Exploring the Lipidome: Current Lipid Extraction Techniques for Mass Spectrometry Analysis


Metabolomics Core Facility–MetCore, Vicepresidency for Research, Universidad de los Andes, Cra 1 No. 18A—12, Bogota 111711, Colombia
*
Author to whom correspondence should be addressed.
Metabolites 2020, 10(6), 231; https://doi.org/10.3390/metabo10060231
Submission received: 7 April 2020
/
Revised: 5 May 2020
/
Accepted: 13 May 2020
/
Published: 3 June 2020
(This article belongs to the Special Issue Mass Spectrometry-Based Metabolomics: Challenges and Applications)
Abstract
:In recent years, high-throughput lipid profiling has contributed to understand the biological, physiological and pathological roles of lipids in living organisms. Across all kingdoms of life, important cell and systemic processes are mediated by lipids including compartmentalization, signaling and energy homeostasis. Despite important advances in liquid chromatography and mass spectrometry, sample extraction procedures remain a bottleneck in lipidomic studies, since the wide structural diversity of lipids imposes a constrain in the type and amount of lipids extracted. Differences in extraction yield across lipid classes can induce a bias on down-stream analysis and outcomes. This review aims to summarize current lipid extraction techniques used for untargeted and targeted studies based on mass spectrometry. Considerations, applications, and limitations of these techniques are discussed when used to extract lipids in complex biological matrices, such as tissues, biofluids, foods, and microorganisms.
1. Introduction
The term lipids generally refers to amphiphilic organic molecules, poorly soluble in water but miscible in organic solvents. Classification and study of lipid species is challenging due to the large chemical and structural diversity, including different hydrocarbon backbone lengths, branching, unsaturations, and functional groups. Since 2005, the International Lipid Classification and Nomenclature Committee (ILCNC) on the initiative of the Lipid Metabolites and Pathways Strategy (LIPID MAPS) consortium defined lipids as “hydrophobic or amphipathic small molecules that originate entirely or in part by carbanion-based condensations of thioesters and/or by carbocation-based condensations of isoprene units” [1,2,3]. Current lipid classification involves eight categories based on chemical functionalities as: (1) glycerolipids (GL), (2) sphingolipids (SP), (3) glycerophospholipids (GP), (4) sterol lipids (ST), (5) fatty acyls (FA), (6) prenol lipids (PR), (7) polyketides (PK), and (8) saccharolipids (SL), where the last two categories are not synthesized by mammals and represent a small proportion of the known lipidome [1,2,3]. Table 1 presents the number of lipid structures per category according to Lipid Maps® Structure Database (LMSD) and Figure 1 shows representative structures for each category.
Once viewed as mere membranes constituents and energy storage reservoirs, nowadays lipids are also recognized for playing crucial roles in diverse biological activities at cellular and systemic levels including: cell signaling, transport, protein trafficking, growth, differentiation, and apoptosis [3,4]. To accomplish these myriad of functions, cells produce lipids with a vast structural complexity, along with a differentiated compartmentalization, location, organization and interaction [5]. Consequently, a particular set of lipids—known as lipidome—characterize each cell, tissue, and biological system [4].
Lipidomes are often are complex mixtures of lipids, with diverse chemical structures that represent the different biological microenvironments where lipids normally play their function in vivo. Therefore, lipidomes are highly susceptible to changes in response to physiological, pathological, and environmental conditions and can indicate an organism status in a particular moment [6]. In fact, abnormalities in the metabolism of lipids have been linked to several human pathologies (e.g., Alzheimer’s disease [7], cancer [8], diabetes [9]), stress response in plants [10] and antibiotic resistance in infectious bacteria [11,12]. For this reason, the study of lipids has represented a valuable tool to elucidate mechanistic insights into all kingdoms of life.
The main analytical platforms for lipid analyses include mass spectrometry (MS) and nuclear magnetic resonance (NMR), where MS-based techniques have been widely used due to their high sensitivity (pM concentrations), availability and speediness in accurate identification, quantification and monitoring of basal lipid profiles in complex biological mixtures [13]. Sample preparation for MS-lipidomics usually includes solvent–protein precipitation, lipid extraction, and solvent evaporation. The initial step of protein precipitation aims to eliminate matrix components that could interfere with the precision and accuracy of the mass analysis, such as proteins and salts. The subsequent step of lipid extraction takes advantage of the hydrophobic properties of lipids to separate them in a non-polar solvent system with or without mechanical assistance (e.g., vortex, microwave, ultrasound). Finally, solvent evaporation allows lipid enrichment and resuspension in a compatible solvent for MS equipment, typically hyphenated to chromatographic separation [14].
Considering the analytical challenge of extracting hundreds of lipidic compounds with a wide range of polarities and concentration levels, there is not a unique method suitable to extract an entire lipidome. Thus, the choice of a particular lipid-extraction protocol must account for its inherent limitations and be tailored to a specific biological matrix, analysis approach and experimental design. The importance of this choice lies in the profound impact on the class of lipids that can be detectable and measured, which could create a bias on the subsequent analysis and findings. In general, two main approaches are used to analyze lipidomes: (1) non-targeted analysis, also known as hypothesis-generating, to simultaneously extract all detectable lipids in a sample regardless of class, concentration or prior identification, for which non-selective methods are used; (2) targeted analysis, or hypothesis-driven, to selectively extract a particular set of known lipids, in order to avoid interferences and enhance extraction efficiency [15].
Furthermore, repeatability and reproducibility during extraction should account for the lack of analyte-specific internal standards to asses lipid concentrations. In lipidomics, and metabolomics in general, multiple samples are analyzed under equivalent conditions to allow further comparison of analyte levels between different groups (e.g., control vs. disease) using univariate and multivariate statistical analysis. For this reason, biological variations most prevail over analytical and random variations in order to avoid data misinterpretation. In light of the vast diversity of lipids structures and its determining role in lipid extraction, this review summarizes current extraction protocols used for isolation of lipid species present in complex biological matrices, including microorganisms, biofluids, plant and animal tissues, and foods. Technical details concerning both, untargeted and targeted approaches, are discussed along with limitations and considerations of lipid extraction protocols employed in MS-based lipidomics.
2. Non-Targeted Analysis
Lipid extraction is without a doubt the major limiting step to analyze the complete set of lipids in biological systems under an untargeted approach. Liquid–liquid extraction (LLE) protocols based on chloroform/methanol mixtures, such as Folch [16] or Bligh and Dyer (BD) [17], have been widely used since the late 1950s with few modifications and still represent the benchmark in the field. Alternative LLE protocols include Methyl tert-butyl ether (MTBE) [18] and Butanol-methanol (BUME) [19] with comparable outcomes for lipid isolation in plasma samples [20,21]. The success of these protocols relies on exploiting the characteristic amphipathic properties of lipids to achieve a differential partition between an aqueous phase and an immiscible organic phase at given temperature, pH and ionic strength. In LLE protocols, separation in two phases is achieved by the hydrophobic and hydrophilic interactions within a system. Hydrophobic interactions are mediated by aliphatic and cyclic hydrocarbon backbones, while hydrophilic ones by polar groups, such as phosphates or carbohydrates. Lipids can also be isolated into one-phase systems, using either a single or a combination of miscible organic solvents for protein precipitation (PPT) and lipid solubilization. Methanol [22], isopropanol [23], and acetonitrile [24] have been proposed as organic systems for untargeted lipidomics.
Solid phase extraction (SPE) is also a feasible option for lipid extraction. It is based on a partition equilibrium involving the adsorption of lipids to a solid phase, which preconcentrates the hydrophobic compounds prior to desorption. Removal of interfering compounds and impurities is achieved by using SPE cartridges, commonly reverse-phase ones. The process comprises of cartridge conditioning, followed by sample loading, cartridge washing and elution. During these stages, aliphatic backbones in lipid structures interact with non-polar stationary phases (e.g., C8, C18) and are retained until an elution solvent is added. Although, SPE protocols are occasionally used for untargeted approaches, they are more typically used for targeted lipidomics. Other strategies for non-targeted lipid extraction are tailored to specific matrices (e.g., plants, cell lines), sampling (e.g., microextraction), and include the use of mechanical assistance like microwaves (MAE), ultrasound (UAE) among others [25]. Reports using solid phase microextraction (SPME) for comprehensive lipid analysis are also worth mentioning [26,27]. SPME employs sorbent-coated rods/fibers where lipids migrate and diffuse directly from a headspace or liquid, to the fiber or rod, eliminating the washing step required on SPE. Then, thermal or solvent desorption is used for lipid elution and analysis by gas chromatography-mass spectrometry (GC-MS) or liquid chromatography-mass spectrometry (LC-MS) respectively [28].
As shown in Figure 1, chemical structures across lipid classes display a large variety of polarities. Encompassing more than 10 units of log P in the octanol–water coefficient as a polarity index, the wide polarity window of lipid species is much wider than any solvent or solvent mixture can cover [15]. In consequence, none of the current protocols is capable to extract all lipid classes simultaneously with high recovery (>80%), and usually the yields of specific compounds are typically sacrificed at the expense of a wider coverage [13]. For instance, chloroform/methanol protocols can extract all lipid classes with recoveries higher than 50%, but present low recoveries for charged and non-polar lipids, like phosphatidic acids (PA) and lysophospholipids (LPA) in plasma [23,29].
Furthermore, the differential recovery across lipid species can interfere with the analysis to different extents. For example, a sub estimation of ST in cerebrospinal fluid is feasible due to its low concentration compared to major constituents FA and SP [31]. In contrast, a similar analysis in plasma would not have a significant impact on ST, considering the its relatively high concentration in that matrix. However, estimation of low-abundant FA could be affected, particularly oxylipins in the pM range [32]. For this reason, it is recommended to explore different extraction protocols if previous reports on the matrix of interest are not available. Crucial factors to be optimized include solvent system and sample-solvent ratio. A good example for method selection and validation can be found in the work of Van Meulebroek et al. [33].
One might tackle the low recovery limitation proposing sequential extraction steps, by using either the same or a complimentary solvent mixture. However, more extraction steps can introduce a higher systematic variability and increase the time of the procedure, which play a crucial factor in large-scale study settings. Furthermore, extraction protocols tend to be minimal in order to preserve sample integrity and content. Non-selective liquid extraction protocols, such as LLE are preferred to extract lipids, since these avoid harsh conditions that can lead to degradation products or cross-products. Nonetheless, there are some concerns about reproducibility of LLE protocols, in particular when MTBE is used given its high volatility [34]. Therefore, single-phase [23,34,35] and even triphasic systems [36] have been proposed to achieve enhanced reproducibility and lipid coverage. Further validation of these methodologies is still required. Table 2 presents a contrast of current lipid extraction protocols used for untargeted MS-based studies of biological matrices. Different parameters were taking into account for the comparison, including coverage, recovery, reproducibility and automatization. Note that lipidomic studies using matrix-assisted laser desorption-ionization (MALDI) as ionization source were not included in this review (recently reviewed by Leopold et al. [37]), since little or no sample preparation is needed.
In recent years, automatized lipid extraction has been proposed to decrease the experimental variability between samples in large batches. Not all solvent–extraction systems are suitable for this task, since some of them are prone to contamination with proteins and other matrix components. This is the case of chloroform/methanol protocols, where in order to get access to the non-polar fraction at the bottom, the injection needle must break through a protein interphase and a polar fraction at the top. In contrast, MTBE and BUME methods use a low-density extraction solvent that locates the hydrophobic fraction at the top of the partition system, there it can be easily sampled by an automatized needle. Moreover, solvent compatibility with the MS system should be considered. For instance, chloroform is well-known for being problematic with liquid chromatography (LC)-MS and it has to be removed by a previous injection. One-phase extraction (OPE) and SPE protocols could also be automatized for high throughput analysis. A recent review by Liu et al. covers the details and advances in analytical methods for MS-based large-scale lipidomics [73].
Another consideration is the high sensitivity of MS instruments, which could play a double-edged sword role in the mass-to-charge analysis of lipid extracts. On the one hand, small sample amounts are enough to detect femtomolar and quantify picomolar concentrations of lipid species. Usually only 10–100 µL of liquid sample or 1–100 mg of solid sample is required for lipid extraction. Then, 5–20 µL of extract are subsequently taken for either LC injection (LC-MS based lipidomics) or direct infusion into an MS (also called shotgun lipidomics). On the other hand, multi-sourced trace impurities coming from biological matrices (e.g., remaining proteins), solvents, preparation devices (e.g., siloxenes and phthalates), and even sample containers (e.g., plasticizers) can also be detected if carried over in the lipid extract.
Overall, the presence of contaminants excerpts an effect on ionization (either enhancement or suppression) and can lead to deterioration of MS instrumentation. This issue is not as critical in LC-MS methods as in shotgun lipidomics, since LC allows separation between lipids and contaminants. Consequently, HPLC (high-performance liquid chromatography) and UPLC (ultra-performance liquid chromatography) are commonly preferred for non-directed lipidomics at the expense of increased solvent consumption and analysis time. Recently, SFC (supercritical fluid chromatography) have been successfully applied as a sample separation step to overcome these drawbacks [74].
Although, a comprehensive discussion of sampling and sample processing falls outside the scope of this review, general guidelines for handling biological matrices are implemented to preserve lipidome integrity. These measures include sample storage at −80 °C, avoid freeze–thaw cycling, and short processing times at 4 °C to minimize unwanted enzymatic and chemical processes. However, some lipid species (e.g., oxylipins, polyphenols, lysoGP) require particular measures due to their susceptibility to oxidation and isomerization. For this reason, antioxidants such as, butylated hydroxytoluene (BHT), butylated hydroxyanisole (BHA), or triphenylphosphonium (TPP), along with buffers are commonly included prior sample homogenization and extraction. Additionally, the use of MS grade solvents, glass vials and glass pipetting tips significantly decrease the incorporation of contaminants.
Another important limitation of any current extraction protocol is the inherent homogenization of lipids coming from different sub-compartments (e.g., tissue region, cell type, organelles). The disruption of in vivo interactions and subsequent reorganization of lipids by hydrophobicity not only hinders the chance to localize and monitor its dynamic changes, but also can impact its stability and reactivity [13]. Therefore, conclusions from these studies most acknowledge this limitation in the biological interpretation of their outcomes.
Overall, lipidomic studies can be powerful hypothesis-generating tools. Extraction conditions should be adjusted and validated for the biological matrix of interest. General guidelines include testing of repeatability, reproducibility, and recovery using at least one representative compound per lipid class. The step by step workflow for protocol selection proposed by Furse et al. [75] can serve as a starting point.
3. Targeted Analysis
For studies focused on a specific subset of lipids, the particular features in their chemical structures drive the selection of the extraction protocol. Isolation and concentration of lipids of interest is achieved by either LLE with narrow polarity windows or SPE cartridges. Compared with untargeted approaches, targeted analysis allows higher sample amounts due to the selective procedures used for enrichment of a particular lipid species. For instance, to study low-abundant oxylipins in plasma, up to 250 µL of the sample are extracted, which represents ten times the typical amount used for untargeted approaches [14].
In general, compounds with a high hydrophobicity index, such as triacylglycerols (TG), diacylglycerols (DG), cholesteryl esters and fatty esters, are commonly extracted with LLE. Non-polar solvents (e.g., cyclohexane, toluene) are preferred over moderately polar solvent mixtures (e.g., chloroform, MTBE) since only the most hydrophobic lipids dissolve in them [75].
The extraction of intermediate hydrophilic species, such as GP and SP, is usually achieved by LLE using polar solvents. Chloroform-based protocols are the most common, followed by a rising popularity of MBTE-based ones. Considering the recovery disparities across lipid species, these protocols are often modified to achieve higher extraction yields, including changes in solvent system, solvent proportions (%v/v), and introduction of mechanic forces. When charged groups are present, another important variable is the pH, since the acid-base character of certain lipids can be used for its extraction and enrichment. For instance, phosphatidic acids (PA), phosphatidylserines (PS) [76] and phosphatidylinositols (PI) [77] can be efficiently extracted using mild acidic conditions. However, pH changes must be optimized to avoid structural re-arrangements in the presence of nucleophiles, given the high electrophilicity of phosphate groups [78].
Finally, isolation of polar lipids has less standardized protocols given both, the similarities in physicochemical properties to common non-lipid metabolites and the complex structure of these compounds (e.g., acylaminosugars, cerebrosides). Typical approaches use water, methanol, or pyrimide as polar solvents in combination with additives [75]. When SPME is used, derivatization reagents are incorporated into the SPME fibers to improve specificity and sensitivity. Table 3 provides a revision of recent lipidomic targeted studies in different biological matrices. A discussion for each lipid category is also presented.
