Renal Chemerin Expression is Induced in Models of Hypertensive Nephropathy and Glomerulonephritis and Correlates with Markers of Inflammation and Fibrosis

Abstract
:1. Introduction
2. Results
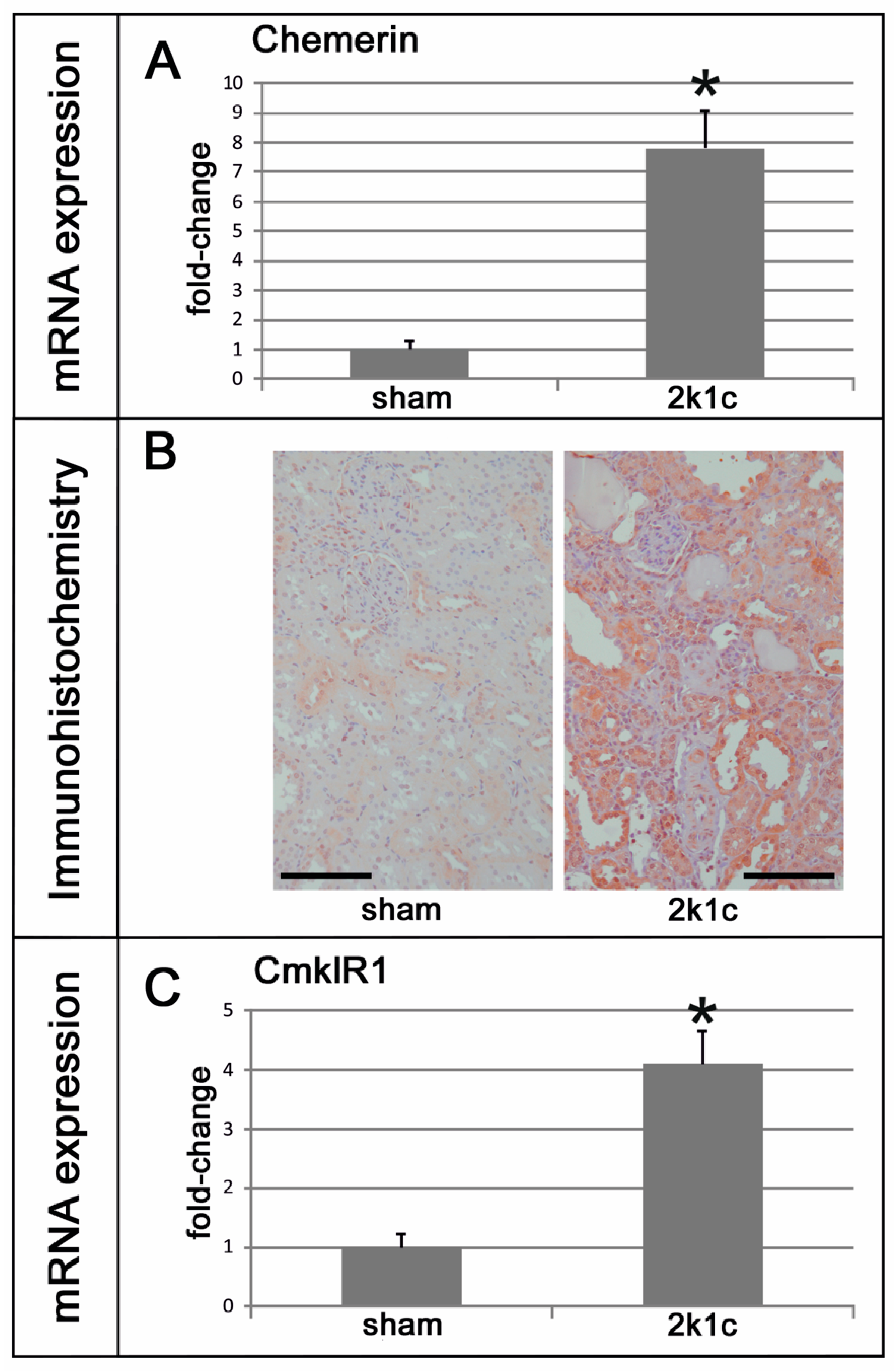
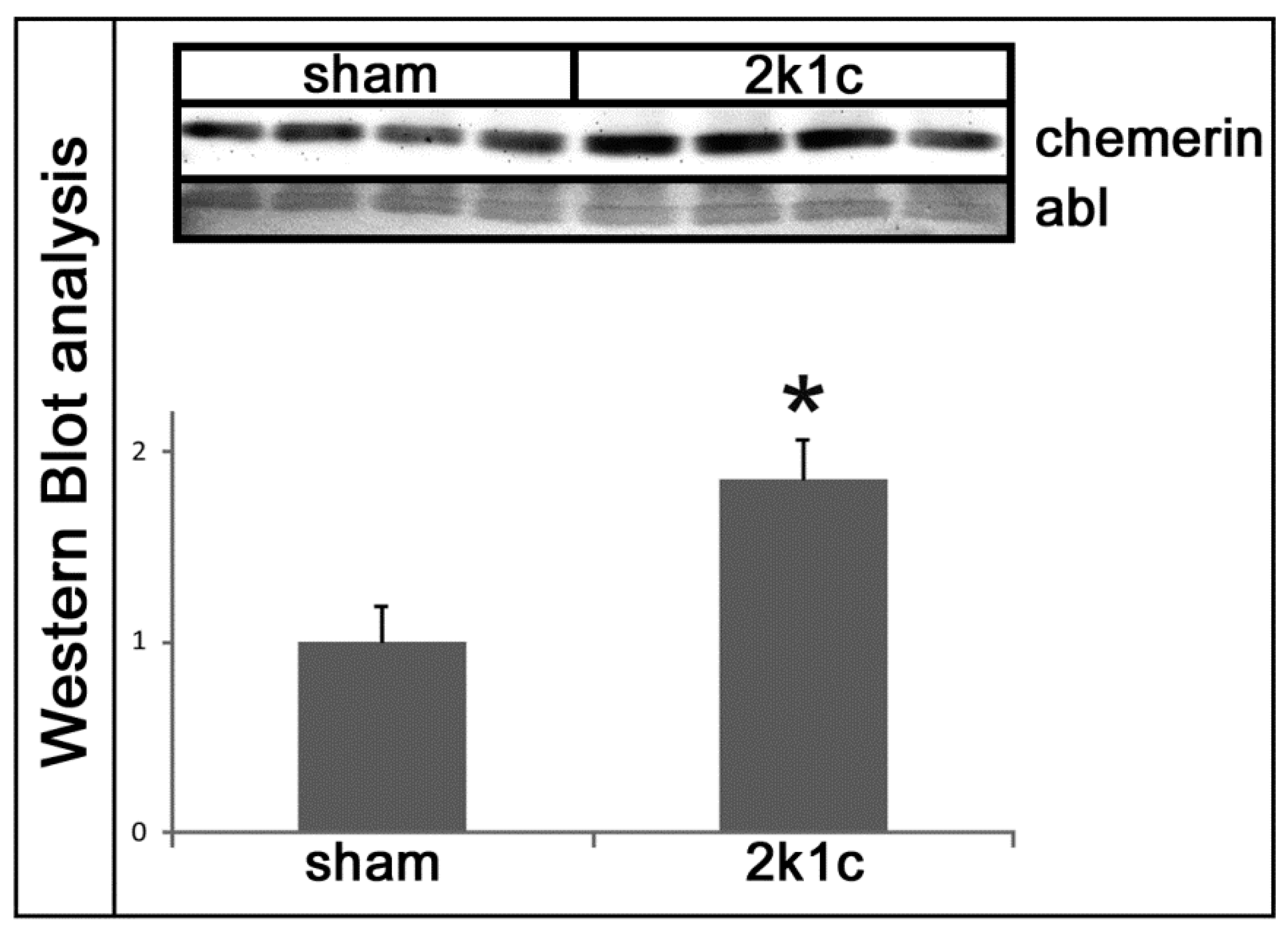
2.1. Chemerin is Induced in Kidneys Exposed to High Blood Pressure
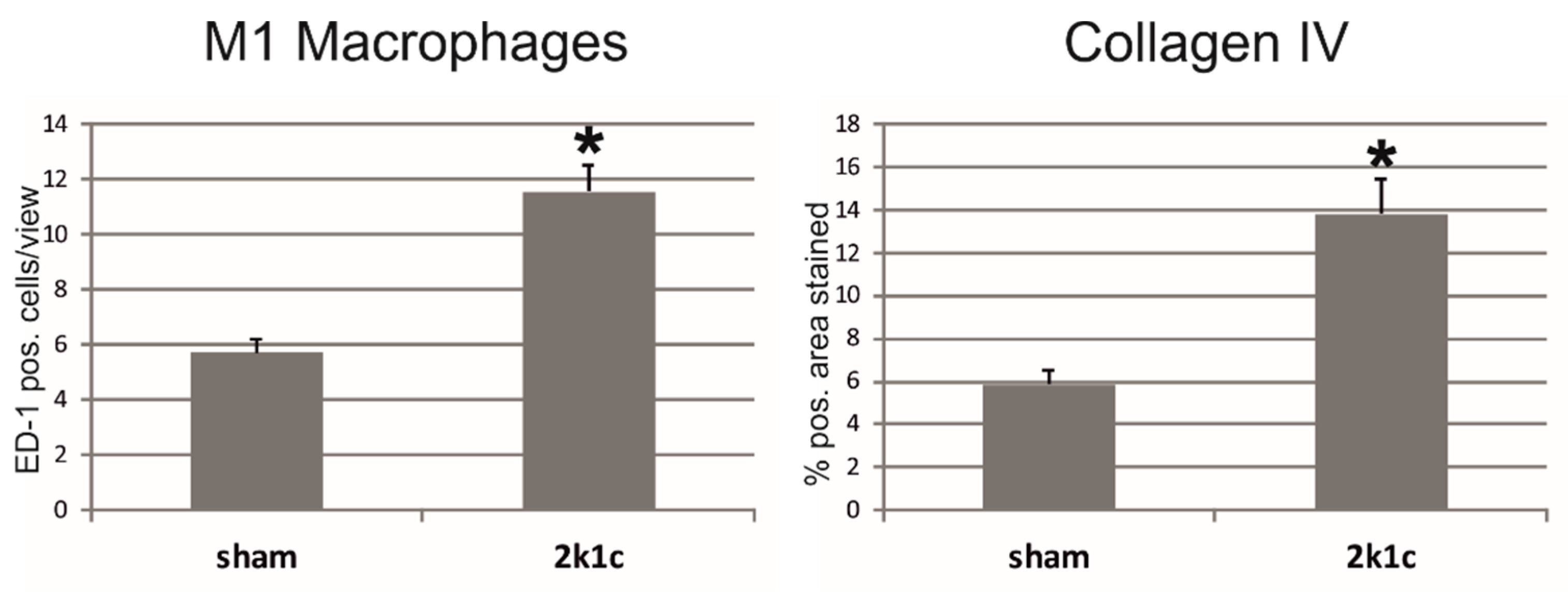
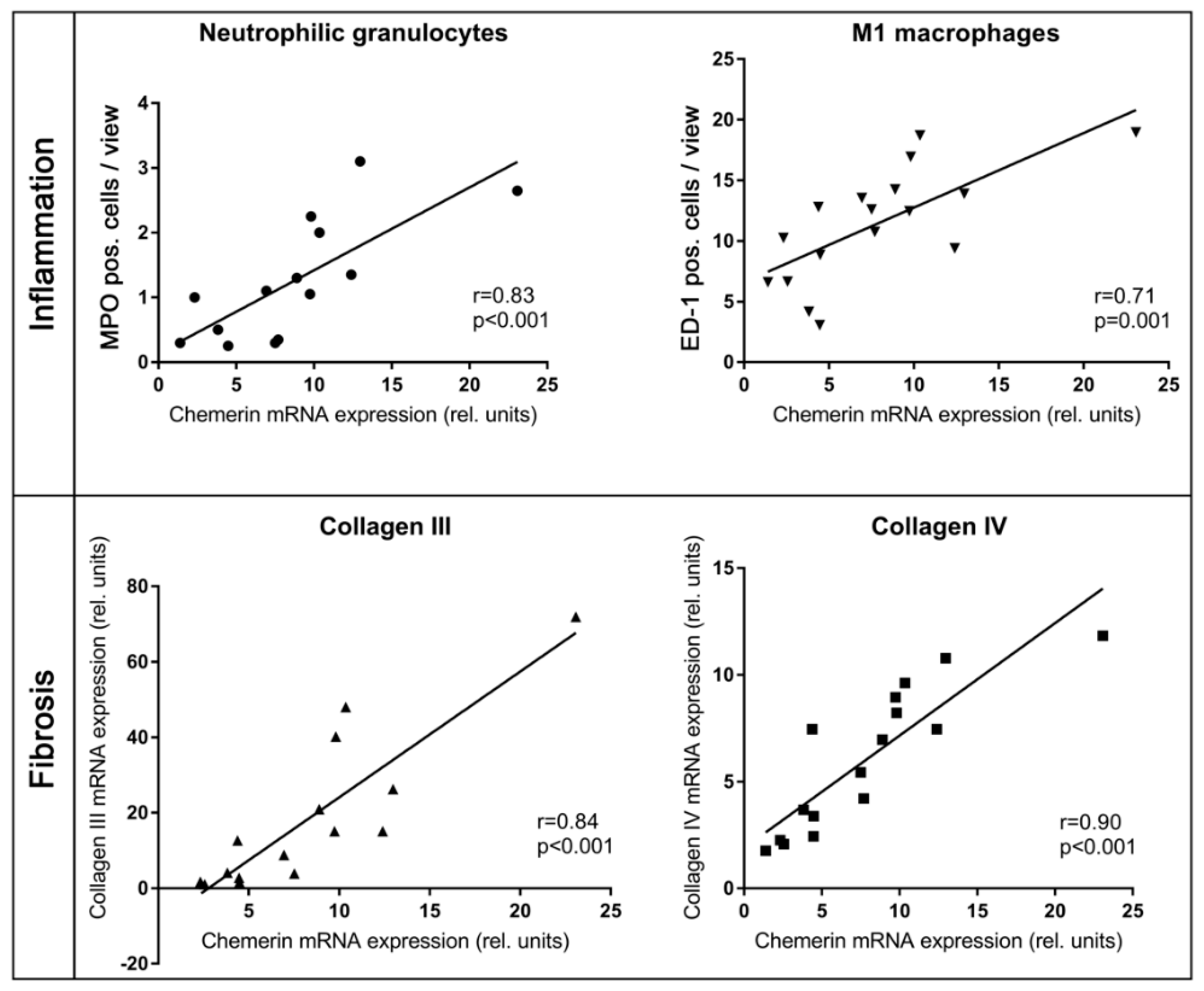
2.2. In Kidneys Exposed to High Blood Pressure, Chemerin Expression Correlates with Markers of Renal Damage, Inflammation, and Fibrosis
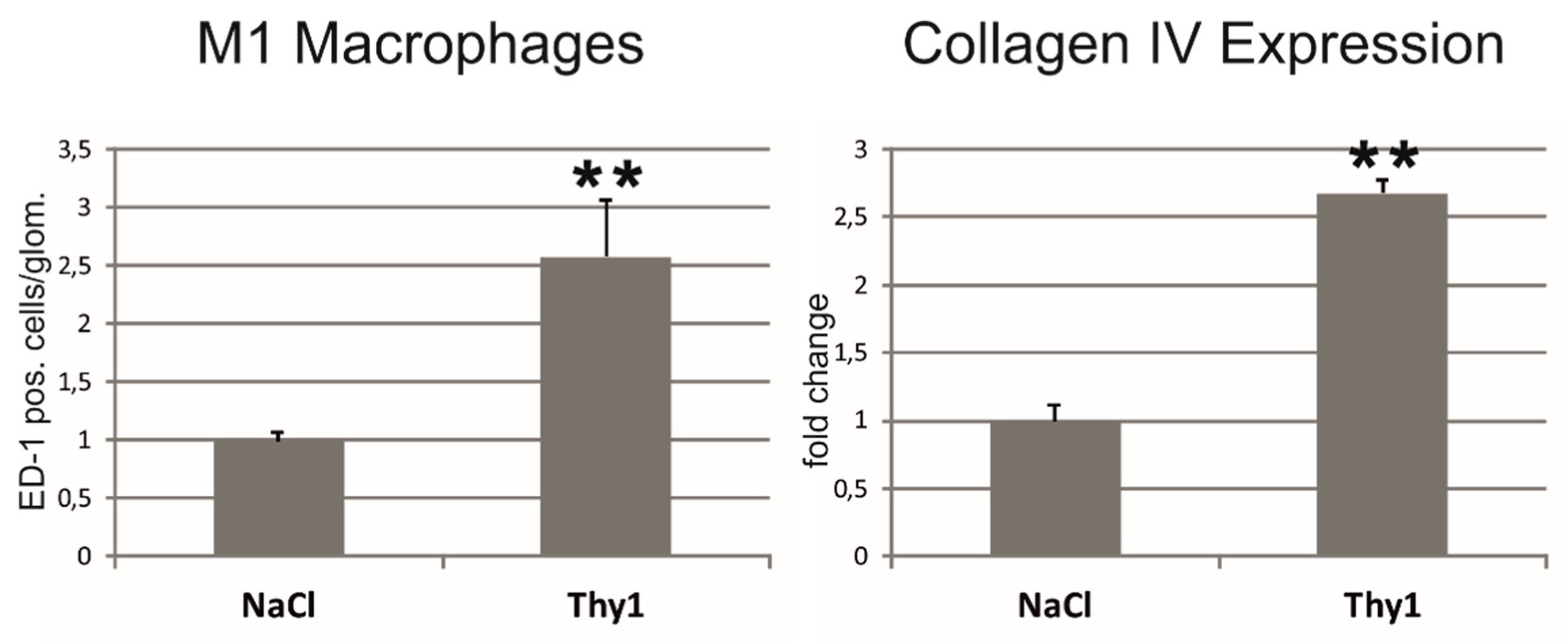
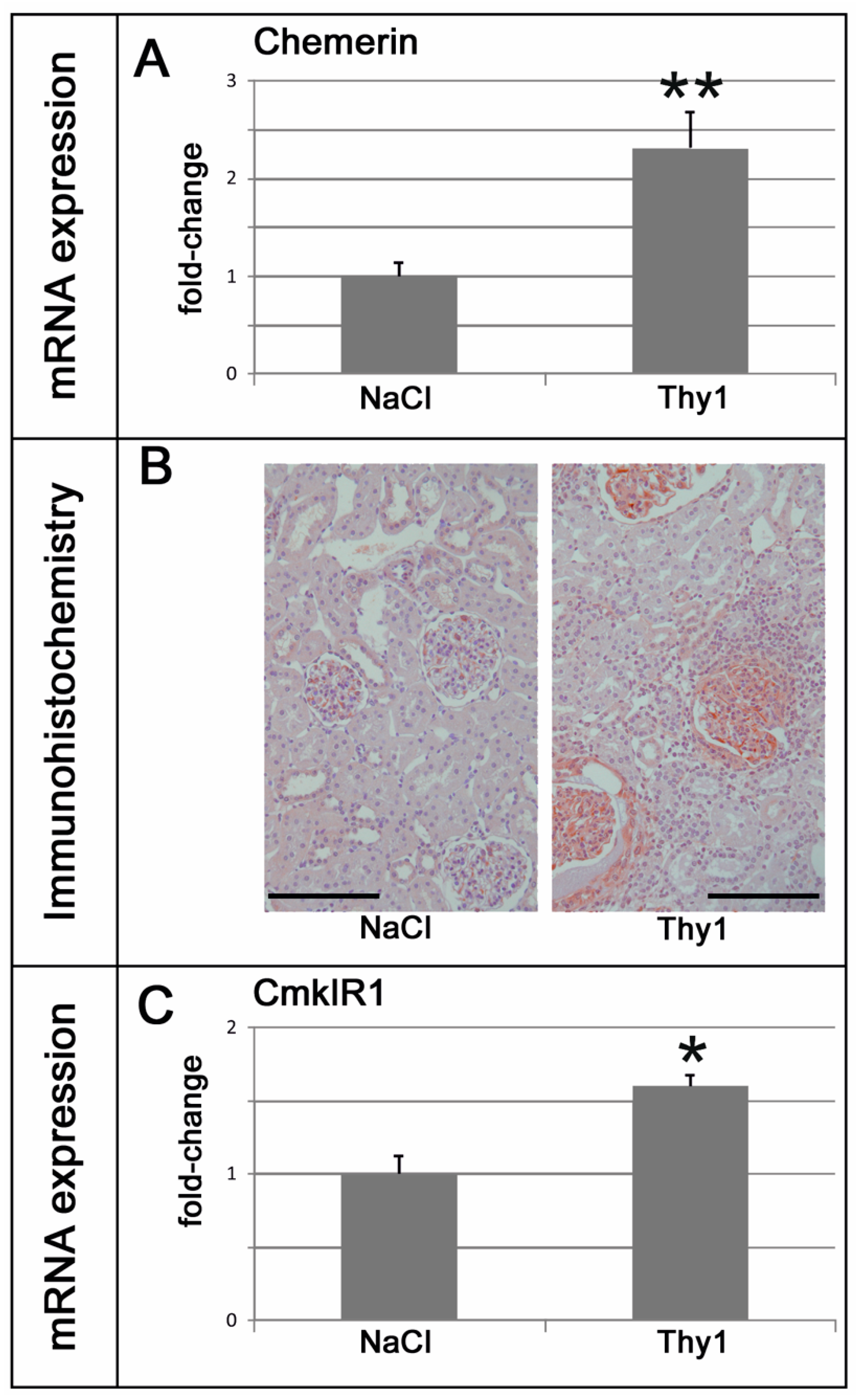
2.3. Chemerin is Induced in Glomeruli Afflicted with Thy1.1 Nephritis and Correlates with Markers of Renal Damage, Inflammation, and Fibrosis
2.4. Chemerin Plasma Levels are Not Increased in Rat Models of Renal Injury
3. Discussion
4. Materials and Methods
4.1. Experimental Procedures
4.2. Blood Pressure Measurements
4.3. Measurement of Serum and Urine Parameters
4.4. Tissue Sampling
4.5. Immunohistochemistry
4.6. Western Blot Analysis
4.7. Real-Time Polymerase Chain Reaction (PCR) Analyses
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Breyer, M.D.; Susztak, K. Developing Treatments for Chronic Kidney Disease in the 21st Century. Semin. Nephrol. 2016, 36, 436–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernandez, T.; Mayadas, T.N. The Changing Landscape of Renal Inflammation. Trends Mol. Med. 2016, 22, 151–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilgers, K.F.; Hartner, A.; Porst, M.; Mai, M.; Wittmann, M.; Hugo, C.; Ganten, D.; Geiger, H.; Veelken, R.; Mann, J.F. Monocyte chemoattractant protein-1 and macrophage infiltration in hypertensive kidney injury. Kidney Int. 2000, 58, 2408–2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafi, T.; Coresh, J. 1 - Chronic Kidney Disease: Definition, Epidemiology, Cost, and Outcomes. In Chronic Kidney Disease, Dialysis, and Transplantation (Fourth Edition), Himmelfarb, J.; Ikizler, T.A., Ed.; Eds. Elsevier: Philadelphia, PA, USA, 2019. [Google Scholar]
- Wada, T.; Matsushima, K.; Kaneko, S. The role of chemokines in glomerulonephritis. Front. Biosci. 2008, 13, 3966–3974. [Google Scholar] [CrossRef] [PubMed]
- Chung, A.C.; Lan, H.Y. Chemokines in renal injury. J. Am. Soc. Nephrol. 2011, 22, 802–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittamer, V.; Franssen, J.D.; Vulcano, M.; Mirjolet, J.F.; Le Poul, E.; Migeotte, I.; Brezillon, S.; Tyldesley, R.; Blanpain, C.; Detheux, M.; et al. Specific recruitment of antigen-presenting cells by chemerin, a novel processed ligand from human inflammatory fluids. J. Exp. Med. 2003, 198, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef]
