Drug-Resistant Fungi: An Emerging Challenge Threatening Our Limited Antifungal Armamentarium

 , , , and
, , , and
Abstract
:1. Introduction
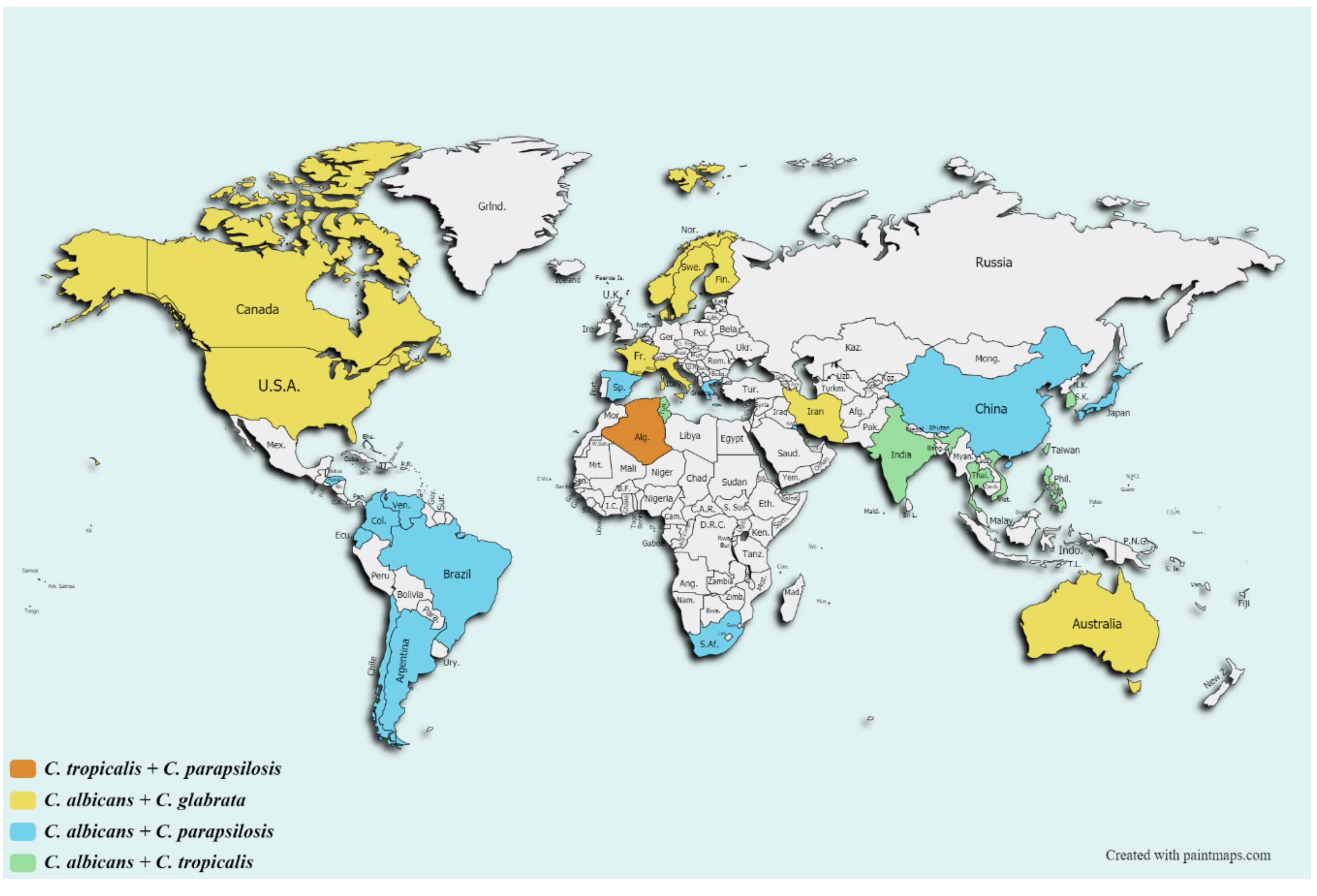
2. Epidemiology and Burden of Antifungal Resistance in Clinically Important Fungi
3. Mechanisms of Antifungal Resistance
3.1. Antifungal Tolerance
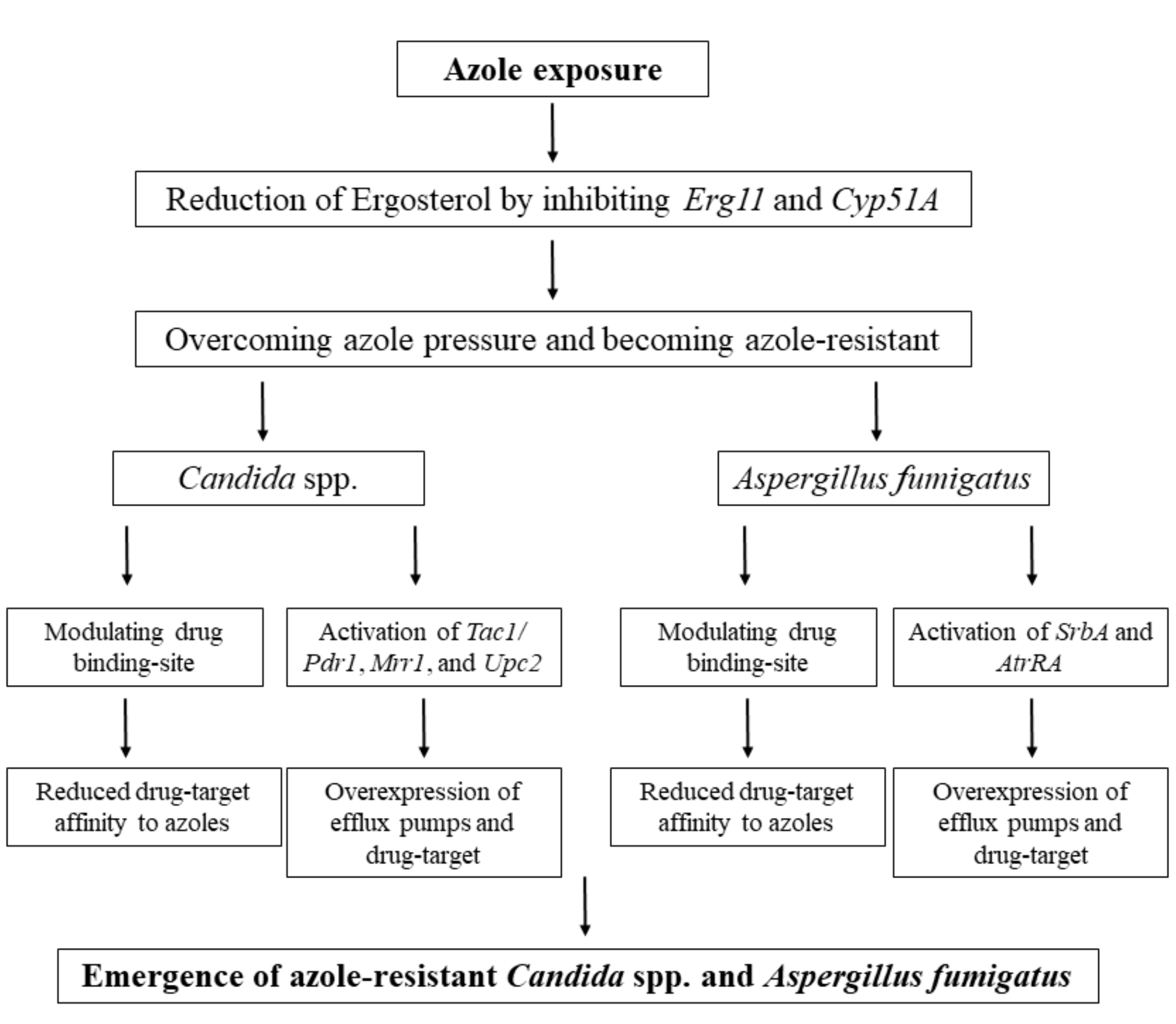
3.2. Azole Resistance
3.2.1. Azole Resistance in the Candida Genus
3.2.2. Azole Resistance in A. fumigatus
3.3. Echinocandin Resistance
3.4. Antifungal Resistance in Cryptococcus and Pneumocystis
4. Antifungal Susceptibility Testing: Current Paradigm, Challenges, and Solutions
5. Emerging Molecular Approaches to Diagnose Resistance, Current State and Challenges
6. Therapeutic Drug Monitoring and Drug Dose Optimization
7. Optimizing Therapy by Species Identification
8. Therapeutic Challenges and how to Overcome Them
9. Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fisher, M.C.; Gurr, S.J.; Cuomo, C.A.; Blehert, D.S.; Jin, H.; Stukenbrock, E.H.; Stajich, J.E.; Kahmann, R.; Boone, C.; Denning, D.W.; et al. Threats posed by the fungal kingdom to humans, wildlife, and agriculture. mBio 2020, 11, e00449-20. [Google Scholar] [CrossRef] [PubMed]
- Rolling, T.; Hohl, T.M.; Zhai, B. Minority report: The intestinal mycobiota in systemic infections. Curr. Opin. Microbiol. 2020, 56, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhai, B.; Ola, M.; Rolling, T.; Tosini, N.L.; Joshowitz, S.; Littmann, E.R.; Amoretti, L.A.; Fontana, E.; Wright, R.J.; Miranda, E.; et al. High-resolution mycobiota analysis reveals dynamic intestinal translocation preceding invasive candidiasis. Nat. Med. 2020, 26, 59–64. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2013, 4, 165rv13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aykut, B.B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; Shadaloey, S.A.; Wu, D.; Preiss, P.; Verma, N.; et al. The fungal mycobiome promotes pancreatic oncogenesis via activation of MBL. Nature 2019, 574, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.M.; Inamine, T.; Hochrath, K.; Chen, P.; Wang, L.; Llorente, C.; Bluemel, S.; Hartmann, P.; Xu, J.; Koyama, Y.; et al. Intestinal fungi contribute to development of alcoholic liver disease. J. Clin. Investig. 2017, 127, 2829–2841. [Google Scholar] [CrossRef] [Green Version]
- Berman, J.; Krysan, D.J. Drug resistance and tolerance in fungi. Nat. Rev. Microbiol. 2020, 18, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 62, e1–e50. [Google Scholar] [CrossRef]
- Patterson, T.F.; Thompson, G.R., 3rd; Denning, D.W.; Fishman, J.A.; Hadley, S.; Herbrecht, R.; Kontoyiannis, D.P.; Marr, K.A.; Morrison, V.A.; Nguyen, M.H.; et al. Practice Guidelines for the Diagnosis and Management of Aspergillosis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 63, e1–e60. [Google Scholar] [CrossRef]
- Cao, C.; Wang, Y.; Husain, S.; Soteropoulos, P.; Xue, C. A mechanosensitive channel governs lipid flippase-mediated echinocandin resistance in Cryptococcus neoformans. mBio 2019, 10, e01952-19. [Google Scholar] [CrossRef] [Green Version]
- Grossman, N.T.; Casadevall, A. Physiological differences in Cryptococcus neoformans strains in vitro versus in vivo and their effects on antifungal susceptibility. Antimicrob. Agents Chemother. 2017, 61, e02108-16. [Google Scholar] [CrossRef] [Green Version]
- Salzer, H.J.F.; Schäfer, G.; Hoenigl, M.; Günther, G.; Hoffmann, C.; Kalsdorf, B.; Alanio, A.; Lange, C. Clinical, diagnostic, and treatment disparities between HIV-infected and non-HIV-infected immunocompromised patients with Pneumocystis jirovecii pneumonia. Respiration 2018, 96, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Salzer, H.J.F.; Heyckendorf, J.; Kalsdorf, B.; Herzmann, C.; Hoffmann, C.; Lange, C. Chronic cough and severe weight loss in a 55-year-old previously healthy man. Clin. Infect. Dis. 2017, 65, 349–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa-de-Oliveira, S.; Rodrigues, A.G. Candida albicans antifungal resistance and tolerance in bloodstream infections: The triad yeast-host-antifungal. Microorganisms 2020, 8, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowen, L.E.; Sanglard, D.; Howard, S.J.; Rogers, P.D.; Perlin, D.S. Mechanisms of antifungal drug resistance. Cold Spring Harb. Perspect. Med. 2014, 5, a019752. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Messina, F.; Marin, E.; Arechavala, A.; Depardo, R.; Walker, L.; Negroni, R.; Santiso, G. Antifungal resistance in clinical isolates of Aspergillus spp.: When local epidemiology breaks the norm. J. Fungi 2019, 5, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamoth, F.; Lockhart, S.R.; Berkow, E.L.; Calandra, T. Changes in the epidemiological landscape of invasive candidiasis. J. Antimicrob. Chemother. 2018, 73, i4–i13. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A.; Diekema, D.J.; Turnidge, J.D.; Castanheira, M.M.; Jones, R.N. Twenty years of the SENTRY antifungal surveillance program: Results for Candida species from 1997–2016. Open Forum Infect. Dis. 2019, 6 (Suppl. 1), S79–S94. [Google Scholar] [CrossRef] [Green Version]
- Latgé, J.P.; Chamilos, G. Aspergillus fumigatus and aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33, e00140-18. [Google Scholar] [CrossRef]
- Verweij, P.E.; Zhang, J.; Debets, A.J.M.; Meis, J.F.; van de Veerdonk, F.L.; Schoustra, S.E.; Zwaan, B.J.; Melchers, W.J.G. In-host adaptation and acquired triazole resistance in Aspergillus fumigatus: A dilemma for clinical management. Lancet Infect. Dis. 2016, 16, e251–e260. [Google Scholar] [CrossRef]
- Chow, N.A.; Muñoz, J.F.; Gade, L.; Berkow, E.L.; Li, X.; Welsh, R.M.; Forsberg, K.; Lockhart, S.R.; Adam, R.; Alanio, A.; et al. Tracing the evolutionary history and global expansion of Candida auris using population genomic analyses. mBio 2020, 11, e03364-19. [Google Scholar] [CrossRef] [PubMed]
- Healey, K.R.; Perlin, D.S. Fungal resistance to echinocandins and the MDR phenomenon in Candida glabrata. J. Fungi 2018, 4, 105. [Google Scholar]
- Huang, L.; Crothers, K.; Atzori, C.; Benfield, T.; Miller, R.; Rabodonirina, M.; Helweg-Larsen, J. Dihydropteroate synthase gene mutations in Pneumocystis and sulfa resistance. Emerg. Infect. Dis. 2004, 10, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Linder, K.A.; Gandhi, T.N.; Miceli, M.H. Treatment failure of isavuconazole in a patient with cryptococcosis. Mycopathologia 2019, 184, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Lamb, D.C.; Corran, A.; Baldwin, B.C.; Kwon-Chung, J.; Kelly, S.L. Resistant P45051A1 activity in azole antifungal tolerant Cryptococcus neoformans from AIDS patients. FEBS Lett. 1995, 368, 326–330. [Google Scholar] [CrossRef] [Green Version]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquerdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect. Dis. 2017, 17, e383–e392. [Google Scholar] [CrossRef]
- Arastehfar, A.; Daneshnia, F.; Salehi, M.; Yaşar, M.; Hoşbul, T.T.; Ilkit, M.; Pan, W.W.; Hagen, F.; Arslan, N.; Türk-Dağı, H.; et al. Low level of antifungal resistance of Candida glabrata blood isolates in Turkey: Fluconazole minimum inhibitory concentration and FKS mutations can predict therapeutic failure. Mycoses 2020, 63, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Prideaux, B.; Nagasaki, Y.; Lee, M.H.; Chen, P.Y.; Blanc, L.; Ho, H.; Clancy, C.J.; Nguyen, M.H.; Dartois, V.; et al. Unraveling drug penetration of echinocandin antifungals at the site of infection in an intra-abdominal abscess model. Antimicrob. Agents Chemother. 2017, 61, e01009-17. [Google Scholar] [CrossRef] [Green Version]
- Müller, F.M.C.; Seidler, M.; Beauvais, A. Aspergillus fumigatus biofilms in the clinical setting. Med. Mycol. 2011, 49, S96–S100. [Google Scholar] [CrossRef] [Green Version]
- Beauvais, A.; Latgé, J.P. Aspergillus biofilm in vitro and in vivo. Microbiol. Spectr. 2015, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Perry, A.M.; Hernday, A.D.; Nobile, C.J. Unraveling how Candida albicans forms sexual biofilms. J. Fungi 2020, 6, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalheiro, M.; Teixeira, M.C. Candida biofilms: Threats, challenges, and promising strategies. Front. Microbiol. 2018, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, D.; Henriques, M.; Silva, S. Portrait of Candida species biofilm regulatory network genes. Trends Microbiol. 2017, 25, 62–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinea, J. Global trends in the distribution of Candida species causing candidemia. Clin. Microbiol. Infect. 2014, 20 (Suppl. 6), 5–10. [Google Scholar] [CrossRef] [Green Version]
- Jeffery-Smith, A.; Taori, S.K.; Schelenz, S.; Jeffery, K.; Johnson, E.M.; Borman, A.; Candida auris Incident Management Team; Manuel, R.; Brown, C.S. Candida auris: A review of the literature. Clin. Microbiol. Rev. 2017, 31, e00029-17. [Google Scholar] [CrossRef] [Green Version]
- Mathur, P.; Hasan, F.; Singh, P.K.; Malhotra, R.; Walia, K.; Chowdhary, A. Five-year profile of candidaemia at an Indian trauma centre: High rates of Candida auris blood stream infections. Mycoses 2018, 61, 674–680. [Google Scholar] [CrossRef] [Green Version]
- Govender, N.P.; Magobo, R.E.; Mpembe, R.; Mhlanga, M.; Matlapeng, P.; Corcoran, C.; Govind, C.; Lowman, W.; Senekal, M.; Thomas, J. Candida auris in South Africa, 2012–2016. Emerg. Infect. Dis. 2018, 24, 2036–2040. [Google Scholar] [CrossRef] [Green Version]
- van Schalkwyk, E.; Mpembe, R.S.; Thomas, J.; Shuping, L.; Ismail, H.; Lowman, W.; Karstaedt, A.S.; Chibabhai, V.; Wadula, J.; Avenant, T.; et al. Epidemiologic shift in candidemia driven by Candida auris, South Africa, 2016–2017. Emerg. Infect. Dis. 2019, 25, 1698–1707. [Google Scholar] [CrossRef] [Green Version]
