Overexpression of α-Synuclein by Oligodendrocytes in Transgenic Mice Does Not Recapitulate the Fibrillar Aggregation Seen in Multiple System Atrophy

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Primary Culture of Cortical Neurons and High Content Analysis
2.3. Primary Culture of Mouse Oligodendrocytes
2.4. Recombinant α-Syn Expression and Purification
2.5. α-syn Fibrillization
2.6. Immunofluorescence
2.7. Brain Sections Staining
2.8. SarkoSpin Fractionation
2.9. Sedimentation Velocity Gradient Fractionation
2.10. Analysis of the Protein Contents of Sarkospin and Velocity Fractions by Filter Trap
2.11. Crosslinking and Western Blot
2.12. Antibodies
3. Results and Discussion
3.1. S129-phosphorylated α-syn Species Found in the Brain of PLP-αSyn Mice Are Distinct from the Amyloid forms Extracted from PD and MSA Brains and from Recombinant PFFs
3.2. S129-phosphorylated Human α-syn Found in PLP-αSyn Mouse Brains Corresponds to Soluble Monomeric and Oligomeric Forms
3.3. Is the Presence of Neuronal Human α-syn in PLP-αSyn Primary Cortical Cultures due to an OL-to-Neuron Transfer of the Protein?
3.4. S129-Phosphorylated Human α-Syn Is Enriched at “Hot Spots” in OL Processes
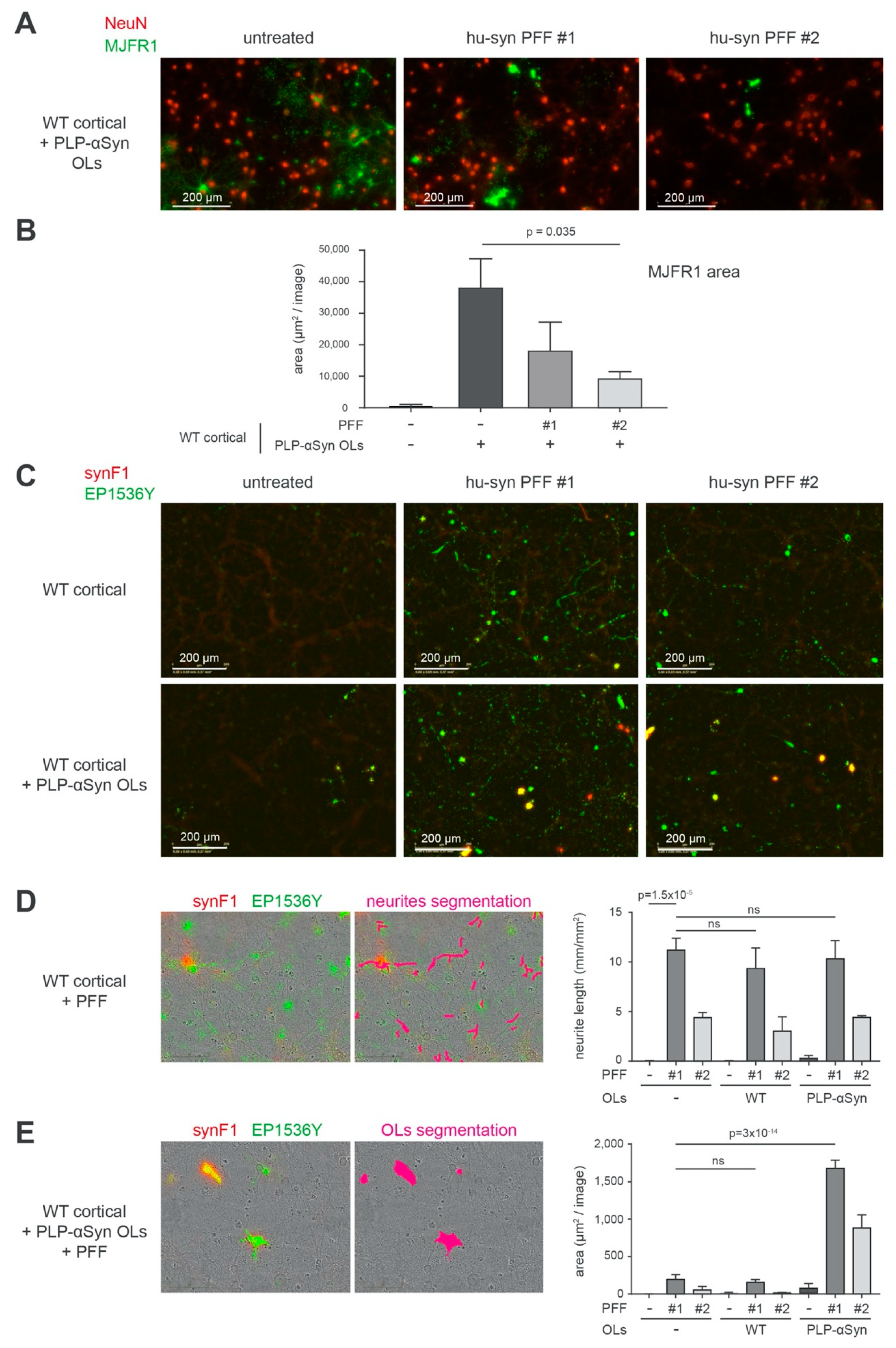
3.5. Aggregation of Human α-Syn Does Not Spontaneously Take Place on OLs from PLP-αSyn Mice but Can Be Experimentally Seeded with PFFs
3.6. Considerations about the Face vs. Predictive Value of the PLP-αSyn Mouse Model
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Papp, M.I.; Kahn, J.E.; Lantos, P.L. Glial cytoplasmic inclusions in the CNS of patients with multiple system atrophy (striatonigral degeneration, olivopontocerebellar atrophy and Shy-Drager syndrome). J. Neurol. Sci. 1989, 94, 79–100. [Google Scholar] [CrossRef]
- Papp, M.I.; Lantos, P.L. The distribution of oligodendroglial inclusions in multiple system atrophy and its relevance to clinical symptomatology. Brain 1994, 117, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Spillantini, M.G.; Crowther, R.A.; Jakes, R.; Cairns, N.J.; Lantos, P.L.; Goedert, M. Filamentous alpha-synuclein inclusions link multiple system atrophy with Parkinson’s disease and dementia with Lewy bodies. Neurosci. Lett. 1998, 251, 205–208. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Yoshimoto, M.; Tsuji, S.; Takahashi, H. A-Synuclein Immunoreactivity in Glial Cytoplasmic Inclusions in Multiple System Atrophy. Neurosci. Lett. 1998, 249, 180–182. [Google Scholar] [CrossRef]
- Meissner, W.G.; Fernagut, P.O.; Dehay, B.; Péran, P.; Le Traon, A.P.; Foubert-Samier, A.; Lopez Cuina, M.; Bezard, E.; Tison, F.; Rascol, O. Multiple System Atrophy: Recent Developments and Future Perspectives. Mov. Disord. 2019, 34, 1629–1642. [Google Scholar] [CrossRef]
