Suppression of Schistosoma japonicum Acetylcholinesterase Affects Parasite Growth and Development

Abstract
:1. Introduction
2. Results
2.1. RNAi-Induced Knockdown of SjAChE
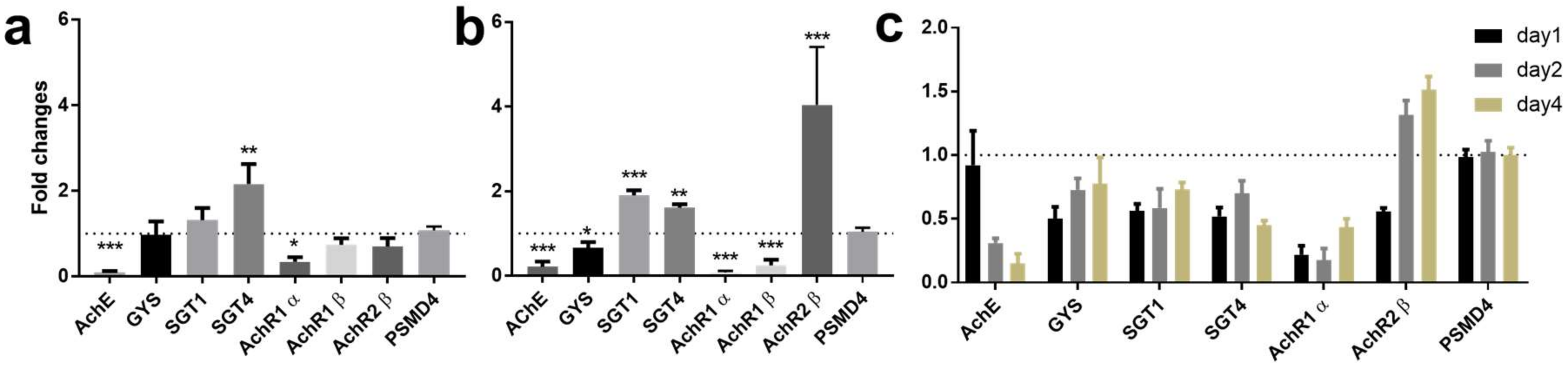
2.1.1. RNAi-Induced Knockdown of SjAChE Regulated Transcription of Nicotinic Acetylcholine Receptors and Genes Involved in Glucose Uptake
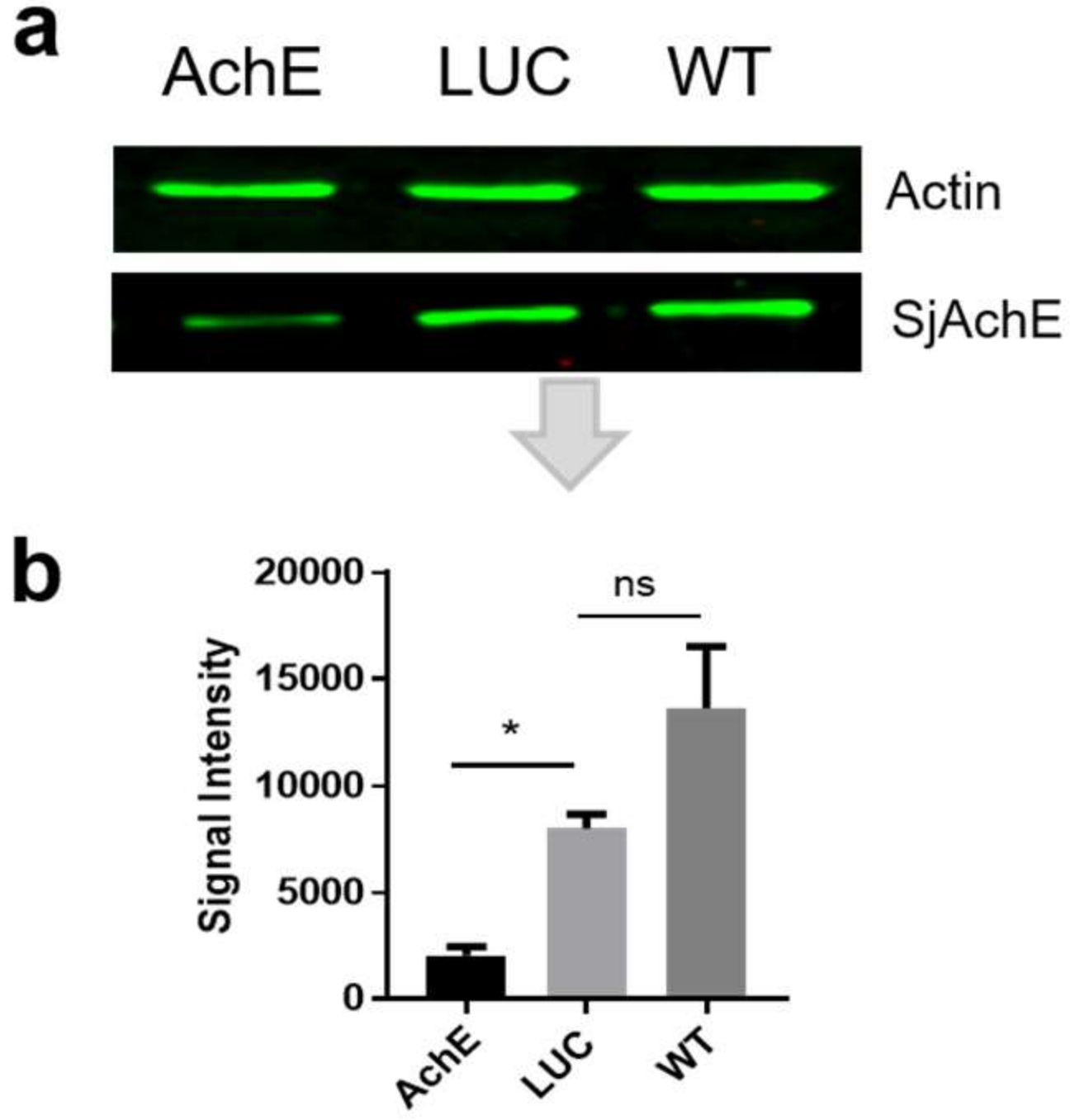
2.1.2. Reduction in SjAChE Protein Expression in Parasites Treated with SjAChE dsRNA
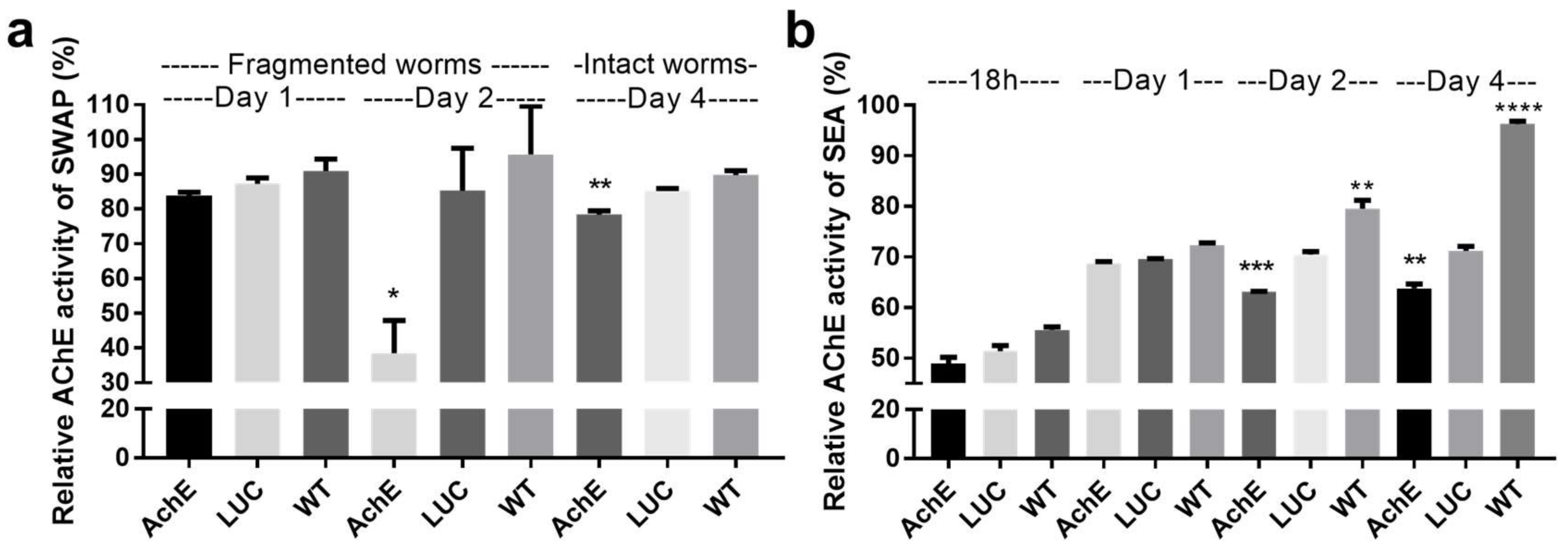
2.1.3. Effect of SjAChE Gene Suppression on AChE Activity of Adults and Eggs Treated with SjAChE dsRNA
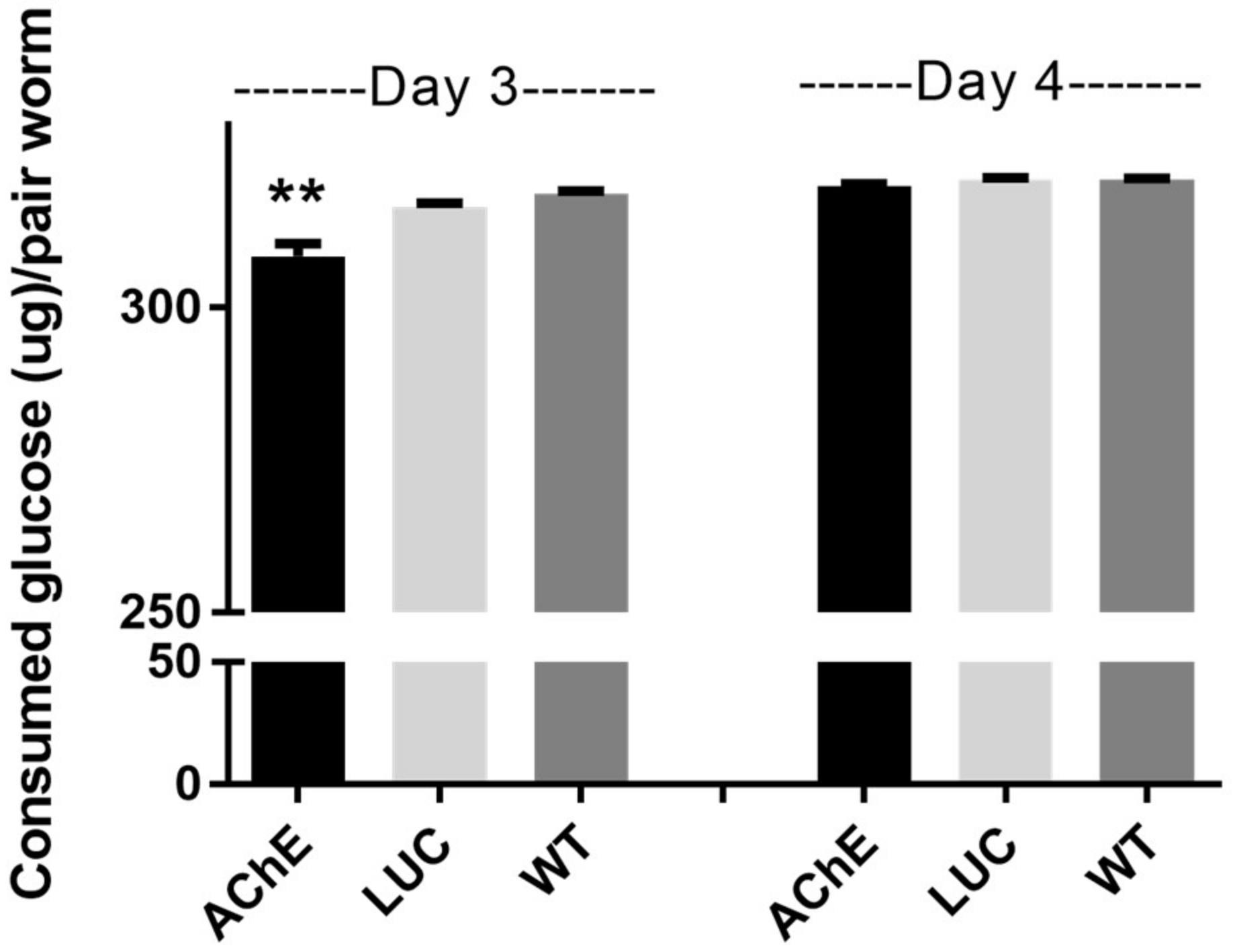
2.1.4. Effect of SjAChE Gene Suppression on Glucose Uptake by Adults
2.2. Immunolocalization of SjAChE in Eggs Trapped in Liver
2.3. Vaccine Efficacy of the SjAChE Protein Induced in Inbred CBA Mice
2.3.1. Worm and Egg Reduction in rSjAChE Vaccinated Mice after Cercarial Challenge
2.3.2. Antibody Responses in Vaccinated Mice
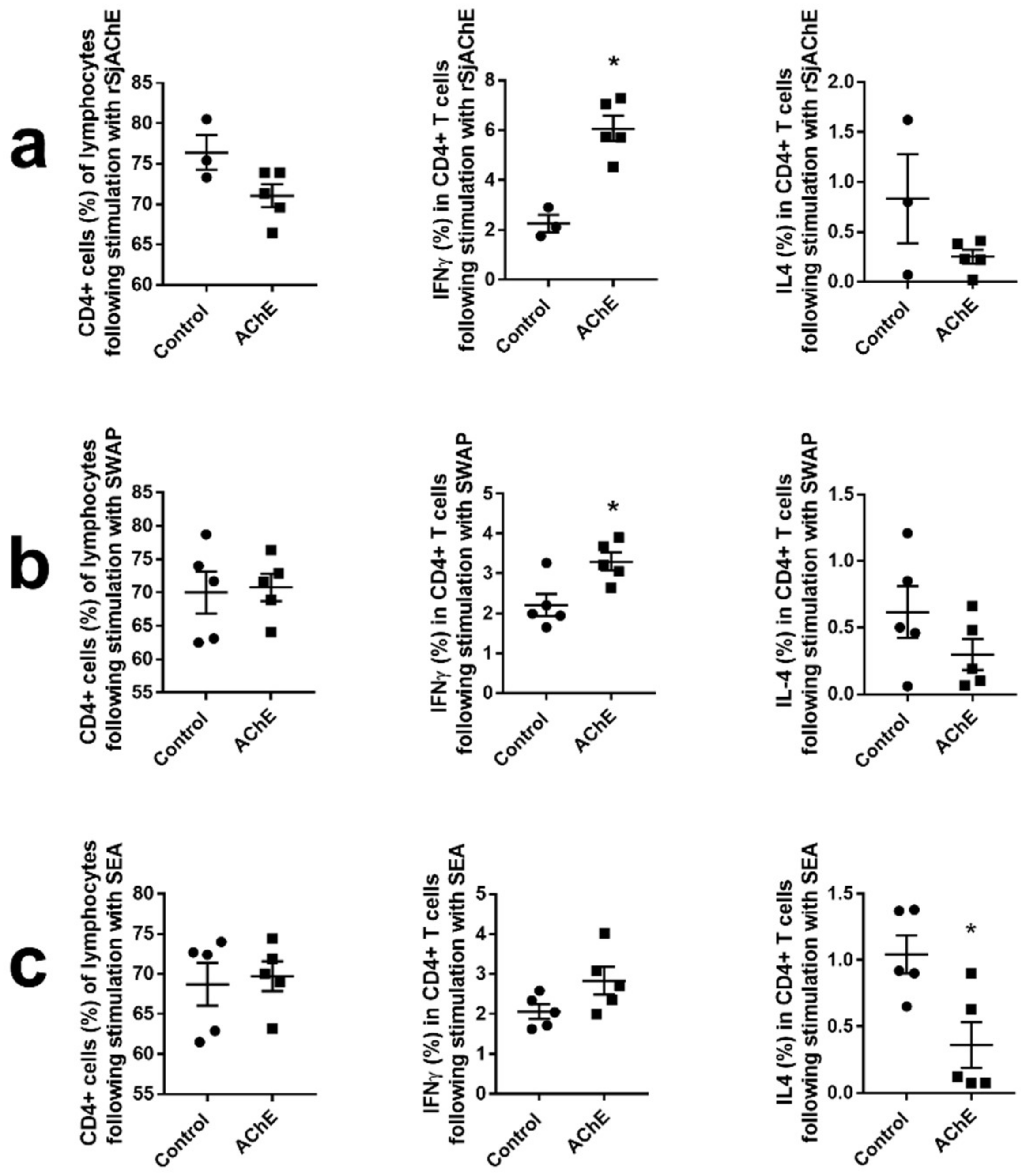
2.3.3. Cytokine Analysis of Vaccinated Mice
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Parasites
4.3. Treatment of Parasites with Double Stranded RNA (dsRNA)
4.4. Glucose Measurement and AChE Activity Assays
4.5. Expression Analysis of SjAChE and Related Genes
4.6. Western-Blot Analysis
4.7. Immunolocalization of SjAChE in Eggs
4.8. Protective Efficacy of the Recombinant SjAChE Vaccine
4.8.1. Recombinant Protein Expression and Purification
4.8.2. Mice Vaccination
4.8.3. ELISA
4.8.4. Worm and Egg Counting and Pathology
4.8.5. Flow Cytometry Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beaumier, C.M.; Gillespie, P.M.; Hotez, P.J.; Bottazzi, M.E. New vaccines for neglected parasitic diseases and dengue. Transl. Res. 2013, 162, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.P.; Martin, R.J. Ion-channels on parasite muscle: Pharmacology and physiology. Invertebr. Neurosci. 