, it builds dome-shaped nests with a side entrance (Sporophila, by contrast, constructs cup-shaped nests). In addition to the three Tiaris species we tested, there are two others, T. olivacea and T. fuliginosa, which were not available to us. We cannot, therefore, exclude the possibility that one or both of these are even more closely related to Darwin's finches than is T. obscura.
If T. obscura is the nearest living relative of Darwin's finches, is it also their ancestor? Comparisons of genetic distances within the Tiaris group, within the geospizines, and between the two groups reveal that the distance between T. obscura and Darwin's finches based on the cytb sequences is on average ∼40 substitutions. This distance is ∼20% smaller than that between T. obscura and its nearest relatives (T. canora, T. bicolor, and M. richardsoni), which amounts, on average, to ∼54 substitutions (47 transitions and seven transversions, with the exception of T. canora, which differs from T. obscura by 45 transitions and 10 transversions). Similar distances are also obtained for pairwise comparisons of the various Tiaris/Melanospiza species with one another. Since the distances from Darwin's finches to T. obscura are only 80% of the distances within the Tiaris group itself, it is very likely that the geospizine ancestor split off from the Tiaris group early in its radiation. These deductions are further supported by the analysis of the numt3 sequences. In phylogenetic reconstructions using numt3, as in those based on cytb, T. obscura is the closest relative of the Darwin's finch group. There are, on average, ∼20 differences (15 transitions and 5 transversions) between T. obscura and any numt3 Darwin's finch sequence. This distance is again comparable with the distances between T. obscura and its closest relatives, T. bicolor and M. richardsoni, which average ∼28 substitutions, consisting of 19 transitions and 9 transversions. (Tiaris canora is, in this case, more distant from T. obscura than in the case of the cytb sequences, having ∼35 differences from the latter.)
There are then two possibilities. Either T. obscura arose early in the radiation and has to this day retained its species identity in Central and South America, while in the Galápagos Archipelago its early descendants underwent a new round of adaptive radiation, or the T. obscura lineage underwent radiation both on the mainland and on the Galápagos Islands following the separation of the two sublineages. In the former case, T. obscura would be the true ancestor of Darwin's finches; in the latter, T. obscura and Darwin's finches would share a most recent common ancestor, but T. obscura itself would not be the actual ancestor of the Darwin's finches. This reasoning is contingent on our correct identification of T. obscura as the closest living relative of the Darwin's finches.
The Origin of Darwin's Finches
Burns (1997) used the biogeographical distribution of the Thraupini, their age as estimated from mtDNA divergence, and the topology of their phylogenetic tree to argue that the group arose on the Caribbean islands and then radiated throughout Central and South America. He estimated that the genera of Thraupini began to diverge from Parulini around 26 MYA and that most of the diversity among the genera then evolved over a time span of approximately 10 Myr. This period was marked by major uplifts of the Andes, which provided the conditions for the adaptive radiation of the tanagers. The divergence within the Thraupini genera followed and may have been influenced by the closure of the Panamanian isthmus ∼3 MYA and the onset of Pleistocene glaciation in the Northern Hemisphere 2–3 MYA.
Using the cytb substitution rate of 2% per site per million years and the 0.5% rate per million years for third-position transversions, we estimate that the divergence within the genus Tiaris occurred 2–3 MYA and was followed shortly afterward by the separation of the geospizine ancestor from this group. The biogeographical distribution of most of the extant Tiaris species is centered on the Caribbean islands (Ridgely and Tudor 1989 ), so presumably the radiation of this genus occurred in this region. Since, however, the distance from Central or South America to the Galápagos Islands is ∼1,000 km shorter than that from the Caribbean islands, it seems more parsimonious to argue that the ancestors of the geospizines first dispersed from their Caribbean cradle to the former region before they undertook the journey to the Galápagos Archipelago.