- Djelloul, M.; Holmqvist, S.; Boza-Serrano, A.; Azevedo, C.; Yeung, M.S.; Goldwurm, S.; Frisén, J.; Deierborg, T.; Roybon, L. Alpha-Synuclein Expression in the Oligodendrocyte Lineage: An in Vitro and in Vivo Study Using Rodent and Human Models. Stem Cell Rep. 2015, 5, 174–184. [Google Scholar] [CrossRef] [Green Version]
- Asi, Y.T.; Simpson, J.E.; Heath, P.R.; Wharton, S.B.; Lees, A.J.; Revesz, T.; Houlden, H.; Holton, J.L. Alpha-synuclein mRNA expression in oligodendrocytes in MSA. Glia 2014, 62, 964–970. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.J.C.; Lundvig, D.M.S.; Huang, Y.; Wei, P.G.; Blumbergs, P.C.; Højrup, P.; Otzen, D.; Halliday, G.M.; Jensen, P.H. P25A Relocalizes in Oligodendroglia From Myelin To Cytoplasmic Inclusions in Multiple System Atrophy. Am. J. Pathol. 2007, 171, 1291–1303. [Google Scholar] [CrossRef] [Green Version]
- Mavroeidi, P.; Arvanitaki, F.; Karakitsou, A.-K.; Vetsi, M.; Kloukina, I.; Zweckstetter, M.; Giller, K.; Becker, S.; Sorrentino, Z.A.; Giasson, B.I.; et al. Endogenous oligodendroglial alpha-synuclein and TPPP/p25α orchestrate alpha-synuclein pathology in experimental multiple system atrophy models. Acta Neuropathol. 2019, 138, 415–441. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Shi, M.; Stewart, T.; Fernagut, P.-O.; Huang, Y.; Tian, C.; Dehay, B.; Atik, A.; Yang, D.; De Giorgi, F.; et al. Reduced oligodendrocyte exosome secretion in multiple system atrophy involves SNARE dysfunction. Brain 2020, 143, 1–18. [Google Scholar] [CrossRef]
- Hasegawa, T.; Kikuchi, A.; Takeda, A. Pathogenesis of multiple system atrophy. Neurol. Clin. Neurosci. 2013, 1, 189–194. [Google Scholar] [CrossRef]
- Reyes, J.F.; Rey, N.L.; Bousset, L.; Melki, R.; Brundin, P.; Angot, E. Alpha-synuclein transfers from neurons to oligodendrocytes. Glia 2014, 62, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, H.; Kowa, H.; Koga, H.; Takata, M.; Satake, W.; Futamura, N.; Funakawa, I.; Jinnai, K.; Takahashi, M.; Kondo, T.; et al. Wide distribution of alpha-synuclein oligomers in multiple system atrophy brain detected by proximity ligation. Acta Neuropathol. 2019, 137, 455–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahle, P.J.; Neumann, M.; Ozmen, L.; Müller, V.; Jacobsen, H.; Spooren, W.; Fuss, B.; Mallon, B.; Macklin, W.B.; Fujiwara, H.; et al. Hyperphosphorylation and insolubility of α-synuclein in transgenic mouse oligodendrocytes. EMBO Rep. 2002, 3, 583–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shults, C.W.; Rockenstein, E.; Crews, L.; Adame, A.; Mante, M.; Larrea, G.; Hashimoto, M.; Song, D.; Iwatsubo, T.; Tsuboi, K.; et al. Neurological and neurodegenerative alterations in a transgenic mouse model expressing human α-synuclein under oligodendrocyte promoter: Implications for multiple system atrophy. J. Neurosci. 