- Bozaoglu, K.; Bolton, K.; McMillan, J.; Zimmet, P.; Jowett, J.; Collier, G.; Walder, K.; Segal, D. Chemerin is a novel adipokine associated with obesity and metabolic syndrome. Endocrinology 2007, 148, 4687–4694. [Google Scholar] [CrossRef]
- Goralski, K.B.; McCarthy, T.C.; Hanniman, E.A.; Zabel, B.A.; Butcher, E.C.; Parlee, S.D.; Muruganandan, S.; Sinal, C.J. Chemerin, a novel adipokine that regulates adipogenesis and adipocyte metabolism. J. Biol. Chem. 2007, 282, 28175–28188. [Google Scholar] [CrossRef] [Green Version]
- Bonomini, M.; Pandolfi, A. Chemerin in renal dysfunction and cardiovascular disease. Vascul. Pharmacol. 2016, 77, 28–34. [Google Scholar] [CrossRef]
- Kaur, J.; Mattu, H.S.; Chatha, K.; Randeva, H.S. Chemerin in human cardiovascular disease. Vascul. Pharmacol. 2018, 110, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Feng, P. Elevated serum chemerin concentrations are associated with renal dysfunction in type 2 diabetic patients. Diabetes Res. Clin. Pract. 2011, 91, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, P.; Sledzinski, T.; Zielinska, H.; Lizakowski, S.; Goyke, E.; Szrok-Wojtkiewicz, S.; Swierczynski, J.; Rutkowski, B. Decrease of serum chemerin concentration in patients with end stage renal disease after successful kidney transplantation. Regul. Pept. 2012, 173, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Pfau, D.; Bachmann, A.; Lossner, U.; Kratzsch, J.; Bluher, M.; Stumvoll, M.; Fasshauer, M. Serum levels of the adipokine chemerin in relation to renal function. Diabetes Care 2010, 33, 171–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaszak, J.; Szolkiewicz, M.; Sucajtys-Szulc, E.; Konarzewski, M.; Lizakowski, S.; Swierczynski, J.; Rutkowski, B. High serum chemerin level in CKD patients is related to kidney function, but not to its adipose tissue overproduction. Ren Fail. 2015, 37, 1033–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.M.; Tang, P.M.; Li, J.; Lan, H.Y. TGF-beta/Smad signaling in renal fibrosis. Front. Physiol. 2015, 6, 82. [Google Scholar] [CrossRef] [Green Version]
- Meran, S.; Steadman, R. Fibroblasts and myofibroblasts in renal fibrosis. Int. J. Exp. Pathol. 2011, 92, 158–167. [Google Scholar] [CrossRef]
- Ernst, M.C.; Sinal, C.J. Chemerin: At the crossroads of inflammation and obesity. Trends Endocrinol. Metab. 2010, 21, 660–667. [Google Scholar] [CrossRef]