The geological events that ultimately led to the closure of the seaway between the North and South American continents lasted from 13 to 1.9 MYA (Haug and Tiedemann 1998 and references therein). The closure was apparently almost complete 3.6 MYA, although the actual landbridge connecting South and North America probably did not form until ∼2.7 MYA. However, marked reorganization of ocean circulation resulting from the shallowing of the seaway had already started 4.6 MYA. Changes in the ocean currents were probably accompanied by alterations in the direction of the prevailing winds, and it may have been one of the strong new wind currents that blew the flock of ancient Tiaris species in the direction of the open waters of the Pacific Ocean and carried it to the Galápagos Archipelago. (Vincek et al. [1997] estimate the size of the founding flock to have been in excess of 30 individuals.) In the Archipelago, the finches must have found conditions not too different from those in their place of origin, and this circumstance may have facilitated the colonization of their new environment. Their occupation of the various ecological niches available on the Archipelago and their dispersion to the different islands triggered a round of adaptive radiation, a process that continues to this day. Both morphological (Grant 1999 ) and molecular (Freeland and Boag 1999a, 1999b ; Petren, Grant, and Grant 1999 ; Sato et al. 1999 ) data indicate that species boundaries have not been fixed firmly. Not only do the species hybridize (Grant 1993 ), but their mtDNA lineages have not yet been sorted out among them (Freeland and Boag 1999a, 1999b ; Sato et al. 1999 ).
Since the dramatic geological and climatic changes in the Caribbean region, as well as those in Central and South America, continued after the departure of the Darwin's finch founding flock and provided the same opportunity for the continuing adaptive radiation of Tiaris ancestors in this region as the flock found on the Galápagos Archipelago, one might expect that the stock from which the founding flock was drawn no longer exists but that both T. obscura and Darwin's finches represent its descendants. If the ancestor of Darwin's finches was a bird resembling T. obscura, possessing a blunt beak, then it evolved on the Galápagos Islands into a warbler-like finch, and one of the lineages secondarily evolved into a blunt-beaked species that gave rise to the remaining extant geospizine species. Alternatively, the common ancestor of both T. obscura and the warbler finches of the Galápagos Islands may have evolved the warbler-like morphology on the mainland. In the absence of a detailed and statistically well supported phylogeny of the genus Tiaris, we are currently unable to reconstruct their morphological evolution and distinguish between these possibilities.
Dan Graur, Reviewing Editor
Abbreviations: cr, control region; cytb, cytochrome b; ME, minimum evolution; ML, maximum likelihood; MP, maximum parsimony; mt, mitochondrial; Myr, million years; NJ, neighbor joining; numt, nuclear mitochondrial; PCR, polymerase chain reaction; Ts, transitions; Tv, transversions.
Keywords: Darwin's finches Thraupini Geospizini Tiaris adaptive radiation speciation
Address for correspondence and reprints: Akie Sato, Max-Planck-Institut für Biologie, Abteilung Immungenetik, Corrensstrasse 42, D-72076 Tübingen, Germany. E-mail: akie.sato@tuebingen.mpg.de.


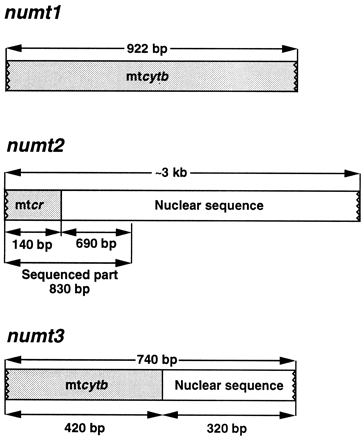
Fig. 1.—Organization of nuclear mitochondrial DNAs (numts) in Darwin's finches and related birds. numt1 is shown here for comparison; it is restricted to Darwin's finches (see Sato et al. 1999 ). mt = mitochondrial; cr = control region; cytb = cytochrome b.
