
Investigation of Neuropathology after Nerve Release in Chronic Constriction Injury of Rat Sciatic Nerve

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
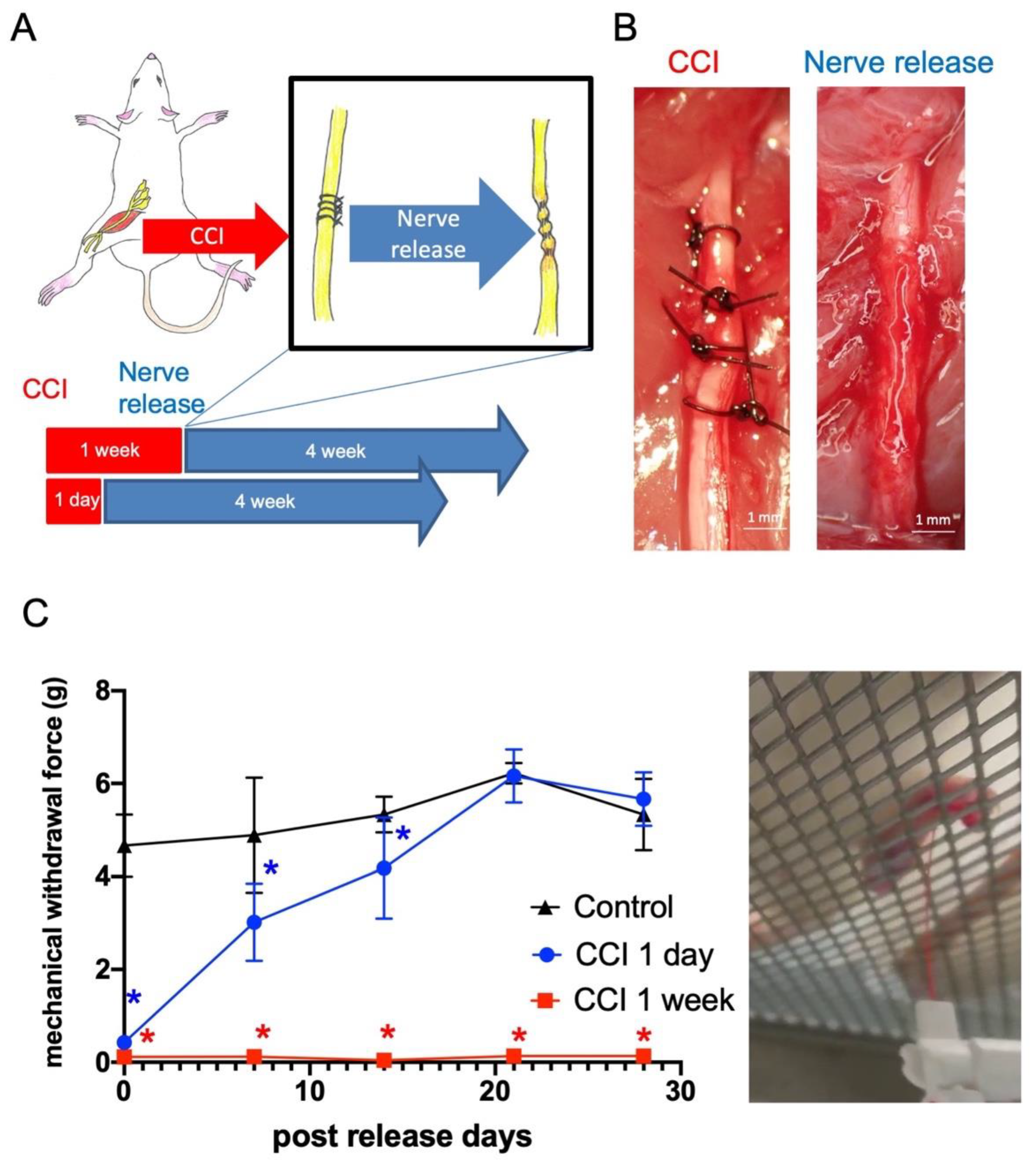
2.1. Late Release of Chronic Nerve Constriction Results in Persistent Neuropathic Pain
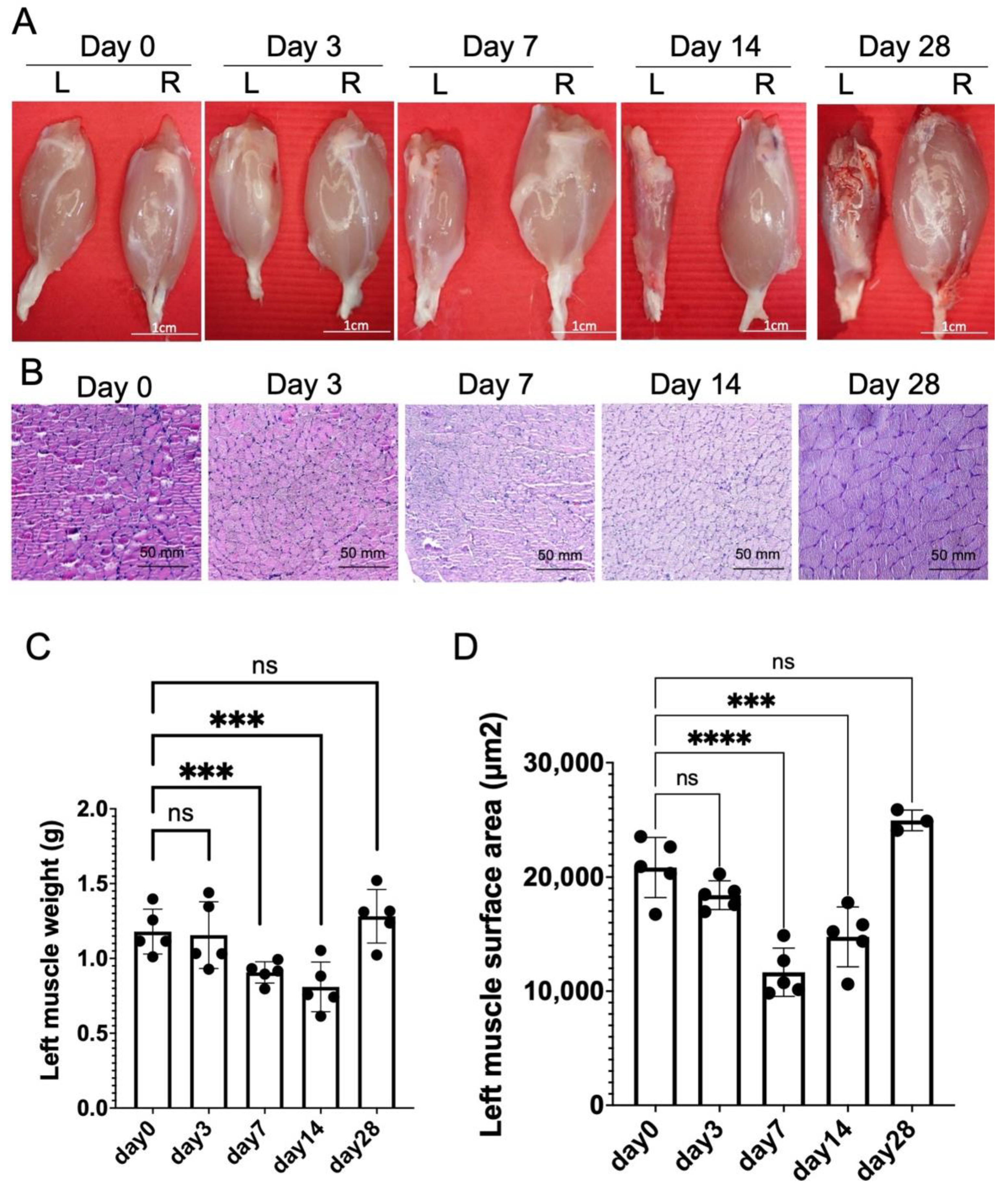
2.2. Chronic Constriction Injury for 1 Week Produces Significant But Reversible Muscle Atrophy
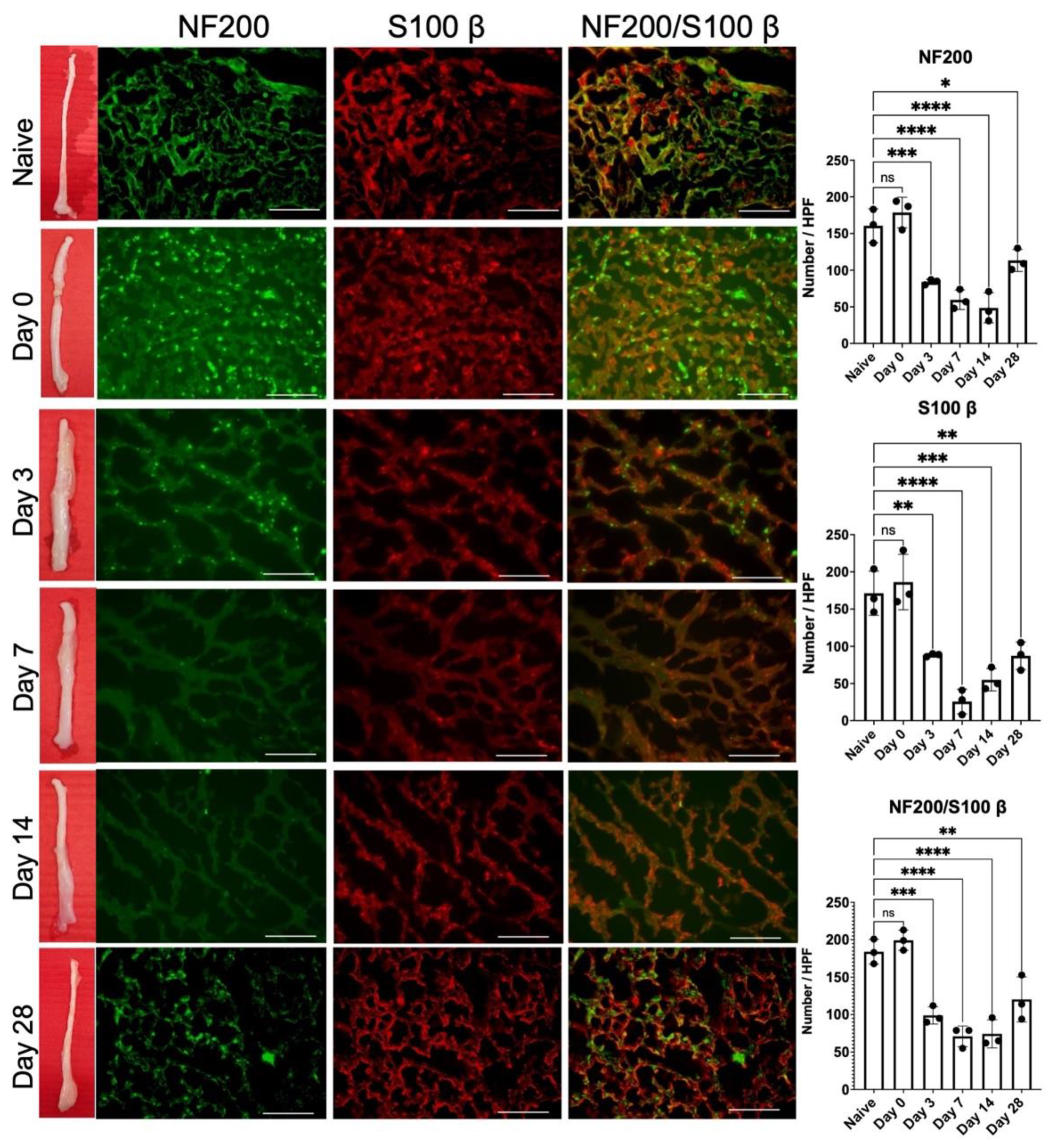
2.3. Release of 1-Week Constricted Nerve Induces Delayed Axon Remyelination
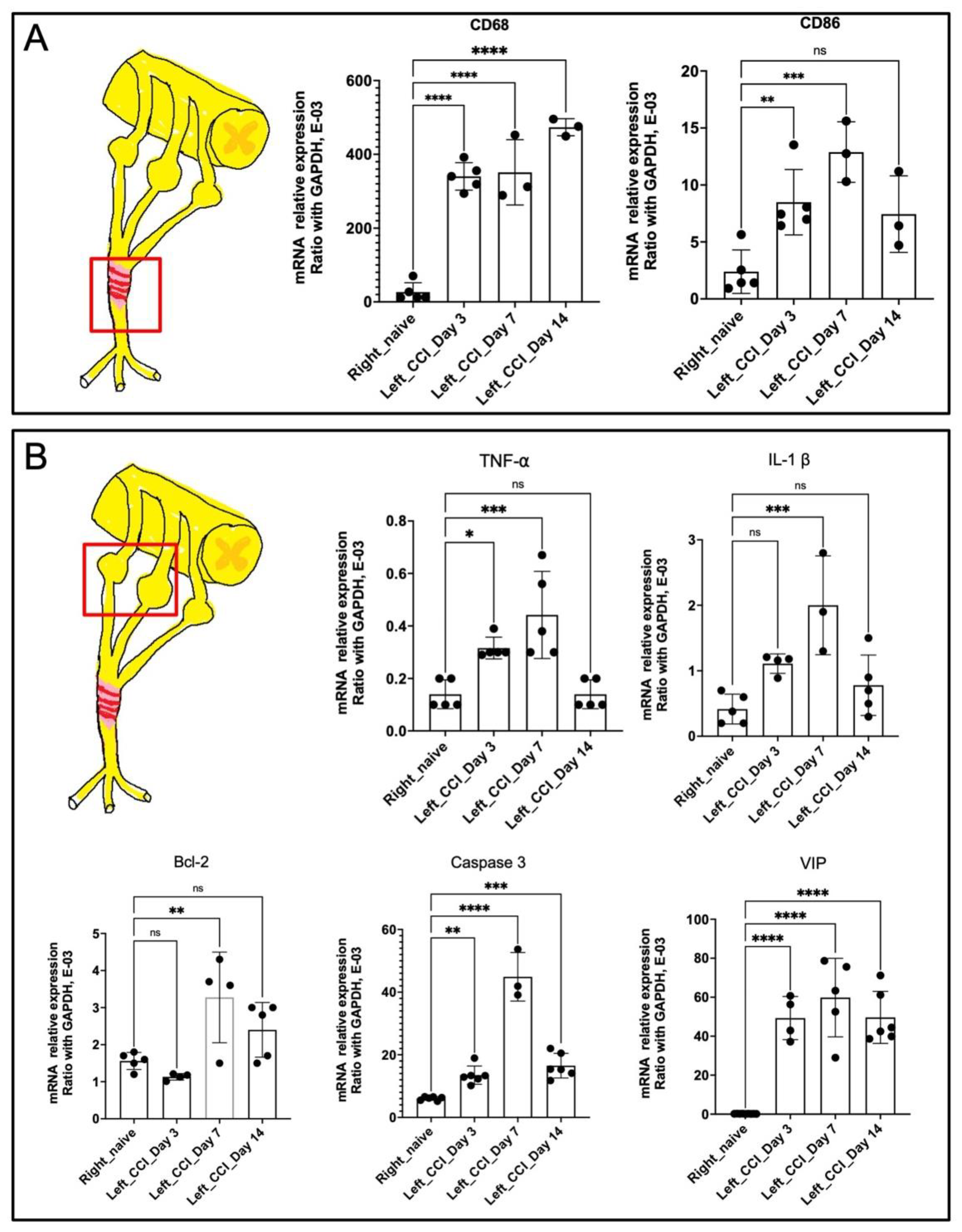
2.4. Release of 1-Week Constricted Nerve Demonstrated Prolonged Neuroinflammation of Injured Nerve and Dorsal Root Ganglion
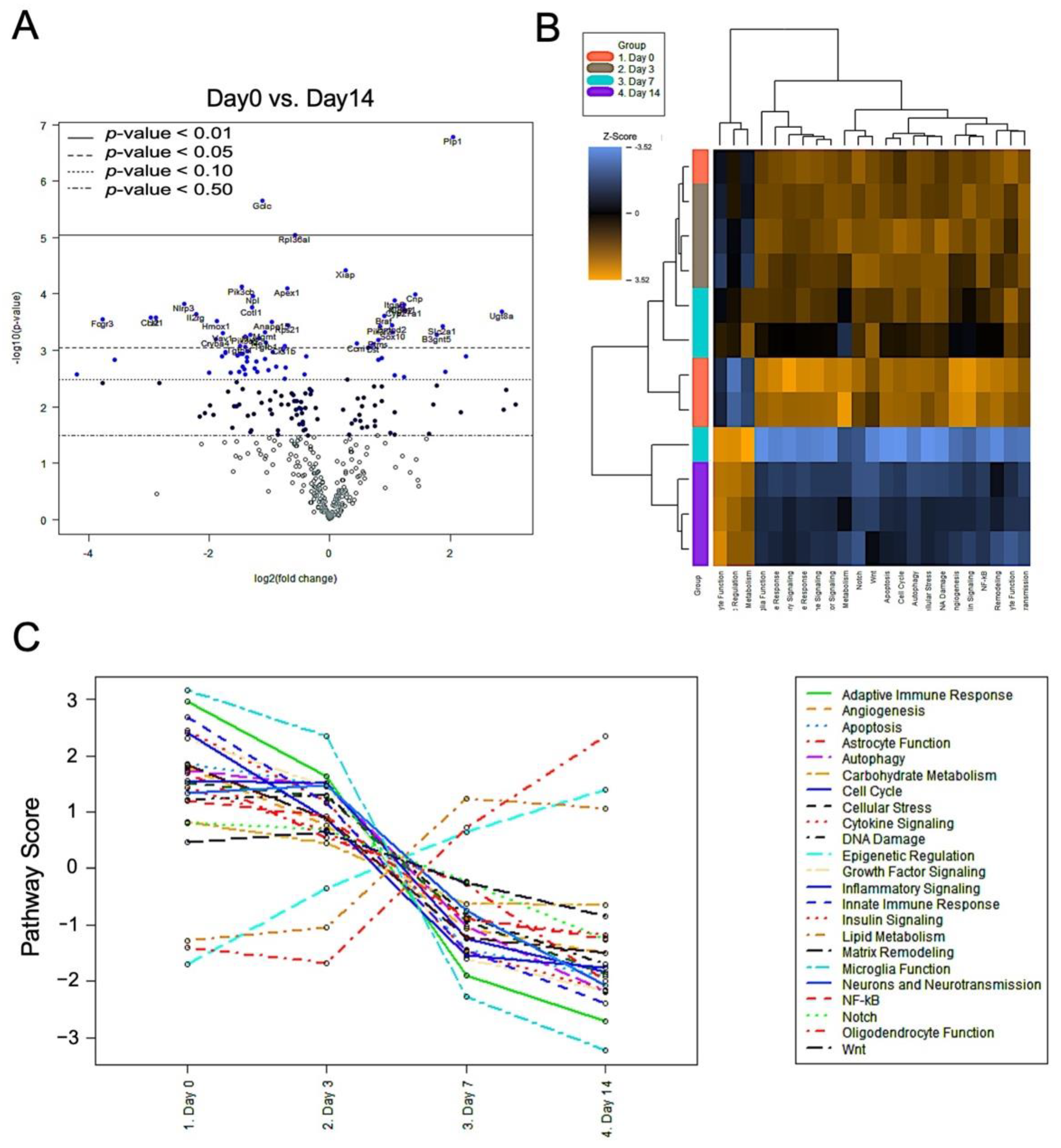
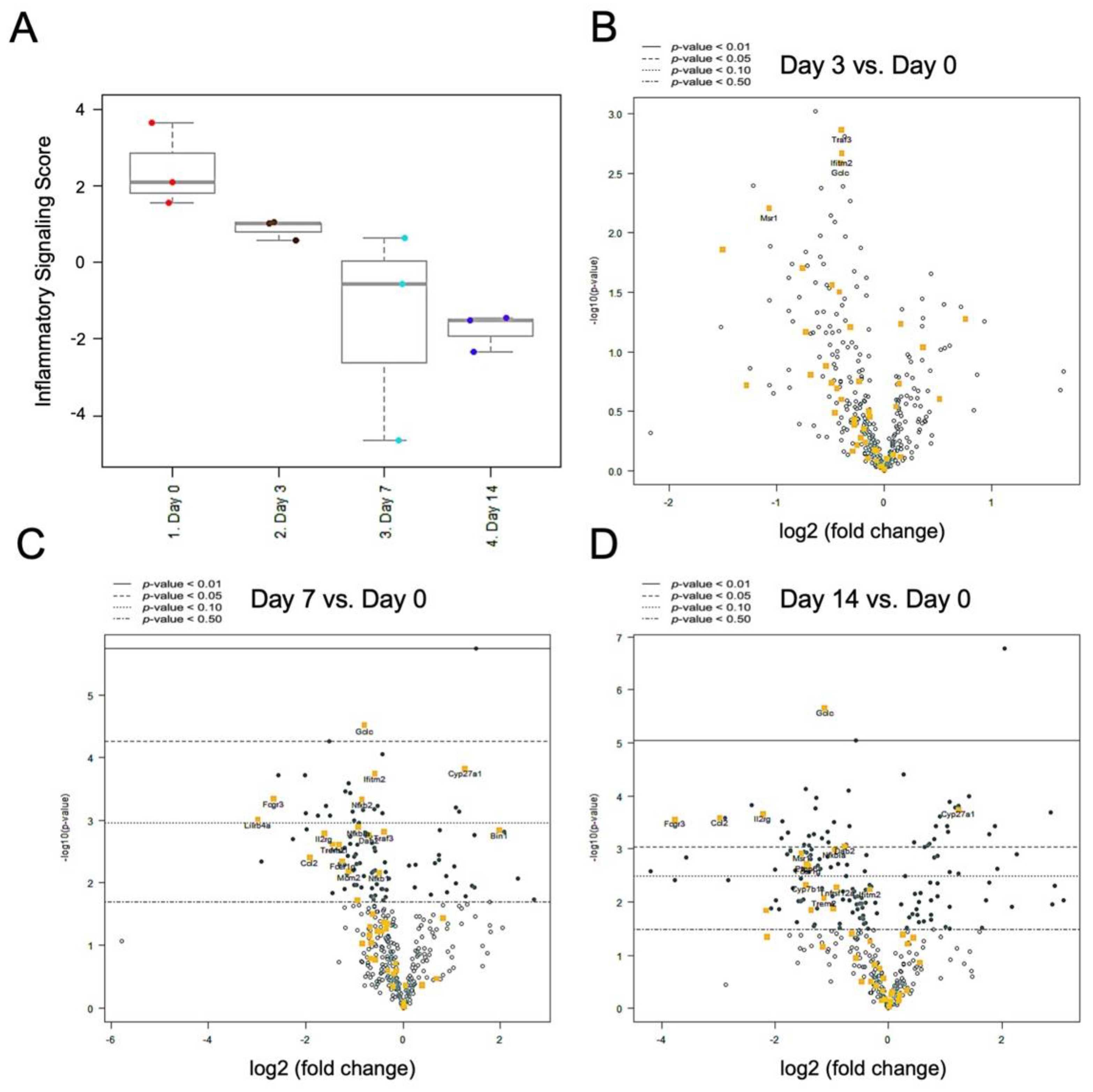
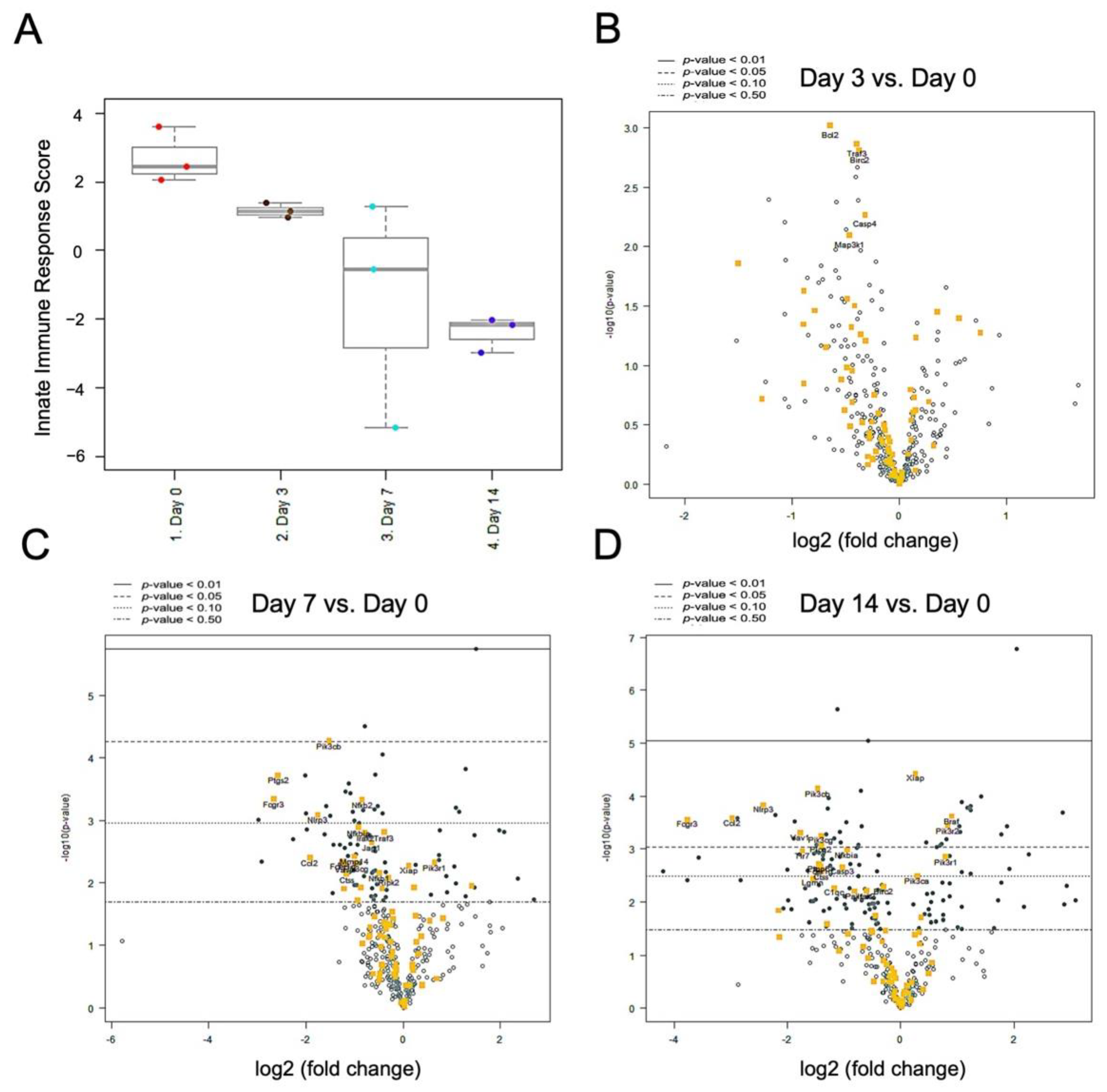
2.5. Nerve Release from CCI Revealed Bimodal Gene Regulation in Inflammatory and Innate Immune Profiling
3. Discussion
4. Materials and Methods
4.1. Animals and Surgical Procedures
4.2. Sensory Assessment
4.3. Wet Muscle Evaluation and Histological Analysis
4.4. Hematoxylin and Eosin Staining
4.5. Immunofluorescent Staining (IF)
4.6. RNA Preparation and Complementary DNA (cDNA) Synthesis
4.7. Quantitative PCR
| Name | Forward Primer/Reverse Primer |
| CD68 | 5′-CTGTTGCGGAAATACAAGCA-3′ 5′-GGCAGCAAGAGAGATTGGTC-3′ |
| CD86 | 5′-CCTCCAGCAGTGGGAAAC-3′ 5′-GTAGGTTTCGGGTATCCTTGC-3′ |
| TNF-α | 5′-TACTCCTCAGAGCCCCCAAT-3′ 5′-TCGTGTGTTTCTGAGCATCG-3′ |
| IL-1β | 5′-CTGTGACTCGTGGGATGATG-3′ 5′-TCCATTGAGGTGGAGAGCTT-3′ |
| BCl-2 | 5′-ACTCTTCAGGGATGGGGTGA-3′ 5′-TGACATCTCCCTGTTGACG-3′ |
| Caspase 3 | 5′-CCGACTTCCTGTATGCTTACTC-3′ 5′-CAGGGAGAAGGACTCAAATTC-3′ |
| VIP | 5′-CAGAAGCAAGCCTCAGTTCC-3′ 5′-GCCTGTCATCCAACCTCACT-3′ |
| GAPDH | 5′-TGGCCTCCAAGGAGTAAGAA-3′ 5′-TGTGAGGGAGATGCTCAGTG-3′ |
4.8. NanoString nCounter Technology for Gene Profiling