- Eyre, D.W.; Sheppard, A.E.; Madder, H.; Moir, I.; Moroney, R.; Quan, T.P.; Griffiths, D.L.; Morgan, M.; Newnham, R.; Sunderland, M.; et al. A Candida auris outbreak and its control in an intensive care setting. N. Engl. J. Med. 2018, 379, 1322–1331. [Google Scholar] [CrossRef]
- Theodoropoulos, N.M.; Bolstorff, B.; Bozorgzadeh, A.; Brandeburg, C.; Cumming, M.; Daly, J.S.; Ellison, R.T., 3rd; Forsberg, K.; Gade, L.; Gibson, L.; et al. Candida auris outbreak involving liver transplant recipients in a surgical intensive care unit. Am. J. Transplant. 2020, 20, 3673–3679. [Google Scholar] [CrossRef]
- Armstrong, P.A.; Rivera, S.M.; Escandon, P.; Caceres, D.H.; Chow, N.; Stuckey, M.J.; Díaz, J.; Gomez, A.; Vélez, N.; Espinosa-Bode, A.; et al. Hospital-associated multicenter outbreak of emerging fungus Candida auris, Colombia, 2016. Emerg. Infect. Dis. 2019, 25, 1339–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, M.V.; Johnson, C.J.; Kernien, J.F.; Patel, T.D.; Lam, B.C.; Cheong, J.Z.A.; Meudt, J.J.; Shanmuganayagam, D.; Kalan, L.R.; Nett, J.E. Candida auris forms high-burden biofilms in skin niche conditions and on porcine skin. mSphere 2020, 5, e00910–e00919. [Google Scholar] [CrossRef] [Green Version]
- Lockhart, S.R.; Iqbal, N.; Cleveland, A.A.; Farley, M.M.; Harrison, L.H.; Bolden, C.B.; Baughman, W.; Stein, B.; Hollick, R.; Park, B.J.; et al. Species identification and antifungal susceptibility testing of Candida bloodstream isolates from population-based surveillance studies in two U.S. cities from 2008 to 2011. J. Clin. Microbiol. 2012, 50, 3435–3442. [Google Scholar] [CrossRef] [Green Version]
- Chapman, B.; Slavin, M.; Marriott, D.; Halliday, C.; Kidd, S.; Arthur, I.; Bak, N.; Heath, C.H.; Kennedy, K.; Morrissey, C.O.; et al. Changing epidemiology of candidaemia in Australia. J. Antimicrob. Chemother. 2017, 72, 1103–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trouvé, C.; Blot, S.; Hayette, M.P.; Jonckheere, S.; Patteet, S.; Rodriguez-Villalobos, H.; Symoens, F.; van Wijngaerden, E.; Lagrou, K. Epidemiology and reporting of candidaemia in Belgium: A multi-centre study. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Astvad, K.M.T.; Johansen, H.K.; Røder, B.L.; Rosenvinge, F.S.; Knudsen, J.D.D.; Lemming, L.; Schønheyder, H.C.; Hare, R.K.; Kristensen, L.; Nielsen, L.; et al. Update from a 12-year nationwide fungaemia surveillance: Increasing intrinsic and acquired resistance causes concern. J. Clin. Microbiol. 2018, 56, e01564-17. [Google Scholar]
- Arastehfar, A.; Daneshnia, F.; Hilmioğlu-Polat, S.; Fang, W.; Yaşar, M.; Polat, F.; Metin, D.Y.; Rigole, P.; Coenye, T.; Ilkit, M.; et al. First report of candidemia clonal outbreak caused by emerging fluconazole-resistant Candida parapsilosis isolates harboring Y132F and/or Y132F+K143R in Turkey. Antimicrob. Agents Chemother. 2020, 64, e01001–e01020. [Google Scholar] [CrossRef]
- Govender, N.P.; Patel, J.; Magobo, R.E.; Naicker, S.; Wadula, J.; Whitelaw, A.; Coovadia, Y.; Kularatne, R.; Govind, C.; Lockhart, S.R.; et al. Emergence of azole-resistant Candida parapsilosis causing bloodstream infection: Results from laboratory-based sentinel surveillance in South Africa. J. Antimicrob. Chemother. 2016, 71, 1994–2004. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.Y.; Chuang, Y.C.; Wu, U.I.; Sun, H.Y.; Wang, J.T.; Sheng, W.H.; Lo, H.J.; Wang, H.Y.; Chen, Y.C.; Chang, S.C. Clonality of fluconazole-nonsusceptible Candida tropicalis in bloodstream infections, Taiwan, 2011–2017. Emerg. Infect. Dis. 2019, 25, 1660–1667. [Google Scholar] [CrossRef] [Green Version]
- Lestrade, P.P.A.; Buil, J.B.; van der Beek, M.T.; Kuijper, E.J.; van Dijk, K.; Kampinga, G.A.; Rijnders, B.J.A.; Vonk, A.G.; de Greeff, S.C.; Schoffelen, A.F.; et al. Paradoxal trends in azole-resistant Aspergillus fumigatus in a National Multicenter Surveillance Program, The Netherlands, 2013–2018. Emerg. Infect. Dis. 2020, 26, 1447–1455. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Chowdhary, A.; Cuenca-Estrella, M.; Fothergill, A.; Fuller, J.; Hagen, F.; Govender, N.; Guarro, J.; Johnson, E.; Lass-Flörl, C.; et al. Cryptococcus neoformans-Cryptococcus gattii species complex: An international study of wild-type susceptibility endpoint and epidemiological cutoff values for amphotericin B and flucytosine. Antimicrob. Agents Chemother. 2012, 56, 3107–3113. [Google Scholar] [CrossRef] [Green Version]
- Vena, A.; Muñoz, P.; Guinea, J.; Escribano, P.; Peláez, T.; Valerio, M.; Bonache, F.; Gago, S.; Álvarez-Uría, A.; Bouza, E. Fluconazole resistance is not a predictor of poor outcome in patients with cryptococcosis. Mycoses 2019, 62, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Chang, T.Y.; Liu, J.W.; Chen, F.J.; Chien, C.C.; Tang, Y.F.; Lu, C.H. Correlation of anti-fungal susceptibility with clinical outcomes in patients with cryptococcal meningitis. BMC Infect. Dis. 2012, 12, 361. [Google Scholar] [CrossRef] [PubMed]
- Bongomin, F.; Oladele, R.O.; Gago, S.; Moore, C.B.; Richardson, M.D. A systematic review of fluconazole resistance in clinical isolates of Cryptococcus species. Mycoses 2018, 61, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Selb, R.; Fuchs, V.; Graf, B.; Hamprecht, A.; Hogardt, M.; Sedlacek, L.; Schwarz, R.; Idelevich, E.A.; Becker, S.L.; Held, J.; et al. Molecular typing and in vitro resistance of Cryptococcus neoformans clinical isolates obtained in Germany between 2011 and 2017. Int. J. Med. Microbiol. 2019, 309, 151336. [Google Scholar] [CrossRef] [PubMed]
- Hagen, F.; Jensen, R.H.; Meis, J.F.; Arendrup, M.C. Molecular epidemiology and in vitro antifungal susceptibility testing of 108 clinical Cryptococcus neoformans sensu lato and Cryptococcus gattii sensu lato isolates from Denmark. Mycoses 2016, 59, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Esteves, F.; de Sousa, B.; Calderón, E.J.; Huang, L.; Badura, R.; Maltez, F.; Bassat, Q.; de Armas, Y.; Antunes, F.; Matos, O. Multicentre study highlighting clinical relevance of new high-throughput methodologies in molecular epidemiology of Pneumocystis jirovecii pneumonia. Clin. Microbiol. Infect. 2016, 22, 566.e9–566.e19. [Google Scholar]
- Ozkoc, S.; Erguden, C.; Bayram-Delibas, S. Absence of dihydropteroate synthase gene mutations in Pneumocystis jirovecii strains isolated from Aegean region of Turkey. Parasitol. Res. 2018, 117, 3103–3108. [Google Scholar] [CrossRef]
- Ponce, C.A.; Chabé, M.; George, C.; Cárdenas, A.; Durán, L.; Guerrero, J.; Bustamante, R.; Matos, O.; Huang, L.; Miller, R.F.; et al. High prevalence of Pneumocystis jirovecii dihydropteroate synthase gene mutations in patients with a first episode of pneumocystis pneumonia in Santiago, Chile, and clinical response to trimethoprim-sulfamethoxazole therapy. Antimicrob. Agents Chemother. 2017, 61, e01290-16. [Google Scholar] [CrossRef] [Green Version]
- Suárez, I.; Roderus, L.; van Gumpel, E.; Jung, N.; Lehmann, C.; Fätkenheuer, G.; Hartmann, P.; Plum, G.; Rybniker, J. Low prevalence of DHFR and DHPS mutations in Pneumocystis jirovecii obtained from a German cohort. Infection 2017, 45, 341–347. [Google Scholar] [CrossRef]
- Tyagi, A.K.; Mirdha, B.R.; Luthra, K.; Guleria, R.; Mohan, A.; Singh, U.B.; Samantaray, J.C.; Dar, L.; Iyer, V.K.; Chaudhry, R. Dihydropteroate synthase (DHPS) gene mutation study in HIV-infected Indian patients with Pneumocystis jirovecii pneumonia. J. Infect. Dev. Ctries. 2010, 4, 761–766. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, A.; Sood, P.; Rudramurthy, S.M.; Chen, S.; Kaur, H.; Capoor, M.; Chhina, D.; Rao, R.; Eshwara, V.K.; Xess, I.; et al. Incidence, characteristics and outcome of ICU-acquired candidemia in India. Intensive Care Med. 2015, 41, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Hii, I.M.; Chang, H.L.; Lin, L.C.; Lee, Y.L.; Liu, Y.M.; Liu, C.E.; Chen, C.H.; Cheng, Y.R.; Chang, C.Y. Changing epidemiology of candidemia in a medical center in middle Taiwan. J. Microbiol. Immunol. Infect. 2015, 48, 306–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.F.; Liu, W.L.; Hsieh, M.H.; Hii, I.M.; Lee, Y.L.; Lin, Y.T.; Ho, M.W.; Liu, C.E.; Chen, Y.H.; Wang, F.D. Epidemiology and antifungal susceptibility of candidemia isolates of non-albicans Candida species from cancer patients. Emerg. Microbes Infect. 2017, 6, e87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, E.J.; Shin, J.H.; Choi, M.J.; Lee, W.G.; Park, Y.J.; Uh, Y.; Kim, S.Y.; Lee, M.K.; Kim, S.H.; Shin, M.G.; et al. Antifungal susceptibilities of bloodstream isolates of Candida species from nine hospitals in Korea: Application of new antifungal breakpoints and relationship to antifungal usage. PLoS ONE 2015, 10, e0118770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.H.; Jung, D.S.; Lee, J.Y.; Kim, H.A.; Ryu, S.Y.; Jung, S.I.; Joo, E.J.; Cheon, S.; Kim, Y.S.; Kim, S.W.; et al. Changing epidemiology of non-albicans candidemia in Korea. J. Infect. Chemother. 2019, 25, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Yang, Y.; Kang, Y.; Zang, B.; Cui, W.; Qin, B.; Qin, Y.; Fang, Q.; Qin, T.; Jiang, D.; et al. Invasive candidiasis in intensive care units in China: A multicentre prospective observational study. J. Antimicrob. Chemother. 2013, 68, 1660–1668. [Google Scholar] [CrossRef] [Green Version]
- Kakeya, H.; Shibata, W.; Yamada, K.; Kaneko, Y. National trends in the Japanese distribution of major Candida species causing candidemia during 2003–2017: A report by the epidemiological investigation committee for human mycoses in Japan. Open Forum Infect. Dis. 2019, 6 (Suppl. 2), S142. [Google Scholar] [CrossRef]
- Tan, T.Y.; Hsu, L.Y.; Alejandria, M.M.; Chaiwarith, R.; Chinniah, T.; Chayakulkeeree, M.; Choudhury, S.; Chen, Y.H.; Shin, J.H.; Kiratisin, P.; et al. Antifungal susceptibility of invasive Candida bloodstream isolates from the Asia-Pacific region. Med. Mycol. 2016, 54, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Khan, Z.; Ahmad, S.; Al-Sweih, N.; Mokaddas, E.; Al-Banwan, K.; Alfouzan, W.; Al-Obaid, I.; Al-Obaid, K.; Asadzadeh, M.; Jeragh, A.; et al. Changing trends in epidemiology and antifungal susceptibility patterns of six bloodstream Candida species isolates over a 12-year period in Kuwait. PLoS ONE 2019, 14, e0216250. [Google Scholar] [CrossRef] [Green Version]