2007, 7, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Kaminsky, R.; Gauvry, N.; Schorderet Weber, S.; Skripsky, T.; Bouvier, J.; Wenger, A.; Schroeder, F.; Desaules, Y.; Hotz, R.; Goebel, T.; et al. Identification of the amino-acetonitrile derivative monepantel (AAD 1566) as a new anthelmintic drug development candidate. Parasitol. Res. 2008, 103, 931–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bueding, E.; Liu, C.L.; Rogers, S.H. Inhibition by metrifonate and dichlorvos of cholinesterases in schistosomes. Br. J. Pharmacol. 1972, 46, 480–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Ridi, R.A.; Tallima, H.A. Novel therapeutic and prevention approaches for schistosomiasis: Review. J. Adv. Res. 2013, 4, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Bergquist, R. Strategies for control of infection and disease: Current practice and future potential. In Schistosomiasis; Mahmoud, A.A.F., Ed.; Imperial College Press: London, UK, 2001. [Google Scholar]
- Salafsky, B.; Fusco, A.C.; Whitley, K.; Nowicki, D.; Ellenberger, B. Schistosoma mansoni: Analysis of cercarial transformation methods. Exp. Parasitol. 1988, 67, 116–127. [Google Scholar] [CrossRef]
- Barker, L.R.; Bueding, E.; Timms, A.R. The possible role of acetylcholine in Schistosoma mansoni. Br. J. Pharmacol. 1966, 26, 656–665. [Google Scholar] [CrossRef]
- Pax, R.A.; Day, T.A.; Miller, C.L.; Bennett, J.L. Neuromuscular physiology and pharmacology of parasitic flatworms. Parasitology 1996, 113, S83–S96. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, B.; Tarrab-Hazdai, R.; Himmeloch, S.; Arnon, R. Acetylcholinesterase from Schistosoma mansoni: Immunological characterization. Immunol. Lett. 1991, 28, 167–174. [Google Scholar] [CrossRef]
- Crabtree, J.E.; Wilson, R.A. Schistosoma mansoni: A scanning electron microscope study of the developing schistosomulum. Parasitology 1980, 81, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.K.; Bentley, G.N.; Oliveros Parra, W.G.; Agnew, A. Molecular characterization of an acetylcholinesterase implicated in the regulation of glucose scavenging by the parasite Schistosoma. FASEB J. 2002, 16, 441–443. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Lester, H.A.; Lummis, S.C. The structural basis of function in Cys-loop receptors. Q. Rev. Biophys. 