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Menorca, R.M.; Fussell, T.S.; Elfar, J.C. Nerve physiology: Mechanisms of injury and recovery. Hand Clin. 2013, 29, 317–330. [Google Scholar] [CrossRef] [Green Version]
- Abzug, J.M.; Jacoby, S.M.; Osterman, A.L. Surgical options for recalcitrant carpal tunnel syndrome with perineural fibrosis. Hand 2012, 7, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Hanewinckel, R.; Ikram, M.A.; Van Doorn, P.A. Peripheral neuropathies. Handb. Clin. Neurol. 2016, 138, 263–282. [Google Scholar]
- Assmus, H.; Antoniadis, G.; Bischoff, C.; Hoffmann, R.; Martini, A.K.; Preissler, P.; Scheglmann, K.; Schwerdtfeger, K.; Wessels, K.D.; Wüstner-Hofmann, M. Cubital tunnel syndrome–A review and management guidelines. Cent. Eur. Neurosurg. 2011, 72, 90–98. [Google Scholar] [CrossRef]
- Dableh, L.J.; Yashpal, K.; Henry, J.L. Neuropathic pain as a process: Reversal of chronification in an animal model. J. Pain Res. 2011, 4, 315–323. [Google Scholar]
- Yousefifard, M.; Rahimi-Movaghar, V.; Baikpour, M.; Ghelichkhani, P.; Hosseini, M.; Jafari, A.; Aziznejad, H.; Tafakhori, A. Early versus late spinal decompression surgery in treatment of traumatic spinal cord injuries; a systematic review and meta-analysis. Emergency 2017, 5, e37–e49. [Google Scholar]
- Rempel, D.; Dahlin, L.; Lundborg, G. Pathophysiology of nerve compression syndromes: Response of peripheral nerves to loading. J. Bone Jt. Surg. Am. 1999, 81, 1600–1610. [Google Scholar] [CrossRef]
- Mackinnon, S.E. Pathophysiology of nerve compression. Hand Clin. 2002, 18, 231–241. [Google Scholar] [CrossRef]
- Mahmoud, M.; El Shafie, S.; Coppola, E.E.; Elfar, J.C. Perforator-based radial forearm fascial flap for management of recurrent carpal tunnel syndrome. J. Hand Surg. Am. 2013, 38, 2151–2158. [Google Scholar] [CrossRef] [PubMed]