- Arita, M.; Ohira, T.; Sun, Y.P.; Elangovan, S.; Chiang, N.; Serhan, C.N. Resolvin E1 selectively interacts with leukotriene B4 receptor BLT1 and ChemR23 to regulate inflammation. J. Immunol. 2007, 178, 3912–3917. [Google Scholar] [CrossRef] [Green Version]
- Vermi, W.; Riboldi, E.; Wittamer, V.; Gentili, F.; Luini, W.; Marrelli, S.; Vecchi, A.; Franssen, J.D.; Communi, D.; Massardi, L.; et al. Role of ChemR23 in directing the migration of myeloid and plasmacytoid dendritic cells to lymphoid organs and inflamed skin. J. Exp. Med. 2005, 201, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Kaur, J.; Adya, R.; Tan, B.K.; Chen, J.; Randeva, H.S. Identification of chemerin receptor (ChemR23) in human endothelial cells: Chemerin-induced endothelial angiogenesis. Biochem. Biophys. Res. Commun. 2010, 391, 1762–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeshima, Y.; Makino, H. Angiogenesis and chronic kidney disease. Fibrogenesis Tissue Repair 2010, 3, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Limbu, M.H.; Wang, Z.; Liu, J.; Liu, L.; Zhang, X.; Chen, P.; Liu, B. MMP-2 and 9 in Chronic Kidney Disease. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.X.; Zhang, H.; Xu, W.H.; Hao, F.; Liu, S.L.; Bai, M.M.; Mu, J.W.; Zhang, H.J. Effect of Irbesartan on Chemerin in the Renal Tissues of Diabetic Rats. Kidney Blood Press Res. 2015, 40, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Goldblatt, H.; Lynch, J.; Hanzal, R.F.; Summerville, W.W. Studies on Experimental Hypertension : I. The Production of Persistent Elevation of Systolic Blood Pressure by Means of Renal Ischemia. J. Exp. Med. 1934, 59, 347–379. [Google Scholar] [CrossRef] [Green Version]
- Goldblatt, H.; Kahn, J.R.; Hanzal, R.F. Studies on Experimental Hypertension : Ix. The Effect on Blood Pressure of Constriction of the Abdominal Aorta above and Below the Site of Origin of Both Main Renal Arteries. J. Exp. Med. 1939, 69, 649–674. [Google Scholar] [CrossRef] [Green Version]
- Hilgers, K.F.; Hartner, A.; Porst, M.; Veelken, R.; Mann, J.F. Angiotensin II type 1 receptor blockade prevents lethal malignant hypertension: Relation to kidney inflammation. Circulation 2001, 104, 1436–1440. [Google Scholar] [CrossRef] [Green Version]
- Wiesel, P.; Mazzolai, L.; Nussberger, J.; Pedrazzini, T. Two-kidney, one clip and one-kidney, one clip hypertension in mice. Hypertension 1997, 29, 1025–1030. [Google Scholar] [CrossRef]
- Corbier, A.; Lecaque, D.; Secchi, J.; Depouez, B.; Hamon, G. Effects of 4 weeks of treatment with trandolapril on renal hypertension and cardiac and vascular hypertrophy in the rat. J. Cardiovasc. Pharmacol. 1994, 23, S26–S29. [Google Scholar] [CrossRef]