2010, 43, 449–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentley, G.N.; Jones, A.K.; Agnew, A. Expression and comparative functional characterisation of recombinant acetylcholinesterase from three species of Schistosoma. Mol. Biochem. Parasitol. 2005, 141, 119–123. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Gobert, G.N.; Du, X.; Pali, G.; Cai, P.; Jones, M.K.; McManus, D.P. Functional characterisation of Schistosoma japonicum acetylcholinesterase. Parasites Vectors 2016, 9, 328. [Google Scholar] [CrossRef] [PubMed]
- Levi-Schaffer, F.; Tarrab-Hazdai, R.; Schryer, M.D.; Arnon, R.; Smolarsky, M. Isolation and partial characterization of the tegumental outer membrane of schistosomula of Schistosoma mansoni. Mol. Biochem. Parasitol. 1984, 13, 283–300. [Google Scholar] [CrossRef]
- Pearson, M.S.; Becker, L.; Driguez, P.; Young, N.D.; Gaze, S.; Mendes, T.; Li, X.H.; Doolan, D.L.; Midzi, N.; Mduluza, T.; et al. Of monkeys and men: Immunomic profiling of sera from humans and non-human primates resistant to schistosomiasis reveals novel potential vaccine candidates. Front. Immunol. 2015, 6, 213. [Google Scholar] [CrossRef] [PubMed]
- Arnon, R.; Silman, I.; Tarrab-Hazdai, R. Acetylcholinesterase of Schistosoma mansoni—Functional correlates. Contributed in honor of Professor Hans Neurath’s 90th birthday. Protein Sci. 1999, 8, 2553–2561. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, S.L.; Fischer, K.; Gobert, G.N.; McManus, D.P. A novel coagulation inhibitor from Schistosoma japonicum. Parasitology 2015, 142, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.K.; Bong, S.H.; Green, K.M.; Holmes, P.; Duke, M.; Loukas, A.; McManus, D.P. Correlative and dynamic imaging of the hatching biology of Schistosoma japonicum from eggs prepared by high pressure freezing. PLoS Negl. Trop. Dis. 2008, 2, e334. [Google Scholar] [CrossRef] [PubMed]