- Soltani, A.M.; Allan, B.J.; Best, M.J.; Mir, H.S.; Panthaki, Z.J. A systematic review of the literature on the outcomes of treatment for recurrent and persistent carpal tunnel syndrome. Plast. Reconstr. Surg. 2013, 132, 114–121. [Google Scholar] [CrossRef]
- Unglaub, F.; Wolf, E.; Goldbach, C.; Hahn, P.; Kroeber, M.W. Subjective and functional outcome after revision surgery in carpal tunnel syndrome. Arch. Orthop. Trauma Surg. 2008, 128, 931–936. [Google Scholar] [CrossRef]
- Spielman, A.F.; Sankaranarayanan, S.; Skowronski, P.; Lessard, A.S.; Panthaki, Z. Recurrent and persistent carpal tunnel syndrome: “Triple-therapy approach”. J. Orthop. 2020, 22, 431–435. [Google Scholar] [CrossRef]
- Sun, P.O.; Selles, R.W.; Jansen, M.C.; Slijper, H.P.; Ulrich, D.J.O.; Walbeehm, E.T. Recurrent and persistent carpal tunnel syndrome: Predicting clinical outcome of revision surgery. J. Neurosurg. 2019, 132, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Dubový, P. Wallerian degeneration and peripheral nerve conditions for both axonal regeneration and neuropathic pain induction. Ann. Anat. 2011, 193, 267–275. [Google Scholar] [CrossRef]
- Schmid, A.B.; Coppieters, M.W.; Ruitenberg, M.J.; McLachlan, E.M. Local and remote immune-mediated inflammation after mild peripheral nerve compression in rats. J. Neuropathol. Exp. Neurol. 2013, 72, 662–680. [Google Scholar] [CrossRef] [Green Version]
- Leung, L.; Cahill, C.M. TNF-alpha and neuropathic pain–A review. J. Neuroinflamm. 2010, 7, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.-H.; Huang, T.-C.; Wang, J.-Y.; Wu, C.-C.; Hsueh, Y.-Y. Controllable forces for reproducible chronic constriction injury mimicking compressive neuropathy in rat sciatic nerve. J. Neurosci. Methods 2020, 335, 108615–108624. [Google Scholar] [CrossRef]
- Bennett, G.J.; Xie, Y.K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef]