- Ozawa, Y.; Kobori, H.; Suzaki, Y.; Navar, L.G. Sustained renal interstitial macrophage infiltration following chronic angiotensin II infusions. Am. J. Physiol. Renal Physiol. 2007, 292, F330–F339. [Google Scholar] [CrossRef]
- Mezzano, S.A.; Aros, C.A.; Droguett, A.; Burgos, M.E.; Ardiles, L.G.; Flores, C.A.; Carpio, D.; Vio, C.P.; Ruiz-Ortega, M.; Egido, J. Renal angiotensin II up-regulation and myofibroblast activation in human membranous nephropathy. Kidney Int. 2003, 64, S39–S45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Chung, A.C.; Huang, X.R.; Lan, H.Y. Angiotensin II induces connective tissue growth factor and collagen I expression via transforming growth factor-beta-dependent and -independent Smad pathways: The role of Smad3. Hypertension 2009, 54, 877–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Border, W.A.; Noble, N.A. Interactions of transforming growth factor-beta and angiotensin II in renal fibrosis. Hypertension 1998, 31, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezzano, S.A.; Ruiz-Ortega, M.; Egido, J. Angiotensin II and renal fibrosis. Hypertension 2001, 38, 635–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, G.; Schneider, A.; Wenzel, U.; Helmchen, U.; Stahl, R.A. Regulation of glomerular TGF-beta expression in the contralateral kidney of two-kidney, one-clip hypertensive rats. J. Am. Soc. Nephrol. 1998, 9, 763–772. [Google Scholar] [PubMed]
- Hoppmann, J.; Perwitz, N.; Meier, B.; Fasshauer, M.; Hadaschik, D.; Lehnert, H.; Klein, J. The balance between gluco- and mineralo-corticoid action critically determines inflammatory adipocyte responses. J. Endocrinol. 2010, 204, 153–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, H.; Hierer, J.; Haas, O.; Forssmann, W.G. Quantification of angiotensin-converting-enzyme-mediated degradation of human chemerin 145-154 in plasma by matrix-assisted laser desorption/ionization-time-of-flight mass spectrometry. Anal. Biochem. 2007, 362, 117–125. [Google Scholar] [CrossRef]
- Urushihara, M.; Kinoshita, Y.; Kondo, S.; Kagami, S. Involvement of the intrarenal renin-angiotensin system in experimental models of glomerulonephritis. J. Biomed. Biotechnol. 2012, 2012, 601786. [Google Scholar] [CrossRef] [Green Version]
- Anders, H.J.; Vielhauer, V.; Schlondorff, D. Chemokines and chemokine receptors are involved in the resolution or progression of renal disease. Kidney Int. 2003, 63, 401–415. [Google Scholar] [CrossRef] [Green Version]