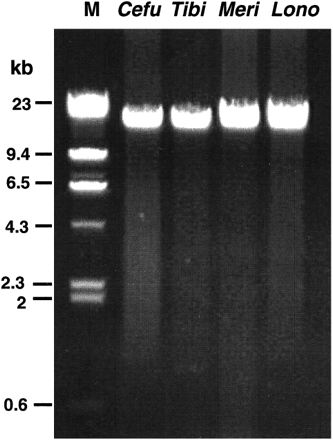
Fig. 2.—Examples of long PCR used to distinguish genuine mtDNA sequences from mtDNA fragments integrated into nuclear DNA (numts). M = size marker (λ phage; indicated sizes are in kb); Cefu = Certhidea fusca; Tibi = Tiaris bicolor; Meri = Melanospiza richardsoni; Lono = Loxigilla noctis.
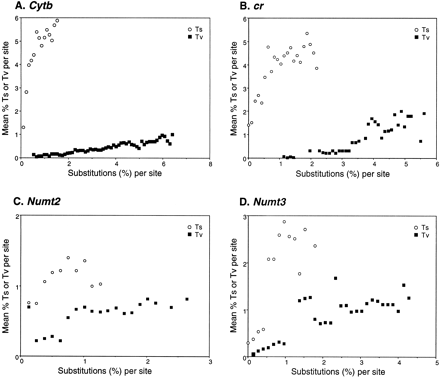
Fig. 3.—Plots of transitions (Ts) or transversions (Tv) in pairwise comparisons at each site of (A) cytb, (B) cr, (C) numt2, and (D) numt3 sequences against the corresponding total percentages of Tv or Ts divergence. The taxa compared were Darwin's finches (except for the Cocos finch numt3), Tiaris obscura, Tiaris canora, Tiaris bicolor, Loxigilla noctalis, and Melanospiza richardsoni. The x axis shows Ts or Tv substitutions per site measured as the uncorrected percentage of sequence divergence. The ordinate axis shows the mean percentage (over all pairwise comparisons) of Tv (filled squares) or Ts (open circles) per site corresponding to a fixed number of Ts or Tv, respectively, found in the same pairwise comparisons
![Fig. 4.—Strict consensus of three best maximum-parsimony trees of the cytb gene sequences. The tree was made using heuristic approaches and indicates topological relationships only. There were 322 parsimony-informative characters (921 sites in total and no gaps). Each of the three best trees found had 1,723 steps and differed only in the position of Loxigilla noctis and Tiaris canora. A number on a node indicates the percentage recovery of that node in 500 bootstrap replications; values below 10% are not shown. Analysis of substitutional saturation indicates that deep divergences may be misconstructed (the Ts-to-Tv ratio falls from 10:1 within the Darwin's finch [DF] group to close to 1:1 in comparisons to nonthraupine finches). There is little evidence of substantial saturation up to and including comparisons between DF and Tiaris, Loxigilla, and Melanospiza species, in which transition : transversion ratios are in the range of 5:1 to 10:1. Plus signs and minus signs following species names indicate the presence or absence of numt2 (first symbol) and numt3 (second symbol). In this figure and in the figures that follow, the sequences are identified by their GenBank accession codes](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/mbe/18/3/10.1093_oxfordjournals.molbev.a003806/2/m_mbev-18-02-20-f04.gif?Expires=1716315062&Signature=oD1fK1o571WTaZKfsHj0FpATqYSWCKEJ3LbrNZ3gz8b5PAS7mklQRxHoqOVjTomMb5CpbRMYK1gYJ9BVSa7ptmerhVwMYndaau1M5QFQI0YIgs65i~OrfZF4NOp11jDwPvCmTC-N9O1ICVQvOpPfTvvDfIuWEF390RCbAkV~zC4MQWB0Y4sg~S2dX8ffbIf4r-n8NBLz2XZgPjffFpZKtUUwxtZCpunBn~ZBXHBYinTnnzwdlpW9ypmcZBIk9LMumeY~MBRFS~VNFaALUxtQJFHe6w7EpXAGK6kEVapk4S40bbODXsrRsoEAJ9ZPjVuL6qACt94cCNgXROofI7xLBg__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Fig. 4.—Strict consensus of three best maximum-parsimony trees of the cytb gene sequences. The tree was made using heuristic approaches and indicates topological relationships only. There were 322 parsimony-informative characters (921 sites in total and no gaps). Each of the three best trees found had 1,723 steps and differed only in the position of Loxigilla noctis and Tiaris canora. A number on a node indicates the percentage recovery of that node in 500 bootstrap replications; values below 10% are not shown. Analysis of substitutional saturation indicates that deep divergences may be misconstructed (the Ts-to-Tv ratio falls from 10:1 within the Darwin's finch [DF] group to close to 1:1 in comparisons to nonthraupine finches). There is little evidence of substantial saturation up to and including comparisons between DF and Tiaris, Loxigilla, and Melanospiza species, in which transition : transversion ratios are in the range of 5:1 to 10:1. Plus signs and minus signs following species names indicate the presence or absence of numt2 (first symbol) and numt3 (second symbol). In this figure and in the figures that follow, the sequences are identified by their GenBank accession codes