- Taj-Aldeen, S.J.; Kolecka, A.; Boesten, R.; Alolaqi, A.; Almaslamani, M.; Chandra, P.; Meis, J.F.; Boekhout, T. Epidemiology of candidemia in Qatar, the Middle East: Performance of MALDI-TOF MS for the identification of Candida species, species distribution, outcome, and susceptibility pattern. Infection 2014, 42, 393–404. [Google Scholar] [CrossRef]
- Mirhendi, H.; Charsizadeh, A.; Eshaghi, H.; Nikmanesh, B.; Arendrup, M.C. Species distribution and antifungal susceptibility profile of Candida isolates from blood and other normally sterile foci from pediatric ICU patients in Tehran, Iran. Med. Mycol. 2020, 58, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Tortorano, A.M.; Prigitano, A.; Lazzarini, C.; Passera, M.; Deiana, M.L.; Cavinato, S.; De Luca, C.; Grancini, A.; Lo Cascio, G.; Ossi, C.; et al. A 1-year prospective survey of candidemia in Italy and changing epidemiology over one decade. Infection 2013, 41, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Puig-Asensio, M.; Padilla, B.; Garnacho-Montero, J.; Zaragoza, O.; Aguado, J.M.; Zaragoza, R.; Montejo, M.; Muñoz, P.; Ruiz-Camps, I.; Cuenca-Estrella, M.; et al. Epidemiology and predictive factors for early and late mortality in Candida bloodstream infections: A population-based surveillance in Spain. Clin. Microbiol. Infect. 2014, 20, O245–O254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimopoulos, G.; Velegraki, A.; Falagas, M.E. A 10-year survey of antifungal susceptibility of candidemia isolates from intensive care unit patients in Greece. Antimicrob. Agents Chemother. 2009, 53, 1242–1244. [Google Scholar] [CrossRef] [Green Version]
- Papadimitriou-Olivgeris, M.; Spiliopoulou, A.; Kolonitsiou, F.; Bartzavali, C.; Lambropoulou, A.; Xaplanteri, P.; Anastassiou, E.D.; Marangos, M.; Spiliopoulou, I.; Christofidou, M. Increasing incidence of candidaemia and shifting epidemiology in favor of Candida non-albicans in a 9-year period (2009–2017) in a university Greek hospital. Infection 2019, 47, 209–216. [Google Scholar] [CrossRef]
- Leroy, O.; Bailly, S.; Gangneux, J.P.; Mira, J.P.; Devos, P.; Dupont, H.; Montravers, P.; Perrigault, P.F.; Constantin, J.M.; Guillemot, D.; et al. Systemic antifungal therapy for proven or suspected invasive candidiasis: The AmarCAND 2 study. Ann. Intensive Care 2016, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Hesstvedt, L.; Gaustad, P.; Andersen, C.T.; Haarr, E.; Hannula, R.; Haukland, H.H.; Hermansen, N.O.; Larssen, K.W.; Mylvaganam, H.; Ranheim, T.E.; et al. Twenty-two years of candidaemia surveillance: Results from a Norwegian national study. Clin. Microbiol. Infect. 2015, 21, 938–945. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, E.; Hammarström, H.; Ataollahy, N.; Kondori, N. Species distribution and antifungal drug susceptibilities of yeasts isolated from the blood samples of patients with candidemia. Sci. Rep. 2019, 9, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Poikonen, E.; Lyytikäinen, O.; Anttila, V.J.; Koivula, I.; Lumio, J.; Kotilainen, P.; Syrjälä, H.; Ruutu, P. Secular trend in candidemia and the use of fluconazole in Finland, 2004–2007. BMC Infect. Dis. 2010, 10, 312. [Google Scholar] [CrossRef] [Green Version]
- Mari, A.H.; Valkonen, M.; Kolho, E.; Friberg, N.; Anttila, V.J. Clinical and microbiological factors associated with mortality in candidemia in adult patients 2007–2016. Infect. Dis. 2019, 51, 824–830. [Google Scholar]
- Sellami, A.; Sellami, H.; Néji, S.; Makni, F.; Abbes, S.; Cheikhrouhou, F.; Chelly, H.; Bouaziz, M.; Hammami, B.; Ben Jemaa, M.; et al. Antifungal susceptibility of bloodstream Candida isolates in Sfax Hospital: Tunisia. Mycopathologia 2011, 171, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Megri, Y.; Arastehfar, A.; Boekhout, T.; Daneshnia, F.; Hörtnagl, C.; Sartori, B.; Hafez, A.; Pan, W.; Lass-Flörl, C.; Hamrioui, B. Candida tropicalis is the most prevalent yeast species causing candidemia in Algeria: The urgent need for antifungal stewardship and infection control measures. Antimicrob. Resist. Infect. Control. 2020, 9, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nucci, M.; Queiroz-Telles, F.; Alvarado-Matute, T.; Tiraboschi, I.N.; Cortes, J.; Zurita, J.; Guzman-Blanco, M.; Santolaya, M.E.; Thompson, L.; Sifuentes-Osornio, J.; et al. Epidemiology of candidemia in Latin America: A laboratory-based survey. PLoS ONE 2013, 8, e59373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remington, T.L.; Isaac, A.; Vickers, D.M.; Fuller, J.; Wrenn-Smith, S. Epidemiology of candidemia at a tertiary Canadian hospital, 2004–2013. Can. J. Infect. Dis. Med. Microbiol. 2018, 3, 14–23. [Google Scholar] [CrossRef]
- Arastehfar, A.; Yazdanpanah, S.; Bakhtiari, M.; Fang, W.; Pan, W.; Mahmoudi, S.; Pakshir, K.; Daneshnia, F.; Boekhout, T.; Ilkit, M.; et al. Epidemiology of candidemia in Shiraz, southern Iran: A prospective multicenter study (2016–2018). Med. Mycol. 2020. [Google Scholar] [CrossRef]
- Kord, M.; Salehi, M.; Khodavaisy, S.; Hashemi, S.J.; Ghazvini, R.D.; Rezaei, S.; Maleki, A.; Elmimoghaddam, A.; Alijani, N.; Alireza Abdollahi, A.; et al. Epidemiology of yeast species causing bloodstream infection in Tehran, Iran (2015–2017); superiority of 21-plex PCR over the Vitek 2 system for yeast identification. J. Med. Microbiol. 2020, 69, 712–720. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous emergence of multidrug resistant Candida auris on 3 continents confirmed by whole-genome sequencing and epidemiological analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Vallabhaneni, S.; Kallen, A.; Tsay, S.; Chow, N.; Welsh, R.; Kerins, J.; Kemble, S.K.; Pacilli, M.; Black, S.R.; Landon, E.; et al. Investigation of the first seven reported cases of Candida auris, a globally emerging invasive, multidrug-resistant fungus—United States, May 2013–August 2016. MMWR. Morb. Mortal. Wkly. Rep. 2016, 65, 1234–1237. [Google Scholar] [CrossRef] [Green Version]
- Sharma, C.; Kumar, N.; Pandey, R.; Meis, J.F.; Chowdhary, A. Whole genome sequencing of emerging multidrug resistant Candida auris isolates in India demonstrates low genetic variation. New Microbes New Infect. 2016, 13, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Kumar, A.V.; Sharma, C.; Prakash, A.; Agarwal, K.; Babu, R.; Dinesh, K.R.; Karim, S.; Singh, S.K.; Hagen, F.; et al. Multidrug-resistant endemic clonal strain of Candida auris in India. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 919–926. [Google Scholar] [CrossRef]
- van der Linden, J.W.M.; Arendrup, M.C.; Warris, A.; Lagrou, K.; Pelloux, H.; Hauser, P.M.; Chryssanthou, E.; Mellado, E.; Kidd, S.E.; Tortorano, A.M.; et al. Prospective multicenter international surveillance of azole resistance in Aspergillus fumigatus. Emerg. Infect. Dis. 2015, 21, 1041–1044. [Google Scholar] [CrossRef] [PubMed]
- Lestrade, P.P.A.; Meis, J.F.; Arends, J.P.; van der Beek, M.T.; de Brauwer, E.; van Dijk, K.; de Greeff, S.C.; Haas, P.J.; Hodiamont, C.J.; Kuijper, E.J.; et al. Diagnosis and management of aspergillosis in the Netherlands: A national survey. Mycoses 2016, 59, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Menendez, O.; Alastruey-Izquierdo, A.; Mellado, E.; Cuenca-Estrella, M. Triazole resistance in Aspergillus spp.: A worldwide problem? J. Fungi 2016, 2, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Rubio, R.; Cuenca-Estrella, M.; Mellado, E. Triazole resistance in Aspergillus species: An emerging problem. Drugs 2017, 77, 599–613. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, E.; Maertens, J.; De Bel, A.; Nulens, E.; Boelens, J.; Surmont, I.; Mertens, A.; Boel, A.; Lagrou, K. Nationwide surveillance of azole resistance in Aspergillus diseases. Antimicrob. Agents Chemother. 2015, 59, 4569–4576. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, K.L.; Jensen, R.H.; Johansen, H.K.; Skov, M.; Pressler, T.; Howard, S.J.; Leatherbarrow, H.; Mellado, E.; Arendrup, M.C. Aspergillus species and other molds in respiratory samples from patients with cystic fibrosis: A laboratory based study with focus on Aspergillus fumigatus azole resistance. J. Clin. Microbiol. 2011, 49, 2243–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakare, N.; Rickerts, V.; Bargon, J.; Just-Nubling, G. Prevalence of Aspergillus fumigatus and other fungal species in the sputum of adult patients with cystic fibrosis. Mycoses 2003, 46, 19–23. [Google Scholar] [CrossRef]
- Burgel, P.R.; Baixench, M.T.; Amsellem, M.; Audureau, E.; Chapron, J.; Kanaan, R.; Honoré, I.; Dupouy-Camet, J.; Dusser, D.; Klaassen, C.H.; et al. High prevalence of azole-resistant Aspergillus fumigatus in adults with cystic fibrosis exposed to itraconazole. Antimicrob. Agents Chemother. 2012, 56, 869–874. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.; van Koningsbruggen-Rietschel, S.; Rietschel, E.; Vehreschild, M.J.; Wisplinghoff, H.; Kronke, M.; Hamprecht, A. Prevalence and molecular characterization of azole resistance in Aspergillus spp. isolates from German cystic fibrosis patients. J. Antimicrob. Chemother. 2014, 69, 1533–1536. [Google Scholar] [CrossRef] [Green Version]
- Alastruey-Izquierdo, A.; Mellado, E.; Peláez, T.; Pemán, J.; Zapico, S.; Alvarez, M.; Rodríguez-Tudela, J.L.; Cuenca-Estrella, M.; FILPOP Study Group. Population-based survey of filamentous fungi and antifungal resistance in Spain (FILPOP Study). Antimicrob. Agents Chemother. 2013, 57, 3380–3387. [Google Scholar] [CrossRef] [Green Version]