- Berriman, M.; Haas, B.J.; LoVerde, P.T.; Wilson, R.A.; Dillon, G.P.; Cerqueira, G.C.; Mashiyama, S.T.; Al-Lazikani, B.; Andrade, L.F.; Ashton, P.D.; et al. The genome of the blood fluke Schistosoma mansoni. Nature 2009, 460, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Young, N.D.; Jex, A.R.; Li, B.; Liu, S.; Yang, L.; Xiong, Z.; Li, Y.; Cantacessi, C.; Hall, R.S.; Xu, X.; et al. Whole-genome sequence of Schistosoma haematobium. Nat. Genet. 2012, 44, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, H.; Chen, X.; Zhang, L.; Wang, K.; Guo, J.; Huang, Z.; Zhang, B.; Huang, W.; Jin, K.; et al. The Schistosoma japonicum genome reveals features of host-parasite interplay. Nature 2009, 460, 345–351. [Google Scholar]
- You, H.; Zhang, W.; Jones, M.K.; Gobert, G.N.; Mulvenna, J.; Rees, G.; Spanevello, M.; Blair, D.; Duke, M.; Brehm, K.; et al. Cloning and characterisation of Schistosoma japonicum insulin receptors. PLoS ONE 2010, 5, e9868. [Google Scholar] [CrossRef] [PubMed]
- Bentley, G.N.; Jones, A.K.; Oliveros Parra, W.G.; Agnew, A. ShAR1alpha and ShAR1beta: Novel putative nicotinic acetylcholine receptor subunits from the platyhelminth blood fluke Schistosoma. Gene 2004, 329, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Camacho, M.; Agnew, A. Schistosoma: Rate of glucose import is altered by acetylcholine interaction with tegumental acetylcholine receptors and acetylcholinesterase. Exp. Parasitol. 1995, 81, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro-dos-Santos, G.; Verjovski-Almeida, S.; Leite, L.C. Schistosomiasis—A century searching for chemotherapeutic drugs. Parasitol. Res. 2006, 99, 505–521. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Stephenson, R.J.; Gobert, G.N.; McManus, D.P. Revisiting glucose uptake and metabolism in schistosomes: New molecular insights for improved schistosomiasis therapies. Front. Genet. 2014, 5, 176. [Google Scholar] [CrossRef] [PubMed]
- Ashton, P.D.; Harrop, R.; Shah, B.; Wilson, R.A. The schistosome egg: Development and secretions. Parasitology 2001, 122, 329–338. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Gobert, G.N.; Duke, M.G.; Zhang, W.; Li, Y.; Jones, M.K.; McManus, D.P. The insulin receptor is a transmission blocking veterinary vaccine target for zoonotic Schistosoma japonicum. Int. J. Parasitol. 