- Ishizaki, M.; Masuda, Y.; Fukuda, Y.; Sugisaki, Y.; Yamanaka, N.; Masugi, Y. Experimental mesangioproliferative glomerulonephritis in rats induced by intravenous administration of anti-thymocyte serum. Acta Pathol. Jpn. 1986, 36, 1191–1203. [Google Scholar] [CrossRef]
- Hartner, A.; Schocklmann, H.; Prols, F.; Muller, U.; Sterzel, R.B. Alpha8 integrin in glomerular mesangial cells and in experimental glomerulonephritis. Kidney Int. 1999, 56, 1468–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, N.; Iseki, K.; Suzuki, S.; Mori, T.; Hagino, S.; Zhang, Y.; Yokoya, S.; Kawasaki, Y.; Suzuki, J.; Isome, M.; et al. Uninephrectomy induces progressive glomerulosclerosis and apoptosis in anti-Thy1 glomerulonephritis. Pathol. Int. 2005, 55, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Kawachi, H.; Iwanaga, T.; Toyabe, S.; Oite, T.; Shimizu, F. Mesangial sclerotic change with persistent proteinuria in rats after two consecutive injections of monoclonal antibody 1-22-3. Clin. Exp. Immunol. 1992, 90, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Qureshi, A.R.; Anderstam, B.; Heimburger, O.; Barany, P.; Lindholm, B.; Stenvinkel, P.; Axelsson, J. Clinical importance of an elevated circulating chemerin level in incident dialysis patients. Nephrol. Dial. Transplant. 2010, 25, 4017–4023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukai, H.; Dai, L.; Chen, Z.; Lindholm, B.; Ripsweden, J.; Brismar, T.B.; Heimburger, O.; Barany, P.; Qureshi, A.R.; Soderberg, M.; et al. Inverse J-shaped relation between coronary arterial calcium density and mortality in advanced chronic kidney disease. Nephrol. Dial. Transplant. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carracedo, M.; Artiach, G.; Witasp, A.; Claria, J.; Carlstrom, M.; Laguna-Fernandez, A.; Stenvinkel, P.; Back, M. The G-protein coupled receptor ChemR23 determines smooth muscle cell phenotypic switching to enhance high phosphate-induced vascular calcification. Cardiovasc Res. 2019, 115, 1557–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artiach, G.; Carracedo, M.; Claria, J.; Laguna-Fernandez, A.; Back, M. Opposing Effects on Vascular Smooth Muscle Cell Proliferation and Macrophage-induced Inflammation Reveal a Protective Role for the Proresolving Lipid Mediator Receptor ChemR23 in Intimal Hyperplasia. Front. Pharmacol. 2018, 9, 1327. [Google Scholar] [CrossRef]
- Laguna-Fernandez, A.; Checa, A.; Carracedo, M.; Artiach, G.; Petri, M.H.; Baumgartner, R.; Forteza, M.J.; Jiang, X.; Andonova, T.; Walker, M.E.; et al. ERV1/ChemR23 Signaling Protects Against Atherosclerosis by Modifying Oxidized Low-Density Lipoprotein Uptake and Phagocytosis in Macrophages. Circulation 2018, 138, 1693–1705. [Google Scholar] [CrossRef]