3.1. Glycerolipids (GL)
GL are neutral lipids composed of one glycerol backbone attached to at least one FA via ether or ester linkage. Structural classification is mainly based on the number of FA bound. Mono-, di-, and triradylglycerols are typically extracted with apolar organic solvents, including octane, cyclohexane, and MTBE. An alternative approach includes supercritical CO2 for high-throughput separation of GL [165]. Pure chloroform and mixtures with methanol have also been reported [84,104,106,108]. Selective isolation of these hydrophobic species is achieved with low degradation, under mild conditions used in the extraction and minor reactivity of GL. Glycosylglycerols are another important subgroup in this class, which are characterized by sugar residues linked to the backbone. BD and Folch methods have been reported for the analysis of glycosylglycerols, such as digalactosyldiacylglycerols [110] and seminolipids [166].
3.2. Sphingolipids (SP)
The common backbone in SP consists of an amino alcohol, known as sphingoid base. Structural diversity arises from different chemical moieties linked to the amino and hydroxy groups. Important members of this family include ceramides, phosphospingolipids, cerebosides, and gangliosides. An up-to-date review by Montefusco et al. summarizes several experimental considerations for quantitative and qualitative analysis of sphingolipids [167]. In general, reports are consistent in comparable SP recoveries using chloroform-methanol-based and MTBE methods, with electrospray (ESI) as ionization source in positive and negative modes [21,23,62]. Mild alkaline and acidic extraction has also been combined with classic LLE protocols. Addition of MeONa [125] and KOH [126] has shown improved recovery and reproducibility in SP extraction, particularly ceramides and phosphosphingolipids. A standardized protocol for SP profiling is proposed by Sullards et al. [126]. For gangliosides, a quantitative approach involves derivatization using 4-(4, 6-Dimethoxy-1, 3, 5-triazin-2-yl)-4-methylmorpholinium chloride (DMTMM) and 2-(2-Pyridilamino)-ethylamine (PAEA) after protein precipitation [131]. The addition of a pyridylamine group increases the ionization efficiency resulting in a 15-fold signal intensity when compared to previously reported methods.
3.3. Glycerophospholipids (GP)
GP basic structure contains a glycerol base linked to one or two FA at the sn-1 and sn-2 positions. A phosphate group is also attached to the sn-3 position. Different bonding (acyl-, alkyl-, or alkenyl), length and unsaturation of the FAs create a wide range of combinations. However, GP are commonly subclassified based on the head group attached to the phosphate. Neutral phosphatidylcholine (PC) and phosphatidylethanolamine (PE) species, and charged ones including phosphatidic acid (PA), phosphatidylglycerol (PG), phosphatidylinositol (PI), phosphatidylserine (PS) and cardiolipins (CL) have precise locations and functions within cell membranes [168]. Most of neutral GP are successfully extracted with traditional procedures using chloroform or MTBE with comparable yields, except for LPCs [21,169]. For polar GPs like PI and LPA acidic conditions (pH 4–6) during extraction are recommended [170]. Tris or citric acid buffers are preferred for this purpose to avoid hydrolysis and interconversion of GP [119]. OPE protocols have also shown improved recoveries in polar GP, particularly PG and PS [23]. As alternative, specific SPE cartridges are commercially available for deproteinization and phospholipid preconcentration, such as HybridSPE-Phospholipid Ultra cartridge® [171] and iSPE®, with reported capability to isolate low-abundant GP [169,172,173].
3.4. Sterol Lipids (ST)
The structural skeleton of ST is based on a four fused ring. Differences in conjugation and position/type of polar functional groups generate a broad spectrum of polarities in this family. Lipid analysis in mammalian systems have focused on steroids, cholesterol and bile acids, while plants systems focus on phytosterols and phytostanols. Thin layer chromatography–flame ionization (TLC-FID) and GC-MS techniques were initially used to determine ST with the requirement of derivatization (e.g., trimethylsilation and methylation). Nonetheless, an increasing number of publications have been implemented LC-MS strategies, since ST analysis can be performed in native state. Commonly, PPT or LLE methods are used for lipid extraction prior SPE in order to avoid interferences and signal suppression. Appropriate SPE elution solvents can resolve cholesterol from derivates like oxysterols and steroids given the differences in number of hydroxyl groups attached to the core ring. Moreover, antioxidants and metal chelators are commonly added to prevent ST oxidation during the extraction procedure [174]. For ST in plants, n-hexane and supercritical CO2 have shown the best yields when combined with mechanical forces [175]. If esterified ST are targeted, a saponification stage under mild conditions can be included to remove fatty acyl groups without generation of artifacts [176].
3.5. Fatty Acyls (FA)
The structure of FA contains repeated series of methylene groups, derived from successive additions of malonyl-CoA or methylmalonyl-CoA to an acetyl-CoA primer. Differences in carbon chain lengths, degree of oxidation, unsaturation, and cyclations are the main sources of structural diversity. Major subclasses include fatty acids, fatty esters, and eicosanoids. Fatty acids have been traditionally derivatized for characterization by GC–MS. However, LC-MS and SFC-MS approaches provide quantitative signals for most fatty acids in biological samples. This allows the use of faster and milder conditions during lipid extraction, like BD and MTBE protocols. For extraction of eicosanoids, further measurements must be taken to avoid lipid degradation and artifact generation. Cold conditions (4 °C) and addition of antioxidants (e.g., BHT, BHA) are commonly introduced to preserve FA integrity within a sample. In a recent review on oxylipin extraction, Liakh et al. contrast PPT, LLE, and SPE methods. A combination of LLE previous SPE seems to be the best approach for preconcentration of oxylipins and other related species [177].
3.6. Prenol Lipids (PR)
PR carbon backbones comprise one or more isoprene units condensed. The number of terpene units and oxidation state of the structure form the basis of their classification. Isoprenoids, quinones and polyprenols are representative members of this class. Besides the well-known derivatization for GC-MS analysis of PR [178], recent OPE and LLE have been introduced for LC-MS methods. One-phase solvent systems containing diethyl ether, methanol, propanol and water are generally employed for the extraction, due to the relative polar character of PR in comparison to other bulk lipid classes. For higher yields of apolar PR like carotenoinds and tocopherols, either hexane or chloroform is added [148,179]. Supercritical fluids have also shown potential to efficiently extract and separate these compounds as reported in food and human serum studies [180]. Given their physicochemical similarities, ST are commonly co-extracted with PR, even after saponification. However, conventional ST precipitation with petroleum ether [181] or SPE enrichment [151] could be used to achieve separation of PRs. Additionally, extraction conditions such as neutral or slightly basic pH, presence of antioxidants and absence of light improve extraction yield and minimize PR degradation.
3.7. Polyketides (PK)
PK are a structural diverse lipid class derived from sequential condensations of ketoacyl groups. PK are characterized by backbones with at least two carbonyls linked by a carbon atom, and also subject to functional modifications including hydroxylation, glycosylation, methylation among others. Further subclassification include macrolides, aromatic polyketides, and flavonoids. The abundance of polar groups and double bounds in PK structures allow the use of polar solvents to narrow down the type of lipids to be extracted. Targeted studies report either ethyl acetate or methanol as OPE solvents. Considering that plants, fungi, and bacteria are the main biological sources of PK, mechanical forces are commonly applied during the extraction. Pressurized systems with [182] or without [183] supercritical fluids increase significantly the PK recovery. SPE methods have also been adopted for PK fractionation and preconcentration using reverse phase cartridges (e.g., Strata™-X(Torrance, CA 90501-1430, USA), Sep-Pak Plus C18), particularly for prymnesins [163] and flavonoids [184]. Other important parameters must be optimized for the extraction, including sample amount, sample-solvent ratio and temperature. Finally, extraction of PK prone to oxidation (e.g., polyphenols) can incorporate β-mercaptoethanol or K2S2O5 to decrease unwanted redox processes.
3.8. Saccharolipids (SL)
SL are distinguished by a sugar backbone linked to FAs via glycosidic bonds. Representative members of this class include lipopolysaccharides (LPS) synthesized by gram-negative bacteria. Several methods have been developed for LPS extraction based on the analytical technique for detection and quantification. For MS studies, phenol-based methods are the standard for SL extraction, including hot phenol extraction (HPE), aqueous phenol-chloroform and aqueous phenol-diethyl ether extraction [185]. Recently, alternative at-room-temperature protocols have been tested to improve efficiency, yield, and lipid integrity. SPE [156] and methanol-chloroform [155] methods are reported in cell membrane characterizations.
SL are distinguished by a sugar backbone linked to FAs via glycosidic bonds. Representative members of this class include lipopolysaccharides (LPS) synthesized by gram-negative bacteria. Several methods have been developed for LPS extraction based on the analytical technique for detection and quantification. For MS studies, phenol-based methods are the standard for SL extraction, including hot phenol extraction (HPE), aqueous phenol-chloroform and aqueous phenol-diethyl ether extraction [185]. Recently, alternative at-room-temperature protocols have been tested to improve efficiency, yield, and lipid integrity. SPE [156] and methanol-chloroform [155] methods are reported in cell membrane characterizations.
4. Conclusions and Future Perspectives
The high-throughput analysis of lipids using mass spectrometry provides a snapshot into complex lipidomes in living organisms. However, lipid identification and quantification are limited by the selected extraction protocol. Structural features not only drive the intermolecular interactions within an extraction system, but also dictates their stability and reactivity. Furthermore, type and amount of biological matrix should be well considered for protocol selection and validation. Despite the efforts of several researchers, the lack of standardized procedures for sample preparation in lipidomics is still a major concern in the field. Therefore, comparative studies contrasting two or more different isolations techniques, along with inclusion of extraction protocols in lipid databases, provide valuable insights and criterion.
Preservation of lipidome integrity must be prioritized through extraction procedures, otherwise lipid extracts would not faithfully retain biological variations and comparisons across samples will be inaccurate. Automatized lipid isolation represents a feasible solution by taking advantage of simple and versatile protocols. Although not all samples are suitable subjects of automatization, advances in SPME-LC online interfaces and combined extraction of amphiphilic and lipophilic compounds in glass-coated microplates are promising.
For untargeted approaches, the main limitations are the narrow coverage and differential yield of extracted lipid species. Meanwhile, repeatability and reproducibility through several extraction steps are constraints faced by targeted approaches. Future lipidomic studies will account for lipid sub-cellular localization, interaction with physiological partners and monitor concentrations changes over time using fast, reproducible, and versatile extraction protocols to assure an accurate depiction of the dynamic changes taking place in biological systems.
Author Contributions
Conceptualization, J.A. and M.P.C.; literature review, J.A., A.R.-O. and M.P.C.; writing—review and editing J.A and M.P.C. All authors accepted the final version of the review. All authors have read and agreed to the published version of the manuscript.
Funding
The APC was funded by Metabolomics Core Facility-MetCore at the Univerisidad de Los Andes a facility that is supported by the vicepresidency for research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fahy, E.; Subramaniam, S.; Brown, H.A.; Glass, C.K.; Merrill, A.H.; Murphy, R.C.; Raetz, C.R.H.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A Comprehensive Classification System for Lipids. J. Lipid Res. 2005, 46, 839–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.H.; Shimizu, T.; Spener, F.; Meer, G.; van Wakelam, M.J.O.; Dennis, E.A. Update of the LIPID MAPS Comprehensive Classification System for Lipids. J. Lipid Res. 2009, 50, S9–S14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahy, E.; Cotter, D.; Sud, M.; Subramaniam, S. Lipid Classification, Structures and Tools. Biochim. Biophys. Acta - Mol. Cell Biol. Lipids 2011, 1811, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Gross, R.W. Shotgun Lipidomics: Electrospray Ionization Mass Spectrometric Analysis and Quantitation of Cellular Lipidomes Directly from Crude Extracts of Biological Samples. Mass Spectrom. Rev. 2005, 24, 367–412. [Google Scholar] [CrossRef]