2005, 25, 10689–10699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazawa, I.; Giasson, B.I.; Sasaki, R.; Zhang, B.; Joyce, S.; Uryu, K.; Trojanowski, J.Q.; Lee, V.M.Y. Mouse model of multiple system atrophy α-synuclein expression in oligodendrocytes causes glial and neuronal degeneration. Neuron 2005, 45, 847–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heras-Garvin, A.; Weckbecker, D.; Ryazanov, S.; Leonov, A.; Griesinger, C.; Giese, A.; Wenning, G.K.; Stefanova, N. Anle138b modulates α-synuclein oligomerization and prevents motor decline and neurodegeneration in a mouse model of multiple system atrophy. Mov. Disord. 2019, 34, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Bassil, F.; Fernagut, P.-O.; Bezard, E.; Pruvost, A.; Leste-Lasserre, T.; Hoang, Q.Q.; Ringe, D.; Petsko, G.A.; Meissner, W.G. Reducing C-terminal truncation mitigates synucleinopathy and neurodegeneration in a transgenic model of multiple system atrophy. Proc. Natl. Acad. Sci. USA 2016, 113, 9593–9598. [Google Scholar] [CrossRef] [Green Version]
- Venezia, S.; Refolo, V.; Polissidis, A.; Stefanis, L.; Wenning, G.K.; Stefanova, N. Toll-like receptor 4 stimulation with monophosphoryl lipid A ameliorates motor deficits and nigral neurodegeneration triggered by extraneuronal α-synucleinopathy. Mol. Neurodegener. 2017, 12, 1–13. [Google Scholar] [CrossRef]
- Bassil, F.; Canron, M.H.; Vital, A.; Bezard, E.; Li, Y.; Greig, N.H.; Gulyani, S.; Kapogiannis, D.; Fernagut, P.O.; Meissner, W.G. Insulin resistance and exendin-4 treatment for multiple system atrophy. Brain 2017, 140, 1420–1436. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Cuina, M.; Guerin, P.A.; Canron, M.H.; Delamarre, A.; Dehay, B.; Bezard, E.; Meissner, W.G.; Fernagut, P.O. Nilotinib Fails to Prevent Synucleinopathy and Cell Loss in a Mouse Model of Multiple System Atrophy. Mov. Disord. 2020, 35, 1–11. [Google Scholar] [CrossRef]
- Fujiwara, H.; Hasegawa, M.; Dohmae, N.; Kawashima, A.; Masliah, E.; Goldberg, M.S.; Shen, J.; Takio, K.; Iwatsubo, T. alpha-Synuclein is phosphorylated in synucleinopathy lesions. Nat. Cell Biol. 2002, 4, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Fujiwara, H.; Nonaka, T.; Wakabayashi, K.; Takahashi, H.; Lee, V.M.Y.; Trojanowski, J.Q.; Mann, D.; Iwatsubo, T. Phosphorylated α-synuclein is ubiquitinated in α-synucleinopathy lesions. J. Biol. Chem. 2002, 277, 49071–49076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bleasel, J.M.; Halliday, G.M.; Kim, W.S. Animal modeling an oligodendrogliopathy--multiple system atrophy. Acta Neuropathol. Commun. 