2012, 42, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Kincaid-Smith, J.; Picard, M.A.L.; Cosseau, C.; Boissier, J.; Severac, D.; Grunau, C.; Toulza, E. Parent-of-Origin-Dependent Gene Expression in Male and Female Schistosome Parasites. Genome Biol. Evol. 2018, 10, 840–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, M.; Alsford, S.; Jones, A.; Agnew, A. Nicotinic acetylcholine receptors on the surface of the blood fluke Schistosoma. Mol. Biochem. Parasitol. 1995, 71, 127–134. [Google Scholar] [CrossRef]
- Camacho, M.; Agnew, A. Glucose uptake rates by Schistosoma mansoni, S. haematobium, and S. bovis adults using a flow in vitro culture system. J. Parasitol. 1995, 81, 637–640. [Google Scholar] [CrossRef] [PubMed]
- Hewitson, J.P.; Hamblin, P.A.; Mountford, A.P. Immunity induced by the radiation-attenuated schistosome vaccine. Parasite Immunol. 2005, 27, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Smythies, L.E.; Coulson, P.S.; Wilson, R.A. Monoclonal antibody to IFN-gamma modifies pulmonary inflammatory responses and abrogates immunity to Schistosoma mansoni in mice vaccinated with attenuated cercariae. J. Immunol. 1992, 149, 3654–3658. [Google Scholar] [PubMed]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Mowen, K.A.; Glimcher, L.H. Signaling pathways in Th2 development. Immunol. Rev. 2004, 202, 203–222. [Google Scholar] [CrossRef] [PubMed]
- McManus, D.P.; Loukas, A. Current status of vaccines for schistosomiasis. Clin. Microbiol. Rev. 2008, 21, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Gobert, G.N.; Moertel, L.; Brindley, P.J.; McManus, D.P. Developmental gene expression profiles of the human pathogen Schistosoma japonicum. BMC Genom. 2009, 10, 128. [Google Scholar] [CrossRef] [PubMed]