- Sato, K.; Yoshizawa, H.; Seki, T.; Shirai, R.; Yamashita, T.; Okano, T.; Shibata, K.; Wakamatsu, M.J.; Mori, Y.; Morita, T.; et al. Chemerin-9, a potent agonist of chemerin receptor (ChemR23), prevents atherogenesis. Clin. Sci. (Lond) 2019, 133, 1779–1796. [Google Scholar] [CrossRef]
- Carracedo, M.; Witasp, A.; Qureshi, A.R.; Laguna-Fernandez, A.; Brismar, T.; Stenvinkel, P.; Back, M. Chemerin inhibits vascular calcification through ChemR23 and is associated with lower coronary calcium in chronic kidney disease. J. Int. Med. 2019, 286, 449–457. [Google Scholar] [CrossRef]
- Hartner, A.; Jagusch, L.; Cordasic, N.; Amann, K.; Veelken, R.; Jacobi, J.; Hilgers, K.F. Impaired Neovascularization and Reduced Capillary Supply in the Malignant vs. Non-malignant Course of Experimental Renovascular Hypertension. Front. Physiol. 2016, 7, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, M.; Hilgers, K.F.; Wagner, J.; Mann, J.F.; Geiger, H. Expression of angiotensin-converting enzyme in renovascular hypertensive rat kidney. Hypertension 1995, 25, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.J.; Mooney, A.; Hughes, J.; Lombardi, D.; Johnson, R.J.; Savill, J. Mesangial cell apoptosis: The major mechanism for resolution of glomerular hypercellularity in experimental mesangial proliferative nephritis. J. Clin. Investig. 1994, 94, 2105–2116. [Google Scholar] [CrossRef] [PubMed]
- Menendez-Castro, C.; Fahlbusch, F.; Cordasic, N.; Amann, K.; Munzel, K.; Plank, C.; Wachtveitl, R.; Rascher, W.; Hilgers, K.F.; Hartner, A. Early and late postnatal myocardial and vascular changes in a protein restriction rat model of intrauterine growth restriction. PLoS ONE 2011, 6, e20369. [Google Scholar] [CrossRef] [Green Version]
- Menendez-Castro, C.; Hilgers, K.F.; Amann, K.; Daniel, C.; Cordasic, N.; Wachtveitl, R.; Fahlbusch, F.; Plank, C.; Dotsch, J.; Rascher, W.; et al. Intrauterine growth restriction leads to a dysregulation of Wilms’ tumour supressor gene 1 (WT1) and to early podocyte alterations. Nephrol Dial. Transplant. 2013, 28, 1407–1417. [Google Scholar] [CrossRef]
- Menendez-Castro, C.; Toka, O.; Fahlbusch, F.; Cordasic, N.; Wachtveitl, R.; Hilgers, K.F.; Rascher, W.; Hartner, A. Impaired myocardial performance in a normotensive rat model of intrauterine growth restriction. Pediatr. Res. 2014, 75, 697–706. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Gibson, U.E.; Heid, C.A.; Williams, P.M. A novel method for real time quantitative RT-PCR. Genome Res. 1996, 6, 995–1001. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological Parameter | Sham | 2k1c | p-Value |