2016, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernagut, P.O.; Meissner, W.G.; Biran, M.; Fantin, M.; Bassil, F.; Franconi, J.M.; Tison, F. Age-related motor dysfunction and neuropathology in a transgenic mouse model of multiple system atrophy. Synapse 2014, 68, 98–106. [Google Scholar] [CrossRef]
- Stefanova, N.; Reindl, M.; Neumann, M.; Haass, C.; Poewe, W.; Kahle, P.J.; Wenning, G.K. Oxidative stress in transgenic mice with oligodendroglial α-synuclein overexpression replicates the characteristic neuropathology of multiple system atrophy. Am. J. Pathol. 2005, 166, 869–876. [Google Scholar] [CrossRef]
- Stemberger, S.; Poewe, W.; Wenning, G.K.; Stefanova, N. Targeted overexpression of human α-synuclein in oligodendroglia induces lesions linked to MSA -like progressive autonomic failure. Exp. Neurol. 2010, 224, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Kuzdas, D.; Stemberger, S.; Gaburro, S.; Stefanova, N.; Singewald, N.; Wenning, G.K. Oligodendroglial alpha-synucleinopathy and MSA-like cardiovascular autonomic failure: Experimental evidence. Exp. Neurol. 2013, 247, 531–536. [Google Scholar] [CrossRef] [Green Version]
- Boudes, M.; Uvin, P.; Pinto, S.; Voets, T.; Fowler, C.J.; Wenning, G.K.; De Ridder, D.; Stefanova, N. Bladder dysfunction in a transgenic mouse model of multiple system atrophy. Mov. Disord. 2013, 28, 347–355. [Google Scholar] [CrossRef] [Green Version]
- De Giorgi, F.; Laferrière, F.; Zinghirino, F.; Faggiani, E.; Lends, A.; Bertoni, M.; Yu, X.; Grélard, A.; Morvan, E.; Habenstein, B.; et al. Novel self-replicating α-synuclein polymorphs that escape ThT monitoring can spontaneously emerge and acutely spread in neurons. Sci. Adv. 2020, 6, eabc4364. [Google Scholar] [CrossRef]
- Laferrière, F.; Maniecka, Z.; Pérez-Berlanga, M.; Hruska-Plochan, M.; Gilhespy, L.; Hock, E.-M.; Wagner, U.; Afroz, T.; Boersema, P.J.; Barmettler, G.; et al. TDP-43 extracted from frontotemporal lobar degeneration subject brains displays distinct aggregate assemblies and neurotoxic effects reflecting disease progression rates. Nat. Neurosci. 2018, 22, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Berlanga, M.; Laferrière, F.; Polymenidou, M. SarkoSpin: A Technique for Biochemical Isolation and Characterization of Pathological TDP-43 Aggregates. Bio-Protocol 2019, 9, 1–12. [Google Scholar]
