Maternal Choline Supplementation during Normal Murine Pregnancy Alters the Placental Epigenome: Results of an Exploratory Study

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Fetal Sex Genotyping
2.3. Placental RNA Extraction
2.4. Placental mRNA Sequencing and Data Analysis
2.5. Placental miRNA Sequencing and Data Analysis
2.6. Placental DNA Extraction
2.7. Promoter Methylation Assay and Data Analysis
2.8. Global DNA Methylation Assay and Data Analysis
3. Results
3.1. Placental Imprinted Gene Expression
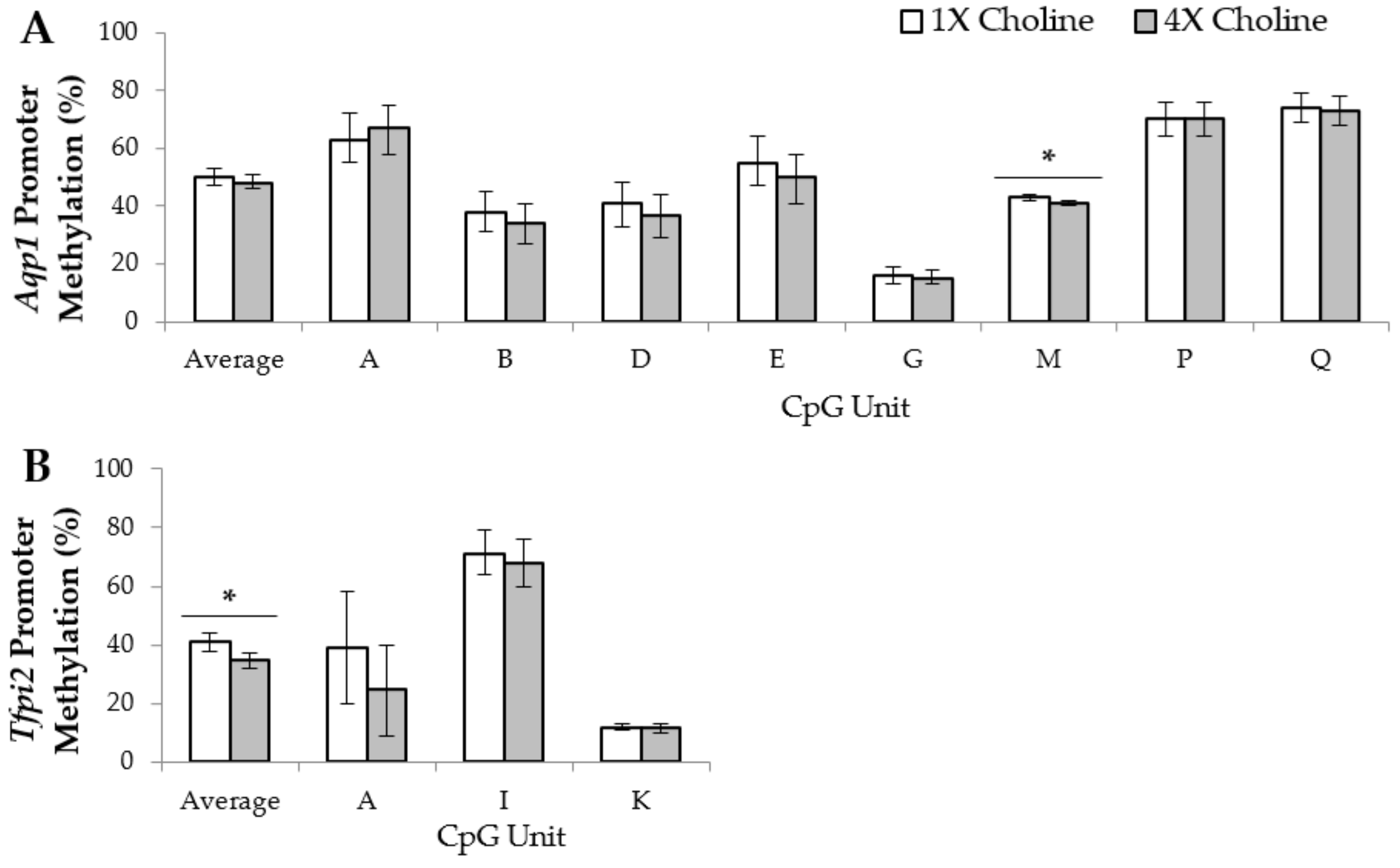
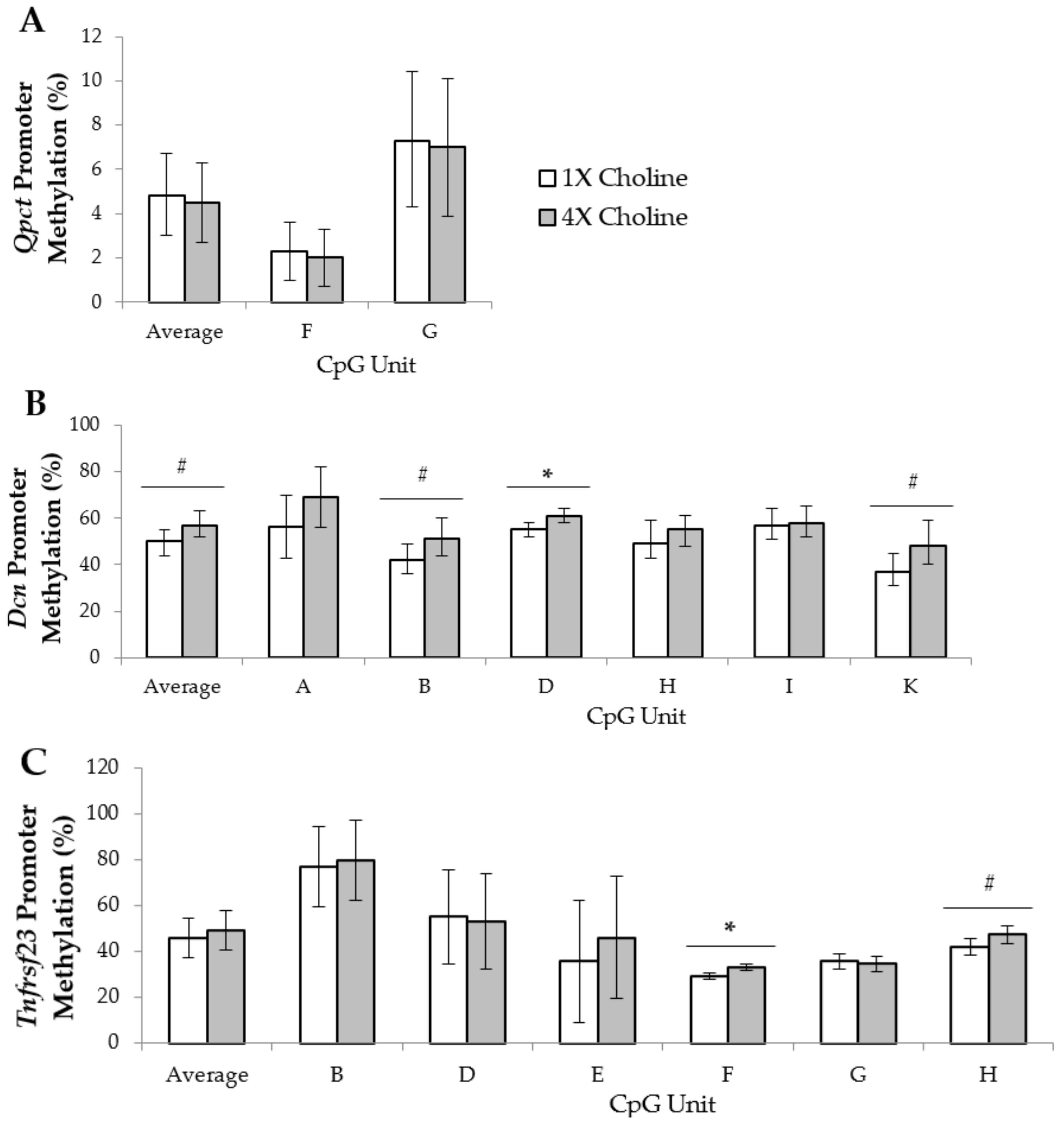
3.2. Placental Gene Promoter Methylation
3.3. Placental Global DNA Methylation
3.4. Placental miRNA Expression
4. Discussion
4.1. Maternal Choline Supplementation Alters the Expression of Several Imprinted Genes in the Placenta
4.2. Altered Promoter-Region Methylation May Be One Epigenetic Mechanism Contributing to the Choline-Induced Differential Expression of the Placental Imprinted Genes
4.3. Maternal Choline Supplementation Increases Global DNA Methylation in the Placenta
4.4. Maternal Choline Supplementation Reduces Placental miR-2137 Abundance, with Downstream Effects on the Expression of Its Potential Target Genes
4.5. Study Limitations
5. Conclusions and Future Directions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Godfrey, K.M.; Costello, P.M.; Lillycrop, K.A. Development, epigenetics and metabolic programming. Nestle Nutr. Inst. Workshop Ser. 2016, 85, 71–80. [Google Scholar] [PubMed]
- Brenseke, B.; Prater, M.R.; Bahamonde, J.; Gutierrez, J.C. Current thoughts on maternal nutrition and fetal programming of the metabolic syndrome. J. Pregnancy 2013, 2013, 368461. [Google Scholar] [CrossRef] [PubMed]
- Chmurzynska, A. Fetal programming: Link between early nutrition, DNA methylation, and complex diseases. Nutr. Rev. 2010, 68, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Longtine, M.S.; Nelson, D.M. Placental dysfunction and fetal programming: The importance of placental size, shape, histopathology, and molecular composition. Semin. Reprod. Med. 2011, 29, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.L.; Ferguson-Smith, A.C. Genomic imprinting effects in a compromised in utero environment: Implications for a healthy pregnancy. Semin. Cell Dev. Biol. 2010, 21, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Coan, P.M.; Burton, G.J.; Ferguson-Smith, A.C. Imprinted genes in the placenta—A review. Placenta 2005, 26 (Suppl. A), S10–S20. [Google Scholar] [CrossRef] [PubMed]