- Alastruey-Izquierdo, A.; Alcazar-Fuoli, L.; Rivero-Menéndez, O.; Ayats, J.; Castro, C.; García-Rodríguez, J.; Goterris-Bonet, L.; Ibáñez-Martínez, E.; Linares-Sicilia, M.J.; Martin-Gomez, M.T.; et al. Molecular identification and susceptibility testing of molds isolated in a prospective surveillance of triazole resistance in Spain (FILPOP2 Study). Antimicrob. Agents Chemother. 2018, 62, e00358-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamoth, F. Aspergillus fumigatus–related species in clinical practice. Front. Microbiol. 2016, 7, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falagas, M.E.; Roussos, N.; Vardakas, K.Z. Relative frequency of albicans and the various non-albicans Candida spp. among candidemia isolates from inpatients in various parts of the world: A systematic review. Int. J. Infect. Dis. 2010, 14, e954–e966. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Dzajic, E.; Jensen, R.H.; Johansen, H.K.; Kjaeldgaard, P.; Knudsen, J.D.; Kristensen, L.; Leitz, C.; Lemming, L.E.; Nielsen, L.; et al. Epidemiological changes with potential implication for antifungal prescription recommendations for fungaemia: Data from a nationwide fungaemia surveillance programme. Clin. Microbiol. Infect. 2013, 19, e343–e353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, R.H.; Hagen, F.; Astvad, K.M.T.; Tyron, A.; Meis, J.F.; Arendrup, M.C. Azole-resistant Aspergillus fumigatus in Denmark: A laboratory-based study on resistance mechanisms and genotypes. Clin. Microbiol. Infect. 2016, 22, e1–e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, K.; Suzuki, J.; Watanabe, A.; Arai, T.; Koiwa, T.; Shinfuku, K.; Narumoto, O.; Kawashima, M.; Fukami, T.; Tamura, A.; et al. High detection rate of azole-resistant Aspergillus fumigatus after treatment with azole antifungal drugs among patients with chronic pulmonary aspergillosis in a single hospital setting with low azole resistance. Med. Mycol. 2020. [Google Scholar] [CrossRef]
- Devoto, T.B.; Hermida-Alva, K.; Posse, G.; Finquelievich, J.L.; García-Effrón, G.; Cuestas, M.L. High prevalence of triazole-resistant Aspergillus fumigatus sensu stricto in an Argentinean cohort of patients with cystic fibrosis. Mycoses 2020, 63, 937–941. [Google Scholar] [CrossRef]
- Alcazar-Fuoli, L.; Mellado, E.; Alastruey-Izquierdo, A.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Aspergillus section Fumigati: Antifungal susceptibility patterns and sequence-based identification. Antimicrob. Agents Chemother. 2018, 52, 1244–1251. [Google Scholar] [CrossRef] [Green Version]
- Whitesell, L.; Robbins, N.; Huang, D.S.; McLellan, C.A.; Shekhar-Guturja, T.; LeBlanc, E.V.; Nation, C.S.; Hui, R.; Hutchinson, A.; Collins, C.; et al. Structural basis for species-selective targeting of Hsp90 in a pathogenic fungus. Nat. Commun. 2019, 10, 402. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Iyer, K.R.; Pardeshi, L.; Muñoz, J.F.; Robbins, N.; Cuomo, C.A.; Wong, K.H.; Cowen, L.E. Genetic analysis of Candida auris implicates Hsp90 in morphogenesis and azole tolerance and Cdr1 in azole resistance. mBio 2019, 10, e02529-18. [Google Scholar] [CrossRef] [Green Version]
- Cowen, L.E.; Singh, S.D.; Köhler, J.R.; Collins, C.; Zaas, A.K.; Schell, W.A.; Aziz, H.; Mylonakis, E.; Perfect, J.R.; Whitesell, L.; et al. Harnessing Hsp90 function as a powerful, broadly effective therapeutic strategy for fungal infectious disease. Proc. Natl. Acad. Sci. USA 2009, 106, 2818–2823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garzon, A.C.; Amado, D.; Robert, E.; Parra Giraldo, C.M.; Le Pape, P. Impact of calmodulin inhibition by fluphenazine on susceptibility, biofilm formation and pathogenicity of caspofungin-resistant Candida glabrata. J. Antimicrob. Chemother. 2020, 75, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Caplan, T.; Lorente-Macías, A.; Stogios, P.J.; Evdokimova, E.; Hyde, S.; Wellington, M.A.; Liston, S.; Iyer, K.R.; Puumala, E.; Shekhar-Guturja, T.; et al. Overcoming fungal echinocandin resistance through inhibition of the non-essential stress kinase Yck2. Cell. Chem. Biol. 2020, 27, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, A.; Ene, I.V.; Bibi, M.; Zakin, S.; Segal, E.S.; Ziv, N.; Dahan, A.M.; Colombo, A.L.; Bennett, R.J.; Berman, J. Antifungal tolerance is a subpopulation effect distinct from resistance and is associated with persistent candidemia. Nat. Commun. 2018, 9, 2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astvad, K.M.T.; Sanglard, D.; Delarze, E.; Hare, R.K.; Arendrup, M.C. Implications of the EUCAST trailing phenomenon in Candida tropicalis for the in vivo susceptibility in invertebrate and murine models. Antimicrob. Agents Chemother. 2018, 62, e01624-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Healey, K.R.; Jiménez-Ortigosa, C.; Shor, E.; Perlin, D.S. Genetic drivers of multidrug resistance in Candida glabrata. Front. Microbiol. 2016, 7, 1995. [Google Scholar] [CrossRef] [Green Version]
- Arastehfar, A.; Hilmioğlu-Polat, S.; Daneshnia, F.; Hafez, A.; Salehi, M.; Polat, F.; Yaşar, M.; Arslan, N.; Hoşbul, T.; Ünal, N.; et al. Recent increase in the prevalence of fluconazole-non-susceptible Candida tropicalis blood isolates in Turkey: Clinical implication of azole-non-susceptible and fluconazole tolerant phenotypes and genotyping. Front. Microbiol. 2020, 11, 587278. [Google Scholar] [CrossRef]
- Gerstein, A.C.; Berman, J. Candida albicans genetic background influences mean and heterogeneity of drug responses and genome stability during evolution in fluconazole. mSphere 2020, 5, e00480-20. [Google Scholar] [CrossRef]
- Schmidt, F.; Thywißen, A.; Goldmann, M.; Cunha, C.; Cseresnyés, Z.; Schmidt, H.; Rafiq, M.; Galiani, S.; Gräler, M.H.; Chamilos, G.; et al. Flotillin-dependent membrane microdomains are required for functional phagolysosomes against fungal infections. Cell. Rep. 2020, 32, 108017. [Google Scholar] [CrossRef]
- Arastehfar, A.; Daneshnia, F.; Zomorodian, K.; Najafzadeh, M.J.; Khodavaisy, S.; Zarrinfar, H.; Hagen, F.; Shahrabadi, Z.Z.; Lackner, M.; Mirhendi, H.; et al. Low level of antifungal resistance in Iranian isolates of Candida glabrata recovered from blood samples in a multicenter study from 2015 to 2018 and potential prognostic values of genotyping and sequencing of PDR1. Antimicrob. Agents Chemother. 2019, 63, e02503-18. [Google Scholar] [CrossRef] [Green Version]
- Arastehfar, A.; Daneshnia, F.; Najafzadeh, M.J.; Hagen, F.; Mahmoudi, S.; Salehi, M.; Zarrinfar, H.; Namvar, Z.; Shahrabadi, Z.Z.; Khodavaisy, S.; et al. Evaluation of molecular epidemiology, clinical characteristics, antifungal susceptibility profiles, and molecular mechanisms of antifungal resistance of Iranian Candida parapsilosis species complex blood isolates. Front. Cell. Infect. Microbiol. 2020, 10, 206. [Google Scholar] [CrossRef] [PubMed]
- Arastehfar, A.; Wickes, B.L.; Ilkit, M.; Pincus, D.H.; Daneshnia, F.; Pan, W.; Fang, W.; Boekhout, T. Identification of mycoses in developing countries. J. Fungi 2019, 5, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimoto, A.T.; Sharma, C.; Rogers, P.D. Molecular and genetic basis of azole antifungal resistance in the opportunistic pathogenic fungus Candida albicans. J. Antimicrob. Chemother. 2020, 75, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Healey, K.R.; Kordalewska, M.; Jiménez-Ortigosa, C.; Singh, A.; Berrío, I.; Chowdhary, A.; Perlin, D.S. Limited ERG11 mutations identified in isolates of Candida auris directly contribute to reduced azole susceptibility. Antimicrob. Agents Chemother. 2018, 62, e01427-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Xiao, M.; Zhang, D.; Huang, J.J.; Wang, H.; Hou, X.; Zhang, L.; Kong, F.; Chen, S.C.A.; Tong, Z.H.; et al. Molecular mechanisms of azole resistance in Candida tropicalis isolates causing invasive candidiasis in China. Clin. Microbiol. Infect. 2019, 25, 885–891. [Google Scholar] [CrossRef]
- Yao, D.; Chen, J.; Chen, W.; Li, Z.; Hu, X. Mechanisms of azole resistance in clinical isolates of Candida glabrata from two hospitals in China. Infect. Drug Resist. 2019, 12, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Branco, J.; Ola, M.; Silva, R.M.; Fonseca, E.; Gomes, N.C.; Martins-Cruz, C.; Silva, A.P.; Silva-Dias, A.; Pina-Vaz, C.; Erraught, C.; et al. Impact of ERG3 mutations and expression of ergosterol genes controlled by UPC2 and NDT80 in Candida parapsilosis azole resistance. Clin. Microbiol. Infect. 2017, 23, 575.e1–575.e8. [Google Scholar] [CrossRef]
- Yang, H.; Tong, J.; Lee, C.W.; Ha, S.; Eom, S.H.; Im, Y.J. Structural mechanism of ergosterol regulation by fungal sterol transcription factor Upc2. Nat. Commun. 2015, 6, 6129. [Google Scholar] [CrossRef] [Green Version]
- Vu, B.G.; Thomas, G.H.; Moye-Rowley, W.S. Evidence that ergosterol biosynthesis modulates activity of the Pdr1 transcription factor in Candida glabrata. mBio 2019, 10, e00934-19. [Google Scholar] [CrossRef] [Green Version]
- Pais, P.; Califórnia, R.; Galocha, M.; Viana, R.; Ola, M.; Cavalheiro, M.; Takahashi-Nakaguchi, A.; Chibana, H.; Butler, G.; Teixeira, M.C. Candida glabrata transcription factor Rpn4 mediates fluconazole resistance through regulation of ergosterol biosynthesis and plasma membrane permeability. Antimicrob. Agents Chemother. 2020, 64, e00554-20. [Google Scholar] [CrossRef]
- Perlin, D.S.; Shor, E.; Zhao, Y. Update on antifungal drug resistance. Curr. Clin. Microbiol. Rep. 2015, 2, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Arastehfar, A.; Lass-Flörl, C.; Garcia-Rubio, R.; Daneshnia, F.; Ilkit, M.; Boekhout, T.; Gabaldón, T.; Perlin, D.S. The quiet and underappreciated rise of drug-resistant invasive fungal pathogens. J. Fungi 2020, 6, 138. [Google Scholar] [CrossRef] [PubMed]