- Laferrière, F.; Tixador, P.; Moudjou, M.; Chapuis, J.; Sibille, P.; Herzog, L.; Reine, F.; Jaumain, E.; Laude, H.; Rezaei, H.; et al. Quaternary structure of pathological prion protein as a determining factor of strain-specific prion replication dynamics. PLoS Pathog 2013, 9, e1003702. [Google Scholar] [CrossRef] [Green Version]
- Refolo, V.; Bez, F.; Polissidis, A.; Kuzdas-Wood, D.; Sturm, E.; Kamaratou, M.; Poewe, W.; Stefanis, L.; Angela Cenci, M.; Romero-Ramos, M.; et al. Progressive striatonigral degeneration in a transgenic mouse model of multiple system atrophy: Translational implications for interventional therapies. Acta Neuropathol. Commun. 2018, 6, 2. [Google Scholar] [CrossRef]
- Schweighauser, M.; Shi, Y.; Tarutani, A.; Kametani, F.; Murzin, A.G.; Ghetti, B.; Matsubara, T.; Tomita, T.; Ando, T.; Hasegawa, K.; et al. Structures of α-synuclein filaments from multiple system atrophy. Nature 2020, 585, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Lashuel, H.A. Do Lewy bodies contain alpha-synuclein fibrils? and Does it matter? A brief history and critical analysis of recent reports. Neurobiol. Dis. 2020, 141, 104876. [Google Scholar]
- Flabeau, O.; Meissner, W.G.; Ozier, A.; Berger, P.; Tison, F.; Fernagut, P.O. Breathing variability and brainstem serotonergic loss in a genetic model of multiple system atrophy. Mov. Disord. 2014, 29, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.R.; Matsuo, Y.; Cashikar, A.G.; Kamitani, T. Role of ser129 phosphorylation of α-synuclein in melanoma cells. J. Cell Sci. 2013, 126, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Volles, M.J.; Lansbury, P.T., Jr. Relationships between the sequence of alpha-synuclein and its membrane affinity, fibrillization propensity and yeast toxicity. J. Mol. Biol. 2007, 366, 1510–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaikath, N.N.; Hmila, I.; Gupta, V.; Erskine, D.; Ingelsson, M.; El-Agnaf, O.M.A. Antibodies against alpha-synuclein: Tools and therapies. J. Neurochem. 2019, 150, 612–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uemura, N.; Uemura, M.T.; Lo, A.; Bassil, F.; Zhang, B.; Luk, K.C.; Lee, V.M.-Y.; Takahashi, R.; Trojanowski, J.Q. Slow progressive accumulation of oligodendroglial alpha-synuclein (α-syn) pathology in synthetic α-syn fibril-induced mouse models of synucleinopathy. J. Neuropathol. Exp. Neurol. 2019, 78, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Shahmoradian, S.H.; Lewis, A.J.; Genoud, C. Lewy pathology in Parkinson’s disease consists of crowded organelles and lipid membranes. Nat. Neurosci. 2019, 22, 1099–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Target | Company | Cat.No | Dilution IF | Dilution IB |
|---|---|---|---|---|---|
| Primary antibodies | |||||
| MJFR-1 | human alpha-synuclein | Abcam | ab138501 | 1: 1000 | 1: 10,000 |