- Kappil, M.A.; Green, B.B.; Armstrong, D.A.; Sharp, A.J.; Lambertini, L.; Marsit, C.J.; Chen, J. Placental expression profile of imprinted genes impacts birth weight. Epigenetics 2015, 10, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Lesseur, C.; Paquette, A.G.; Marsit, C.J. Epigenetic regulation of infant neurobehavioral outcomes. Med. Epigenet. 2014, 2, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.M.; Cleal, J.K.; Ntani, G.; Crozier, S.R.; Mahon, P.A.; Robinson, S.M.; Harvey, N.C.; Cooper, C.; Inskip, H.M.; Godfrey, K.M.; et al. Relationship between placental expression of the imprinted phlda2 gene, intrauterine skeletal growth and childhood bone mass. Bone 2012, 50, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Lycoudi, A.; Mavreli, D.; Mavrou, A.; Papantoniou, N.; Kolialexi, A. Mirnas in pregnancy-related complications. Expert Rev. Mol. Diagn. 2015, 15, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Shruti, K.; Shrey, K.; Vibha, R. Micro RNAs: Tiny sequences with enormous potential. Biochem. Biophys. Res. Commun. 2011, 407, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Bidarimath, M.; Khalaj, K.; Wessels, J.M.; Tayade, C. MicroRNAs, immune cells and pregnancy. Cell. Mol. Immunol. 2014, 11, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Santa, L.M.; Teshima, L.Y.; Forero, J.V.; Giraldo, A.O. AngiomiRs: Potential biomarkers of pregnancy’s vascular pathologies. J. Pregnancy 2015, 2015, 320386. [Google Scholar] [PubMed]
- Higashijima, A.; Miura, K.; Mishima, H.; Kinoshita, A.; Jo, O.; Abe, S.; Hasegawa, Y.; Miura, S.; Yamasaki, K.; Yoshida, A.; et al. Characterization of placenta-specific microRNAs in fetal growth restriction pregnancy. Prenat. Diagn. 2013, 33, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Maccani, M.A.; Padbury, J.F.; Marsit, C.J. MiR-16 and miR-21 expression in the placenta is associated with fetal growth. PLoS ONE 2011, 6, e21210. [Google Scholar] [CrossRef] [PubMed]
- Kwan, S.T.C.; King, J.H.; Caudill, M.A. Choline and placental trophoblast development. In Human Placental Trophoblasts: Impact of Maternal Nutrition; Duttaroy, A.K., Basak, S., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 209–230. [Google Scholar]
- Kwan, S.T.C.; King, J.H.; Yan, J.; Jiang, X.; Wei, E.; Fomin, V.G.; Roberson, M.S.; Caudill, M.A. Maternal choline supplementation during murine pregnancy modulates placental markers of inflammation, apoptosis and vascularization in a fetal sex-dependent manner. Placenta 2017, 53, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Bar, H.Y.; Yan, J.; Jones, S.; Brannon, P.M.; West, A.A.; Perry, C.A.; Ganti, A.; Pressman, E.; Devapatla, S.; et al. A higher maternal choline intake among third-trimester pregnant women lowers placental and circulating concentrations of the antiangiogenic factor fms-like tyrosine kinase-1 (sflt1). FASEB J. 2013, 27, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Kwan, S.T.C.; King, J.H.; Yan, J.; Wang, Z.; Jiang, X.; Hutzler, J.S.; Klein, H.R.; Brenna, J.T.; Roberson, M.S.; Caudill, M.A. Maternal choline supplementation modulates placental nutrient transport and metabolism in late gestation of mouse pregnancy. J. Nutr. 2017, 147, 2083–2092. [Google Scholar] [CrossRef] [PubMed]