- Arastehfar, A.; Daneshnia, F.; Hafez, A.; Khodavaisy, S.; Najafzadeh, M.J.; Charsizadeh, A.; Zarrinfar, H.; Salehi, M.; Shahrabadi, Z.Z.; Sasani, E.; et al. Antifungal susceptibility, genotyping, resistance mechanism, and clinical profile of Candida tropicalis blood isolates. Med. Mycol. 2020, 58, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Grossman, N.T.; Pham, C.D.; Cleveland, A.A.; Lockhart, S.R. Molecular mechanisms of fluconazole resistance in Candida parapsilosis isolates from a U.S. surveillance system. Antimicrob. Agents Chemother. 2015, 59, 1030–1037. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.J.; Kim, Y.J.; Yong, D.; Byun, J.H.; Kim, T.S.; Chang, Y.S.; Choi, M.J.; Byeon, S.A.; Won, E.J.; Kim, S.H.; et al. Fluconazole-resistant Candida parapsilosis bloodstream isolates with Y132F mutation in ERG11 gene, South Korea. Emerg. Infect. Dis. 2018, 24, 1768–1770. [Google Scholar] [CrossRef] [Green Version]
- Rybak, J.M.; Muñoz, J.M.; Barker, K.S.; Parker, J.E.; Esquivel, B.D.; Berkow, E.L.; Lockhart, S.R.; Gade, L.; Palmer, G.E.; White, T.C. Mutations in TAC1B: A novel genetic determinant of clinical fluconazole resistance in Candida auris. mBio 2020, 11, e00365-20. [Google Scholar] [CrossRef]
- Hou, X.; Xiao, M.; Wang, H.; Yu, S.Y.; Zhang, G.; Zhao, Y.; Xu, Y.C. Profiling of PDR1 and MSH2 in Candida glabrata bloodstream isolates from a multicenter study in China. Antimicrob. Agents Chemother. 2018, 62, e00153-18. [Google Scholar] [CrossRef] [Green Version]
- Cavalheiro, M.; Costa, C.; Silva-Dias, A.; Miranda, I.M.; Wang, C.; Pais, P.; Pinto, S.N.; Mil-Homens, D.; Sato-Okamoto, M.; Takahashi-Nakaguchi, A.; et al. A transcriptomics approach to unveiling the mechanisms of in vitro evolution towards fluconazole resistance of a Candida glabrata clinical isolate. Antimicrob. Agents Chemother. 2018, 63, e00995-18. [Google Scholar] [CrossRef] [Green Version]
- Vale-Silva, L.; Ischer, F.; Leibundgut-Landmann, S.; Sanglard, D. Gain-of-function mutations in PDR1, a regulator of antifungal drug resistance in Candida glabrata, control adherence to host cells. Infect. Immun. 2013, 81, 1709–1720. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, S.; Ischer, F.; Calabrese, D.; Posteraro, B.; Sanguinetti, M.; Fadda, G.; Rohde, B.; Bauser, C.; Bader, O.; Sanglard, D. Gain of function mutations in CgPDR1 of Candida glabrata not only mediate antifungal resistance but also enhance virulence. PLoS Pathog. 2009, 5, e1000268. [Google Scholar] [CrossRef] [Green Version]
- Dudakova, A.; Spiess, B.; Tangwattanachuleeporn, M.; Sasse, C.; Buchheidt, D.; Weig, M.; Groβ, U.; Bader, O. Molecular tools for the detection and deduction of azole antifungal drug resistance phenotypes in Aspergillus species. Clin. Microbiol. Rev. 2017, 30, 1065–1091. [Google Scholar] [CrossRef] [Green Version]
- Willger, S.D.; Puttikamonkul, S.; Kim, K.H.; Burritt, J.B.; Grahl, N.; Metzler, L.J.; Barbuch, R.; Bard, M.; Lawrence, C.B.; Cramer, R.A., Jr. A sterol-regulatory element binding protein is required for cell polarity, hypoxia adaptation, azole drug resistance, and virulence in Aspergillus fumigatus. PLoS Pathog. 2008, 4, e1000200. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara, D.; Miura, D.; Shimizu, K.; Paul, S.; Ohba, A.; Gonoi, T.; Watanabe, A.; Kamei, K.; Shintani, T.; Moye-Rowley, W.S.; et al. A novel Zn2-Cys6 transcription factor AtrR plays a key role in an azole resistance mechanism of Aspergillus fumigatus by co-regulating cyp51A and cdr1B expressions. PLoS Pathog. 2017, 13, e1006096. [Google Scholar] [CrossRef]
- Gsaller, F.; Hortschansky, P.; Furukawa, T.; Carr, P.D.; Rash, B.; Capilla, J.; Müller, C.; Bracher, F.; Bowyer, P.; Haas, H.; et al. Sterol biosynthesis and azole tolerance is governed by the opposing actions of SrbA and the CCAAT binding complex. PLoS Pathog. 2016, 12, e1005775. [Google Scholar]
- Camps, S.M.T.; Dutilh, B.E.; Arendrup, M.C.; Rijs, A.J.M.M.; Snelders, E.; Huynen, M.A.; Verweij, P.E.; Melchers, W.J.G. Discovery of a HapE mutation that causes azole resistance in Aspergillus fumigatus through whole genome sequencing and sexual crossing. PLoS ONE 2012, 7, e50034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ukai, Y.; Kuroiwa, M.; Kurihara, N.; Naruse, H.; Homma, T.; Maki, H.; Naito, A. Contributions of yap1 mutation and subsequent atrF upregulation to voriconazole resistance in Aspergillus flavus. Antimicrob. Agents Chemother. 2018, 62, e01216-18. [Google Scholar] [CrossRef] [Green Version]
- dos Reis, T.F.; Pereira Silva, L.; Alves de Castro, P.; do Carmo, R.A.; Mendes Marini, M.; da Silveira, J.F.; Ferreira, B.H.; Rodrigues, F.; Lind, A.L.; Rokas, A.; et al. The Aspergillus fumigatus mismatch repair MSH2 homolog is important for virulence and azole resistance. mSphere 2019, 4, e00416–e00419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rybak, J.M.; Ge, W.; Wiederhold, N.P.; Parker, J.E.; Kelly, S.L.; Rogers, P.D.; Fortwendel, J.R. Mutations in hmg1, challenging the paradigm of clinical triazole resistance in Aspergillus fumigatus. mBio 2019, 10, e00437-19. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Zhai, P.; Zhang, Y.; Zhang, C.; Sang, H.; Han, G.; Keller, N.P.; Lu, L. The Aspergillus fumigatus damage resistance protein family coordinately regulates ergosterol biosynthesis and azole susceptibility. mBio 2016, 7, e01919-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, T.; van Rhijn, N.; Fraczek, M.; Gsaller, F.; Davies, E.; Carr, P.; Gago, S.; Fortune-Grant, R.; Rahman, S.; Gilsenan, J.M.; et al. The negative cofactor 2 complex is a key regulator of drug resistance in Aspergillus fumigatus. Nat. Commun. 2020, 11, 427. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Perlin, D.S. Echinocandin resistance: An emerging clinical problem? Curr. Opin. Infect. Dis. 2014, 27, 484–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Ortigosa, C.; Moore, C.; Denning, D.W.; Perlin, D.S. Emergence of echinocandin resistance due to a point mutation in the fks1 gene of Aspergillus fumigatus in a patient with chronic pulmonary aspergillosis. Antimicrob. Agents Chemother. 2017, 61, e01277-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, C.D.; Iqbal, N.; Bolden, C.B.; Kuykendall, R.J.; Harrison, L.H.; Farley, M.M.; Schaffner, W.; Beldavs, Z.G.; Chiller, T.M.; Park, B.J.; et al. Role of FKS mutations in Candida glabrata: MIC values, echinocandin resistance, and multidrug resistance. Antimicrob. Agents Chemother. 2014, 58, 4690–4696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perlin, D.S. Resistance to echinocandin-class antifungal drugs. Drug Resist. Updat. 2007, 10, 121–130. [Google Scholar] [CrossRef] [Green Version]
- García-Effrón, G.; Katiyar, S.K.; Park, S.; Edlind, T.D.; Perlin, D.S. A naturally occurring proline-to-alanine amino acid change in Fks1p in Candida parapsilosis, Candida orthopsilosis, and Candida metapsilosis accounts for reduced echinocandin susceptibility. Antimicrob. Agents Chemother. 2008, 52, 2305–2312. [Google Scholar] [CrossRef] [Green Version]
- Arastehfar, A.; Daneshnia, F.; Hilmioğlu-Polat, S.; Ilkit, M.; Yaşar, M.; Polat, F.; Metin, D.Y.; Dökümcü, Ü.Z.; Pan, W.; Hagen, F.; et al. Genetically-related micafungin-resistant C. parapsilosis blood isolates harboring a novel mutation R658G in hotspot1-Fks1p: A new challenge? J. Antimicrob. Chemother. 2020. [Google Scholar] [CrossRef]
- Verweij, P.E.; Ananda-Rajah, M.; Andes, D.; Arendrup, M.C.; Brüggemann, R.J.; Chowdhary, A.; Cornely, O.A.; Denning, D.W.; Groll, A.H.; Izumikawa, K.; et al. International expert opinion on the management of infection caused by azole-resistant Aspergillus fumigatus. Drug Resist. Updat. 2015, 21–22, 30–40. [Google Scholar] [CrossRef]
- Satish, S.; Jiménez-Ortigosa, C.; Zhao, Y.; Lee, M.H.; Dolgov, E.; Krüger, T.; Park, S.; Denning, D.W.; Kniemeyer, O.; Brakhage, A.A.; et al. Stress-induced changes in the lipid microenvironment of β-(1,3)-D-glucan synthase cause clinically important echinocandin resistance in Aspergillus fumigatus. mBio 2019, 10, e00779-19. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Healey, K.R.; Shor, E.; Kordalewska, M.; Jiménez-Ortigosa, C.; Paderu, P.; Xiao, M.; Wang, H.; Zhao, Y.; Lin, L.Y.; et al. Novel FKS1 and FKS2 modifications in a high-level echinocandin resistant clinical isolate of Candida glabrata. Emerg. Microbes Infect. 2019, 8, 1619–1625. [Google Scholar] [CrossRef] [Green Version]
- Kano, R.; Okubo, M.; Hasegawa, A.; Kamata, H. Multi-azole-resistant strains of Cryptococcus neoformans var. grubii isolated from a FLZ-resistant strain by culturing in medium containing voriconazole. Med. Mycol. 2017, 55, 877–882. [Google Scholar]
- Kano, R.; Sugita, T.; Kamata, H. Antifungal susceptibility of clinical isolates and artificially produced multi-azole-resistant strains of Cryptococcus neoformans (formaly: Cryptococcus grubii) to ravuconazole. Med. Mycol. J. 2020, 61, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Mondon, P.; Petter, R.; Amalfitano, G.; Luzzati, R.; Concia, E.; Polacheck, I.; Kwon-Chung, K.J. Heteroresistance to fluconazole and voriconazole in Cryptococcus neoformans. Antimicrob. Agents Chemother. 1999, 43, 1856–1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sionov, E.; Chang, Y.C.; Kwon-Chung, K.J. Azole heteroresistance in Cryptococcus neoformans: Emergence of resistant clones with chromosomal disomy in the mouse brain during fluconazole treatment. Antimicrob. Agents Chemother. 2013, 57, 5127–5130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hope, W.; Stone, N.R.H.; Johnson, A.; McEntee, L.; Farrington, N.; Santoro-Castelazo, A.; Liu, X.; Lucaci, A.; Hughes, M.; Oliver, J.D.; et al. Fluconazole monotherapy is a suboptimal option for initial treatment of cryptococcal meningitis because of emergence of resistance. mBio 2019, 10, e02575-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billmyre, R.B.; Applen-Clancey, S.; Li, L.X.; Doering, T.L.; Heitman, J. 5-fluorocytosine resistance is associated with hypermutation and alterations in capsule biosynthesis in Cryptococcus. Nat. Commun. 2020, 11, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, N.R.H.; Rhodes, J.; Fisher, M.C.; Mfinanga, S.; Kivuyo, S.; Rugemalila, J.; Segal, E.S.; Needleman, L.; Molloy, S.F.; Kwon-Chung, J.; et al. Dynamic ploidy changes drive fluconazole resistance in human cryptococcal meningitis. J. Clin. Investig. 2019, 129, 999–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maligie, M.A.; Selitrennikoff, C.P. Cryptococcus neoformans resistance to echinocandins: (1,3) beta-glucan synthase activity is sensitive to echinocandins. Antimicrob. Agents Chemother. 2005, 49, 2851–2856. [Google Scholar] [CrossRef] [Green Version]