- Van Meer, G.; de Kroon, A.I.P.M. Lipid Map of the Mammalian Cell. J. Cell Sci. 2011, 124, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.D. Lipidomics: A Global Approach to Lipid Analysis in Biological Systems. J. Lipid Res. 2006, 47, 2101–2111. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.W.; Braidy, N.; Poljak, A.; Pickford, R.; Thambisetty, M.; Sachdev, P.S. Dysregulation of Lipids in Alzheimer’s Disease and Their Role as Potential Biomarkers. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2017, 13, 810–827. [Google Scholar] [CrossRef]
- Perrotti, F.; Rosa, C.; Cicalini, I.; Sacchetta, P.; Del Boccio, P.; Genovesi, D.; Pieragostino, D. Advances in Lipidomics for Cancer Biomarkers Discovery. Int. J. Mol. Sci. 2016, 17, 1992. [Google Scholar] [CrossRef] [Green Version]
- Mika, A.; Sledzinski , T.; Stepnowski, P. Current Progress of Lipid Analysis in Metabolic Diseases by Mass Spectrometry Methods. Curr. Med. Chem. 2019, 60–103. [Google Scholar] [CrossRef]
- Tenenboim, H.; Burgos, A.; Willmitzer, L.; Brotman, Y. Using Lipidomics for Expanding the Knowledge on Lipid Metabolism in Plants. Biochimie 2016, 130, 91–96. [Google Scholar] [CrossRef]
- Appala, K.; Bimpeh, K.; Freeman, C.; Hines, K.M. Recent Applications of Mass Spectrometry in Bacterial Lipidomics. Anal. Bioanal. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Hewelt-Belka, W.; Nakonieczna, J.; Belka, M.; Bączek, T.; Namieśnik, J.; Kot-Wasik, A. Untargeted Lipidomics Reveals Differences in the Lipid Pattern among Clinical Isolates of Staphylococcus Aureus Resistant and Sensitive to Antibiotics. J. Proteome Res. 2016, 15, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, C.; Han, X. Tutorial on Lipidomics. Anal. Chim. Acta 2019, 1061, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Cajka, T.; Fiehn, O. Comprehensive Analysis of Lipids in Biological Systems by Liquid Chromatography-Mass Spectrometry. TrAC Trends Anal. Chem. 2014, 61, 192–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cajka, T.; Fiehn, O. Toward Merging Untargeted and Targeted Methods in Mass Spectrometry-Based Metabolomics and Lipidomics. Anal. Chem. 2016, 88, 524–545. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid Extraction by Methyl-Tert-Butyl Ether for High-Throughput Lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Löfgren, L.; Ståhlman, M.; Forsberg, G.-B.; Saarinen, S.; Nilsson, R.; Hansson, G.I. The BUME Method: A Novel Automated Chloroform-Free 96-Well Total Lipid Extraction Method for Blood Plasma. J. Lipid Res. 2012, 53, 1690–1700. [Google Scholar] [CrossRef] [Green Version]
- Sostare, J.; Di Guida, R.; Kirwan, J.; Chalal, K.; Palmer, E.; Dunn, W.B.; Viant, M.R. Comparison of Modified Matyash Method to Conventional Solvent Systems for Polar Metabolite and Lipid Extractions. Anal. Chim. Acta 2018, 1037, 301–315. [Google Scholar] [CrossRef]
- Ulmer, C.Z.; Jones, C.M.; Yost, R.A.; Garrett, T.J.; Bowden, J.A. Optimization of Folch, Bligh-Dyer, and Matyash Sample-to-Extraction Solvent Ratios for Human Plasma-Based Lipidomics Studies. Anal. Chim. Acta 2018, 1037, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Satomi, Y.; Hirayama, M.; Kobayashi, H. One-Step Lipid Extraction for Plasma Lipidomics Analysis by Liquid Chromatography Mass Spectrometry. J. Chromatogr. B 2017, 1063, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Sarafian, M.H.; Gaudin, M.; Lewis, M.R.; Martin, F.-P.; Holmes, E.; Nicholson, J.K.; Dumas, M.-E. Objective Set of Criteria for Optimization of Sample Preparation Procedures for Ultra-High Throughput Untargeted Blood Plasma Lipid Profiling by Ultra Performance Liquid Chromatography–Mass Spectrometry. Anal. Chem. 2014, 86, 5766–5774. [Google Scholar] [CrossRef] [PubMed]
- Zein Elabdeen, H.R.; Mustafa, M.; Szklenar, M.; Rühl, R.; Ali, R.; Bolstad, A.I. Ratio of Pro-Resolving and Pro-Inflammatory Lipid Mediator Precursors as Potential Markers for Aggressive Periodontitis. PLoS ONE 2013, 8, e70838. [Google Scholar] [CrossRef] [Green Version]
- Jurowski, K.; Kochan, K.; Walczak, J.; Barańska, M.; Piekoszewski, W.; Buszewski, B. Comprehensive Review of Trends and Analytical Strategies Applied for Biological Samples Preparation and Storage in Modern Medical Lipidomics: State of the Art. TrAC Trends Anal. Chem. 2017, 86, 276–289. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, L.; Steffen, D.; Ye, T.; Raftery, D. Metabolic Profiling of Gender: Headspace-SPME/GC–MS and 1H NMR Analysis of Urine. Metabolomics 2012, 8, 323–334. [Google Scholar] [CrossRef]
- Silva, C.L.; Passos, M.; Câmara, J.S. Solid Phase Microextraction, Mass Spectrometry and Metabolomic Approaches for Detection of Potential Urinary Cancer Biomarkers—A Powerful Strategy for Breast Cancer Diagnosis. Talanta 2012, 89, 360–368. [Google Scholar] [CrossRef]
- Pawliszyn, J. Theory of Solid-Phase Microextraction. J. Chromatogr. Sci. 2000, 38, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, C.; Han, R.H.; Han, X. Novel Advances in Shotgun Lipidomics for Biology and Medicine. Prog. Lipid Res. 2016, 61, 83–108. [Google Scholar] [CrossRef] [Green Version]
- Reichardt, C.; Welton, T.; Appendix, A. Properties, Purification, and Use of Organic Solvents. In Solvents and Solvent Effects in Organic Chemistry; Wiley: Hoboken, NJ, USA, 2010; pp. 549–586. [Google Scholar] [CrossRef]
- Seyer, A.; Boudah, S.; Broudin, S.; Junot, C.; Colsch, B. Annotation of the Human Cerebrospinal Fluid Lipidome Using High Resolution Mass Spectrometry and a Dedicated Data Processing Workflow. Metabolomics 2016, 12, 91. [Google Scholar] [CrossRef] [Green Version]
- Burla, B.; Arita, M.; Arita, M.; Bendt, A.K.; Cazenave-Gassiot, A.; Dennis, E.A.; Ekroos, K.; Han, X.; Ikeda, K.; Liebisch, G.; et al. MS-Based Lipidomics of Human Blood Plasma—A Community-Initiated Position Paper to Develop Accepted Guidelines. J. Lipid Res. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Meulebroek, L.; De Paepe, E.; Vercruysse, V.; Pomian, B.; Bos, S.; Lapauw, B.; Vanhaecke, L. Holistic Lipidomics of the Human Gut Phenotype Using Validated Ultra-High-Performance Liquid Chromatography Coupled to Hybrid Orbitrap Mass Spectrometry. Anal. Chem. 2017, 89, 12502–12510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, M.W.K.; Braidy, N.; Pickford, R.; Sachdev, P.S.; Poljak, A. Comparison of Single Phase and Biphasic Extraction Protocols for Lipidomic Studies Using Human Plasma. Front. Neurol. 2019, 10, 879. [Google Scholar] [CrossRef]
- Gardner, M.S.; Kuklenyik, Z.; Lehtikoski, A.; Carter, K.A.; McWilliams, L.G.; Kusovschi, J.; Bierbaum, K.; Jones, J.I.; Rees, J.; Reis, G.; et al. Development and Application of a High Throughput One-Pot Extraction Protocol for Quantitative LC-MS/MS Analysis of Phospholipids in Serum and Lipoprotein Fractions in Normolipidemic and Dyslipidemic Subjects. J. Chromatogr. B 2019, 1118–1119, 137–147. [Google Scholar] [CrossRef]
- Shibusawa, Y.; Yamakawa, Y.; Noji, R.; Yanagida, A.; Shindo, H.; Ito, Y. Three-Phase Solvent Systems for Comprehensive Separation of a Wide Variety of Compounds by High-Speed Counter-Current Chromatography. J. Chromatogr. A 2006, 1133, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Leopold, J.; Popkova, Y.; Engel, K.M.; Schiller, J. Recent Developments of Useful MALDI Matrices for the Mass Spectrometric Characterization of Lipids. Biomolecules 2018, 8, 173. [Google Scholar] [CrossRef] [Green Version]
- Alshehry, H.Z.; Barlow, K.C.; Weir, M.J.; Zhou, Y.; McConville, J.M.; Meikle, J.P. An Efficient Single Phase Method for the Extraction of Plasma Lipids. Metabolites 2015, 5, 389. [Google Scholar] [CrossRef]
- Alshehry, Z.H.; Mundra, P.A.; Barlow, C.K.; Mellett, N.A.; Wong, G.; McConville, M.J.; Simes, J.; Tonkin, A.M.; Sullivan, D.R.; Barnes, E.H.; et al. Plasma Lipidomic Profiles Improve on Traditional Risk Factors for the Prediction of Cardiovascular Events in Type 2 Diabetes Mellitus. Circulation 2016, 134, 1637–1650. [Google Scholar] [CrossRef] [Green Version]
- Meikle, P.J.; Gerard, W.; Despina, T.; Barlow, C.K.; Weir, J.M.; Christopher, M.J.; MacIntosh, G.L.; Benjamin, G.; Linda, S.; Adam, K.; et al. Plasma Lipidomic Analysis of Stable and Unstable Coronary Artery Disease. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2723–2732. [Google Scholar] [CrossRef] [Green Version]
- Iriondo, A.; Tainta, M.; Saldias, J.; Arriba, M.; Ochoa, B.; Goñi, F.M.; Martinez-Lage, P.; Abad-García, B. Isopropanol Extraction for Cerebrospinal Fluid Lipidomic Profiling Analysis. Talanta 2019, 195, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Rombouts, C.; De Spiegeleer, M.; Van Meulebroek, L.; De Vos, W.H.; Vanhaecke, L. Validated Comprehensive Metabolomics and Lipidomics Analysis of Colon Tissue and Cell Lines. Anal. Chim. Acta 2019, 1066, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Birjandi, A.P.; Bojko, B.; Ning, Z.; Figeys, D.; Pawliszyn, J. High Throughput Solid Phase Microextraction: A New Alternative for Analysis of Cellular Lipidome? J. Chromatogr. B 2017, 1043, 12–19. [Google Scholar] [CrossRef]
- Astarita, G.; Ahmed, F.; Piomelli, D. Lipidomic Analysis of Biological Samples by Liquid Chromatography Coupled to Mass Spectrometry BT—Lipidomics: Volume 1: Methods and Protocols; Armstrong, D., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 201–219. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Guo, H.; Wu, T.; Tao, N.; Wang, X.; Zhong, J. Effect of Three Types of Thermal Processing Methods on the Lipidomics Profile of Tilapia Fillets by UPLC-Q-Extractive Orbitrap Mass Spectrometry. Food Chem. 2019, 298, 125029. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Xiang, X.; Jin, Y.; Wang, N.; Ma, M. High-Throughput Lipidomic Profiling of High-Density Lipoprotein from Egg Yolk (EYHDL): Comparison Based on UPLC-MS/MS and GC–MS. Eur. Food Res. Technol. 2019, 245, 1665–1675. [Google Scholar] [CrossRef]
- Chen, J.; Nichols, K.K.; Wilson, L.; Barnes, S.; Nichols, J.J. Untargeted Lipidomic Analysis of Human Tears: A New Approach for Quantification of O-Acyl-Omega Hydroxy Fatty Acids. Ocul. Surf. 2019, 17, 347–355. [Google Scholar] [CrossRef]
- Martins-Noguerol, R.; Moreno-Pérez, J.A.; Acket, S.; Makni, S.; Garcés, R.; Troncoso-Ponce, A.; Salas, J.J.; Thomasset, B.; Martínez-Force, E. Lipidomic Analysis of Plastidial Octanoyltransferase Mutants of Arabidopsis Thaliana. Metabolites 2019, 9, 209. [Google Scholar] [CrossRef] [Green Version]
- Min, H.K.; Lim, S.; Chung, B.C.; Moon, M.H. Shotgun Lipidomics for Candidate Biomarkers of Urinary Phospholipids in Prostate Cancer. Anal. Bioanal. Chem. 2011, 399, 823–830. [Google Scholar] [CrossRef]
- Gregory, K.E.; Bird, S.S.; Gross, V.S.; Marur, V.R.; Lazarev, A.V.; Walker, W.A.; Kristal, B.S. Method Development for Fecal Lipidomics Profiling. Anal. Chem. 2013, 85, 1114–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Löfgren, L.; Forsberg, G.-B.; Ståhlman, M. The BUME Method: A New Rapid and Simple Chloroform-Free Method for Total Lipid Extraction of Animal Tissue. Sci. Rep. 2016, 6, 27688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrizo, D.; Nerín, I.; Domeño, C.; Alfaro, P.; Nerín, C. Direct Screening of Tobacco Indicators in Urine and Saliva by Atmospheric Pressure Solid Analysis Probe Coupled to Quadrupole-Time of Flight Mass Spectrometry (ASAP-MS-Q-TOF-). J. Pharm. Biomed. Anal. 2016, 124, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Burgos, A.; Szymanski, J.; Seiwert, B.; Degenkolbe, T.; Hannah, M.A.; Giavalisco, P.; Willmitzer, L. Analysis of Short-Term Changes in the Arabidopsis Thaliana Glycerolipidome in Response to Temperature and Light. Plant J. 2011, 66, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Kamide, Y.; Hirai, M.Y.; Saito, K. Plant Lipidomics Based on Hydrophilic Interaction Chromatography Coupled to Ion Trap Time-of-Flight Mass Spectrometry. Metabolomics 2013, 9, 121–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cala, M.P.; Aldana, J.; Medina, J.; Sánchez, J.; Guio, J.; Wist, J.; Meesters, R.J.W. Multiplatform Plasma Metabolic and Lipid Fingerprinting of Breast Cancer: A Pilot Control-Case Study in Colombian Hispanic Women. PLoS ONE 2018, 13, e0190958. [Google Scholar] [CrossRef] [Green Version]
- Cala, M.; Aldana, J.; Sánchez, J.; Guio, J.; Meesters, R.J.W. Urinary Metabolite and Lipid Alterations in Colombian Hispanic Women with Breast Cancer: A Pilot Study. J. Pharm. Biomed. Anal. 2018, 152, 234–241. [Google Scholar] [CrossRef]
- Navarro-Reig, M.; Tauler, R.; Iriondo-Frias, G.; Jaumot, J. Untargeted Lipidomic Evaluation of Hydric and Heat Stresses on Rice Growth. J. Chromatogr. B 2019, 1104, 148–156. [Google Scholar] [CrossRef]
- Villaseñor, A.; Garcia-Perez, I.; Garcia, A.; Posma, J.M.; Fernández-López, M.; Nicholas, A.J.; Modi, N.; Holmes, E.; Barbas, C. Breast Milk Metabolome Characterization in a Single-Phase Extraction, Multiplatform Analytical Approach. Anal. Chem. 2014, 86, 8245–8252. [Google Scholar] [CrossRef] [PubMed]
- Hummel, J.; Segu, S.; Li, Y.; Irgang, S.; Jueppner, J.; Giavalisco, P. Ultra Performance Liquid Chromatography and High Resolution Mass Spectrometry for the Analysis of Plant Lipids. Front. Plant Sci. 2011, 2, 54. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.S.; Raju, R.; Ray, S.; Kshirsagar, R.; Gilbert, A.; Zang, L.; Karger, B.L. Lipidomics of CHO Cell Bioprocessing: Relation to Cell Growth and Specific Productivity of a Monoclonal Antibody. Biotechnol. J. 2018, 13, 1700745. [Google Scholar] [CrossRef]