- King, J.H.; Kwan, S.T.C.; Yan, J.; Klatt, K.C.; Jiang, X.; Roberson, M.S.; Caudill, M.A. Maternal choline supplementation alters fetal growth patterns in a mouse model of placental insufficiency. Nutrients 2017, 9, 765. [Google Scholar] [CrossRef] [PubMed]
- Coan, P.M.; Ferguson-Smith, A.C.; Burton, G.J. Developmental dynamics of the definitive mouse placenta assessed by stereology. Biol. Reprod. 2004, 70, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Coan, P.M.; Angiolini, E.; Sandovici, I.; Burton, G.J.; Constancia, M.; Fowden, A.L. Adaptations in placental nutrient transfer capacity to meet fetal growth demands depend on placental size in mice. J. Physiol. 2008, 586, 4567–4576. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. Tophat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; Goff, L.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotechnol. 2013, 31, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Williamson, C.M.; Blake, A.; Thomas, S.; Beechey, C.V.; Hancock, J.; Cattanach, B.M.; Peters, J. World Wide Web Site—Mouse Imprinting Data and References. Available online: http://www.Har.Mrc.Ac.Uk/research/genomic_imprinting/ (accessed on 26 January 2017).
- Friedlander, M.R.; Chen, W.; Adamidi, C.; Maaskola, J.; Einspanier, R.; Knespel, S.; Rajewsky, N. Discovering microRNAs from deep sequencing data using mirdeep. Nat. Biotechnol. 2008, 26, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Alexander, K.; Clark, A.G.; Grimson, A.; Yu, H. Integrated network analysis reveals distinct regulatory roles of transcription factors and microRNAs. RNA (New York, NY) 2016, 22, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Wissink, E.M.; Smith, N.L.; Spektor, R.; Rudd, B.D.; Grimson, A. Micrornas and their targets are differentially regulated in adult and neonatal mouse cd8+ t cells. Genetics 2015, 201, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Huang, X.; Muruganujan, A.; Tang, H.; Mills, C.; Kang, D.; Thomas, P.D. Panther version 11: Expanded annotation data from gene ontology and reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2017, 45, D183–D189. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the panther classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Ehrich, M.; Nelson, M.R.; Stanssens, P.; Zabeau, M.; Liloglou, T.; Xinarianos, G.; Cantor, C.R.; Field, J.K.; van den Boom, D. Quantitative high-throughput analysis of DNA methylation patterns by base-specific cleavage and mass spectrometry. Proc. Natl. Acad. Sci. USA 2005, 102, 15785–15790. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Yan, J.; West, A.A.; Perry, C.A.; Malysheva, O.V.; Devapatla, S.; Pressman, E.; Vermeylen, F.; Caudill, M.A. Maternal choline intake alters the epigenetic state of fetal cortisol-regulating genes in humans. FASEB J. 2012, 26, 3563–3574. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; James, S.R.; Kazim, L.; Karpf, A.R. Specific method for the determination of genomic DNA methylation by liquid chromatography-electrospray ionization tandem mass spectrometry. Anal. Chem. 2005, 77, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Buckberry, S.; Bianco-Miotto, T.; Bent, S.J.; Dekker, G.A.; Roberts, C.T. Integrative transcriptome meta-analysis reveals widespread sex-biased gene expression at the human fetal-maternal interface. Mol. Hum. Reprod. 2014, 20, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; He, H.; Liu, H.; Liu, Q.; Zhang, L.; Liu, B.; Sugimoto, K.; Wu, Q. Aquaporin-1, a new maternally expressed gene, regulates placental development in the mouse. Biol. Reprod. 2016, 95, 40. [Google Scholar] [CrossRef] [PubMed]
- McMinn, J.; Wei, M.; Schupf, N.; Cusmai, J.; Johnson, E.B.; Smith, A.C.; Weksberg, R.; Thaker, H.M.; Tycko, B. Unbalanced placental expression of imprinted genes in human intrauterine growth restriction. Placenta 2006, 27, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Fons, C.; Campistol, J. Creatine defects and central nervous system. Semin. Pediatr. Neurol. 2016, 23, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Ma, D.; Sun, F.; Liang, W.; Liu, R.; Shen, W.; Wang, H.; Ji, Y.; Hu, R.; Liu, R.; et al. Over-expression of TFPI-2 promotes atherosclerotic plaque stability by inhibiting MMPs in apoE−/− mice. Int. J. Cardiol. 2013, 168, 1691–1697. [Google Scholar] [CrossRef] [PubMed]
- Udagawa, K.; Miyagi, Y.; Hirahara, F.; Miyagi, E.; Nagashima, Y.; Minaguchi, H.; Misugi, K.; Yasumitsu, H.; Miyazaki, K. Specific expression of PP5/TFPI2 mRNA by syncytiotrophoblasts in human placenta as revealed by in situ hybridization. Placenta 1998, 19, 217–223. [Google Scholar] [CrossRef]
- Mahnke-Zizelman, D.K.; D’Cunha, J.; Wojnar, J.M.; Brogley, M.A.; Sabina, R.L. Regulation of rat AMP deaminase 3 (isoform C) by development and skeletal muscle fibre type. Biochem. J. 1997, 326 Pt 2, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.; Funasaka, K.; Obayashi, T.; Miyahara, R.; Hirooka, Y.; Hamaguchi, M.; Goto, H.; Senga, T. AMPD3 is associated with the malignant characteristics of gastrointestinal stromal tumors. Oncol. Lett. 2017, 13, 1281–1287. [Google Scholar] [CrossRef] [PubMed]
- Guillomot, M.; Campion, E.; Prezelin, A.; Sandra, O.; Hue, I.; Le Bourhis, D.; Richard, C.; Biase, F.H.; Rabel, C.; Wallace, R.; et al. Spatial and temporal changes of decorin, type I collagen and fibronectin expression in normal and clone bovine placenta. Placenta 2014, 35, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Farrell, G.C.; Larter, C.Z.; Hou, J.Y.; Zhang, R.H.; Yeh, M.M.; Williams, J.; dela Pena, A.; Francisco, R.; Osvath, S.R.; Brooling, J.; et al. Apoptosis in experimental nash is associated with p53 activation and trail receptor expression. J. Gastroenterol. Hepatol. 2009, 24, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Clark, L.; Wei, M.; Cattoretti, G.; Mendelsohn, C.; Tycko, B. The tnfrh1 (tnfrsf23) gene is weakly imprinted in several organs and expressed at the trophoblast-decidua interface. BMC Genet. 2002, 3, 11. [Google Scholar] [CrossRef]