| EP1536Y | pS129 phospho-synuclein | Abcam | ab51253 | 1: 500 | 1: 5000 |
| Syn1 clone 42 | human and murine alpha-synuclein | BD Biosciences | 610787 | 1: 500 | 1: 2000 |
| D37A6 | murine alpha-synuclein | Cell Signaling | #4179 | 1: 200 | 1: 2000 |
| SynF1 | aggregated alpha-synuclein | BioLegend | 847802 | 1: 500 | 1: 10,000 |
| syn211 | human alpha-synuclein | Abcam | ab80627 | 1: 500 | n/a |
| MBP | myelin basic protein | Abcam | ab218011 | 1: 200 | 1: 2000 |
| CNPase | cyclic nucleotide phospho. | Abcam | ab6319 | 1: 500 | n/a |
| Sox10 | sox10 protein | Abcam | ab155279 | 1: 200 | n/a |
| NeuN | neuronal nuclei protein | Merck Millipore | MAB377 | 1: 500 | n/a |
| Actin | beta-actin | Sigma | A5316 | n/a | 1: 10,000 |
| Secondary antibodies | |||||
| Goat anti-mouse HRP | mouse IgG (H + L) | Jackson Immuno | 115-035-146 | n/a | 1: 10,000 |
| Goat anti-rabbit HRP | rabbit IgG (H + L) | Jackson Immuno | 111-035-144 | n/a | 1: 10,000 |
| Goat anti-mouse IRDye 680RD | mouse IgG (H + L) | LI-COR | 926-68070 | n/a | 1: 5000 |
| Goat anti-rabbit IRDye 800CW | rabbit IgG (H + L) | LI-COR | 926-32211 | n/a | 1: 5000 |
| Donkey anti-mouse Alexa 488 | mouse IgG (H + L) | Thermo Fisher Sci. | A-21202 | 1: 500 | n/a |
| Goat anti-rabbit Alexa 488 | rabbit IgG (H + L) | Thermo Fisher Sci. | A-11008 | 1: 500 | n/a |
| Donkey anti-mouse Alexa 594 | mouse IgG (H + L) | Thermo Fisher Sci. | A-21203 | 1: 500 | n/a |
| Donkey anti-rabbit Alexa 594 | rabbit IgG (H + L) | Thermo Fisher Sci. | A-21207 | 1: 500 | n/a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laferrière, F.; He, X.; Zinghirino, F.; Doudnikoff, E.; Faggiani, E.; Meissner, W.G.; Bezard, E.; De Giorgi, F.; Ichas, F. Overexpression of α-Synuclein by Oligodendrocytes in Transgenic Mice Does Not Recapitulate the Fibrillar Aggregation Seen in Multiple System Atrophy. Cells 2020, 9, 2371. https://doi.org/10.3390/cells9112371
Laferrière F, He X, Zinghirino F, Doudnikoff E, Faggiani E, Meissner WG, Bezard E, De Giorgi F, Ichas F. Overexpression of α-Synuclein by Oligodendrocytes in Transgenic Mice Does Not Recapitulate the Fibrillar Aggregation Seen in Multiple System Atrophy. Cells. 2020; 9(11):2371. https://doi.org/10.3390/cells9112371
Chicago/Turabian StyleLaferrière, Florent, Xin He, Federica Zinghirino, Evelyne Doudnikoff, Emilie Faggiani, Wassilios G. Meissner, Erwan Bezard, Francesca De Giorgi, and François Ichas. 2020. "Overexpression of α-Synuclein by Oligodendrocytes in Transgenic Mice Does Not Recapitulate the Fibrillar Aggregation Seen in Multiple System Atrophy" Cells 9, no. 11: 2371. https://doi.org/10.3390/cells9112371