- You, H.; McManus, D.P.; Hu, W.; Smout, M.J.; Brindley, P.J.; Gobert, G.N. Transcriptional responses of in vivo praziquantel exposure in schistosomes identifies a functional role for calcium signalling pathway member CamKII. PLoS Pathog. 2013, 9, e1003254. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.E.; Rinaldi, G.; Gobert, G.N.; Kines, K.J.; Tort, J.F.; Brindley, P.J. RNA interference of Schistosoma mansoni cathepsin D, the apical enzyme of the hemoglobin proteolysis cascade. Mol. Biochem. Parasitol. 2008, 157, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, G.; Morales, M.E.; Alrefaei, Y.N.; Cancela, M.; Castillo, E.; Dalton, J.P.; Tort, J.F.; Brindley, P.J. RNA interference targeting leucine aminopeptidase blocks hatching of Schistosoma mansoni eggs. Mol. Biochem. Parasitol. 2009, 167, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, G.; Suttiprapa, S.; Brindley, P.J. Quantitative retrotransposon anchored PCR confirms transduction efficiency of transgenes in adult Schistosoma mansoni. Mol. Biochem. Parasitol. 2011, 177, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, G.; Okatcha, T.I.; Popratiloff, A.; Ayuk, M.A.; Suttiprapa, S.; Mann, V.H.; Liang, Y.S.; Lewis, F.A.; Loukas, A.; Brindley, P.J. Genetic manipulation of Schistosoma haematobium, the neglected schistosome. PLoS Negl. Trop. Dis. 2011, 5, e1348. [Google Scholar] [CrossRef] [PubMed]
- Boros, D.L.; Warren, K.S. Delayed hypersensitivity-type granuloma formation and dermal reaction induced and elicited by a soluble factor isolated from Schistosoma mansoni eggs. J. Exp. Med. 1970, 132, 488–507. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Cai, P.; Hou, N.; Piao, X.; Wang, H.; Hung, T.; Chen, Q. Genome-wide identification and characterization of a panel of house-keeping genes in Schistosoma japonicum. Mol. Biochem. Parasitol. 2012, 182, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Kyhse-Andersen, J. Electroblotting of multiple gels: A simple apparatus without buffer tank for rapid transfer of proteins from polyacrylamide to nitrocellulose. J. Biochem. Biophys. Methods. 1984, 10, 203–209. [Google Scholar] [CrossRef]