- Guo, J.; He, H.; Liu, Q.; Zhang, F.; Lv, J.; Zeng, T.; Gu, N.; Wu, Q. Identification and epigenetic analysis of a maternally imprinted gene Qpct. Mol. Cells 2015, 38, 859–865. [Google Scholar] [PubMed]
- Vaiman, D.; Calicchio, R.; Miralles, F. Landscape of transcriptional deregulations in the preeclamptic placenta. PLoS ONE 2013, 8, e65498. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, Z.; Simon, I. DNA methylation and gene expression. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Sandell, L.L.; Guan, X.J.; Ingram, R.; Tilghman, S.M. Gatm, a creatine synthesis enzyme, is imprinted in mouse placenta. Proc. Natl. Acad. Sci. USA 2003, 100, 4622–4627. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Zhou, W.; Fang, B.; Lu, W.; Loro, E.; Damle, M.; Ding, G.; Jager, J.; Zhang, S.; Zhang, Y.; et al. Dissociation of muscle insulin sensitivity from exercise endurance in mice by HDAC3 depletion. Nat. Med. 2017, 23, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.S.; Power, B.E.; Molloy, P.L. DNA hypomethylation and human diseases. Biochim. Biophys. Acta 2007, 1775, 138–162. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm-Benartzi, C.S.; Houseman, E.A.; Maccani, M.A.; Poage, G.M.; Koestler, D.C.; Langevin, S.M.; Gagne, L.A.; Banister, C.E.; Padbury, J.F.; Marsit, C.J. In utero exposures, infant growth, and DNA methylation of repetitive elements and developmentally related genes in human placenta. Environ. Health Perspect. 2012, 120, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.S. Sex-specific placental responses in fetal development. Endocrinology 2015, 156, 3422–3434. [Google Scholar] [CrossRef] [PubMed]
- Blusztajn, J.K.; Mellott, T.J. Neuroprotective actions of perinatal choline nutrition. Clin. Chem. Lab. Med. CCLM/FESCC 2013, 51, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.Y.; Briggs, D.I.; Vickers, M.H. Increased systolic blood pressure in rat offspring following a maternal low-protein diet is normalized by maternal dietary choline supplementation. J. Dev. Orig. Health Dis. 2012, 3, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Michel, V.; Singh, R.K.; Bakovic, M. The impact of choline availability on muscle lipid metabolism. Food Funct. 2011, 2, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Kular, J.; Tickner, J.C.; Pavlos, N.J.; Viola, H.M.; Abel, T.; Lim, B.S.; Yang, X.; Chen, H.; Cook, R.; Hool, L.C.; et al. Choline kinase beta mutant mice exhibit reduced phosphocholine, elevated osteoclast activity, and low bone mass. J. Biol. Chem. 2015, 290, 1729–1742. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D.; Zhang, P.; Gonzalez-Freire, M.; Moaddel, R.; Trehan, I.; Maleta, K.M.; Ordiz, M.I.; Ferrucci, L.; Manary, M.J. The association of serum choline with linear growth failure in young children from rural malawi. Am. J. Clin. Nutr. 2016, 104, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Huck, O.; Al-Hashemi, J.; Poidevin, L.; Poch, O.; Davideau, J.L.; Tenenbaum, H.; Amar, S. Identification and characterization of microRNA differentially expressed in macrophages exposed to porphyromonas gingivalis infection. Infect. Immunol. 2017, 85, e00771-16. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Ye, Y.; Qi, J.; Tan, X.; Zhang, Y.; Ma, Y.; Li, Y. Age and sex differences in microRNAs expression during the process of thymus aging. Acta Biochim. Biophys. Sin. 2017, 49, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Meissner, L.; Gallozzi, M.; Balbi, M.; Schwarzmaier, S.; Tiedt, S.; Terpolilli, N.A.; Plesnila, N. Temporal profile of microRNA expression in contused cortex after traumatic brain injury in mice. J. Neurotrauma 2016, 33, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zang, G.; Zhang, G.; Wang, H.; Zhang, X.; Johnston, N.; Min, W.; Luke, P.; Jevnikar, A.; Haig, A.; et al. MicroRNA and mRNA signatures in ischemia reperfusion injury in heart transplantation. PLoS ONE 2013, 8, e79805. [Google Scholar] [CrossRef] [PubMed]
- McPherson, N.O.; Fullston, T.; Kang, W.X.; Sandeman, L.Y.; Corbett, M.A.; Owens, J.A.; Lane, M. Paternal under-nutrition programs metabolic syndrome in offspring which can be reversed by antioxidant/vitamin food fortification in fathers. Sci. Rep. 2016, 6, 27010. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Sun, X.J.; Zhang, Y.L.; Kuang, Y.; Hu, C.Q.; Wu, W.L.; Shen, S.H.; Du, T.T.; Li, H.; He, F.; et al. Histone h3 lysine 36 methyltransferase hypb/setd2 is required for embryonic vascular remodeling. Proc. Natl. Acad. Sci. USA 2010, 107, 2956–2961. [Google Scholar] [CrossRef] [PubMed]
- Wierzowiecka, B.; Gomulkiewicz, A.; Cwynar-Zajac, L.; Olbromski, M.; Grzegrzolka, J.; Kobierzycki, C.; Podhorska-Okolow, M.; Dziegiel, P. Expression of metallothionein and vascular endothelial growth factor isoforms in breast cancer cells. In Vivo (Athens, Greece) 2016, 30, 271–278. [Google Scholar]
- Solberg, H.; Rinkenberger, J.; Dano, K.; Werb, Z.; Lund, L.R. A functional overlap of plasminogen and mmps regulates vascularization during placental development. Development (Cambridge, England) 2003, 130, 4439–4450. [Google Scholar] [CrossRef]