- Dijkstra, J.R.; Meek, M.F.; Robinson, P.H.; Gramsbergen, A. Methods to evaluate functional nerve recovery in adult rats: Walking track analysis, video analysis and the withdrawal reflex. J. Neurosci. Methods 2000, 96, 89–96. [Google Scholar] [CrossRef]
- Chen, L.E.; Seaber, A.V.; Glisson, R.R.; Davies, H.; Murrell, G.A.; Anthony, D.C.; Urbaniak, J.R. The functional recovery of peripheral nerves following defined acute crush injuries. J. Orthop. Res. 1992, 10, 657–664. [Google Scholar] [CrossRef]
- Pettersson, L.M.; Danielsen, N.; Dahlin, L.B. Altered behavioural responses and functional recovery in rats following sciatic nerve compression and early vs late decompression. J. Plast. Surg. Hand Surg. 2016, 50, 321–330. [Google Scholar] [CrossRef]
- Hu, J.; Tu, Y.; Ding, Z.; Chen, Z.; Dellon, A.L.; Lineaweaver, W.C.; Zhang, F. Alteration of sciatic nerve histology and electrical function after compression and after neurolysis in a diabetic rat model. Ann. Plast. Surg. 2018, 81, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Ceyhan, D.; Kocman, A.E.; Yildirim, E.; Ozatik, O.; Aydin, S.; Kose, A. Comparison of the Effects of Curcumin, Tramadol and Surgical Treatments on Neuropathic Pain Induced by Chronic Constriction Injury in Rats. Turk. Neurosurg. 2018, 28, 288–295. [Google Scholar] [CrossRef] [PubMed]
- George, A.; Marziniak, M.; Schäfers, M.; Toyka, K.V.; Sommer, C. Thalidomide treatment in chronic constrictive neuropathy decreases endoneurial tumor necrosis factor-alpha, increases interleukin-10 and has long-term effects on spinal cord dorsal horn met-enkephalin. Pain 2000, 88, 267–275. [Google Scholar] [CrossRef]
- Orhan, C.E.; Onal, A.; Ulker, S. Antihyperalgesic and antiallodynic effect of sirolimus in neuropathic pain and the role of cytokines in this effect. Neurosci. Lett. 2010, 481, 17–20. [Google Scholar] [CrossRef]