- Zhang, H.; Gao, Y.; Sun, J.; Fan, S.; Yao, X.; Ran, X.; Zheng, C.; Huang, M.; Bi, H. Optimization of Lipid Extraction and Analytical Protocols for UHPLC-ESI-HRMS-Based Lipidomic Analysis of Adherent Mammalian Cancer Cells. Anal. Bioanal. Chem. 2017, 409, 5349–5358. [Google Scholar] [CrossRef]
- Calderón, C.; Sanwald, C.; Schlotterbeck, J.; Drotleff, B.; Lämmerhofer, M. Comparison of Simple Monophasic versus Classical Biphasic Extraction Protocols for Comprehensive UHPLC-MS/MS Lipidomic Analysis of Hela Cells. Anal. Chim. Acta 2019, 1048, 66–74. [Google Scholar] [CrossRef]
- Nilsson, A.K.; Sjöbom, U.; Christenson, K.; Hellström, A. Lipid Profiling of Suction Blister Fluid: Comparison of Lipids in Interstitial Fluid and Plasma. Lipids Health Dis. 2019, 18, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwood, D.J.; Dos Santos, M.S.; Huang, S.; Russell, M.R.G.; Collinson, L.M.; MacRae, J.I.; West, A.; Jiang, H.; Gutierrez, M.G. Subcellular Antibiotic Visualization Reveals a Dynamic Drug Reservoir in Infected Macrophages. Science 2019, 364, 1279–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, S.; Barnes, J.M.; Young, S.A.; Garcia, D.M.; Tolley, H.D.; Kauwe, J.S.K.; Graves, S.W. Discovery and Confirmation of Diagnostic Serum Lipid Biomarkers for Alzheimer’s Disease Using Direct Infusion Mass Spectrometry. J. Alzheimer’s Dis. 2017, 59, 277–290. [Google Scholar]
- Gulati, S.; Ekland, E.H.; Ruggles, K.V.; Chan, R.B.; Jayabalasingham, B.; Zhou, B.; Mantel, P.-Y.; Lee, M.C.S.; Spottiswoode, N.; Coburn-Flynn, O.; et al. Profiling the Essential Nature of Lipid Metabolism in Asexual Blood and Gametocyte Stages of Plasmodium Falciparum. Cell Host Microbe 2015, 18, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Uchikata, T.; Matsubara, A.; Fukusaki, E.; Bamba, T. High-Throughput Phospholipid Profiling System Based on Supercritical Fluid Extraction–Supercritical Fluid Chromatography/Mass Spectrometry for Dried Plasma Spot Analysis. J. Chromatogr. A 2012, 1250, 69–75. [Google Scholar] [CrossRef]
- Lísa, M.; Holčapek, M. UHPSFC/ESI-MS Analysis of Lipids BT—Clinical Metabolomics: Methods and Protocols; Giera, M., Ed.; Springer: New York, NY, USA, 2018; pp. 73–82. [Google Scholar] [CrossRef]
- Yang, Y.; Zhong, Q.; Zhang, H.; Mo, C.; Yao, J.; Huang, T.; Zhou, T.; Tan, W. Lipidomics Study of the Protective Effects of Isosteviol Sodium on Stroke Rats Using Ultra High-Performance Supercritical Fluid Chromatography Coupling with Ion-Trap and Time-of-Flight Tandem Mass Spectrometry. J. Pharm. Biomed. Anal. 2018, 157, 145–155. [Google Scholar] [CrossRef]
- Bojko, B.; Wąsowicz, M.; Pawliszyn, J. Metabolic Profiling of Plasma from Cardiac Surgical Patients Concurrently Administered with Tranexamic Acid: DI-SPME–LC–MS Analysis. J. Pharm. Anal. 2014, 4, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Bessonneau, V.; Bojko, B.; Pawliszyn, J. Analysis of Human Saliva Metabolome by Direct Immersion Solid-Phase Microextraction LC and Benchtop Orbitrap MS. Bioanalysis 2013, 5, 783–792. [Google Scholar] [CrossRef]
- Garwolińska, D.; Hewelt-Belka, W.; Namieśnik, J.; Kot-Wasik, A. Rapid Characterization of the Human Breast Milk Lipidome Using a Solid-Phase Microextraction and Liquid Chromatography–Mass Spectrometry-Based Approach. J. Proteome Res. 2017, 16, 3200–3208. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, L.; Shi, X.; Xu, G. New Advances in Analytical Methods for Mass Spectrometry-Based Large-Scale Metabolomics Study. TrAC Trends Anal. Chem. 2019, 121, 115665. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, Y.; Yang, J.; Ye, F.; Zhou, T.; Gongke, L. Advances of Supercritical Fluid Chromatography in Lipid Profiling. J. Pharm. Anal. 2019, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Furse, S.; Egmond, M.R.; Killian, J.A. Isolation of Lipids from Biological Samples. Mol. Membr. Biol. 2015, 32, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Sethi, S.; Brietzke, E. Recent Advances in Lipidomics: Analytical and Clinical Perspectives. Prostaglandins Other Lipid Mediat. 2017, 128–129, 8–16. [Google Scholar] [CrossRef] [PubMed]
- König, S.; Ischebeck, T.; Lerche, J.; Stenzel, I.; Heilmann, I. Salt-Stress-Induced Association of Phosphatidylinositol 4,5-Bisphosphate with Clathrin-Coated Vesicles in Plants. Biochem. J. 2008, 415, 387–399. [Google Scholar] [CrossRef] [Green Version]
- Lubary, M.; Hofland, G.W.; ter Horst, J.H. A Process for the Production of a Diacylglycerol-Based Milk Fat Analogue. Eur. J. Lipid Sci. Technol. 2011, 113, 459–468. [Google Scholar] [CrossRef]
- Schlotterbeck, J.; Cebo, M.; Kolb, A.; Lämmerhofer, M. Quantitative Analysis of Chemoresistance-Inducing Fatty Acid in Food Supplements Using UHPLC–ESI-MS/MS. Anal. Bioanal. Chem. 2019, 411, 479–491. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, H.; Zhu, M.-J. A Sensitive GC/MS Detection Method for Analyzing Microbial Metabolites Short Chain Fatty Acids in Fecal and Serum Samples. Talanta 2019, 196, 249–254. [Google Scholar] [CrossRef]
- Pérez-Navarro, J.; Da Ros, A.; Masuero, D.; Izquierdo-Cañas, P.M.; Hermosín-Gutiérrez, I.; Gómez-Alonso, S.; Mattivi, F.; Vrhovsek, U. LC-MS/MS Analysis of Free Fatty Acid Composition and Other Lipids in Skins and Seeds of Vitis Vinifera Grape Cultivars. Food Res. Int. 2019, 125, 108556. [Google Scholar] [CrossRef]
- Mehari, B.; Redi-Abshiro, M.; Chandravanshi, B.S.; Combrinck, S.; McCrindle, R.; Atlabachew, M. GC-MS Profiling of Fatty Acids in Green Coffee (Coffea arabica L.) Beans and Chemometric Modeling for Tracing Geographical Origins from Ethiopia. J. Sci. Food Agric. 2019, 99, 3811–3823. [Google Scholar] [CrossRef]
- Zarai, Z.; Eddehech, A.; Rigano, F.; Oteri, M.; Micalizzi, G.; Dugo, P.; Mondello, L.; Cacciola, F. Characterization of Monoacylglycerols and Diacylglycerols Rich in Polyunsaturated Fatty Acids Produced by Hydrolysis of Musteleus Mustelus Liver Oil Catalyzed by an Immobilized Bacterial Lipase. J. Chromatogr. A 2020, 1613, 460692. [Google Scholar] [CrossRef]
- Gašparović, B.; Penezić, A.; Lampitt, R.S.; Sudasinghe, N.; Schaub, T. Free Fatty Acids, Tri-, Di- and Monoacylglycerol Production and Depth-Related Cycling in the Northeast Atlantic. Mar. Chem. 2016, 186, 101–109. [Google Scholar] [CrossRef]
- Jensen, J.R.; Pitcher, M.H.; Yuan, Z.X.; Ramsden, C.E.; Domenichiello, A.F. Concentrations of Oxidized Linoleic Acid Derived Lipid Mediators in the Amygdala and Periaqueductal Grey Are Reduced in a Mouse Model of Chronic Inflammatory Pain. Prostaglandins, Leukot. Essent. Fat. Acids 2018, 135, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Rund, K.M.; Ostermann, A.I.; Kutzner, L.; Galano, J.-M.; Oger, C.; Vigor, C.; Wecklein, S.; Seiwert, N.; Durand, T.; Schebb, N.H. Development of an LC-ESI(-)-MS/MS Method for the Simultaneous Quantification of 35 Isoprostanes and Isofurans Derived from the Major N3- and N6-PUFAs. Anal. Chim. Acta 2018, 1037, 63–74. [Google Scholar] [CrossRef]
- Sanak, M.; Gielicz, A.; Nagraba, K.; Kaszuba, M.; Kumik, J.; Szczeklik, A. Targeted Eicosanoids Lipidomics of Exhaled Breath Condensate in Healthy Subjects. J. Chromatogr. B 2010, 878, 1796–1800. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.M.; Poloyac, S.M.; Anderson, K.B.; Waddell, B.L.; Messamore, E.; Yao, J.K. A Rapid UPLC-MS/MS Assay for Eicosanoids in Human Plasma: Application to Evaluate Niacin Responsivity. Prostaglandins Leukot. Essent. Fat. Acids 2018, 136, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Cummings, M.; Massey, K.A.; Mappa, G.; Wilkinson, N.; Hutson, R.; Munot, S.; Saidi, S.; Nugent, D.; Broadhead, T.; Wright, A.I.; et al. Integrated Eicosanoid Lipidomics and Gene Expression Reveal Decreased Prostaglandin Catabolism and Increased 5-Lipoxygenase Expression in Aggressive Subtypes of Endometrial Cancer. J. Pathol. 2019, 247, 21–34. [Google Scholar] [CrossRef]
- Gobo, L.A.; de Carvalho, L.M.; Temp, F.; Viana, C.; Mello, C.F. A Rapid Method for Identification and Quantification of Prostaglandins in Cerebral Tissues by UHPLC-ESI-MS/MS for the Lipidomic in Vivo Studies. Anal. Biochem. 2018, 545, 98–103. [Google Scholar] [CrossRef]
- Dumlao, D.S.; Buczynski, M.W.; Norris, P.C.; Harkewicz, R.; Dennis, E.A. High-Throughput Lipidomic Analysis of Fatty Acid Derived Eicosanoids and N-Acylethanolamines. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 724–736. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.M.; Donnelly, M.K.; Crago, E.A.; Roman, D.M.; Sherwood, P.R.; Horowitz, M.B.; Poloyac, S.M. Rapid, Simultaneous Quantitation of Mono and Dioxygenated Metabolites of Arachidonic Acid in Human CSF and Rat Brain. J. Chromatogr. B 2009, 877, 3991–4000. [Google Scholar] [CrossRef] [Green Version]
- Teav, T.; Gallart-Ayala, H.; van der Velpen, V.; Mehl, F.; Henry, H.; Ivanisevic, J. Merged Targeted Quantification and Untargeted Profiling for Comprehensive Assessment of Acylcarnitine and Amino Acid Metabolism. Anal. Chem. 2019, 91, 11757–11769. [Google Scholar] [CrossRef]
- Wang, M.; Yang, R.; Mu, H.; Zeng, J.; Zhang, T.; Zhou, W.; Wang, S.; Tang, Y.; Li, H.; Zhang, C.; et al. A Simple and Precise Method for Measurement of Serum Free Carnitine and Acylcarnitines by Isotope Dilution HILIC-ESI-MS/MS. Int. J. Mass Spectrom. 2019, 446, 116208. [Google Scholar] [CrossRef]
- Yang, Y.; Sadri, H.; Prehn, C.; Adamski, J.; Rehage, J.; Dänicke, S.; Saremi, B.; Sauerwein, H. Acylcarnitine Profiles in Serum and Muscle of Dairy Cows Receiving Conjugated Linoleic Acids or a Control Fat Supplement during Early Lactation. J. Dairy Sci. 2019, 102, 754–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.T.; MahmoudianDehkordi, S.; Bhattacharyya, S.; Arnold, M.; Liu, D.; Neavin, D.; Moseley, M.A.; Thompson, J.W.; Williams, L.S.J.; Louie, G.; et al. Acylcarnitine Metabolomic Profiles Inform Clinically-Defined Major Depressive Phenotypes. J. Affect. Disord. 2020, 264, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhang, Q. Simultaneous Quantification of Free Fatty Acids and Acylcarnitines in Plasma Samples Using Dansylhydrazine Labeling and Liquid Chromatography–Triple Quadrupole Mass Spectrometry. Anal. Bioanal. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kolar, M.J.; Nelson, A.T.; Chang, T.; Ertunc, M.E.; Christy, M.P.; Ohlsson, L.; Härröd, M.; Kahn, B.B.; Siegel, D.; Saghatelian, A. Faster Protocol for Endogenous Fatty Acid Esters of Hydroxy Fatty Acid (FAHFA) Measurements. Anal. Chem. 2018, 90, 5358–5365. [Google Scholar] [CrossRef]
- Zhu, Q.-F.; Yan, J.-W.; Zhang, T.-Y.; Xiao, H.-M.; Feng, Y.-Q. Comprehensive Screening and Identification of Fatty Acid Esters of Hydroxy Fatty Acids in Plant Tissues by Chemical Isotope Labeling-Assisted Liquid Chromatography–Mass Spectrometry. Anal. Chem. 2018, 90, 10056–10063. [Google Scholar] [CrossRef]
- Zhu, Q.-F.; Yan, J.-W.; Ni, J.; Feng, Y.-Q. FAHFA Footprint in the Visceral Fat of Mice across Their Lifespan. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158639. [Google Scholar] [CrossRef]
- Hu, C.; Wang, M.; Duan, Q.; Han, X. Sensitive Analysis of Fatty Acid Esters of Hydroxy Fatty Acids in Biological Lipid Extracts by Shotgun Lipidomics after One-Step Derivatization. Anal. Chim. Acta 2020, 1105, 105–111. [Google Scholar] [CrossRef]
- Tan, B.; William Yu, Y.; Francesca Monn, M.; Velocity Hughes, H.; O’Dell, D.K.; Michael Walker, J. Targeted Lipidomics Approach for Endogenous N-Acyl Amino Acids in Rat Brain Tissue. J. Chromatogr. B 2009, 877, 2890–2894. [Google Scholar] [CrossRef]
- Zhu, M.; Xu, X.; Hou, Y.; Han, J.; Wang, J.; Zheng, Q.; Hao, H. Boronic Derivatization of Monoacylglycerol and Monitoring in Biofluids. Anal. Chem. 2019, 91, 6724–6729. [Google Scholar] [CrossRef]
- Gao, F.; McDaniel, J.; Chen, Y.E.; Rockwell, H.; Lynes, D.M.; Tseng, Y.-H.; Sarangarajan, R.; Narain, R.N.; Kiebish, A.M. Monoacylglycerol Analysis Using MS/MSALL Quadruple Time of Flight Mass Spectrometry. Metabolites 2016, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Ertl, V.M.; Höring, M.; Schött, H.-F.; Blücher, C.; Kjølbæk, L.; Astrup, A.; Burkhardt, R.; Liebisch, G. Quantification of Diacylglycerol and Triacylglycerol Species in Human Fecal Samples by Flow Injection Fourier Transform Mass Spectrometry. Anal. Bioanal. Chem. 2020. [Google Scholar] [CrossRef] [Green Version]
- Rashid, R.; Cazenave-Gassiot, A.; Gao, I.H.; Nair, Z.J.; Kumar, J.K.; Gao, L.; Kline, K.A.; Wenk, M.R. Comprehensive Analysis of Phospholipids and Glycolipids in the Opportunistic Pathogen Enterococcus Faecalis. PLoS ONE 2017, 12, e0175886. [Google Scholar] [CrossRef] [Green Version]
- Harrison, K.A.; Bergman, B.C. HPLC-MS/MS Methods for Diacylglycerol and Sphingolipid Molecular Species in Skeletal Muscle BT—High-Throughput Metabolomics: Methods and Protocols; D’Alessandro, A., Ed.; Springer: New York, NY, USA, 2019; pp. 137–152. [Google Scholar] [CrossRef]
- Bird, S.S.; Marur, V.R.; Sniatynski, M.J.; Greenberg, H.K.; Kristal, B.S. Serum Lipidomics Profiling Using LC–MS and High-Energy Collisional Dissociation Fragmentation: Focus on Triglyceride Detection and Characterization. Anal. Chem. 2011, 83, 6648–6657. [Google Scholar] [CrossRef] [Green Version]
- Zeb, A. Triacylglycerols Composition, Oxidation and Oxidation Compounds in Camellia Oil Using Liquid Chromatography–Mass Spectrometry. Chem. Phys. Lipids 2012, 165, 608–614. [Google Scholar] [CrossRef]
- Zheng, G.; Li, W. Profiling Membrane Glycerolipids during γ-Ray-Induced Membrane Injury. BMC Plant Biol. 2017, 17, 203. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Su, X.; Luo, Q.; Xu, J.; Chen, J.; Yan, X.; Chen, H. Profiles of Glycerolipids in Pyropia Haitanensis and Their Changes Responding to Agaro-Oligosaccharides. J. Appl. Phycol. 2014, 26, 2397–2404. [Google Scholar] [CrossRef]