- Bizet, A.A.; Liu, K.; Tran-Khanh, N.; Saksena, A.; Vorstenbosch, J.; Finnson, K.W.; Buschmann, M.D.; Philip, A. The TGF-beta co-receptor, CD109, promotes internalization and degradation of TGF-beta receptors. Biochim. Biophys. Acta 2011, 1813, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Faura Tellez, G.; Vandepoele, K.; Brouwer, U.; Koning, H.; Elderman, R.M.; Hackett, T.L.; Willemse, B.W.; Holloway, J.; Van Roy, F.; Koppelman, G.H.; et al. Protocadherin-1 binds to smad3 and suppresses TGF-beta1-induced gene transcription. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 309, L725–L735. [Google Scholar] [CrossRef] [PubMed]
- Lo, P.K.; Chen, J.Y.; Lo, W.C.; Chen, B.F.; Hsin, J.P.; Tang, P.P.; Wang, F.F. Identification of a novel mouse p53 target gene dda3. Oncogene 1999, 18, 7765–7774. [Google Scholar] [CrossRef] [PubMed]
- Kalkat, M.; Garcia, J.; Ebrahimi, J.; Melland-Smith, M.; Todros, T.; Post, M.; Caniggia, I. Placental autophagy regulation by the bok-mcl1 rheostat. Autophagy 2013, 9, 2140–2153. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, X.; Su, L. Parthenolide induces apoptosis via TNFRSF10B and PMAIP1 pathways in human lung cancer cells. J. Exp. Clin. Cancer Res. CR 2014, 33, 3. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Harada, Y.; Watanabe, R.; Inutake, Y.; Ogawa, S.; Onuki, K.; Kagaya, S.; Tanabe, K.; Kishimoto, H.; Abe, R. Cd28 stimulation triggers nf-kappab activation through the carma1-pkctheta-grb2/gads axis. Int. Immunol. 2008, 20, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Heazell, A.E.; Crocker, I.P. Live and let die—Regulation of villous trophoblast apoptosis in normal and abnormal pregnancies. Placenta 2008, 29, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Ten Dijke, P.; Franzen, P.; Yamashita, H.; Ichijo, H.; Heldin, C.H.; Miyazono, K. Serine/threonine kinase receptors. Prog. Growth Factor Res. 1994, 5, 55–72. [Google Scholar] [CrossRef]
- Jones, R.L.; Stoikos, C.; Findlay, J.K.; Salamonsen, L.A. Tgf-beta superfamily expression and actions in the endometrium and placenta. Reproduction (Cambridge, England) 2006, 132, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Poplawski, A.; Binder, H. Feasibility of sample size calculation for RNA-seq studies. Brief. Bioinform. 2017, bbw144. [Google Scholar] [CrossRef] [PubMed]
- Adalsteinsson, B.T.; Gudnason, H.; Aspelund, T.; Harris, T.B.; Launer, L.J.; Eiriksdottir, G.; Smith, A.V.; Gudnason, V. Heterogeneity in white blood cells has potential to confound DNA methylation measurements. PLoS ONE 2012, 7, e46705. [Google Scholar] [CrossRef] [PubMed]
- Monk, D.; Wagschal, A.; Arnaud, P.; Muller, P.S.; Parker-Katiraee, L.; Bourc’his, D.; Scherer, S.W.; Feil, R.; Stanier, P.; Moore, G.E. Comparative analysis of human chromosome 7q21 and mouse proximal chromosome 6 reveals a placental-specific imprinted gene, TFPI2/TFPI2, which requires EHMT2 and EED for allelic-silencing. Genome Res. 2008, 18, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Monk, D. Genomic imprinting in the human placenta. Am. J. Obstet. Gynecol. 2015, 213, S152–S162. [Google Scholar] [CrossRef] [PubMed]