- van Duin, D.; Casadevall, A.; Nosanchuk, J.D. Melanization of Cryptococcus neoformans and Histoplasma capsulatum reduces their susceptibilities to amphotericin B and caspofungin. Antimicrob. Agents Chemother. 2002, 46, 3394–3400. [Google Scholar] [CrossRef] [Green Version]
- Kaneshiro, E.S. Sterol metabolism in the opportunistic pathogen Pneumocystis: Advances and new insights. Lipids 2004, 39, 753–761. [Google Scholar] [CrossRef]
- Porollo, A.; Meller, J.; Joshi, Y.; Jaiswal, V.; Smulian, A.G.; Cushion, M.T. Analysis of current antifungal agents and their targets within the Pneumocystis carinii genome. Curr. Drug Targets 2012, 13, 1575–1585. [Google Scholar] [CrossRef] [Green Version]
- Volpe, F.; Dyer, M.; Scaife, J.G.; Darby, G.; Stammers, D.K.; Delves, C.J. The multifunctional folic acid synthesis fas gene of Pneumocystis carinii appears to encode dihydropteroate synthase and hydroxymethyldihydropterin pyrophosphokinase. Gene 1992, 112, 213–218. [Google Scholar] [CrossRef]
- Ma, L.; Borio, L.; Masur, H.; Kovacs, J.A. Pneumocystis carinii dihydropteroate synthase but not dihydrofolate reductase gene mutations correlate with prior trimethoprim-sulfamethoxazole or dapsone use. J. Infect. Dis. 1999, 180, 1969–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, C.R.; Poole, C.; Kazanjian, P.; Meshnick, S.R. Sulfa use, dihydropteroate synthase mutation, and Pneumocystis jirovecii pneumonia. Emerg. Infect. Dis. 2004, 10, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Mirdha, B.R.; Guleria, R.; Kabra, S.K.; Mohan, A.; Chaudhry, R.; Kumar, L.; Dwivedi, S.N.; Agarwal, S.K. Novel dihydropteroate synthase gene mutation in Pneumocystis jirovecii among HIV-infected patients in India: Putative association with drug resistance and mortality. J. Glob. Antimicrob. Resist. 2019, 17, 236–239. [Google Scholar] [CrossRef] [PubMed]
- Crothers, K.; Beard, C.B.; Turner, J.; Groner, G.; Fox, M.; Morris, A.; Eiser, S.; Huang, L. Severity and outcome of HIV-associated Pneumocystis pneumonia containing Pneumocystis jirovecii dihydropteroate synthase gene mutaion. AIDS 2005, 19, 801–805. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Mirdha, B.R.; Guleria, R.; Kabra, S.K.; Mohan, A.; Chaudhry, R.; Kumar, L.; Dwivedi, S.N.; Agarwal, S.K. Genetic polymorphisms associated with treatment failure and mortality in pediatric pneumocystosis. Sci. Rep. 2019, 9, 1192. [Google Scholar] [CrossRef] [Green Version]
- Knabl, L.; Lass-Flörl, C. Antifungal susceptibility testing in Candida species: Current methods and promising new tools for shortening the turnaround time. Expert Rev. Anti Infect. Ther. 2020, 18, 779–787. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Cuenca-Estrella, M.; Lass-Flörl, C.; Hope, W.; EUCAST-AFST Collaborators. EUCAST technical note on the EUCAST definitive document EDef 7.2: Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for yeasts EDef 7.2 (EUCAST-AFST). Clin. Microbiol. Infect. 2012, 18, E246–E247. [Google Scholar] [CrossRef] [Green Version]
- Eggimann, P.; Que, Y.A.; Revelly, J.P.; Pagani, J.L. Preventing invasive Candida infections. Where could we do better? J. Hosp. Infect. 2015, 89, 302–308. [Google Scholar] [CrossRef] [Green Version]
- Berkow, E.L.; Lockhart, S.R.; Ostrosky-Zeichner, L. Antifungal susceptibility testing: Current approaches. Clin. Microbiol. Rev. 2020, 33, e00069-19. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Meletiadis, J.; Mouton, J.W.; Guinea, J.; Cuenca-Estrella, M.; Lagrou, K.; Howard, S.J. Subcommittee on Antifungal Susceptibility Testing (AFST) of the ESCMID European Committee for Antimicrobial Susceptibility Testing (EUCAST). EUCAST technical note on isavuconazole breakpoints for Aspergillus, itraconazole breakpoints for Candida and updates for the antifungal susceptibility testing method documents. Clin. Microbiol. Infect. 2016, 22, 571.e1–571.e4. [Google Scholar]
- Clinical and Laboratory Standards Institute. M27-A3: Reference Method for Broth Microdilution Antifungal Susceptibility Testing for Yeasts; CLSI: Wayne, PA, USA, 2019. [Google Scholar]
- Clinical and Laboratory Standards Institute. M38-A2: Reference Method for Broth Microdilution Antifungal Susceptibility Testing of Filamenteous Fungi; CLSI: Wayne, PA, USA, 2019. [Google Scholar]
- Meletiadis, J.; Siopi, M.; Kanioura, L.; Jørgensen, K.M.; Perlin, D.S.; Mouton, J.W.; Arendrup, M.C. Development and multicentre validation of an agar-based screening method for echinocandin susceptibility testing of Aspergillus species. J. Antimicrob. Chemother. 2019, 74, 2247–2254. [Google Scholar] [CrossRef] [PubMed]
- Delma, F.Z.; Al-Hatmi, A.M.S.; Buil, J.B.; van der Lee, H.; Tehupeiory-Kooreman, M.; de Hoog, G.S.; Meletiadis, J.; Verweij, P.E. Comparison of MIC Test Strip and Sensititre YeastOne with the CLSI and EUCAST broth microdilution reference methods for in vitro antifungal susceptibility testing of Cryptococcus neoformans. Antimicrob. Agents Chemother. 2020, 64, e02261-19. [Google Scholar] [CrossRef]
- Bassetti, M.; Vena, A.; Bouza, E.; Peghin, M.; Muñoz, P.; Righi, E.; Pea, F.; Lackner, M.; Lass-Flörl, C. Antifungal susceptibility testing in Candida, Aspergillus and Cryptococcus infections: Are the MICs useful for clinicians? Clin. Microbiol. Infect. 2020, 26, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Vatanshenassan, M.; Boekhout, T.; Lass-Flörl, C.; Lackner, M.; Schubert, S.; Kostrzewa, M.; Sparbier, K. Proof of concept for MBT ASTRA, a rapid matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS)-based method to detect caspofungin resistance in Candida albicans and Candida glabrata. J. Clin. Microbiol. 2018, 56, e00420-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatanshenassan, M.; Arastehfar, A.; Boekhout, T.; Berman, J.; Lass-Flörl, C.; Sparbier, K.; Kostrzewa, M. Anidulafungin susceptibility testing of Candida glabrata isolates from blood cultures by the MALDI Biotyper Antibiotic (Antifungal) susceptibility test rapid assay. Antimicrob. Agents Chemother. 2019, 63, e00554-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ksiezopolska, E.; Gabaldón, T. Evolutionary emergence of drug resistance in Candida opportunistic pathogens. Genes 2018, 9, 461. [Google Scholar] [CrossRef] [Green Version]
- Consortium, O.; Gabaldón, T. Recent trends in molecular diagnostics of yeast infections: From PCR to NGS. FEMS Microbiol. Rev. 2019, 43, 517–547. [Google Scholar] [CrossRef] [Green Version]
- Greninger, A.L. The challenge of diagnostic metagenomics. Expert Rev. Mol. Diagn. 2018, 18, 605–615. [Google Scholar] [CrossRef] [Green Version]
- Kidd, S.E.; Chen, S.C.A.; Meyer, W.; Halliday, C.L. A new age in molecular diagnostics for invasive fungal disease: Are we ready? Front. Microbiol. 2020, 10, 2903. [Google Scholar] [CrossRef] [Green Version]
- Perlin, D.S.; Wiederhold, N.P. Culture-independent molecular methods for detection of antifungal resistance mechanisms and fungal identification. J. Infect. Dis. 2017, 216, S458–S465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frade, J.P.; Warnock, D.W.; Arthington-Skaggs, B.A. Rapid quantification of drug resistance gene expression in Candida albicans by reverse transcriptase LightCycler PCR and fluorescent probe hybridization. J. Clin. Microbiol. 2004, 42, 2085–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.F.; Sammons, L.R.; Zhang, X.; Suffis, S.D.; Su, Q.; Myers, T.G.; Marr, K.A.; Bennett, J.E. Microarray and molecular analyses of the azole resistance mechanism in Candida glabrata oropharyngeal isolates. Antimicrob. Agents Chemother. 2010, 54, 3308–3317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, C.; Nelson-Sathi, S.; Singh, A.; Radhakrishna Pillai, M.; Chowdhary, A. Genomic perspective of triazole resistance in clinical and environmental Aspergillus fumigatus isolates without cyp51A mutations. Fungal Genet. Biol. 2019, 132, 103265. [Google Scholar] [CrossRef] [PubMed]
- De Backer, M.D.; Ilyina, T.; Ma, X.J.; Vandoninck, S.; Luyten, W.H.; Vanden Bossche, H. Genomic profiling of the response of Candida albicans to itraconazole treatment using a DNA microarray. Antimicrob. Agents Chemother. 2001, 45, 1660–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montesinos, I.; Delforge, M.L.; Ajjaham, F.; Brancart, F.; Hites, M.; Jacobs, F.; Denis, O. Evaluation of a new commercial real-time PCR assay for diagnosis of Pneumocystis jirovecii pneumonia and identification of dihydropteroate synthase (DHPS) mutations. Diagn. Microbiol. Infect. Dis. 2017, 87, 32–36. [Google Scholar] [CrossRef]
- Stoesser, N.; Batty, E.M.; Eyre, D.W.; Morgan, M.; Wyllie, D.H.; Del Ojo, C.E.; Johnson, J.R.; Walker, A.S.; Peto, T.E.A.; Crook, D.W. Predicting antimicrobial susceptibilities for Escherichia coli and Klebsiella pneumoniae isolates using whole genomic sequence data. J. Antimicrob. Chemother. 2013, 68, 2234–2244. [Google Scholar] [CrossRef]
- Ferreira, I.; Beisken, S.; Lueftinger, L.; Weinmaier, T.; Klein, M.; Bacher, J.; Patel, R.; Arndt von Haeseler, A.; Posch, A.E. Species identification and antibiotic resistance prediction by analysis of whole-genome sequence data by use of ARESdb: An analysis of isolates from the unyvero lower respiratory tract infection trial. J. Clin. Microbiol. 2020, 58, e00273-20. [Google Scholar] [CrossRef] [Green Version]