- Hu, Q.; Fang, L.; Li, F.; Thomas, S.; Yang, Z. Hyperbaric oxygenation treatment alleviates CCI-induced neuropathic pain and decreases spinal apoptosis. Eur. J. Pain 2015, 19, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, A.D.; Popovich, P.G.; Ramer, M.S. Wallerian degeneration: Gaining perspective on inflammatory events after peripheral nerve injury. J. Neuroinflamm. 2011, 8, 110–122. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.L.; Lassner, F.; Bader, A.; Becker, M.; Walter, G.F.; Berger, A. Cellular activity of resident macrophages during Wallerian degeneration. Microsurgery 2000, 20, 255–261. [Google Scholar] [CrossRef]
- Mueller, M.; Leonhard, C.; Wacker, K.; Ringelstein, E.B.; Okabe, M.; Hickey, W.F.; Kiefer, R. Macrophage response to peripheral nerve injury: The quantitative contribution of resident and hematogenous macrophages. Lab. Investig. 2003, 83, 175–185. [Google Scholar] [CrossRef]
- Griffin, J.W.; George, R.; Ho, T. Macrophage systems in peripheral nerves. A review. J. Neuropathol. Exp. Neurol. 1993, 52, 553–560. [Google Scholar] [CrossRef]
- Stevens, A.M.; Liu, L.; Bertovich, D.; Janjic, J.M.; Pollock, J.A. Differential Expression of Neuroinflammatory mRNAs in the Rat Sciatic Nerve Following Chronic Constriction Injury and Pain-Relieving Nanoemulsion NSAID Delivery to Infiltrating Macrophages. Int. J. Mol. Sci. 2019, 20, 5269–5292. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Eisenach, J.C. Four PGE2 EP receptors are up-regulated in injured nerve following partial sciatic nerve ligation. Exp. Neurol. 2003, 183, 581–592. [Google Scholar] [CrossRef]
- Martin, S.L.; Reid, A.J.; Verkhratsky, A.; Magnaghi, V.; Faroni, A. Gene expression changes in dorsal root ganglia following peripheral nerve injury: Roles in inflammation, cell death and nociception. Neural Regen. Res. 2019, 14, 939–947. [Google Scholar]