- Ressurreicao, M.; Kirk, R.S.; Rollinson, D.; Emery, A.M.; Page, N.M.; Walker, A.J. Sensory Protein Kinase Signaling in Schistosoma mansoni Cercariae: Host Location and Invasion. J. Infect. Dis. 2015, 212, 1787–1797. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Weerakoon, K.G.; Mu, Y.; Olveda, D.U.; Piao, X.; Liu, S.; Olveda, R.M.; Chen, Q.; Ross, A.G.; McManus, D.P. A Parallel Comparison of Antigen Candidates for Development of an Optimized Serological Diagnosis of Schistosomiasis Japonica in the Philippines. EBioMedicine 2017, 24, 237–246. [Google Scholar] [CrossRef] [PubMed]
- McManus, D.P.; Wong, J.Y.; Zhou, J.; Cai, C.; Zeng, Q.; Smyth, D.; Li, Y.; Kalinna, B.H.; Duke, M.J.; Yi, X. Recombinant paramyosin (rec-Sj-97) tested for immunogenicity and vaccine efficacy against Schistosoma japonicum in mice and water buffaloes. Vaccine 2001, 20, 870–878. [Google Scholar] [CrossRef]
- Dalton, J.P.; Day, S.R.; Drew, A.C.; Brindley, P.J. A method for the isolation of schistosome eggs and miracidia free of contaminating host tissues. Parasitology 1997, 115, 29–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurberg, A.D.; Goncalves, T.; Costa, T.A.; de Mattos, A.C.; Pascarelli, B.M.; de Manso, P.P.; Ribeiro-Alves, M.; Pelajo-Machado, M.; Peralta, J.M.; Coelho, P.M.; et al. The embryonic development of Schistosoma mansoni eggs: Proposal for a new staging system. Dev. Genes Evol. 2009, 219, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Ricciardi, A.; Dalton, J.P.; Ndao, M. Evaluation of the immune response and protective efficacy of Schistosoma mansoni Cathepsin B in mice using CpG dinucleotides as adjuvant. Vaccine 2015, 33, 346–353. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Immunoglobulin | Number Adult Worms Mean ± S.E. | Mean Length of Adult Worms (mm) Mean ± S.E. % Reduction (p Value) | Liver Eggs/g Mean ± S.E. % Reduction (p Value) | Liver Granuloma Density (%) Mean ± S.E. % Reduction (p Value) | Intestinal eggs/g Mean ± S.E. % Reduction (p Value) | Maturity of Intestinal Eggs in Stage V (%) Mean ± S.E. % Decrease (p Value) | Faecal Eggs/g/F Mean ± S.E. % Reduction (p Value) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Before Challenge | Before Perfusion | Stage I | Stage II | Stage III | Stage IV | Stage V | |||||||