- Gupta, C.L.; Tiwari, R.K.; Cytryn, E. Platforms for elucidating antibiotic resistance in single genomes and complex metagenomes. Environ. Int. 2020, 138, 105667. [Google Scholar] [CrossRef]
- Graf, E.H.; Pancholi, P. Appropriate use and future directions of molecular diagnostic testing. Curr. Infect. Dis. Rep. 2020, 22, 5. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.Y.; Miller, S.A. Clinical metagenomics. Nat. Rev. Genet. 2019, 20, 341–355. [Google Scholar] [CrossRef]
- Rice, J.P.; Seifert, M.; Moser, K.S.; Rodwell, T.C. Performance of the Xpert MTB/RIF assay for the diagnosis of pulmonary tuberculosis and rifampin resistance in a low-incidence, high-resource setting. PLoS ONE 2017, 12, e0186139. [Google Scholar] [CrossRef] [Green Version]
- Ullmann, A.J.; Aguado, J.M.; Arikan-Akdagli, S.; Denning, D.W.; Groll, A.H.; Lagrou, K.; Lass-Flörl, C.; Lewis, R.E.; Munoz, P.; Verweij, P.E.; et al. Diagnosis and management of Aspergillus diseases: Executive summary of the 2017 ESCMID-ECMM-ERS guideline. Clin. Microbiol. Infect. 2018, 24 (Suppl. 1), e1–e38. [Google Scholar] [CrossRef] [Green Version]
- Tissot, F.; Agrawal, S.; Pagano, L.; Petrikkos, G.; Groll, A.H.; Skiada, A.; Lass-Flörl, C.; Calandra, T.; Viscoli, C.; Herbrecht, R. ECIL-6 guidelines for the treatment of invasive candidiasis, aspergillosis and mucormycosis in leukemia and hematopoietic stem cell transplant patients. Hematologica 2017, 102, 433–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denning, D.W.; Cadranel, J.; Beigelman-Aubry, C.; Ader, F.; Chakrabarti, A.; Blot, S.; Ullmann, A.J.; Dimopoulos, G.; Lange, C.; European Society for Clinical Microbiology and Infectious Diseases and European Respiratory Society. Chronic pulmonary aspergillosis: Rationale and clinical guidelines for diagnosis and management. Eur. Respir. J. 2016, 47, 45–68. [Google Scholar] [CrossRef]
- Cuenca-Estrella, M.; Verweij, P.E.; Arendrup, M.C.; Arikan-Akdagli, S.; Bille, J.; Donnelly, J.P.; Jensen, E.H.; Lass-Flörl, C.; Richardson, M.D.; Akova, M.; et al. ESCMID* guideline for the diagnosis and management of Candida diseases 2012: Diagnostic procedures. Clin. Microbiol. Infect. 2012, 18 (Suppl. 7), 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andes, D.; Marchillo, K.; Conklin, R.; Krishna, G.; Ezzet, F.; Cacciapuoti, A.; Loebenberg, D. Pharmacodynamics of a new triazole, posaconazole, in a murine model of disseminated candidiasis. Antimicrob. Agents Chemother. 2004, 48, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Howard, S.J.; Lestner, J.M.; Sharp, A.; Gregson, L.; Goodwin, J.; Slater, J.; Majithiya, J.B.; Warn, P.A.; Hope, W.W. Pharmacokinetics and pharmacodynamics of posaconazole for invasive pulmonary aspergillosis: Clinical implications for antifungal therapy. J. Infect. Dis. 2011, 203, 1324–1332. [Google Scholar] [CrossRef] [Green Version]
- Mavridou, E.; Brüggemann, R.J.; Melchers, W.J.; Mouton, J.W.; Verweij, P.E. Efficacy of posaconazole against three clinical Aspergillus fumigatus isolates with mutations in the cyp51A gene. Antimicrob. Agents Chemother. 2010, 54, 860–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinnollareddy, M.; Peake, S.L.; Roberts, M.S.; Lipman, J.; Roberts, J.A. Using pharmacokinetics and pharmacodynamics to optimise dosing of antifungal agents in critically ill patients: A systematic review. Int. J. Antimicrob. Agents. 2012, 39, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lutsar, I.; Hodges, M.R.; Tomaszewski, K.; Troke, P.F.; Wood, N.D. Safety of voriconazole and dose individualization. Clin. Infect. Dis. 2003, 36, 1087–1088. [Google Scholar] [CrossRef] [PubMed]
- Dolton, M.J.; Ray, J.E.; Chen, S.C.A.; Ng, K.; Pont, L.; McLahlan, A.J. Multicenter study of posaconazole therapeutic drug monitoring: Exposure-response relationship and factors affecting concentration. Antimicrob. Agents Chemother. 2012, 56, 5503–5510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.H.; Colangelo, P.M.; Gobburu, J.V.S. Exposure-response of posaconazole used for prophylaxis against invasive fungal infections: Evaluating the need to adjust doses based on drug concentrations in plasma. Clin. Pharmacol. Ther. 2010, 88, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, C.; Panzali, A.; Passi, A.; Borlenghi, E.; Lamorgese, C.; Petullà, M.; Re, A.; Caimi, L.; Rossi, G. Serum posaconazole levels during acute myeloid leukaemia induction therapy: Correlations with breakthrough invasive fungal infections. Mycoses 2015, 58, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Mitsani, D.; Nguyen, M.H.; Shields, R.K.; Toyoda, Y.; Kwak, E.J.; Silveira, F.P.; Pilewski, J.M.; Crespo, M.M.; Bermudez, C.; Bhama, J.K.; et al. Prospective, observational study of voriconazole therapeutic drug monitoring among lung transplant recipients receiving prophylaxis: Factors impacting levels of and associations between serum troughs, efficacy, and toxicity. Antimicrob. Agents Chemother. 2012, 56, 2371–2377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifilio, S.; Singhal, S.; Williams, S.; Frankfurt, O.; Gordon, L.; Evens, A.; Winter, J.; Talmann, M.; Pi, J.; Mehta, J. Breakthrough fungal infections after allogeneic hematopoietic stem cell transplantation in patients on prophylactic voriconazole. Bone Marrow Transplant. 2007, 40, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Kuo, S.F.; Wang, H.C.; Wu, C.J.; Lin, Y.S.; Li, W.S.; Lee, C.H. Azole resistance in Aspergillus species in Southern Taiwan: An epidemiological surveillance study. Mycoses 2019, 62, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Tsuchido, Y.; Tanaka, M.; Nakano, S.; Yamamoto, M.; Matsumura, Y.; Nagao, M. Prospective multicenter surveillance of clinically isolated Aspergillus species revealed azole-resistant Aspergillus fumigatus isolates with TR34/L98H mutation in the Kyoto and Shiga regions of Japan. Med. Mycol. 2019, 57, 997–1003. [Google Scholar] [CrossRef]
- Beer, K.D.; Farnon, E.C.; Jain, S.; Jamerson, C.; Lineberger, S.; Miller, J.; Berkow, E.L.; Lockhart, S.R.; Chiller, T.; Jackson, B.R. Multidrug-resistant Aspergillus fumigatus carrying mutations linked to environmental fungicide exposure—Three States, 2010–2017. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 1064–1067. [Google Scholar] [CrossRef] [Green Version]
- Resendiz Sharpe, A.; Lagrou, K.; Meis, J.F.; Chowdhary, A.; Lockhart, S.R.; Verweij, P.E.; ISHAM/ECMM Aspergillus Resistance Surveillance Working Group. Triazole resistance surveillance in Aspergillus fumigatus. Med. Mycol. 2018, 56 (Suppl. 1), 83–92. [Google Scholar] [CrossRef] [Green Version]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; De Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal agents in agriculture: Friends and foes of public health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snelders, E.; Camps, S.M.T.; Karawajczyk, A.; Schaftenaar, G.; Kema, G.H.J.; van der Lee, H.A.; Klaassen, C.H.; Melchers, W.J.G.; Verweij, P.E. Triazole fungicides can induce cross-resistance to medical triazoles in Aspergillus fumigatus. PLoS ONE 2012, 7, e31801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Paolo, M.; Dave, K.; Vijayasingam, A.; Sheth, R.; Luke, E.; Scourfield, A.; Nwankwo, L.; Schelenz, S.; Elborn, J.S.; Armstrong-James, D.; et al. Azoles therapeutic drug monitoring and fungal antimicrobial resistance in adults with cystic fibrosis. Eur. Respir. J. 2018, 52 (Suppl. 62), PA 3409. [Google Scholar]
- Seyedmousavi, S.; Meletiadis, J.; Melchers, W.J.; Rijs, A.J.M.M.; Mouton, J.W.; Verweij, P.E. In vitro interaction of voriconazole and anidulafungin against triazole-resistant Aspergillus fumigatus. Antimicrob. Agents Chemother. 2013, 57, 796–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavridou, E.; Bruggemann, R.J.; Melchers, W.J.; Verweij, P.E.; Mouton, J.W. Impact of cyp51A mutations on the pharmacokinetic and pharmacodynamic properties of voriconazole in a murine model of disseminated aspergillosis. Antimicrob. Agents Chemother. 2010, 54, 4758–4764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schauwvlieghe, A.F.A.D.; Buil, J.B.; Verweij, P.E.; Hoek, R.A.S.; Cornelissen, J.J.; Blijlevens, N.M.A.; Henriet, S.S.V.; Rijnders, B.J.A.; Brüggemann, R.J.M. High-dose posaconazole for azole-resistant aspergillosis and other difficult-to-treat mould infections. Mycoses 2020, 63, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Arastehfar, A.; Fang, W.; Pan, W.; Lackner, M.; Liao, W.; Badiee, P.; Zomorodian, K.; Badali, H.; Hagen, F.; Lass-Flörl, C.; et al. YEAST PANEL multiplex PCR for identification of clinically important yeast species: Stepwise diagnostic strategy, useful for developing countries. Diagn. Microbiol. Infect. Dis. 2019, 93, 112–119. [Google Scholar] [CrossRef]
- Arastehfar, A.; Daneshnia, F.; Farahyar, S.; Fang, W.; Salimi, M.; Salehi, M.; Hagen, F.; Weihua, P.; Roudbary, M.; Boekhout, T. Incidence and spectrum of yeast species isolated from the oral cavity of Iranian patients suffering from hematological malignancies. J. Oral Microbiol. 2019, 11, 1601061. [Google Scholar] [CrossRef] [Green Version]
- Arastehfar, A.; Daneshnia, F.; Kord, M.; Roudbary, M.; Zarrinfar, H.; Fang, W.; Hashemi, S.J.; Najafzadeh, M.J.; Khodavaisy, S.; Pan, W.; et al. Comparison of 21-Plex PCR and API 20C AUX, MALDI-TOF MS, and rDNA sequencing for a wide range of clinically isolated yeast species: Improved identification by combining 21-Plex PCR and API 20C AUX as an alternative strategy for developing countries. Front. Cell. Infect. Microbiol. 2019, 9, 21. [Google Scholar] [CrossRef]
- Clinical alert to U.S. Healthcare Facilities: Global Emergence of Invasive Infections Caused by the Multidrug-Resistant Yeast Candida auris. Atlanta: Centers for Disease Control and Prevention; September 2017; Clinical Update. Available online: https://www.cdc.gov/fungal/candida-auris/c-auris-alert-09-17.html (accessed on 21 December 2018).