- Fregnan, F.; Muratori, L.; Simoes, A.R.; Giacobini-Robecchi, M.G.; Raimondo, S. Role of inflammatory cytokines in peripheral nerve injury. Neural Regen. Res. 2012, 7, 2259–2266. [Google Scholar]
- Momeni, H.R.; Soleimani Mehranjani, M.; Shariatzadeh, M.A.; Haddadi, M. Caspase-mediated apoptosis in sensory neurons of cultured dorsal root Ganglia in adult mouse. Cell J. 2013, 15, 212–217. [Google Scholar]
- Wiberg, R.; Novikova, L.N.; Kingham, P.J. Evaluation of apoptotic pathways in dorsal root ganglion neurons following peripheral nerve injury. Neuroreport 2018, 29, 779–785. [Google Scholar] [CrossRef]
- Fu, C.; Yin, Z.; Yu, D.; Yang, Z. Substance P and calcitonin gene-related peptide expression in dorsal root ganglia in sciatic nerve injury rats. Neural Regen. Res. 2013, 8, 3124–3130. [Google Scholar] [PubMed]
- Ma, W.; Bisby, M.A. Partial and complete sciatic nerve injuries induce similar increases of neuropeptide Y and vasoactive intestinal peptide immunoreactivities in primary sensory neurons and their central projections. Neuroscience 1998, 86, 1217–1234. [Google Scholar] [CrossRef]
- Sah, D.W.; Ossipo, M.H.; Porreca, F. Neurotrophic factors as novel therapeutics for neuropathic pain. Nat. Rev. Drug Discov. 2003, 2, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Pandak, W.M.; Erickson, S.K.; Ma, Y.; Yin, L.; Hylemon, P.; Ren, S. Biosynthesis of the regulatory oxysterol, 5-cholesten-3beta,25-diol 3-sulfate, in hepatocytes. J. Lipid Res. 2007, 48, 2587–2596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutemberezi, V.; Buisseret, B.; Masquelier, J.; Guillemot-Legris, O.; Alhouayek, M.; Muccioli, G.G. Oxysterol levels and metabolism in the course of neuroinflammation: Insights from in vitro and in vivo models. J. Neuroinflamm. 2018, 15, 74–89. [Google Scholar] [CrossRef]
- Kurschus, F.C.; Wanke, F. EBI2–Sensor for dihydroxycholesterol gradients in neuroinflammation. Biochimie 2018, 153, 52–55. [Google Scholar] [CrossRef] [PubMed]