|---|---|---|---|
| Rel. right kidney weight (mg/g body weight) | 3.21 ± 0.06 | 4.87 ± 0.25 | <0.001 |
| Serum urea (mg/dL) | 37.74 ± 1.13 | 80.92 ± 11.26 | <0.001 |
| Serum creatinine (mg/dL) | 0.195 ± 0.007 | 0.335 ± 0.038 | <0.001 |
| Rel. left ventricular weight (mg/g body weight) | 2.05 ± 0.04 | 3.03 ± 0.21 | 0.003 |
| Mean arterial blood pressure (mm Hg) | 113.2 ± 2.6 | 203.7 ± 5.0 | 0.014 |
| Cell Type | Sham | 2k1c | p-Value |
|---|---|---|---|
| M2 macrophages (CD163 pos. cells/cortical view) | 0.09 ± 0.04 | 0.86 ± 0.20 | <0.001 |
| Neutrophil granulocytes (myeloperoxidase pos. cells/cortical view) | 0.53 ± 0.15 | 1.49 ± 0.30 | 0.039 |
| Total T-cells (CD3 pos. cells/cortical view) | 1.23 ± 0.47 | 4.88 ± 1.81 | 0.046 |
| Helper T-cells (CD4 pos. cells/cortical view) | 11.56 ± 3.37 | 75.45 ± 14.60 | 0.002 |
| Cytotoxic T-cells (CD8a pos. cells/cortical view) | 3.12 ± 0.38 | 4.76 ± 0.35 | 0.456 |
| Fibrotic Marker | Sham | 2k1c | p-Value |
|---|---|---|---|
| TGFβ-1 expression (fold change) | 1.00 ± 0.25 | 3.69 ± 0.44 | 0.001 |
| Smooth muscle actin expression (fold change) | 1.00 ± 0.21 | 5.89 ± 0.96 | 0.014 |
| Activated fibroblasts (smooth muscle actin pos. cells/cortical view) | 0.26 ± 0.04 | 5.74 ± 1.50 | 0.002 |
| Fibronectin expression (fold change) | 1.00 ± 0.24 | 8.81 ± 1.55 | 0.003 |
| Collagen I expression (fold change) | 1.00 ± 0.32 | 4.42 ± 0.78 | 0.024 |
| Collagen I stain (% pos. cells/cortical view) | 4.66 ± 0.44 | 7.98 ± 1.14 | 0.001 |
| Collagen III expression (fold change) | 1.00 ± 0.49 | 18.25 ± 5.32 | 0.004 |
| Collagen IV expression (fold change) | 1.00 ± 0.22 | 6.03 ± 0.83 | 0.001 |
| Chemerin (mRNA Expression) | r | p-Value |
|---|---|---|
| Serum creatinine (mg/dL) | 0.62 | 0.009 |
| Serum urea (mg/dL) | 0.77 | <0.001 |
| Mean arterial blood pressure (mm Hg) | 0.42 | 0.12 |
| M1 macrophages (ED-1 pos. cells/view) | 0.71 | 0.001 |
| M2 macrophages (CD163 pos. cells/view) | 0.51 | 0.16 |
| Total T-cells (CD3 pos. cells/view) | 0.27 | 0.40 |
| Cytotoxic T-cells (CD8a pos. cells/view) | 0.47 | 0.11 |
| Helper T-cells (CD4 pos. cells/view) | −0.14 | 0.63 |
| Neutrophil granulocytes (MPO pos. cells/view) | 0.83 | <0.001 |
| Activated myofibroblasts (SMA-pos. cells/view) | 0.74 | 0.003 |
| Smooth muscle actin (mRNA expression) | 0.65 | 0.005 |
| Fibronectin (mRNA expression) | 0.86 | <0.001 |
| Collagen I (% pos. area stained) | 0.65 | 0.043 |