- European Centre for Disease Prevention and Control. Candida auris in Healthcare Settings—Europe—First Update 23 April 2018. Stockholm: ECDC. 2018. Available online: https://ecdc.europa.eu/sites/portal/files/documents/RRA-Candida-auris-European-Union-countries-first-update.pdf (accessed on 23 April 2018).
- Chowdhary, A.; Sharma, C.; Meis, J.F. Candida auris: A rapidly emerging cause of hospital-acquired multidrug-resistant fungal infections globally. PLoS Pathog. 2017, 13, e1006290. [Google Scholar] [CrossRef]
- Aruanno, M.; Glampedakis, E.; Lamoth, F. Echinocandins for the treatment of invasive aspergillosis: From laboratory to bedside. Antimicrob. Agents Chemother. 2019, 63, e00399-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiederhold, N.P.; Locke, J.B.; Daruwala, P.; Bartizal, K. Rezafungin (CD101) demonstrates potent in vitro activity against Aspergillus, including azole-resistant Aspergillus fumigatus isolates and cryptic species. J. Antimicrob. Chemother. 2018, 73, 3063–3067. [Google Scholar] [CrossRef]
- Cornely, O.A.; Hoenigl, M.; Lass-Flörl, C.; Chen, S.C.A.; Kontoyiannis, D.P.; Morrissey, C.O.; Thompson, G.R., 3rd; Mycoses Study Group Education and Research Consortium (MSG-ERC) and the European Confederation of Medical Mycology (ECMM). Defining breakthrough invasive fungal infection–Position paper of the mycoses study group education and research consortium and the European Confederation of Medical Mycology. Mycoses 2019, 62, 716–729. [Google Scholar] [PubMed]
- Jenks, J.D.; Mehta, S.R.; Hoenigl, M. Broad spectrum triazoles for invasive mould infections in adults: Which drug and when? Med. Mycol. 2019, 57 (Suppl. 2), S168–S178. [Google Scholar] [CrossRef]
- Verweij, P.E.; Chowdhary, A.; Melchers, W.J.; Meis, J.F. Azole resistance in Aspergillus fumigatus: Can we retain the clinical use of mold-active antifungal azoles? Clin. Infect. Dis. 2016, 62, 362–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schauwvlieghe, A.F.A.D.; de Jonge, N.; van Dijk, K.; Verweij, P.E.; Brüggemann, R.J.; Biemond, B.J.; Bart, A.; von dem Borne, P.A.; Verbon, A.; van der Beek, M.T.; et al. The diagnosis and treatment of invasive aspergillosis in Dutch haematology units facing a rapidly increasing prevalence of azole-resistance. A nationwide survey and rationale for the DB-MSG 002 study protocol. Mycoses 2018, 61, 656–664. [Google Scholar] [CrossRef]
- Zurl, C.; Waller, M.; Schwameis, F.; Muhr, T.; Bauer, N.; Zollner-Schwetz, I.; Valentin, T.; Meinitzer, A.; Ullrich, E.; Wunsch, S.; et al. Isavuconazole treatment in a mixed patient cohort with invasive fungal infections: Outcome, tolerability and clinical implications of isavuconazole plasma concentrations. J. Fungi 2020, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Jenks, J.D.; Seidel, D.; Cornely, O.A.; Chen, S.; van Hal, S.; Kauffman, C.; Miceli, M.H.; Heinemann, M.; Christner, M.; Jover-Sáenz, A.; et al. Voriconazole plus terbinafine combination antifungal therapy for invasive Lomentospora prolificans infections: Analysis of 41 patients from the FungiScope® registry 2008–2019. Clin. Microbiol. Infect. 2020, 26, 784.e1–784.e5. [Google Scholar] [CrossRef]
- Tsitsopoulou, A.; Posso, R.; Vale, L.; Bebb, S.; Johnson, E.; White, P.L. Determination of the prevalence of triazole resistance in environmental Aspergillus fumigatus strains isolated in South Wales, UK. Front. Microbiol. 2018, 9, 1395. [Google Scholar] [CrossRef]
- Zoran, T.; Sartori, B.; Sappl, L.; Aigner, M.; Sánchez-Reus, F.; Rezusta, A.; Chowdhary, A.; Taj-Aldeen, S.J.; Arendrup, M.C.; Oliveri, S.; et al. Azole-resistance in Aspergillus terreus and related species: An emerging problem or a rare phenomenon? Front. Microbiol. 2018, 9, 516. [Google Scholar] [CrossRef]
- Kupferschmidt, K. New drugs target growing threat of fatal fungi. Science 2019, 366, 407. [Google Scholar] [CrossRef] [PubMed]
- Arastehfar, A.; Carvalho, A.; van de Veerdonk, F.L.; Jenks, J.D.; Koehler, P.; Krause, R.; Cornely, O.A.; Perlin, D.S.; Lass-Flörl, C.; Hoenigl, M. COVID-19 associated pulmonary aspergillosis (CAPA)–From immunology to treatment. J. Fungi 2020, 6, 91. [Google Scholar] [CrossRef]
- Siles, S.A.; Srinivasan, A.; Pierce, C.G.; Lopez-Ribot, J.L.; Ramasubramanian, A.K. High-throughput screening of a collection of known pharmacologically active small compounds for identification of Candida albicans biofilm inhibitors. Antimicrob. Agents Chemother. 2013, 57, 3681–3687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Zilbermintz, L.; Martchenko, M. Repurposing FDA approved drugs against the human fungal pathogen, Candida albicans. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousfi, H.; Ranque, S.; Cassagne, C.; Rolain, J.M.; Bittar, F. Identification of repositionable drugs with novel antimycotic activity by screening the Prestwick Chemical Library against emerging invasive moulds. J. Glob. Antimicrob. Resist. 2020, 21, 314–317. [Google Scholar] [CrossRef] [PubMed]
- Wall, G.; Chaturvedi, A.K.; Wormley, F.L., Jr.; Wiederhold, N.P.; Patterson, H.P.; Patterson, T.F.; Lopez-Ribot, J.L. Screening a repurposing library for inhibitors of multidrug-resistant Candida auris identifies ebselen as a repositionable candidate for antifungal drug development. Antimicrob. Agents Chemother. 2018, 62, e01084-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Species | Prevalence (%) | Azole Resistance Rate (%) | Echinocandin Resistance Rate (%) | References |
|---|---|---|---|---|
| C. albicans | >20.9–70 | 0–7.8 | 0–7 | [43,44,46,48,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87] |
| C. glabrata | <15 | 0–21 | 0–23.1 | [48,62,67,69,70,74,75,76,78,84] |
| 15–38 | 0–76 | 0–100 | [43,44,46,63,66,68,71,73,77,79,80,81,82,83,85,86,87] | |
| C. tropicalis | <10 | 0.6–31.5 | 0–8 | [62,63,64,65,66,67,69,70,71,82,83] |
| 10–49 | 0–66.6 | 0–10 | [43,44,46,48,68,72,73,74,75,76,77,78,79,80,81,84,85,86,87] | |
| C. parapsilosis | <15 | 0–8.3 | 0–10.8 | [46,62,63,77,78,79,80,81,82,85] |
| 15–37 | 0–53 | 0–1.6 | [43,44,48,65,71,73,74,75,76,83,84,86,87] | |
| C. auris | 0–14 | >90–100 | 0–40 | [38,62,88,89,90,91] |
| A. fumigatus | 33.2–92 | <2–30 | 0 | [92,93,94,95,96,97,98,99,100,101,102] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arastehfar, A.; Gabaldón, T.; Garcia-Rubio, R.; Jenks, J.D.; Hoenigl, M.; Salzer, H.J.F.; Ilkit, M.; Lass-Flörl, C.; Perlin, D.S. Drug-Resistant Fungi: An Emerging Challenge Threatening Our Limited Antifungal Armamentarium. Antibiotics 2020, 9, 877. https://doi.org/10.3390/antibiotics9120877
Arastehfar A, Gabaldón T, Garcia-Rubio R, Jenks JD, Hoenigl M, Salzer HJF, Ilkit M, Lass-Flörl C, Perlin DS. Drug-Resistant Fungi: An Emerging Challenge Threatening Our Limited Antifungal Armamentarium. Antibiotics. 2020; 9(12):877. https://doi.org/10.3390/antibiotics9120877
Chicago/Turabian StyleArastehfar, Amir, Toni Gabaldón, Rocio Garcia-Rubio, Jeffrey D. Jenks, Martin Hoenigl, Helmut J. F. Salzer, Macit Ilkit, Cornelia Lass-Flörl, and David S. Perlin. 2020. "Drug-Resistant Fungi: An Emerging Challenge Threatening Our Limited Antifungal Armamentarium" Antibiotics 9, no. 12: 877. https://doi.org/10.3390/antibiotics9120877