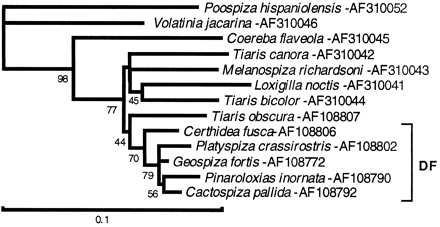
Fig. 5.—Maximum-likelihood tree obtained using cytb sequences of those species found to be most closely related to the Darwin's finch (DF) group by various tree-drawing methods applied to the four DNA sequence data sets. Poospiza hispaniolensis is included as an outgroup. The estimated transition : transversion ratio is 4.747 (κ = 9.390), and the α parameter is 0.202. Numbers below nodes show bootstrap recovery in 500 replications. During bootstrapping, trees with approximate likelihoods of 5% or farther away from the target score were rejected without additional iteration. The log likelihood of the tree shown is −3,129.739. The scale bar indicates the number of substitutions per site
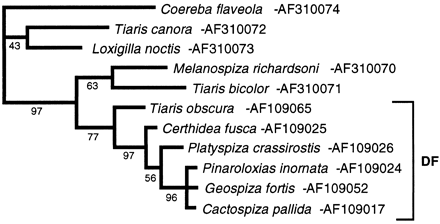
Fig. 6.—Strict consensus of two best maximum-parsimony trees of the control regions (crs) of selected Darwin's finches (DF) and their closest relatives. Each of the two best trees had 404 steps and differed only in the placement of the Cocos finch within the Darwin's finch group. Coereba flaveola is always more distant than the Tiaris group from the Darwin's finches and is used here to root the tree. Transversions are weighted five times transitions for all cr sites. Removal of the weighting or an alternative selection of Darwin's finch representatives does not alter the topology of the tree. A number below a node indicates the percentage recovery of that node in 500 bootstrap replications

Fig. 7.—Maximum-likelihood reconstruction of numt3 phylogeny. The initial tree, branch length, and rate variations were obtained from a minimum-evolution tree. The DNA segment has an estimated transition : transversion ratio of 2.839 (κ = 5.687) and a γ shape parameter of 0.563. Numbers below nodes show bootstrap recovery in 500 replications. During bootstrapping, trees with approximate likelihoods of 5% or farther away from the target score were rejected without additional iteration. The tree obtained has a log likelihood of −2,120.820. The scale bar indicates the number of substitutions per site. DF = Darwin's finches
We are grateful to Dr. Laura Arcos-Teran and Dr. Alberto Padilla, Departmento de Biologia, Pontifica Universitad Catolica del Ecuador, Quito, Ecuador, and to the Instituto Ecuatoriano Forestal y de Areas Naturales y Vida Silvestre (INEFAN) for their help and support in obtaining all the necessary permits. We thank the Galápagos and Costa Rica National Parks Services and the Charles Darwin Research Station (Galápagos); the staff on Cocos Island and the Smithsonian Tropical Research Institute for administrative and logistical support; the National Science Foundation for financial support to B.R.G. and P.R.G.; Drs. Brian James and Donald Anthony, The Forestry Department, Ministry of Agriculture, Lands, Fisheries and Forestry, Castries, St. Lucia, West Indies, and Juan Carlos Matheus for assistance in capturing and identification of birds; Ms. Sabine Rosner, Ms. Gudrun Labib, and Ms. Ljubica Sanader for technical assistance; and Ms. Jane Kraushaar for editorial assistance. We dedicate this paper to Luis F. Baptista, recently deceased. He was an outstanding avian ethologist whose behavioral studies of finches led to the investigation reported in this contribution.