| Control | IgG, IgG1, IgG2a, IgG2c. and IgG3 1:50 | - | (F) 5.9 ± 0.7 | (F) 10.4 ± 0.2 | 41,886 ± 6001 | 30 ± 3.9 | 64,483 ± 9998 | 3.4 ± 1.8 | 15.6 ± 3 | 27.7 ± 2 | 31.6 ± 2 | 21.6 ± 2 | 1398 ± 474 |
| (M) 8.5 ± 0.8 | (M) 6.6 ± 0.2 | ||||||||||||
| rSjAChE | IgG 1:819,200 IgG1 1:409,600 IgG2a 1:819,200 IgG2b 1:409,600 IgG2c 1:12,800 IgG3 1:6400 | - | (F) 5.3 ± 1.05 | (F) 9.8 ± 0.17 | 38,335 ± 7280 | 17.8 ± 2 41% ** p = 0.009 | 61,724 ± 12,008 | 9.8 ± 3.3 | 25 ± 3.7 | 34 ± 2.6 | 25 ± 0.9 ↓21% * p = 0.02 | 5.8 ± 0.8 ↓73% *** p = 0.00006 | 1039 ± 480 25.6% ns p = 0.77 |
| (M) 5.8±0.9 32% * p = 0.03 | (M) 6.8 ± 0.19 | ||||||||||||
| Weeks | OD450 | IgG1/IgG2a Ratio | |||
|---|---|---|---|---|---|
| IgG1 | IgG2a | ||||
| rSjAChE | Control | rSjAChE | Control | ||
| 4 (Third-vaccination) | 1.237 ± 0.07 | 0.13 ± 0.0035 | 0.859 ± 0.024 | 0.14 ± 0.002 | 1.441 |
| 6 (Pre-Challenge) | 1.4 ± 0.016 | 0.15 ± 0.0005 | 1.382 ± 0.022 | 0.15 ± 0.0005 | 1.013 |
| 8 | 0.98 ± 0.025 | 0.14 ± 0.0006 | 1.303 ± 0.003 | 0.13 ± 0.002 | 0.751 |
| 10 | 0.99 ± 0.003 | 0.14 ± 0.0007 | 1.21 ± 0.029 | 0.13 ± 0.0006 | 0.819 |
| 12 (Pre-Perfusion) | 0.46 ± 0.007 | 0.16 ± 0.0004 | 0.827 ± 0.06 | 0.13 ± 0.0007 | 0.56 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
You, H.; Liu, C.; Du, X.; Nawaratna, S.; Rivera, V.; Harvie, M.; Jones, M.; McManus, D.P. Suppression of Schistosoma japonicum Acetylcholinesterase Affects Parasite Growth and Development. Int. J. Mol. Sci. 2018, 19, 2426. https://doi.org/10.3390/ijms19082426
You H, Liu C, Du X, Nawaratna S, Rivera V, Harvie M, Jones M, McManus DP. Suppression of Schistosoma japonicum Acetylcholinesterase Affects Parasite Growth and Development. International Journal of Molecular Sciences. 2018; 19(8):2426. https://doi.org/10.3390/ijms19082426
Chicago/Turabian StyleYou, Hong, Chang Liu, Xiaofeng Du, Sujeevi Nawaratna, Vanessa Rivera, Marina Harvie, Malcolm Jones, and Donald P. McManus. 2018. "Suppression of Schistosoma japonicum Acetylcholinesterase Affects Parasite Growth and Development" International Journal of Molecular Sciences 19, no. 8: 2426. https://doi.org/10.3390/ijms19082426