- Ginjala, V. Gene imprinting gateway. Genome Biol. 2001, 2, reports2009. [Google Scholar]
- Jirtle, R.L. Geneimprint. Available online: http://www.geneimprint.com/site/genes-by-species (accessed on 26 March 2018).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Fold Change | False Discovery Rate (FDR) Value |
|---|---|---|---|
| Aqp1 a | aquaporin 1 | 2.25 | 0.009 |
| Tfpi2 b | tissue factor pathway inhibitor 2 | 2.17 | 0.009 |
| Ampd3 b | adenosine monophosphate deaminase 3 | 1.63 | 0.009 |
| Gatm b | glycine amidinotransferase | 1.65 | 0.054 |
| Gene Symbol | Gene Name | Fold Change | FDR Value |
|---|---|---|---|
| Qpct | glutaminyl cyclase | 0.46 | 0.012 |
| Dcn | decorin | 0.58 | 0.012 |
| Tnfrsf23 | tumor necrosis factor receptor superfamily, member 23 | 0.62 | 0.012 |
| Biological Processes | p-Values |
|---|---|
| regulation of transcription from RNA polymerase II promoter (GO: 0006357) | <0.01 |
| transcription from RNA polymerase II promoter (GO: 0006366) | <0.01 |
| developmental process (GO: 0032502) | <0.01 |
| muscle organ development (GO: 0007517) | 0.01 |
| transcription, DNA-dependent (GO: 0006351) | 0.01 |
| segment specification (GO: 0007379) | 0.01 |
| nervous system development (GO: 0007399) | 0.01 |
| system development (GO: 0048731) | 0.02 |
| RNA metabolic process (GO: 0016070) | 0.02 |
| mesoderm development (GO: 0007498) | 0.05 |
| ectoderm development (GO: 0007398) | 0.05 |
| Female Placentas | Gene Symbol | Gene Name | Fold Difference |
| Gja4 | gap junction protein, alpha 4 | 1.33 | |
| Psrc1 | proline/serine-rich coiled-coil 1 | 1.49 | |
| Cd109 | CD109 antigen | 1.55 | |
| Mt3 | metallothionein 3 | 3.85 | |
| Plg | plasminogen | 3.92 | |
| Male Placentas | Pcdh1 | protocadherin 1 | 1.25 |
| Pmaip1 | phorbol-12-myristate-13-acetate-induced protein 1 | 1.39 | |
| Cd28 | CD28 antigen | 1.64 | |
| Mt3 | metallothionein 3 | 1.69 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwan, S.T.; King, J.H.; Grenier, J.K.; Yan, J.; Jiang, X.; Roberson, M.S.; Caudill, M.A. Maternal Choline Supplementation during Normal Murine Pregnancy Alters the Placental Epigenome: Results of an Exploratory Study. Nutrients 2018, 10, 417. https://doi.org/10.3390/nu10040417
Kwan ST, King JH, Grenier JK, Yan J, Jiang X, Roberson MS, Caudill MA. Maternal Choline Supplementation during Normal Murine Pregnancy Alters the Placental Epigenome: Results of an Exploratory Study. Nutrients. 2018; 10(4):417. https://doi.org/10.3390/nu10040417
Chicago/Turabian StyleKwan, Sze Ting (Cecilia), Julia H. King, Jennifer K. Grenier, Jian Yan, Xinyin Jiang, Mark S. Roberson, and Marie A. Caudill. 2018. "Maternal Choline Supplementation during Normal Murine Pregnancy Alters the Placental Epigenome: Results of an Exploratory Study" Nutrients 10, no. 4: 417. https://doi.org/10.3390/nu10040417