literature cited
Baker, A. J., and H. D. Marshall.
Bates, J. M.
Bledsoe, A. H.
Bowman, R. I.
Burns, K. J.
———.
Clark, G. A. J.
Dickey, D. R., and A. J. Van Rossem.
Feldman, R. A., L. A. Freed, J. G. Groth, and R. L. Cann.
Felsenstein, J.
Freeland, J. R., and P. T. Boag. 1999a. Phylogenetics of Darwin's finches: paraphyly in the tree finches, and two divergent lineages in the warbler finch. Auk 116:577–587.
———. 1999b. The mitochondrial and nuclear genetic homogeneity of the phenotypically diverse Darwin's ground finches. Evolution 53:1553–1563.
Gould, J.
Grant, P. R.
———.
———.
Grant, P. R., B. R. Grant, and K. Petren.
Groth, J. G.
Hasegawa, M., H. Kishino, and T.-A. Yano.
Haug, G. H., and R. Tiedemann.
Hillis, D. M., J. P. Huelsenbeck, and D. L. Swofford.
Irwin, D. M., T. D. Kocher, and A. C. Wilson.
Jo, N.
Kaiser, K. A.
Kimura, M.
Klicka, J., K. P. Johnson, and S. M. Lanyon.
Kumar, S., K. Tamura, and M. Nei.
Lanyon, S. M., and K. E. Omland.
Lockhart, P. J., M. A. Steel, M. D. Hendy, and D. Penny.
Lopez, J. V., N. Yuhki, R. Masuda, W. Modi, and S. J. O'Brien.
Lowe, P. R.
Marshall, H. D., and A. J. Baker.
———.
———.
Moore, W. S., and V. R. DeFilippis.
Olson, S. L.
Omland, K. E., S. M. Lanyon, and S. J. Fritz.
Paynter, R. A. Jr.
Petren, K., B. R. Grant, and P. R. Grant.
Quinn, T. W.
Ridgely, R. S., and G. Tudor.
Rothschild, W., and E. Hartert.
———.
Roy, M. S., J. C. Torres-Mura, and F. Hertel.
Rzhetsky, A., and M. Nei.
Saitou, N., and M. Nei.
Sato, A., F. Figueroa, C. O'hUigin, N. Steck, and J. Klein.
Sato, A., C. O'hUigin, F. Figueroa, P. R. Grant, B. R. Grant, H. Tichy, and J. Klein.
Shields, G. F., and A. C. Wilson.
Sibley, C. G., and J. E. Ahlquist.
Sibley, C. G., and B. L. Monroe.
Snodgrass, R. E., and E. Heller.
Steadman, D.
Sushkin, P. P.
———.
Swofford, D. L.
Tamura, K., and M. Nei.
Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin, and D. G. Higgins.
Trail, P. W., and L. F. Baptista.
Tsuzuki, T., H. Nomiyama, C. Setoyama, S. Maeda, and K. Shimada.
Vincek, V., C. O'hUigin, Y. Satta, N. Takahata, P. T. Boag, P. R. Grant, B. R. Grant, and J. Klein.
Webster, J. D., and J. R. Webster.
Wetmore, A., R. F. Pasquier, and S. L. Olson.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}