- Knific, T.; Vouk, K.; Smrkolj, Š.; Prehn, C.; Adamski, J.; Rižner, T.L. Models Including Plasma Levels of Sphingomyelins and Phosphatidylcholines as Diagnostic and Prognostic Biomarkers of Endometrial Cancer. J. Steroid Biochem. Mol. Biol. 2018, 178, 312–321. [Google Scholar] [CrossRef]
- Yan, Y.; Du, Z.; Chen, C.; Li, J.; Xiong, X.; Zhang, Y.; Jiang, H. Lysophospholipid Profiles of Apolipoprotein E-Deficient Mice Reveal Potential Lipid Biomarkers Associated with Atherosclerosis Progression Using Validated UPLC-QTRAP-MS/MS-Based Lipidomics Approach. J. Pharm. Biomed. Anal. 2019, 171, 148–157. [Google Scholar] [CrossRef]
- Uhl, O.; Glaser, C.; Demmelmair, H.; Koletzko, B. Reversed Phase LC/MS/MS Method for Targeted Quantification of Glycerophospholipid Molecular Species in Plasma. J. Chromatogr. B 2011, 879, 3556–3564. [Google Scholar] [CrossRef]
- Wang, X.; Xu, Y.; Song, X.; Jia, Q.; Zhang, X.; Qian, Y.; Qiu, J. Analysis of Glycerophospholipid Metabolism after Exposure to PCB153 in PC12 Cells through Targeted Lipidomics by UHPLC-MS/MS. Ecotoxicol. Environ. Saf. 2019, 169, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Deeley, J.M.; Thomas, M.C.; Truscott, R.J.W.; Mitchell, T.W.; Blanksby, S.J. Identification of Abundant Alkyl Ether Glycerophospholipids in the Human Lens by Tandem Mass Spectrometry Techniques. Anal. Chem. 2009, 81, 1920–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- She, Y.; Song, J.; Yang, E.; Zhao, L.; Zhong, Y.; Rui, W.; Feng, Y.; Wu, X. Rapid Identification of Glycerophospholipids from RAW264.7 Cells by UPLC/ESI -QTOF-MS. Biomed. Chromatogr. 2014, 28, 1744–1755. [Google Scholar] [CrossRef]
- She, Y.; Zheng, Q.; Xiao, X.; Wu, X.; Feng, Y. An Analysis on the Suppression of NO and PGE2 by Diphenylheptane A and Its Effect on Glycerophospholipids of Lipopolysaccharide-Induced RAW264.7 Cells with UPLC/ESI-QTOF-MS. Anal. Bioanal. Chem. 2016, 408, 3185–3201. [Google Scholar] [CrossRef]
- Sun, T.; Pawlowski, S.; Johnson, M.E. Highly Efficient Microscale Purification of Glycerophospholipids by Microfluidic Cell Lysis and Lipid Extraction for Lipidomics Profiling. Anal. Chem. 2011, 83, 6628–6634. [Google Scholar] [CrossRef] [Green Version]
- Maciel, E.; Neves, B.M.; Santinha, D.; Reis, A.; Domingues, P.; Teresa Cruz, M.; Pitt, A.R.; Spickett, C.M.; Domingues, M.R.M. Detection of Phosphatidylserine with a Modified Polar Head Group in Human Keratinocytes Exposed to the Radical Generator AAPH. Arch. Biochem. Biophys. 2014, 548, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Stübiger, G.; Aldover-Macasaet, E.; Bicker, W.; Sobal, G.; Willfort-Ehringer, A.; Pock, K.; Bochkov, V.; Widhalm, K.; Belgacem, O. Targeted Profiling of Atherogenic Phospholipids in Human Plasma and Lipoproteins of Hyperlipidemic Patients Using MALDI-QIT-TOF-MS/MS. Atherosclerosis 2012, 224, 177–186. [Google Scholar] [CrossRef]
- Basit, A.; Piomelli, D.; Armirotti, A. Rapid Evaluation of 25 Key Sphingolipids and Phosphosphingolipids in Human Plasma by LC-MS/MS. Anal. Bioanal. Chem. 2015, 407, 5189–5198. [Google Scholar] [CrossRef] [Green Version]
- Mi, S.; Zhao, Y.-Y.; Dielschneider, R.F.; Gibson, S.B.; Curtis, J.M. An LC/MS/MS Method for the Simultaneous Determination of Individual Sphingolipid Species in B Cells. J. Chromatogr. B 2016, 1031, 50–60. [Google Scholar] [CrossRef]
- Chipeaux, C.; de Person, M.; Burguet, N.; Billette de Villemeur, T.; Rose, C.; Belmatoug, N.; Héron, S.; Le Van Kim, C.; Franco, M.; Moussa, F. Optimization of Ultra-High Pressure Liquid Chromatography–Tandem Mass Spectrometry Determination in Plasma and Red Blood Cells of Four Sphingolipids and Their Evaluation as Biomarker Candidates of Gaucher’s Disease. J. Chromatogr. A 2017, 1525, 116–125. [Google Scholar] [CrossRef]
- Li, J.; Hu, C.; Zhao, X.; Dai, W.; Chen, S.; Lu, X.; Xu, G. Large-Scaled Human Serum Sphingolipid Profiling by Using Reversed-Phase Liquid Chromatography Coupled with Dynamic Multiple Reaction Monitoring of Mass Spectrometry: Method Development and Application in Hepatocellular Carcinoma. J. Chromatogr. A 2013, 1320, 103–110. [Google Scholar] [CrossRef]
- Sullards, M.C.; Liu, Y.; Chen, Y.; Merrill, A.H. Analysis of Mammalian Sphingolipids by Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS) and Tissue Imaging Mass Spectrometry (TIMS). Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 838–853. [Google Scholar] [CrossRef] [Green Version]
- Bui, H.H.; Leohr, J.K.; Kuo, M.-S. Analysis of Sphingolipids in Extracted Human Plasma Using Liquid Chromatography Electrospray Ionization Tandem Mass Spectrometry. Anal. Biochem. 2012, 423, 187–194. [Google Scholar] [CrossRef]
- Scherer, M.; Böttcher, A.; Schmitz, G.; Liebisch, G. Sphingolipid Profiling of Human Plasma and FPLC-Separated Lipoprotein Fractions by Hydrophilic Interaction Chromatography Tandem Mass Spectrometry. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 68–75. [Google Scholar] [CrossRef]
- T’Kindt, R.; Jorge, L.; Dumont, E.; Couturon, P.; David, F.; Sandra, P.; Sandra, K. Profiling and Characterizing Skin Ceramides Using Reversed-Phase Liquid Chromatography–Quadrupole Time-of-Flight Mass Spectrometry. Anal. Chem. 2012, 84, 403–411. [Google Scholar] [CrossRef]
- Liao, H.W.; Lin, S.W.; Lin, Y.T.; Lee, C.H.; Kuo, C.H. Identification of Potential Sphingomyelin Markers for the Estimation of Hematocrit in Dried Blood Spots via a Lipidomic Strategy. Anal. Chim. Acta 2018, 1003, 34–41. [Google Scholar] [CrossRef]
- Huang, Q.; Liu, D.; Xin, B.; Cechner, K.; Zhou, X.; Wang, H.; Zhou, A. Quantification of Monosialogangliosides in Human Plasma through Chemical Derivatization for Signal Enhancement in LC–ESI-MS. Anal. Chim. Acta 2016, 929, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hájek, R.; Jirásko, R.; Lísa, M.; Cífková, E.; Holčapek, M. Hydrophilic Interaction Liquid Chromatography–Mass Spectrometry Characterization of Gangliosides in Biological Samples. Anal. Chem. 2017, 89, 12425–12432. [Google Scholar] [CrossRef] [PubMed]
- Broughton, R.; Ruíz-Lopez, N.; Hassall, K.L.; Martínez-Force, E.; Garcés, R.; Salas, J.J.; Beaudoin, F. New Insights in the Composition of Wax and Sterol Esters in Common and Mutant Sunflower Oils Revealed by ESI-MS/MS. Food Chem. 2018, 269, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Reinicke, M.; Schröter, J.; Müller-Klieser, D.; Helmschrodt, C.; Ceglarek, U. Free Oxysterols and Bile Acids Including Conjugates—Simultaneous Quantification in Human Plasma and Cerebrospinal Fluid by Liquid Chromatography-Tandem Mass Spectrometry. Anal. Chim. Acta 2018, 1037, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Piqueras, M.; Theotoka, D.; Sarohia, G.S.; Bhattacharya, S.K. Analyses of Cholesterol and Derivatives in Ocular Tissues Using LC-MS/MS Methods BT—Metabolomics: Methods and Protocols; Bhattacharya, S.K., Ed.; Springer: New York, NY, USA, 2019; pp. 53–59. [Google Scholar] [CrossRef]
- Kunz, S.; Matysik, S. A Comprehensive Method to Determine Sterol Species in Human Faeces by GC-Triple Quadrupole MS. J. Steroid Biochem. Mol. Biol. 2019, 190, 99–103. [Google Scholar] [CrossRef]
- Schött, H.-F.; Krautbauer, S.; Höring, M.; Liebisch, G.; Matysik, S. A Validated, Fast Method for Quantification of Sterols and Gut Microbiome Derived 5α/β-Stanols in Human Feces by Isotope Dilution LC–High-Resolution MS. Anal. Chem. 2018, 90, 8487–8494. [Google Scholar] [CrossRef]
- Ceglarek, U.; Dittrich, J.; Leopold, J.; Helmschrodt, C.; Becker, S.; Staab, H.; Richter, O.; Rohm, S.; Aust, G. Free Cholesterol, Cholesterol Precursor and Plant Sterol Levels in Atherosclerotic Plaques Are Independently Associated with Symptomatic Advanced Carotid Artery Stenosis. Atherosclerosis 2020, 295, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Takeshima, M.; Ogihara, H.M.; Kataoka, H. Sterol Characteristics in Silkworm Brain and Various Tissues Characterized by Precise Sterol Profiling Using LC-MS/MS. Int. J. Mol. Sci. 2019, 20, 4840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matysik, S.; Liebisch, G. Quantification of Steroid Hormones in Human Serum by Liquid Chromatography-High Resolution Tandem Mass Spectrometry. J. Chromatogr. A 2017, 1526, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.-Y.; Lee, H.S.; Kim, J.H.; Lee, J.H.; Choi, M.H. Supported Liquid Extraction Coupled to Gas Chromatography-Selective Mass Spectrometric Scan Modes for Serum Steroid Profiling. Anal. Chim. Acta 2018, 1037, 281–292. [Google Scholar] [CrossRef]
- Drotleff, B.; Hallschmid, M.; Lämmerhofer, M. Quantification of Steroid Hormones in Plasma Using a Surrogate Calibrant Approach and UHPLC-ESI-QTOF-MS/MS with SWATH-Acquisition Combined with Untargeted Profiling. Anal. Chim. Acta 2018, 1022, 70–80. [Google Scholar] [CrossRef]
- Hu, T.; An, Z.; Shi, C.; Li, P.; Liu, L. A Sensitive and Efficient Method for Simultaneous Profiling of Bile Acids and Fatty Acids by UPLC-MS/MS. J. Pharm. Biomed. Anal. 2020, 178, 112815. [Google Scholar] [CrossRef] [PubMed]
- LeníĿek, M.; Vecka, M.; Žížalová, K.; Vítek, L. Comparison of Simple Extraction Procedures in Liquid Chromatography⿿mass Spectrometry Based Determination of Serum 7α-Hydroxy-4-Cholesten-3-One, a Surrogate Marker of Bile Acid Synthesis. J. Chromatogr. B 2016, 1033–1034, 317–320. [Google Scholar] [CrossRef]
- Ulaszewska, M.M.; Mancini, A.; Garcia-Aloy, M.; Del Bubba, M.; Tuohy, K.M.; Vrhovsek, U. Isotopic Dilution Method for Bile Acid Profiling Reveals New Sulfate Glycine-Conjugated Dihydroxy Bile Acids and Glucuronide Bile Acids in Serum. J. Pharm. Biomed. Anal. 2019, 173, 1–17. [Google Scholar] [CrossRef]
- Franco, P.; Porru, E.; Fiori, J.; Gioiello, A.; Cerra, B.; Roda, G.; Caliceti, C.; Simoni, P.; Roda, A. Identification and Quantification of Oxo-Bile Acids in Human Faeces with Liquid Chromatography–Mass Spectrometry: A Potent Tool for Human Gut Acidic Sterolbiome Studies. J. Chromatogr. A 2019, 1585, 70–81. [Google Scholar] [CrossRef]
- Higashi, T.; Watanabe, S.; Tomaru, K.; Yamazaki, W.; Yoshizawa, K.; Ogawa, S.; Nagao, H.; Minato, K.; Maekawa, M.; Mano, N. Unconjugated Bile Acids in Rat Brain: Analytical Method Based on LC/ESI-MS/MS with Chemical Derivatization and Estimation of Their Origin by Comparison to Serum Levels. Steroids 2017, 125, 107–113. [Google Scholar] [CrossRef]
- Stinco, C.M.; Benítez-González, A.M.; Meléndez-Martínez, A.J.; Hernanz, D.; Vicario, I.M. Simultaneous Determination of Dietary Isoprenoids (Carotenoids, Chlorophylls and Tocopherols) in Human Faeces by Rapid Resolution Liquid Chromatography. J. Chromatogr. A 2019, 1583, 63–72. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, T.; Xiang, T.; Dong, Y.; Zhang, J.; Zhang, L. Quantitation of Isoprenoids for Natural Rubber Biosynthesis in Natural Rubber Latex by Liquid Chromatography with Tandem Mass Spectrometry. J. Chromatogr. A 2018, 1558, 115–119. [Google Scholar] [CrossRef]
- Baidoo, E.E.K.; Wang, G.; Joshua, C.J.; Benites, V.T.; Keasling, J.D. Liquid Chromatography and Mass Spectrometry Analysis of Isoprenoid Intermediates in Escherichia Coli BT—Microbial Metabolomics: Methods and Protocols; Baidoo, E.E.K., Ed.; Springer: New York, NY, USA, 2019; pp. 209–224. [Google Scholar] [CrossRef]
- Chhonker, Y.S.; Haney, S.L.; Bala, V.; Holstein, S.A.; Murry, D.J. Simultaneous Quantitation of Isoprenoid Pyrophosphates in Plasma and Cancer Cells Using LC-MS/MS. Molecules 2018, 23, 3275. [Google Scholar] [CrossRef] [Green Version]
- Lim, D.; Ikeda, A.; Vu, K.K.-T.; Yamaguchi, K.T.; Tyner, T.R.; Hasson, A.S. Method Development for the Measurement of Quinone Levels in Urine. J. Chromatogr. B 2011, 879, 3592–3598. [Google Scholar] [CrossRef]
- Kaiser, P.; Geyer, R.; Surmann, P.; Fuhrmann, H. LC–MS Method for Screening Unknown Microbial Carotenoids and Isoprenoid Quinones. J. Microbiol. Methods 2012, 88, 28–34. [Google Scholar] [CrossRef]
- Lukasiewicz, J.; Jachymek, W.; Niedziela, T.; Kenne, L.; Lugowski, C. Structural Analysis of the Lipid A Isolated from Hafnia Alvei 32 and PCM 1192 Lipopolysaccharides. J. Lipid Res. 2010, 51, 564–574. [Google Scholar] [CrossRef] [Green Version]
- Sándor, V.; Dörnyei, Á.; Makszin, L.; Kilár, F.; Péterfi, Z.; Kocsis, B.; Kilár, A. Characterization of Complex, Heterogeneous Lipid A Samples Using HPLC–MS/MS Technique I. Overall Analysis with Respect to Acylation, Phosphorylation and Isobaric Distribution. J. Mass Spectrom. 2016, 51, 1043–1063. [Google Scholar] [CrossRef]
- Sawyer, W.S.; Wang, L.; Uehara, T.; Tamrakar, P.; Prathapam, R.; Mostafavi, M.; Metzger IV, L.E.; Feng, B.; Baxter Rath, C.M. Targeted Lipopolysaccharide Biosynthetic Intermediate Analysis with Normal-Phase Liquid Chromatography Mass Spectrometry. PLoS ONE 2019, 14, e0211803. [Google Scholar] [CrossRef]
- Paulus, C.; Rebets, Y.; Zapp, J.; Rückert, C.; Kalinowski, J.; Luzhetskyy, A. New Alpiniamides From Streptomyces Sp. IB2014/011-12 Assembled by an Unusual Hybrid Non-Ribosomal Peptide Synthetase Trans-AT Polyketide Synthase Enzyme. Front. Microbiol. 2018, 9, 1959. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Xu, S.; Jiang, F.; Liu, W. Genomic-Driven Discovery of an Amidinohydrolase Involved in the Biosynthesis of Mediomycin A. Appl. Microbiol. Biotechnol. 2018, 102, 2225–2234. [Google Scholar] [CrossRef] [PubMed]
- Bhan, N.; Cress, B.F.; Linhardt, R.J.; Koffas, M. Expanding the Chemical Space of Polyketides through Structure-Guided Mutagenesis of Vitis Vinifera Stilbene Synthase. Biochimie 2015, 115, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, G.; Lucini, L.; Giuberti, G.; Bhumireddy, S.R.; Mandal, R.; Trevisan, M.; Wishart, D.S. Transformation of Polyphenols Found in Pigmented Gluten-Free Flours during in Vitro Large Intestinal Fermentation. Food Chem. 2019, 298, 125068. [Google Scholar] [CrossRef] [PubMed]