- Testa, G.; Staurenghi, E.; Zerbinati, C.; Gargiulo, S.; Iuliano, L.; Giaccone, G.; Fantò, F.; Poli, G.; Leonarduzzi, G.; Gamba, P. Changes in brain oxysterols at different stages of Alzheimer’s disease: Their involvement in neuroinflammation. Redox Biol. 2016, 10, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Li, W.; Zheng, R.; Zhao, B.; Zhang, Y.; Zhao, D.; Zhao, C.; Yan, C.; Zhao, Y. Cerebrotendinous xanthomatosis with peripheral neuropathy: A clinical and neurophysiological study in Chinese population. Ann. Transl. Med. 2020, 8, 1372–1384. [Google Scholar] [CrossRef]
- Ginanneschi, F.; Mignarri, A.; Mondelli, M.; Gallus, G.N.; Del Puppo, M.; Giorgi, S.; Federico, A.; Rossi, A.; Dotti, M.T. Polyneuropathy in cerebrotendinous xanthomatosis and response to treatment with chenodeoxycholic acid. J. Neurol. 2013, 260, 268–274. [Google Scholar] [CrossRef]
- Liu, X.; Cui, X.; Guan, G.; Dong, Y.; Zhang, Z. microRNA-192-5p is involved in nerve repair in rats with peripheral nerve injury by regulating XIAP. Cell Cycle 2020, 19, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Huo, D.S.; Sun, J.F.; Cai, Z.P.; Yan, X.S.; Wang, H.; Jia, J.X.; Yang, Z.J. The protective mechanisms underlying Ginsenoside Rg1 effects on rat sciatic nerve injury. J. Toxicol. Environ. Health A 2019, 82, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Serrano, C.; Simonetti, S.; Hernandez, J.; Valverde, C.; Carles, J.; Bague, S.; Orellana, R.; Somoza, R.; Cajal, S.R.Y.; Romagosa, C. BRAF V600E mutations in benign and malignant peripheral nerve sheath tumors. J. Clin. Oncol. 2010, 28, 10043. [Google Scholar] [CrossRef]