| Collagen I (mRNA expression) | 0.83 | <0.001 |
| Collagen III (mRNA expression) | 0.84 | <0.001 |
| Collagen IV (% pos. area stained) | 0.64 | 0.014 |
| Collagen IV (mRNA expression) | 0.90 | <0.001 |
| TGFβ-1 (mRNA expression) | 0.73 | 0.001 |
| CmklR1 (mRNA expression) | 0.89 | <0.001 |
| Damage Marker | NaCl | Thy1 | p-Value |
|---|---|---|---|
| Rel. right kidney weight (mg/g body weight) | 4.30 ± 0.15 | 8.57 ± 0.18 | <0.01 |
| Albuminuria (mg/24 h) | 0.80 ± 0.19 | 725.79 ± 303.13 | <0.01 |
| Serum creatinine (mg/dL) | 0.19 ± 0.01 | 0.34 ± 0.03 | <0.01 |
| Cytotoxic T-cells (CD8a pos. cells/cortical view) | 3.51 ± 0.56 | 5.72 ± 0.41 | n.s. |
| Helper T-cells (CD4 pos. cells/cortical view) | 3.05 ± 0.61 | 6.23 ± 2.39 | n.s. |
| Mean arterial blood pressure (mm Hg) | 118.5 ± 1.5 | 123.7 ± 6.3 | n.s. |
| Rel. left ventricular weight (mg/g body weight) | 2.23 ± 0.11 | 2.49 ± 0.09 | n.s. |
| Chemerin (mRNA Expression) | r | p-Value |
|---|---|---|
| CmklR1 (mRNA expression) | 0.89 | 0.001 |
| Serum creatinine (mg/dL) | 0.73 | 0.017 |
| Albuminuria (mg/24h) | 0.73 | 0.016 |
| Mean arterial blood pressure (mm Hg) | 0.42 | 0.262 |
| M1 macrophages (ED-1 pos. cells/view) | 0.72 | 0.019 |
| Cytotoxic T-cells (CD8a pos. cells/view) | 0.58 | 0.082 |
| Helper T-cells (CD4 pos. cells/view) | 0.39 | 0.266 |
| Collagen IV (mRNA expression) | 0.83 | 0.005 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mocker, A.; Hilgers, K.F.; Cordasic, N.; Wachtveitl, R.; Menendez-Castro, C.; Woelfle, J.; Hartner, A.; Fahlbusch, F.B. Renal Chemerin Expression is Induced in Models of Hypertensive Nephropathy and Glomerulonephritis and Correlates with Markers of Inflammation and Fibrosis. Int. J. Mol. Sci. 2019, 20, 6240. https://doi.org/10.3390/ijms20246240
Mocker A, Hilgers KF, Cordasic N, Wachtveitl R, Menendez-Castro C, Woelfle J, Hartner A, Fahlbusch FB. Renal Chemerin Expression is Induced in Models of Hypertensive Nephropathy and Glomerulonephritis and Correlates with Markers of Inflammation and Fibrosis. International Journal of Molecular Sciences. 2019; 20(24):6240. https://doi.org/10.3390/ijms20246240
Chicago/Turabian StyleMocker, Alexander, Karl F. Hilgers, Nada Cordasic, Rainer Wachtveitl, Carlos Menendez-Castro, Joachim Woelfle, Andrea Hartner, and Fabian B. Fahlbusch. 2019. "Renal Chemerin Expression is Induced in Models of Hypertensive Nephropathy and Glomerulonephritis and Correlates with Markers of Inflammation and Fibrosis" International Journal of Molecular Sciences 20, no. 24: 6240. https://doi.org/10.3390/ijms20246240