- Wen, Q.; Li, H.-L.; Mai, S.-Y.; Tan, Y.-F.; Chen, F. Tissue Distribution of Active Principles from Alpiniae Oxyphyllae Fructus Extract: An Experimental Study in Rats. Curr. Pharm. Anal. 2019, 286–293. [Google Scholar] [CrossRef]
- Chen, F.; Li, H.-L.; Tan, Y.-F.; Li, Y.-H.; Lai, W.-Y.; Guan, W.-W.; Zhang, J.-Q.; Zhao, Y.-S.; Qin, Z.-M. Identification of Known Chemicals and Their Metabolites from Alpinia Oxyphylla Fruit Extract in Rat Plasma Using Liquid Chromatography/Tandem Mass Spectrometry (LC–MS/MS) with Selected Reaction Monitoring. J. Pharm. Biomed. Anal. 2014, 97, 166–177. [Google Scholar] [CrossRef]
- Manning, S.R.; La Claire II, J.W. Isolation of Polyketides from Prymnesium Parvum (Haptophyta) and Their Detection by Liquid Chromatography/Mass Spectrometry Metabolic Fingerprint Analysis. Anal. Biochem. 2013, 442, 189–195. [Google Scholar] [CrossRef]
- Rodríguez-López, C.E.; Hernández-Brenes, C.; Treviño, V.; Díaz de la Garza, R.I. Avocado Fruit Maturation and Ripening: Dynamics of Aliphatic Acetogenins and Lipidomic Profiles from Mesocarp, Idioblasts and Seed. BMC Plant Biol. 2017, 17, 159. [Google Scholar] [CrossRef] [Green Version]
- Lísa, M.; Holčapek, M. High-Throughput and Comprehensive Lipidomic Analysis Using Ultrahigh-Performance Supercritical Fluid Chromatography–Mass Spectrometry. Anal. Chem. 2015, 87, 7187–7195. [Google Scholar] [CrossRef]
- Wood, P.L.; Scoggin, K.; Ball, B.A.; Troedsson, M.H.; Squires, E.L. Lipidomics of Equine Sperm and Seminal Plasma: Identification of Amphiphilic (O-Acyl)-ω-Hydroxy-Fatty Acids. Theriogenology 2016, 86, 1212–1221. [Google Scholar] [CrossRef]
- Montefusco, D.J.; Allegood, J.C.; Spiegel, S.; Cowart, L.A. Non-Alcoholic Fatty Liver Disease: Insights from Sphingolipidomics. Biochem. Biophys. Res. Commun. 2018, 504, 608–616. [Google Scholar] [CrossRef]
- Yang, Y.; Lee, M.; Fairn, G.D. Phospholipid Subcellular Localization and Dynamics. J. Biol. Chem. 2018, 293, 6230–6240. [Google Scholar] [CrossRef] [Green Version]
- López-Bascón, M.A.; Calderón-Santiago, M.; Sánchez-Ceinos, J.; Fernández-Vega, A.; Guzmán-Ruiz, R.; López-Miranda, J.; Malagon, M.M.; Priego-Capote, F. Influence of Sample Preparation on Lipidomics Analysis of Polar Lipids in Adipose Tissue. Talanta 2018, 177, 86–93. [Google Scholar] [CrossRef]
- Bollinger, J.G.; Ii, H.; Sadilek, M.; Gelb, M.H. Improved Method for the Quantification of Lysophospholipids Including Enol Ether Species by Liquid Chromatography-Tandem Mass Spectrometry. J. Lipid Res. 2010, 51, 440–447. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Kalra, H.; Gupta, A.; Raut, B.; Hussain, A.; Rahman, M.A. HybridSPE: A Novel Technique to Reduce Phospholipid-Based Matrix Effect in LC-ESI-MS Bioanalysis. J. Pharm. Bioallied Sci. 2012, 4, 267–275. [Google Scholar] [CrossRef]
- Criado-Navarro, I.; Mena-Bravo, A.; Calderón-Santiago, M.; Priego-Capote, F. Determination of Glycerophospholipids in Vegetable Edible Oils: Proof of Concept to Discriminate Olive Oil Categories. Food Chem. 2019, 299, 125136. [Google Scholar] [CrossRef]
- Helmer, P.O.; Korf, A.; Hayen, H. Analysis of Artificially Oxidized Cardiolipins and Monolyso-Cardiolipins via Liquid Chromatography/High-Resolution Mass Spectrometry and Kendrick Mass Defect Plots after Hydrophilic Interaction Liquid Chromatography Based Sample Preparation. Rapid Commun. Mass Spectrom. 2020, 34, e8566. [Google Scholar] [CrossRef] [Green Version]
- John, C.; Werner, P.; Worthmann, A.; Wegner, K.; Tödter, K.; Scheja, L.; Rohn, S.; Heeren, J.; Fischer, M. A Liquid Chromatography-Tandem Mass Spectrometry-Based Method for the Simultaneous Determination of Hydroxy Sterols and Bile Acids. J. Chromatogr. A 2014, 1371, 184–195. [Google Scholar] [CrossRef]
- Ms, U.; Ferdosh, S.; Haque Akanda, M.J.; Ghafoor, K.; Rukshana, A.H.; Ali, M.E.; Kamaruzzaman, B.Y.; Fauzi, M.B.; Sharifudin Shaarani, H.S.; Islam Sarker, M.Z. Techniques for the Extraction of Phytosterols and Their Benefits in Human Health: A Review. Sep. Sci. Technol. 2018, 53, 2206–2223. [Google Scholar] [CrossRef]
- McDonald, J.G.; Thompson, B.M.; McCrum, E.C.; Russell, D.W. Extraction and Analysis of Sterols in Biological Matrices by High Performance Liquid Chromatography Electrospray Ionization Mass Spectrometry. Methods Enzymol. 2007, 432, 145–170. [Google Scholar] [CrossRef]
- Liakh, I.; Pakiet, A.; Sledzinski, T.; Mika, A. Modern Methods of Sample Preparation for the Analysis of Oxylipins in Biological Samples. Molecules 2019, 24, 1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodama, H. GC-MS Methods for Tobacco Constituents BT—Gas Chromatography/Mass Spectrometry; Linskens, H.F., Jackson, J.F., Eds.; Springer: Berlin/Heidelberg, Germany, 1986; pp. 277–298. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Stinco, C.M.; Brahm, P.M.; Vicario, I.M. Analysis of Carotenoids and Tocopherols in Plant Matrices and Assessment of Their In Vitro Antioxidant Capacity BT—Plant Isoprenoids: Methods and Protocols; Rodríguez-Concepción, M., Ed.; Springer: New York, NY, USA, 2014; pp. 77–97. [Google Scholar] [CrossRef]
- Yamada, T.; Bamba, T. Lipid Profiling by Supercritical Fluid Chromatography/Mass Spectrometry BT—Lipidomics; Wood, P., Ed.; Springer: New York, NY, USA, 2017; pp. 109–131. [Google Scholar] [CrossRef]
- Britton, G.; Young, A.J. Methods for the Isolation and Analysis of Carotenoids BT—Carotenoids in Photosynthesis; Young, A.J., Britton, G., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 409–457. [Google Scholar] [CrossRef]
- Hadj-Mahammed, M.; Badjah-Hadj-Ahmed, Y.; Meklati, B.Y. Behaviour of Polymethoxylated and Polyhydroxylated Flavones by Carbon Dioxide Supercritical Fluid Chromatography with Flame Ionization and Fourier Transform Infrared Detectors. Phytochem. Anal. 1993, 4, 275–278. [Google Scholar] [CrossRef]
- Lebeau, J.; Venkatachalam, M.; Fouillaud, M.; Petit, T.; Vinale, F.; Dufossé, L.; Caro, Y. Production and New Extraction Method of Polyketide Red Pigments Produced by Ascomycetous Fungi from Terrestrial and Marine Habitats. J. Fungi (Basel Switz.) 2017, 3, 34. [Google Scholar] [CrossRef] [Green Version]
- Nieman, D.C.; Kay, C.D.; Rathore, A.S.; Grace, M.H.; Strauch, R.C.; Stephan, E.H.; Sakaguchi, C.A.; Lila, M.A. Increased Plasma Levels of Gut-Derived Phenolics Linked to Walking and Running Following Two Weeks of Flavonoid Supplementation. Nutrients 2018, 10, 1718. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.P.; Tran, L.V.H.; Namgoong, H.; Kim, Y.-H. Applications of Different Solvents and Conditions for Differential Extraction of Lipopolysaccharide in Gram-Negative Bacteria. J. Microbiol. 2019, 57, 644–654. [Google Scholar] [CrossRef]
Figure 1.
LIPID MAPS categories and representative structures with calculated octanol/water partition coefficient (log P) using ChemAxon. Reported log P of solvents used in lipidomics are indicated below [30]. Color code represents relative polarity, non-polar (blue), and polar (red). Example of classes corresponds to Glycerolipids, DG(16:0/16:0/0:0)—L02010001; Sphingolipids, SP(16:0/16:0)—LMGP01010564; Glicerophospholipids, PC(16:0/16:0)—LMGP01010564; Sterol lipids, Cholesterol—LMST01010001; Fatty acyls, C16:0—LMFA01010001; Prenol lipids, 2E,6E-farnesol—LMPR0103010001; Polyketides, Pinosylvin—LMPK13090001; Saccharolipids, 2,3-di-0-hexanoyl-α-glucopyranose—LMSL05000001.
Figure 1.
LIPID MAPS categories and representative structures with calculated octanol/water partition coefficient (log P) using ChemAxon. Reported log P of solvents used in lipidomics are indicated below [30]. Color code represents relative polarity, non-polar (blue), and polar (red). Example of classes corresponds to Glycerolipids, DG(16:0/16:0/0:0)—L02010001; Sphingolipids, SP(16:0/16:0)—LMGP01010564; Glicerophospholipids, PC(16:0/16:0)—LMGP01010564; Sterol lipids, Cholesterol—LMST01010001; Fatty acyls, C16:0—LMFA01010001; Prenol lipids, 2E,6E-farnesol—LMPR0103010001; Polyketides, Pinosylvin—LMPK13090001; Saccharolipids, 2,3-di-0-hexanoyl-α-glucopyranose—LMSL05000001.


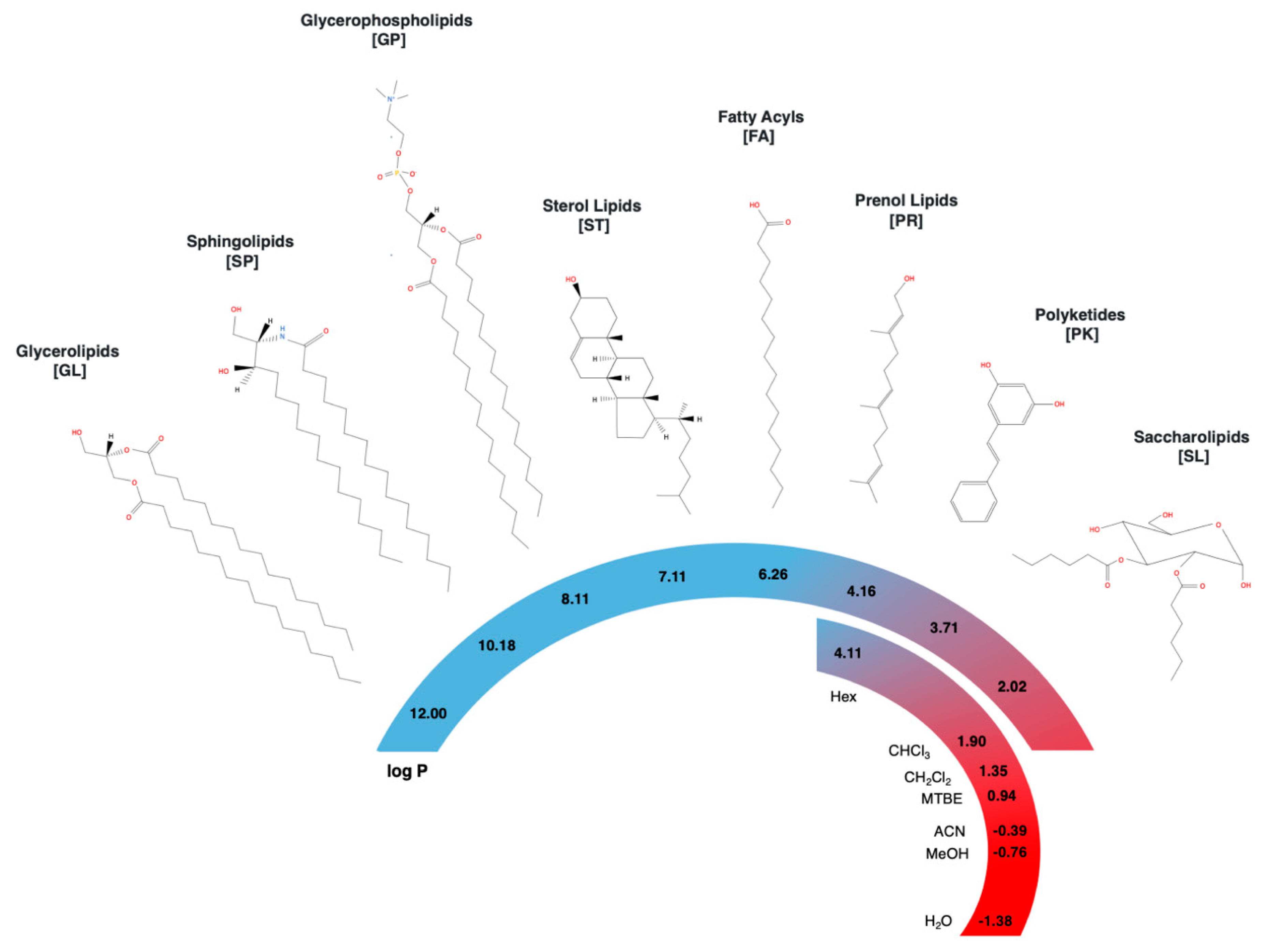{kind=link}
Table 1.
Number of lipids structures per representative lipid category.
| Lipid Category | Main Subclasses | Log P Range a | LIPID Maps b | |||
|---|---|---|---|---|---|---|
| Curated | Computationally-Generated | All | ||||
| Fatty Acyls [FA] | Fatty Acids and Conjugates, Eicosanoids, Docosanoids, Fatty esters, Fatty amides, Fatty nitriles, Fatty ethers, Fatty acyl glycosides, Acylcarnitines. | −5–15 | 7644 | 1792 | 9436 | |
| Glycerolipids [GL] | Monoradylglycerols, Diradylglycerols, Triradylglycerols, Glycosylmonoradylglycerols, Glycosyldiradylglycerols. | 5–35 | 232 | 7379 | 7611 | |
| Glycerophospholipids [GP] | Glycerophosphocholines, Glycerophosphoethanolamines, Glycerophosphoserines, Glycerophosphoglycerols, Glycerophosphoglycerophosphates, Glycerophosphoinositols, Oxidized glycerophospholipids, Cardiolipins. | 5–25 | 1607 | 8312 | 9919 | |
| Sphingolipids [SP] | Sphingoid bases, Ceramides, Phosphosphingolipids, Neutral glycosphingolipids, Acidic glycosphingolipids, Basic glycosphingolipids. | 5–25 | 1410 | 3176 | 4586 | |
| Sterol lipids [ST] | Sterols, Steroids, Secosteroids, Bile acids and derivatives, Steroid conjugates. | 0–20 | 2829 | 2829 | ||
| Prenol lipids [PR] | Isoprenoids, Quinones and hydroquinones, Polyprenols. | 0–20 | 1352 | 1352 | ||
| Sacccharolipids [SL] | Acylaminosugars, Acylaminosugar glycans, Acyltrehaloses. | 0–30 | 22 | 1294 | 1316 | |
| Polyketides [PK] | Linear polyketides, Macrolides and lactone polyketides, Linear tetracyclines, Polyether antibiotics, Aflatoxins, Flavonoids, Aromatic polyketides. | 0–15 | 6810 | 6810 | ||
| TOTAL | 21,906 | 21,953 | 43,859 | |||
a Octanol/water partition coefficient (log P) calculated using ChemAxon. b Data taken from Lipid Maps® Structure Database (LMSD) in the 05/02/2020 update.
Table 2.
Comparison between extraction methods used in mass spectrometry (MS)-based untargeted lipidomics.
Table 2.
Comparison between extraction methods used in mass spectrometry (MS)-based untargeted lipidomics.
| Extraction Method | Principle | Protocol Name | System | Lipid Coverage | Lipid Recovery | Solvent-Efficiency | Carry-Over Free | Time-Efficiency | Repeatability | Versatility | Automatization | Biological Matrix | Platform | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PPT | Precipitation | OPE | iPrOH, ButOH/MeOH (2:2) CHCl3/MeOH (2:2) | Plasma | HPLC-ESI-QTRAP | [38,39,40] | ||||||||
| CSF | UHPLC-ESI-QTOF | [41] | ||||||||||||
| LLE | Partition | Folch * | CHCl3/MeOH (2:1) | Cell line/tissue | UHPLC-ESI-Q-Exactive Orbitrap | [42] | ||||||||
| CSF | HPLC-ESI-Q-Exactive Orbitrap | [43] | ||||||||||||
| Folch * BD * | CHCl3/MeOH (2:1) CHCl3 or CH2Cl2/MeOH (1:2) | Plasma/CSF Food | HPLC-ESI-Ion trap | [44] | ||||||||||
| UHPLC-Q-Exactive Orbitrap | [45,46] | |||||||||||||
| Tears | UHPLC-TripleTOF | [47] | ||||||||||||
| Plants | HPLC-ESI-QTOF | [48] | ||||||||||||
| Urine | UHPLC-ESI-3D-Ion trap | [49] | ||||||||||||
| Feces | HPLC-ESI-Q-Exactive Orbitrap | [50] | ||||||||||||
| Cell culture | HPLC-ESI-Q-Exactive Orbitrap | [43] | ||||||||||||
| BD * MTBE * | CHCl3 or CH2Cl2/MeOH (1:2) MTBE/MeOH (5:1.5) | Animal tissue | DI-ESI-QTRAP | [51] | ||||||||||
| Urine/saliva | ASAP-QTOF | [52] | ||||||||||||
| Plant tissue | UHPLC-ESI-IT-TOF | [53,54] | ||||||||||||
| Feces | HPLC-ESI-Q-Exactive Orbitrap | [50] | ||||||||||||
| Plasma | HPLC-ESI-QTOF | [55] | ||||||||||||
| MTBE * BUME | MTBE/MeOH (5:1.5) 1. ButOH/MeOH (3:1) 2. Hep/EtOAc (3:1) 3. 1% AcOH | Urine | HPLC-ESI-QTOF | [56] | ||||||||||
| Food | UHPLC-ESI-TOF | [57] | ||||||||||||
| Breast Milk | HPLC-ESI-QTOF | [58] | ||||||||||||
| Plant tissue | UHPLC-ESI-Q-Exactive Orbitrap | [59] | ||||||||||||
| Cell line | HPLC-LTQ-Orbitrap | [60] | ||||||||||||
| Plant tissue | UHPLC-ESI-Q-Exactive Orbitrap | [61] | ||||||||||||
| Cell line Plasma/SBF Animal tissue | UHPLC-ESI-QTOF | [62] | ||||||||||||
| HPLC-ESI-QqQ | [63] | |||||||||||||
| DI-ESI-QTRAP | [51] | |||||||||||||
| BUME IPA/Hex * | 1. ButOH/MeOH (3:1) 2. Hep/EtOAc (3:1) 3. 1% AcOHiPrOH/Hex (3:2) | Cell lines | UHPLC-ESI-Q-Exactive Orbitrap | [64] | ||||||||||
| Cell lines | UHPLC-ESI-Q-Exactive Orbitrap | [61] | ||||||||||||
| Cell lines | UHPLC-ESI-QTOF | [62] | ||||||||||||
| IPA/Hex * SFE | iPrOH/Hex (3:2) Supercritical CO2 | Serum Parasites | DI-ESI-TOF | [65] | ||||||||||
| HPLC-ESI-QqQ | [66] | |||||||||||||
| Plasma | SFE-ESI-QTRAP | [67] | ||||||||||||
| Parasites | UHPSFC-ESI-IT-TOF | [68] | ||||||||||||
| SPE | Coated fibers and capillary tubes (sorbent cartridge). | Animal tissue Plasma | UHPSFC-ESI-IT-TOF | [69] | ||||||||||
| HPLC-ESI-Q-Exactive Orbitrap | [70] | |||||||||||||
| SPE | Adsorbance | SPE SPME | Coated fibers and capillary tubes (sorbent cartridge). Diameter-reduced sorbent-coated rods/fibers | Saliva | HPLC-ESI-Q-Exactive Orbitrap | [71] | ||||||||
| Breast Milk | HPLC-ESI-QTOF | [72] | ||||||||||||
| Cell lines | HPLC-ESI-Q-Exactive Orbitrap | [43] | ||||||||||||
| Urine | GC-EI-Q | [26,27] |
* Water is added for phase separation. OPE: One-phase extraction; PPT: Protein precipitation; BD: Bligh and Dyer; SFE: Supercritical fluid extraction; SPE: Solid-phase extraction; SPME: Solid-phase microextraction; BUME: Butanol-methanol; MTBE: Methyl tert-butyl ether; DI: Direct infusion; ASAP: Atmospheric Solids Analysis Probe; Color reference: Dark blue: Good; Blue: Fair; Light blue: Poor. References were selected for studies conducted during the 2009–2019 period.