- Kaplan, H.G.; Rostad, S.; Ross, J.S.; Ali, S.M.; Millis, S.Z. Genomic Profiling in Patients with Malignant Peripheral Nerve Sheath Tumors Reveals Multiple Pathways with Targetable Mutations. J. Natl. Compr. Cancer Netw. 2018, 16, 967–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, L.; Li, R.; Li, D.; Wang, B.; Lu, Y.; Li, P.; Yu, F.; Jin, Y.; Ni, X.; Wu, Y.; et al. FGF10 Enhances Peripheral Nerve Regeneration via the Preactivation of the PI3K/Akt Signaling-Mediated Antioxidant Response. Front. Pharmacol. 2019, 10, 1224–1236. [Google Scholar] [CrossRef]
- Eastel, J.M.; Lam, K.W.; Lee, N.L.; Lok, W.Y.; Tsang, A.H.F.; Pei, X.M.; Chan, A.K.C.; Cho, W.C.S.; Wong, S.C.C. Application of NanoString technologies in companion diagnostic development. Expert Rev. Mol. Diagn. 2019, 19, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, J.E.; Zygelyte, E.; Grenier, J.K.; Edwards, M.G.; Cheetham, J. Temporal changes in macrophage phenotype after peripheral nerve injury. J. Neuroinflamm. 2018, 15, 185–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-H.; Wu, C.-C.; Lin, S.-C.; Tseng, W.-L.; Huang, T.-C.; Yadav, A.; Lu, F.-I.; Liu, Y.-H.; Lin, S.-P.; Hsueh, Y.-Y. Investigation of Neuropathology after Nerve Release in Chronic Constriction Injury of Rat Sciatic Nerve. Int. J. Mol. Sci. 2021, 22, 4746. https://doi.org/10.3390/ijms22094746
Chen S-H, Wu C-C, Lin S-C, Tseng W-L, Huang T-C, Yadav A, Lu F-I, Liu Y-H, Lin S-P, Hsueh Y-Y. Investigation of Neuropathology after Nerve Release in Chronic Constriction Injury of Rat Sciatic Nerve. International Journal of Molecular Sciences. 2021; 22(9):4746. https://doi.org/10.3390/ijms22094746
Chicago/Turabian StyleChen, Szu-Han, Chia-Ching Wu, Sheng-Che Lin, Wan-Ling Tseng, Tzu-Chieh Huang, Anjali Yadav, Fu-I Lu, Ya-Hsin Liu, Shau-Ping Lin, and Yuan-Yu Hsueh. 2021. "Investigation of Neuropathology after Nerve Release in Chronic Constriction Injury of Rat Sciatic Nerve" International Journal of Molecular Sciences 22, no. 9: 4746. https://doi.org/10.3390/ijms22094746