Table 3.
Reports of targeted extraction of lipids by category.
| Lipid Class | Extraction Method | System | Biological Matrix (Sample Amount) | Platform | Ref |
|---|---|---|---|---|---|
| Fatty Acyls | |||||
| Fatty Acids and Conjugates | |||||
| 1. LLE, 2. SPE | 1. Modified Bligh and Dyer, CH3Cl - MeOH - H2O (1:1:1) with 2 mM HCl; 2. 3-aminopropyl silica gel/ACN-AcOH (49:1) | Food supplements (3 mL) | UHPLC-ESI-QTOF | [79] | |
| LLE | H2O with 0.5 M HCl - anhydrous Et2O (1:1) + Deriv. with BSTFA | Feces (30 mg) | GC-Q | [80] | |
| LLE | Modified Folch, CH3Cl-MeOH-H2O (2:1:0.8) + BHT | Grapes-skins-seeds (0.1 g) | UHPLC-ESI-QqQ | [81] | |
| LLE | Modified Folch, CH3Cl - MeOH - 0.73% NaCl (2:1:0.6) + Deriv. with 1% H2SO4 in MeOH | Green coffee (0.5 g) | GC-Q | [82] | |
| LLE | Hex - 0.5% NaOH in MEOH and 14% BF3 in MeOH - Sat. NaCl (1.3:2:4) | Shark liver oil (25 mg) | GC-Q | [83] | |
| 1. Filter pre-conc. 2. LLE | 1. Whatman GF/F filters, 2. CH2Cl2 - MeOH - H2O (1.5:3:1) | Oceanic water (5–10 L) | DI-ESI- FT-ICR MS | [84] | |
| Octadecanoids | |||||
| 1. OPE, 2. SPE | 1. MeOH + BHT; 2. < 8% MeOH/Polymeric RP/MeOH + BHT | Mice amygdala tissue | UHPLC-ESI-QTRAP | [85] | |
| Eicosanoids | |||||
| 1. LLE, 2. Hydrolysis, 3. SPE | 1. MTBE-MeOH-0.15 M NH4OAc (2:1:1) + BHT; 2. MeOH-H2O-10M NaOH (1:1:1.2); 3. Reac.Mix. + AcOH + 0.1 M Na2HPO4 buffer (pH 6)/AEC/tOAc-Hex (75:25) + 1%AcOH | Human cells (1 × 107) | HPLC-ESI-QTRAP | [86] | |
| LLE | Sample + AcOH (pH 3.5) - MTBE (2:1) + Deriv. for GC | Breath condensate (2 mL) | HPLC-ESI-QTRAP GC/NICI–MS | [87] | |
| SPE | MeOH (<17%) + Na2HPO4 buffer (pH 6)/AEC/EtOAc - Hex (75:25) with 1% AcOH | Plasma (500 µL) | UHPLC-ESI-QTRAP | [86] | |
| SPE | H2O + BHT/Polymeric RP/MeOH | Plasma (500 µL) | UPLC-ESI-QqQ | [88] | |
| SPE | 15% MeOH + 0.1 M HCl (pH 3.0)/C18/Methyl formate | Mouse brain tissue (25–75 mg) | HPLC-ESI-QqQ | [89,90] | |
| SPE | 10% MeOH/Polymeric RP/MeOH | Human cells-animal tissues | HPLC-ESI-QTRAP | [91] | |
| 1. OPE, 2. SPE | 1. MeOH + BHT; 2. < 8% MeOH/Polymeric RP/MeOH + BHT | Mice brain tissue | UHPLC-ESI-QTRAP | [85] | |
| SPE | 0.12 M PP buffer + 5 mM MgCl2 + BHT/Polymeric RP/MeOH | Human CSF and rat cortex | UPLC-ESI-QqQ | [92] | |
| Fatty Esters | |||||
| OPE | MeOH - 0.1% FA (0.8:1) | Plasma (20 µL)-urine (5 µL)-CSF (20 µL) | UHPLC-HESI-QTRAP | [93] | |
| 1. OPE, 2. OPE | 1. MeOH - H2O (4:1) + 0.1% FA; 2. MeOH - 0.1% FA (0.8:1) | Human brain tissue (10 mg) | UHPLC-HESI-Q-Orbitrap | [93] | |
| OPE | iPrOH | Serum (10 µL) | HPLC-ESI-QqQ | [94] | |
| OPE | AbsoluteIDQ p180 Kit | Serum (10 µL) | HPLC-ESI-QqQ | [95] | |
| OPE | AbsoluteIDQ p180 Kit | Plasma | UPLC-ESI-QqQ | [96] | |
| OPE | ACN + Deriv. with DnsHz | Plasma | UHPLC-ESI-QqQ | [97] | |
| 1. LLE, 2. SPE | 1. CHCl3 - MeOH - PBS (2:1:1); 2./CHCl3/Silica NP/EtOAc | Adipose tissue (150 mg) | HPLC-ESI-QqQ | [98] | |
| 1. LLE, 2. SPE | 1. CHCl3 - MeOH - PBS (2:1:0.86); 2./CHCl3/Silica NP/EtOAc | Serum or plasma (200 µL) | HPLC-ESI-QqQ | [98] | |
| 1. LLE, 2. SPE | 1. Bligh and Dyer, CHCl3-MeOH - H2O (2:1:1); 2. 0.1% NH4OH in ACN/AEC/1% FA in Acetone + AMPP | Plant leaves (100 mg) | UHPLC-ESI-QqQ | [99] | |
| SPE | 0.1% NH4OH in ACN (1:2)/AEC/1% FA in Acetone + Deriv. DMED and d4-DMED | Mice adipose tissue (100 mg) | UHPLC-ESI-QqQ | [100] | |
| 1. LLE, 2. SPE | 1. Modified Bligh and Dyer, CHCl3 - MeOH (1:1); 2. CHCl3/Silica NP/EtOAc | Plasma (100 µL), liver | nanoESI-QqQ | [101] | |
| Fatty amides | |||||
| SPE | 10% MeOH/Polymeric RP/MeOH | Human cell lines or animal tissues | HPLC-ESI-QTRAP | [91] | |
| SPE | MeOH - H2O (1:2.3)/C18/MeOH | Rat brain | HPLC-ESI-QqQ | [102] | |
| Glycerolipids | |||||
| Monoradylglycerols | |||||
| OPE | MeOH + Deriv. with d4-NPB and 3-NPB | Cells-Tissue (60 mg)-Serum (50 µL) | UPLC-ESI-QTOF | [103] | |
| OPE | iPrOH | Shark liver oil (10 mg) | HPLC-ESI-QqQ | [83] | |
| 1. Filter pre-conc. 2. LLE | 1. Whatman GF/F filters, 2. CH2Cl2 - MeOH - H2O (1.5:3:1) | Oceanic water (5–10 L) | DI-ESI- FT-ICR MS | [84] | |
| LLE | CHCl3 - MeOH - 25 M LiCl (1:1:1) | Plasma (25 µL) | DI-ESI-QTOF | [104] | |
| Diradylglycerols | |||||
| 1. OPE, 2. LLE | 1. 70% iPrOH, 2. Bligh and Dyer | Feces (2 mg) | FIA-ESI-Q-Orbitrap | [105] | |
| OPE | iPrOH | Shark liver oil (10 mg) | HPLC-ESI-QqQ | [83] | |
| LLE | Modified Bligh and Dyer, CHCl3 - MeOH - H2O (1:1:0.8) | Bacterial cells (OD: 0.3, 0.5, 0.8, 1.3) | HPLC-ESI-LTQ-Orbitrap | [106] | |
| LLE | Modified Matyash, MTBE - MeOH - H2O (3:0.9:0.75) | Skeletal muscle (50 mg) | HPLC-API-QqQ | [107] | |
| 1. Filter pre-conc. 2. LLE | 1. Whatman GF/F filters, 2. CH2Cl2 - MeOH - H2O (1.5:3:1) | Oceanic water (5–10 L) | DI-ESI- FT-ICR MS | [84] | |
| Triradylglycerols | |||||
| LLE | Modified Bligh and Dyer, CH2Cl2 - MeOH - H2O (1.5:3:1) | Serum (30 µL) | HPLC-HESI-Orbitrap | [108] | |
| OPE | EtOH - MTBE - DCM (70:15:15) | Size-fractionated serum (20 µL) | UPLC-ESI-QTRAP | [35] | |
| LLE | Modified Bligh and Dyer, CHCl3 - MeOH - H2O (1:1:0.8) | Bacterial cells (OD: 0.3, 0.5, 0.8, 1.3) | HPLC-ESI-LTQ-Orbitrap | [106] | |
| OPE | Acetone | Vegetal Oil (40 mg) | HPLC-ESI-Quadrupole | [109] | |
| 1. Filter pre-conc. 2. LLE | 1. Whatman GF/F filters, 2. CH2Cl2 - MeOH - H2O (1.5:3:1) | Oceanic water (5–10 L) | DI-ESI- FT-ICR MS | [84] | |
| 1. OPE, 2. LLE | 1. 70% iPrOH, 2. Bligh and Dyer | Feces (2 mg) | FIA-ESI-Q-Orbitrap | [105] | |
| Glycosyldiradylglycerols | |||||
| 1. pre-treat, 2. LLE | 1. iPrOH (75 °C) + BHT; 2. CHCl3 - MeOH (2:1) | Vegetal tissue (2–3 plant rosettes) | ESI-QqQ | [110] | |
| LLE | Bligh and Dyer | Algae tissue (30 L culture) | UPLC-ESI-QTOF | [111] | |
| Glycerophospholipids | |||||
| Glycerophosphocholines | |||||
| OPE | AbsoluteIDQ p150 Kit | Plasma (10 µL) | FIA-ESI-QTRAP | [112] | |
| OPE | ACN | Plasma (20 µL) | UPLC-ESI-QTRAP | [113] | |
| OPE | MeOH + BHT | Plasma (10 µL) | HPLC-ESI-QTRAP | [114] | |
| LLE | CH2Cl2 - MeOH (2:1) | Cytosol (100 µL) | UPLC-ESI-QTRAP | [115] | |
| LLE | Modified Folch, CHCl3 - MeOH - 0.15 M NaCl (2:1:0.8) + BHT | Lenses | ESI-QqQ | [116] | |
| LLE | Modified Bligh and Dyer, CHCl3 - MeOH - 0.1N HCl (1:1:1) | Human cells (2 × 106) | UPLC-ESI-QTOF | [117,118] | |
| µChip-SPE | Lysozyme in 20 mM Tris-HCl (pH 7.5)/Silica beads/MeOH | Bacterial cells (10µL) | nanoESI-QTOF | [119] | |
| OPE | EtOH - MTBE - DCM (7:1.5:1.5) | Size-fractionated serum (20 µL) | UPLC-ESI-QTRAP | [35] | |
| LLE | Bligh and Dyer | Algae tissue (30 L culture) | UPLC-ESI-QTOF | [111] | |
| Glycerophosphoethanolamines | |||||
| OPE | ACN | Plasma (20 µL) | UPLC-ESI-QTRAP | [113] | |
| OPE | MeOH + BHT | Plasma (20 µL) | HPLC-ESI-QTRAP | [114] | |
| LLE | CH2Cl2 - MeOH (2:1) | Cytosol (100 µL) | UPLC-ESI-QTRAP | [115] | |
| LLE | Modified Folch, CHCl3 - MeOH - 0.15 M NaCl (2:1:0.8) + BHT | Lenses | DI-ESI-QqQ | [116] | |
| LLE | Modified Bligh and Dyer, CHCl3 - MeOH - 0.1N HCl (1:1:1) | Human cells (2 X 106) | UPLC-ESI-QTOF | [117,118] | |
| µChip-SPE | Lysozyme in 20 mM Tris-HCl (pH 7.5)/Silica beads/MeOH | Bacterial cells (10 µL) | nanoESI-QTOF | [119] | |
| OPE | EtOH - MTBE - DCM (7:1.5:1.5) | Size-fractionated serum (20 µL) | UPLC-ESI-QTRAP | [35] | |
| LLE | Bligh and Dyer | Algae tissue (30 L culture) | UPLC-ESI-QTOF | [111] | |
| Glycerophosphoserines | |||||
| OPE | ACN | Plasma (20 µL) | UPLC-ESI-QTRAP | [113] | |
| LLE | CH2Cl2 - MeOH (2:1) | Cytosol (100 µL) | UPLC-ESI-QTRAP | [115] | |
| LLE | Bligh and Dyer | Human cells (0.4 × 106) | ESI-QTRAP | [120] | |
| LLE | Modified Folch, CHCl3 - MeOH - 0.15 M NaCl (2:1:0.8) + BHT | Lenses | ESI-QqQ | [116] | |
| µChip-SPE | Lysozyme in 20 mM Tris-HCl (pH 7.5)/Silica beads/MeOH | Bacterial cells (10µL) | nanoESI-QTOF | [119] | |
| Glycerophosphoglycerols | |||||
| OPE | ACN | Plasma (20 µL) | UPLC-ESI-QTRAP | [113] | |
| LLE | Modified Bligh and Dyer, CHCl3 - MeOH - 0.1N HCl (1:1:1) | Human cells (0.4 × 106) | UPLC-ESI-QTOF | [117,118] | |
| µChip-SPE | Lysozyme in 20 mM Tris-HCl (pH 7.5)/Silica beads/MeOH | Bacterial cells (10 µL) | nanoESI-QTOF | [119] | |
| LLE | Modified Bligh and Dyer, CHCl3 - MeOH - H2O (1:1:0.8) | Bacterial cells (OD: 0.3, 0.5, 0.8, 1.3) | UPLC-ESI-QqQ | [106] | |
| LLE | Bligh and Dyer | Algae tissue (30 L culture) | UPLC-ESI-QTOF | [111] | |
| Glycerophosphoinositols | |||||
| OPE | ACN | Plasma (20 µL) | UPLC-ESI-QTRAP | [113] | |
| LLE | CH2Cl2 - MeOH (2:1) | Cytosol (100 µL) | UPLC-ESI-QTRAP | [115] | |
| LLE | Modified Bligh and Dyer, CHCl3 - MeOH - 0.1N HCl (1:1:1) | Human cells (2 × 106) | UPLC-ESI-QTOF | [117,118] | |
| µChip-SPE | Lysozyme in 20 mM Tris-HCl (pH 7.5)/Silica beads/MeOH | Bacteria Cells (10 µL) | nanoESI-QTOF | [119] | |
| OPE | EtOH - MTBE - DCM (7:1.5:1.5) | Size-fractionated serum (20 µL) | UPLC-ESI-QTRAP | [35] | |
| Glycerophosphates | |||||
| OPE | ACN | Plasma (20 µL) | UPLC-ESI-QTRAP | [113] | |
| µChip-SPE | Lysozyme in 20 mM Tris-HCl (pH 7.5)/Silica beads/MeOH | Bacterial cells (10µL) | nanoESI-QTOF | [119] | |
| Glycerophosphoglycerophosphoglycerols | |||||
| LLE | Modified Bligh and Dyer, CHCl3 - MeOH - H2O (1:1:0.8) | Bacterial cells (OD: 0.3, 0.5, 0.8, 1.3) | HPLC-ESI-QTOF | [106] | |
| Oxidized glycerophospholipids | |||||
| µHP-SPE | MeOH/C18 spin column/MeOH - 0.2% FA (82:18) | Plasma (20 µL) | HPLC-ESI-QTRAP | [121] | |
| Sphingolipids | |||||
| Sphingoid bases | |||||
| LLE | Modified Bligh and Dyer, CHCl3 - MeOH -H2O (1.8:2:0.8) + 0.1% TFA | Plasma (50 µL) | UHPLC-ESI-QqQ | [122] | |
| LLE | Bligh and Dyer, CHCl3 - MeOH - H2O (1:2:0.8) | B Cells (80 µL cell suspension) | HPLC-ESI-QTRAP | [123] | |
| LLE | MTBE - MeOH - H2O (3:0.9:0.8) | Plasma-Red blood cells (50 µL) | UHPLC-ESI-QqQ | [124] | |
| LLE + Transesterification | 0.25M MeONa in MeOH - MTBE - H2O (1.3:4:1) + AcOH (pH 7) | Serum (40 µL) | UHPLC-ESI-QqQ | [125] | |
| LLE + Sap. | CH2Cl2 - MeOH - KOH 1M in MeOH (1:2:0.3) | Cells (106)-Tissue (1–10 mg) | LC-MS/MS techniques | [126] | |
| LLE | CH 2Cl2 - MeOH (1:1) with 0.25% DEA | Plasma (25 µL) | HPLC-ESI-QqQ | [127] | |
| LLE | ButOH - 40 mM Na2HPO4 + CA (pH4) (1:1) | Plasma (75 µL) | HPLC-ESI-QTRAP | [128] | |
| Ceramides | |||||
| LLE | Modified Bligh and Dyer, CHCl3 - MeOH -H2O (1.8:2:0.8) + 0.1% TFA | Plasma (50 µL) | UHPLC-ESI-QqQ | [122] | |
| LLE | Bligh and Dyer, CHCl3 - MeOH - H2O (1:2:0.8) | B Cells (80 µL cell suspension) | HPLC-ESI-QTRAP | [123] | |
| LLE + Sap. | CHCl3 - MeOH - 1M KOH in MeOH (1:2:0.3) | Cells (106)-Tissue (1–10 mg) | LC-MS/MS techniques | [126] | |
| LLE | CH2Cl2 - MeOH (1:1) with 0.25% DEA | Plasma (25 µL) | HPLC-ESI-QqQ | [127] | |
| LLE + Transesterification | 0.25M MeONa in MeOH - MTBE - H2O (1.3:4:1) + AcOH (pH 7) | Serum (40 µL) | UHPLC-ESI-QqQ | [125] | |
| LLE | ButOH - 40 mM Na2HPO4 + CA (pH4) (1:1) | Plasma (75 µl) | HPLC-ESI-QTRAP | [128] | |
| 1. OPE, 2. SPE | 1.MeOH, 2.Hex-iPrOH (11:1)/aminopropyl silica cartridges/Hex-CHCl3-MeOH (80:10:10) | Skin (Three patches) | UHPLC-ESI-QTOF | [129] | |
| Phosphosphingolipids | |||||
| LLE | Modified Bligh and Dyer, CHCl3 - MeOH -H2O (1.8:2:0.8) + 0.1% TFA | Plasma (50 µL) | UHPLC-ESI-QqQ | [122] | |
| LLE | CHCl3 - MeOH - 1M KOH in MeOH (1:2:0.3) | Cells (106)-Tissue (1–10 mg) | LC-MS/MS techniques | [126] | |
| LLE + Transesterification | 0.25M MeONa in MeOH - MTBE - H2O (1.3:4:1) + AcOH (pH 7) | Serum (40 µL) | UHPLC-ESI-QqQ | [125] | |
| OPE | MeOH | Whole blood (15 µL on DBS) | UHPLC-ESI-TOF | [130] | |
| OPE | AbsoluteIDQ p150 Kit | Plasma (10 µL) | FIA-ESI-QTRAP | [112] | |
| Neutral glycosphingolipids | |||||
| LLE | Bligh and Dyer, CHCl3 - MeOH - H2O (1:2:0.8) | B Cells (80 µL cell suspension) | HPLC-ESI-QTRAP | [123] | |
| LLE | CHCl3 - MeOH - 1M KOH in MeOH (1:2:0.3) | Cells (106)-Tissue (1–10 mg) | LC-MS/MS techniques | [126] | |
| LLE | ButOH - 40 mM Na2HPO4 + CA (pH 4) (1:1) | Plasma (75 µL) | HPLC-ESI-QTRAP | [128] | |
| LLE | MTBE - MeOH - H2O (3:0.9:0.8) | Plasma-Red blood cells (50 µL) | UHPLC-ESI-QqQ | [124] | |
| Acidic glycosphingolipids | |||||
| LLE | CHCl3 - MeOH - H2O (0.8:1:1) + Deriv. with PAEA and DMTMM to aq. phase | Plasma (20 µL) | UHPLC-ESI-QTRAP | [131] | |
| 1. LLE, 2. SPE | 1. Modified Folch, CHCl3 - MeOH -H2O (2:1:0.6) 2.C18/MeOH to aq. phase | Plasma (200 µL)-Human tissues (25 mg) | HPLC-ESI-QTOF | [132] | |
| LLE | CH2Cl2 - MeOH (1:1) with 0.25% DEA | Plasma (25 µL) | HPLC-ESI-QqQ | [127] | |
| Sterol lipids | |||||
| Sterols | |||||
| SPE | Hex - Et2O (99:1)/NP silica cartridge/Hex - Et2O (99:1) | Sunflower oils (200 mg) | ESI-QTRAP-QqQ | [133] | |
| SPE | iPrOH/Polymeric RP/MeOH - 0.02% FA (10:90) | Plasma-Serum-CSF (100 µL) | HPLC-ESI-QTRAP | [134] | |
| LLE | Bligh and Dyer like method + Deriv. with AcCl - CHCl3 (1:5) | Ocular Tissue | HPLC-ESI-QTrap-Orbitrap | [135] | |
| 1. OPE + Esterif. 2. LLE | 1. iPrOH 70% + 5M NaOH (1M HCl), 2. iOct - Reaction Mix. (2:1) + Deriv. with MSTFA | Feces (2 g) | GC-QqQ | [136] | |
| 1. OPE + Esterif. 2. LLE | 1. iPrOH 70%+5M NaOH (1M HCl), 2. iOct-Reaction Mix.(2:1)+Deriv. with DMG+DMAP | Feces (2 g) | UHPL-HESI-Q-Orbitrap | [137] | |
| LLE | Hex - iPrOH - 0.47M Na2SO4 (2:3:1.5) + BHT | Atherosclerotic plaques (10 mg) | HPLC-APCI-QqQ | [138] | |
| 1. OPE, 2. LLE | 1. 50 mM Tris-HCl (pH 7.5), 150 mM NaCl and 2mM EGTA, 2. Modified Bligh and Dyer, CHCl3 - MeOH - Sample (1:1:0.9) | Silkworm tissues | HPLC-ESI-QqQ | [139] | |
| Steroids | |||||
| LLE | MTBE | Serum (100 µl) | HPLC-ESI-QTRAP | [140] | |
| SLE | Acetate buffer (pH 5.2)/Diat. Earth/CH2Cl2 + Deriv. with MSTFA - NH4I − DTE (500:4:2) | Serum (100 µl) | GC-QqQ | [141] | |
| SPE | 5% H3PO4/Polymeric RP/MeOH | Plasma (100 µL) | HPLC-ESI-TripleTOF | [142] | |
| Bile acids and derivatives | |||||
| OPE | Methanol | Rat serum (10 μL) | UPLC-ESI-QTRAP/QTOF | [143] | |
| A. OPE or D. LLE | A. ACN; D: ACN - 400 g/L NH4SO4 - H2O (1:1:0.35) | Serum (100 µL) | HPLC-ESI-QqQ | [144] | |
| PD-SPE | Ostro 96 well plates/cold ACN with 1% of FA | Plasma-Serum (100 µL) | UHPLC-ESI-QqQ | [145] | |
| SPE | iPrOH/Polymeric RP/MeOH - 0.02% FA (10:90) | Plasma-Serum-CSF (100 µL) | HPLC-ESI-QTRAP | [134] | |
| OPE | iPrOH | Feces (1 g) | HPLC-ESI-QqQ | [146] | |
| 1. OPE, 2. SPE | 1. Saline solution - (60 °C) Ethanol (1:1); 2. Extract - H2O (1:10)/Polymeric RP/MeOH | Rat brain tissue (1.5–1.8 g) | HPLC-ESI-QqQ | [147] | |
| Prenol lipids | |||||
| Isoprenoids | |||||
| LLE | Et2O - PBS (pH = 7.4) - EtOH (1:1:0.2) | Feces (0.3–0.5 g) | HPLC-DAD | [148] | |
| OPE | ACN - MeOH - H2O (2:1:1) | Natural rubber (1.5 g) | HPLC-ESI-QqQ | [149] | |
| OPE | Methanol - H2O (1:1) | Bacterial cells (1.5 mL, OD: 5) | HPLC-ESI-TOF | [150] | |
| OPE | Methanol - H2O (1:3) | Bacterial supernatant (1.5 mL) | HPLC-ESI-TOF | [150] | |
| OPE | iPrOH - 100 mM NH4HCO3 (pH 7.4) (1:1) | Human cells | UPLC-ESI-QqQ | [151] | |
| SPE | 2% FA/Polymeric RP/Hex - iPrOH - NH4OH (12:7:1) | Plasma (300 µL) | UPLC-ESI-QqQ | [151] | |
| Quinones and hydroquinones | |||||
| LLE | Saturated K2CO3 - CH2Cl2 (1:1) | Urine (Rats: 2 mL Human: 10 mL) | GC-Q | [152] | |
| LLE | CHCl3 - MeOH (3:7) - cold 10% NaCl + BHT | Bacterial cells (10–50 mg) | HPLC-APCI-QTRAP | [153] | |
| Saccharolipids | |||||
| Acylaminosugars | |||||
| 1. LLE, 2. LLE | 1. 45% Phenol - H2O; 2. CHCl3 - MeOH - H2O (2:1:3) + Deriv. aq phase. | Bacterial cells (2 g/mL) | GC-Q | [154] | |
| LLE | 50% Phenol - H2O | Acetone-dried bacteria | UHPLC-ESI-QTOF | [155] | |
| SPE | Genlantis SoluLyse detergent/AEC/5% NH4OH in MeOH | Supernatant cell lysate (2×900 μL) | UPLC-ESI-QTRAP | [156] | |
| Polyketides | |||||
| Linear polyketides | |||||
| LLE | EtOAc | Bacterial supernatant (10 mL) | HPLC-ESI-LTQ-Orbitrap | [157] | |
| OPE | Acetone - MeOH (1:1) | Bacterial cells (10 mL culture) | HPLC-ESI-LTQ-Orbitrap | [157] | |
| OPE | MeOH | Bacterial supernatant (10 mL) | UPLC-ESI-QTOF | [158] | |
| Aromatic polyketides | |||||
| LLE | EtOAc + 1% HCl | Enzymatic mixture (500 µL) | HPLC-ESI-LTQ-Orbitrap | [159] | |
| Flavonoids | |||||
| OPE | 70% MeOH + 3% FA | Digested cooked flour (1 mL) | UHPLC-HESI-QTRAP | [160] | |
| LLE | EtOAc | Blood-Tissue (50 µL) | UFLC-ESI-QqQ | [161,162] | |
| Polyether antibiotics | |||||
| 1. OPE, 2. OPE, 3. LLE, 4. SPE | 1. MeOH; 2. 80% PrOH; 3. EtOAc - H2O (1:1); 4. aq. phase/C18/80% PrOH | Microalga cells pellet (125–150 mL culture) | UPLC-nanoESI-QTOF | [163] | |
| Annonaceae acetogenins | |||||
| 1. OPE, 2. LLE | 1. Acetone; 2. CH2Cl2 - H2O (1:1) | Avocado mesocarp (2g)-cotyledons (1g) | HPLC-ESI-TOF | [164] | |
AcCl: Acetyl chloride, Ace: Acetone, ACN: Acetonitrile, AcOH: Acetic acid, AEC: Anion exchange cartridge, AMPP: N-[4-(aminomethyl)phenyl]pyridinium, APCI: Atmospheric pressure chemical ionization, API: Atmospheric pressure ionization, aq.: aqueous, BHT: Butylated hydroxytoluene, BHT: Butylhydroxytoluene, BSTFA: N,O-Bis(trimethylsilyl)trifluoroacetamide, ButOH: n-Butanol, CA: Citric acid, Conc.: Concentration, CSF: cerebrospinal fluid, DAD: Diode Array Detection, DEA: Diethylamine, Deriv.: Derivatization, Diat.: Diatomaceous, DI: direct-infusion, DMAP: N,N-dimethylpyridin-4-amine, DMED: N,N-dimethylethylenediamine, DMG: N,N-dimethylglycine, DMTMM: 4-(4, 6-Dimethoxy-1,3,5-triazin-2-yl)-4-methylmorpholinium chloride, DnsHz: Dansylhydrazine, DTE: Dithioerythritol, EDTA: Ethylenediaminetetraacetic acid, EGTA: ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid, eq.: Equivalent, Et2O: Diethyl ether, ESI: Electrospray ionization, EtOAc: Ethyl acetate, FA: Formic acid, FIA: Flow Injection Analysis, FT-ICR MS: Fourier transform ion cyclotron resonance mass spectrometry, HESI: Heated electrospray ionization mode, Hex: n-Hexane, iOct: 2,2,4-trimethylpentane (isooctane), iPrOH: 2-propanol, LLE: Liquid–liquid Extraction, LTQ: Linear Trap Quadrupole, MeONa: Sodium methoxide, MF: Methyl formate, MSTFA: N-Methyl-N-trimethylsilyl-trifluoracetamide, MTBE: Methyl tert-butyl ether, nanoESI: nano electrospray ionization, NH4OAc: Ammonium acetate, NP: Normal phase, NPB: 3-nitrophenylboronic acid, OPE: One Phase Extraction, PAEA: 2-(2-Pyridilamino)-ethylamine, PBS: Phosphate Buffered Saline, PC: Protein content, PD-SPE: Phospholipid depletion solid phase extraction, PGWAT: Perigonadal white adipose tissue, PP: Potassium Phosphate, PrOH: 1-propanol, Q: Quadrupole, RP: Reverse Phase, SLE: Supported Liquid Extraction, SPE: Solid Phase Extraction, TFA: Trifluoroacetic acid, TOF: Time of flight, Tris-HCl: Tris(hydroxymethyl)aminomethane hydrochloride, UFLC: Ultra Fast Liquid Chromatography, µChip-SPE: Microchip-based SPE, µHP-SPE: micro-preparative high-performance solid-phase extraction. References were selected for studies conducted during the 2011–2020 period.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Aldana, J.; Romero-Otero, A.; Cala, M.P. Exploring the Lipidome: Current Lipid Extraction Techniques for Mass Spectrometry Analysis. Metabolites 2020, 10, 231. https://doi.org/10.3390/metabo10060231
AMA Style
Aldana J, Romero-Otero A, Cala MP. Exploring the Lipidome: Current Lipid Extraction Techniques for Mass Spectrometry Analysis. Metabolites. 2020; 10(6):231. https://doi.org/10.3390/metabo10060231
Chicago/Turabian StyleAldana, Julian, Adriana Romero-Otero, and Mónica P. Cala. 2020. "Exploring the Lipidome: Current Lipid Extraction Techniques for Mass Spectrometry Analysis" Metabolites 10, no. 6: 231. https://doi.org/10.3390/metabo10060231
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.